Modeling Climate Change Impacts on Rangeland Productivity and Livestock Population Dynamics in Nkayi District, Zimbabwe

 ,
,
Abstract
:1. Introduction
Objectives
- To analyse the trends in biomass (herbaceous, shrub and tree layers) production in relation to rainfall in Nkayi district (Zimbabwe) over a period of 30 years (1980–2010).
- To assess the effect of potential changes in climatic variables (rainfall, temperature, and atmospheric carbon dioxide) on rangeland productivity and livestock population dynamics on a temporal and spatial scale.
- To analyse how the biomass production from the tree, shrub and grass layers vary spatially in different times of the year.
2. Methodology
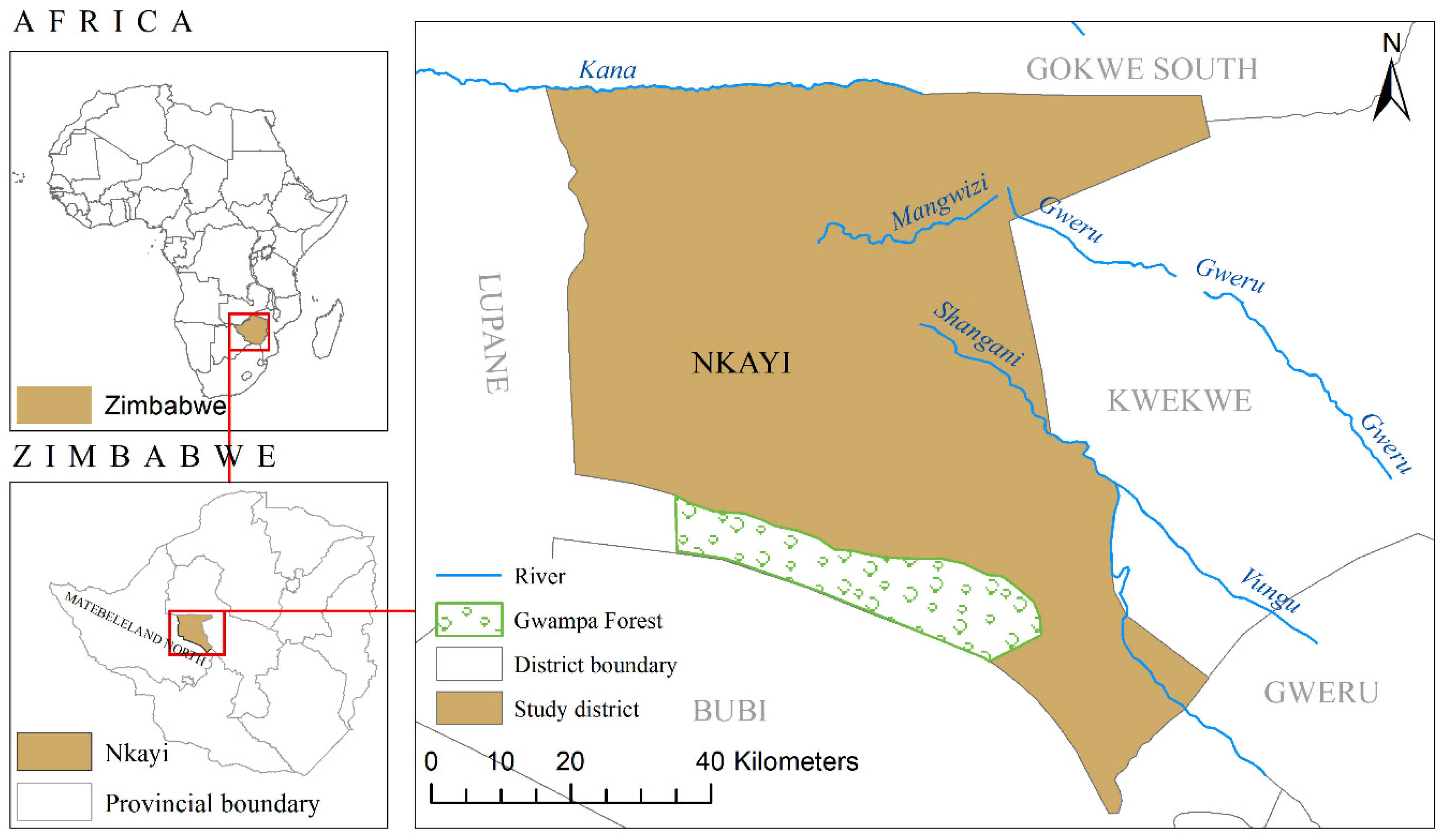
2.1. Study Area
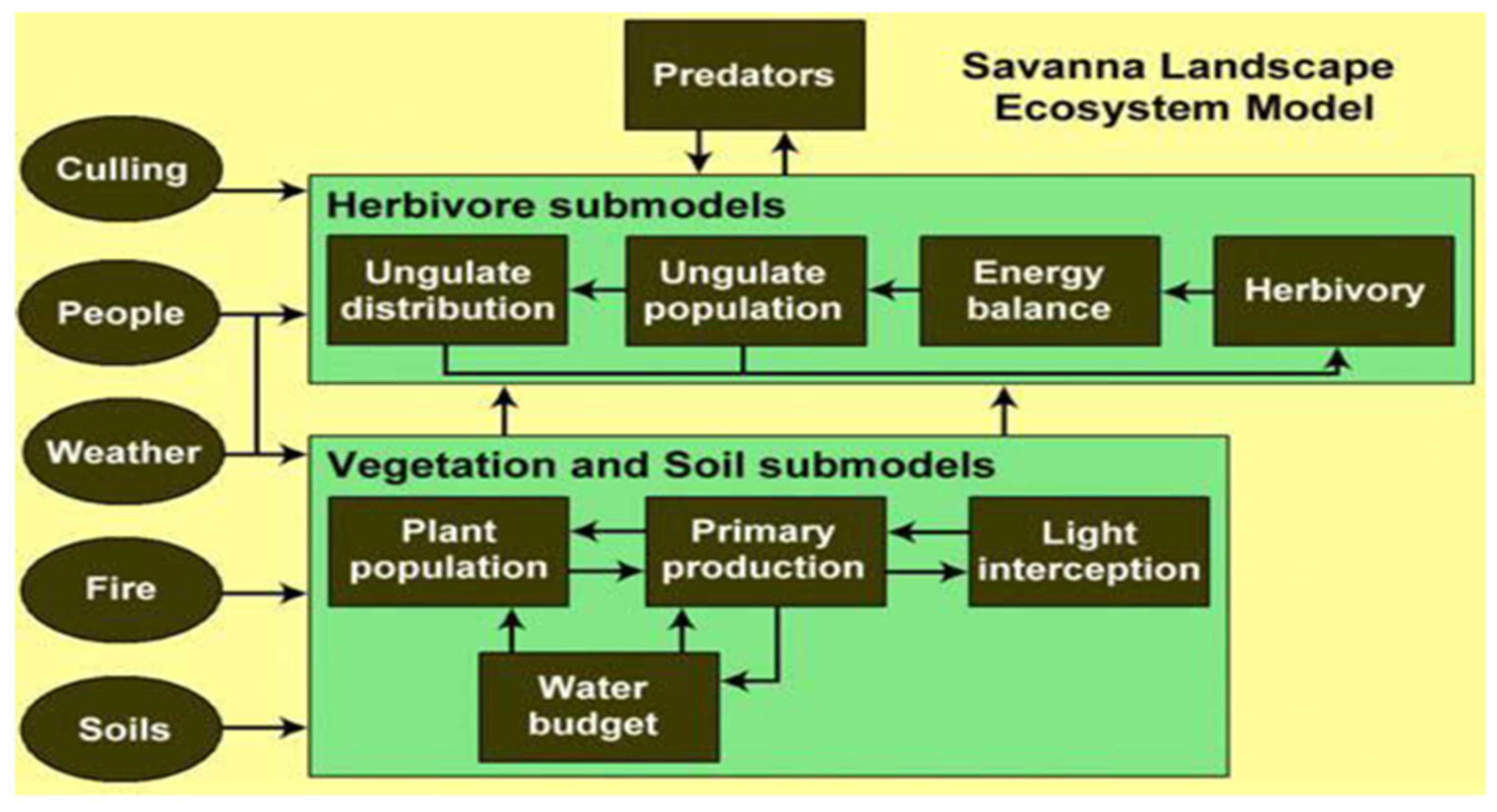
2.2. The SAVANNA Modeling System
2.3. Model Parameterization and Calibration
2.4. Climate Change Scenarios
3. Results
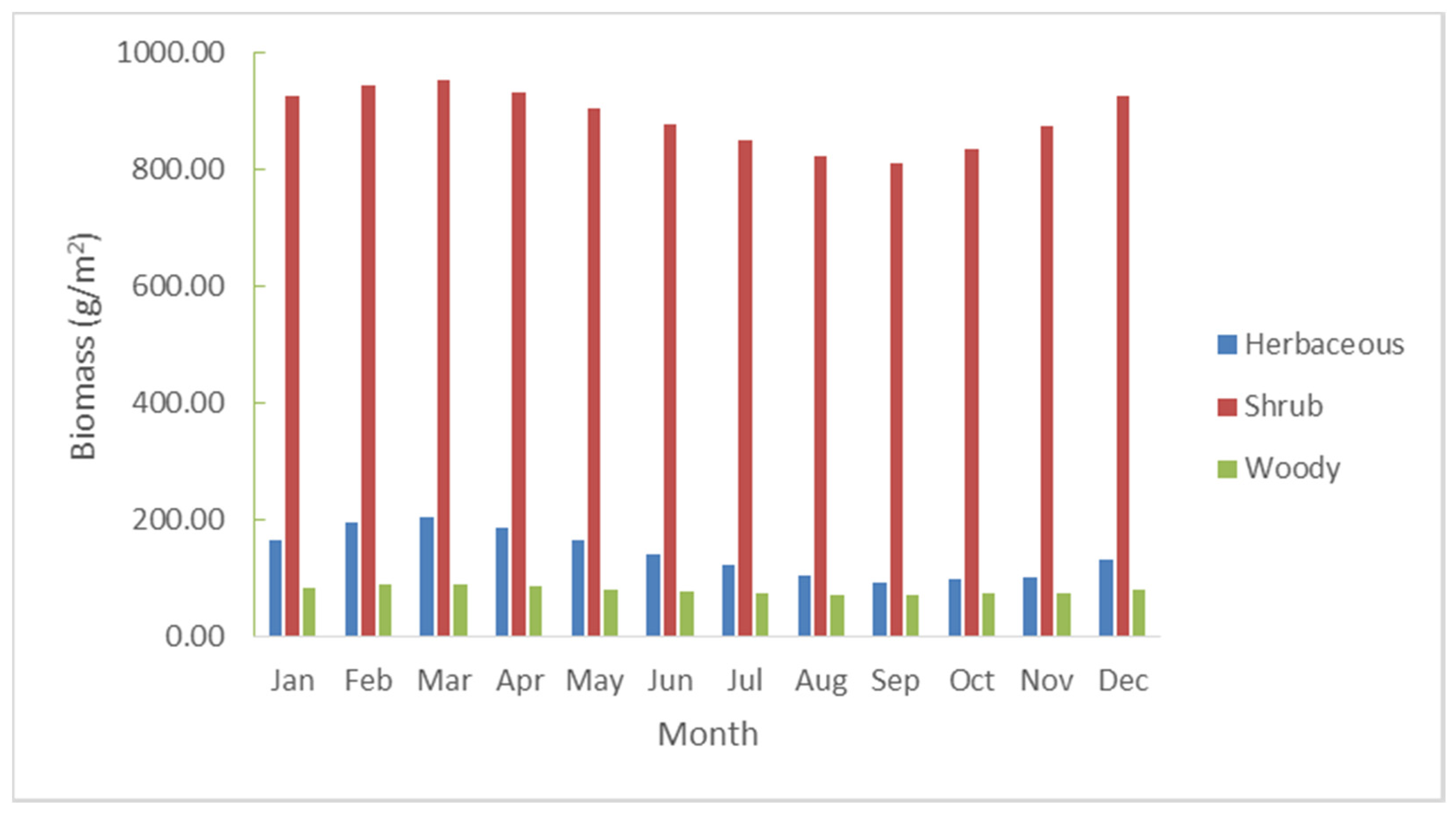
3.1. Historical Scenario: Biomass Production over the Entire Nkayi District
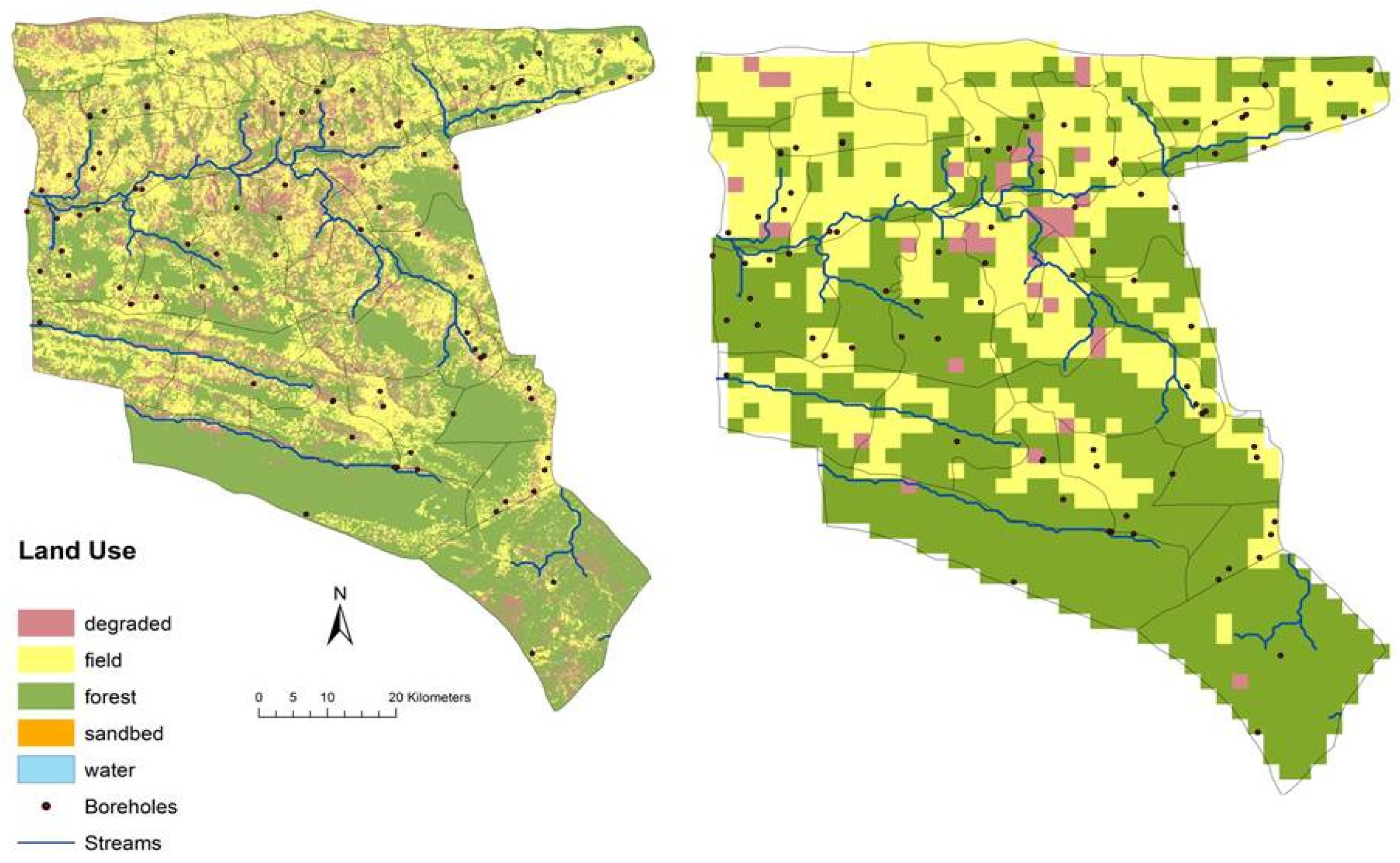
3.2. Historical Scenario: The Spatial Distribution of Green Biomass Production
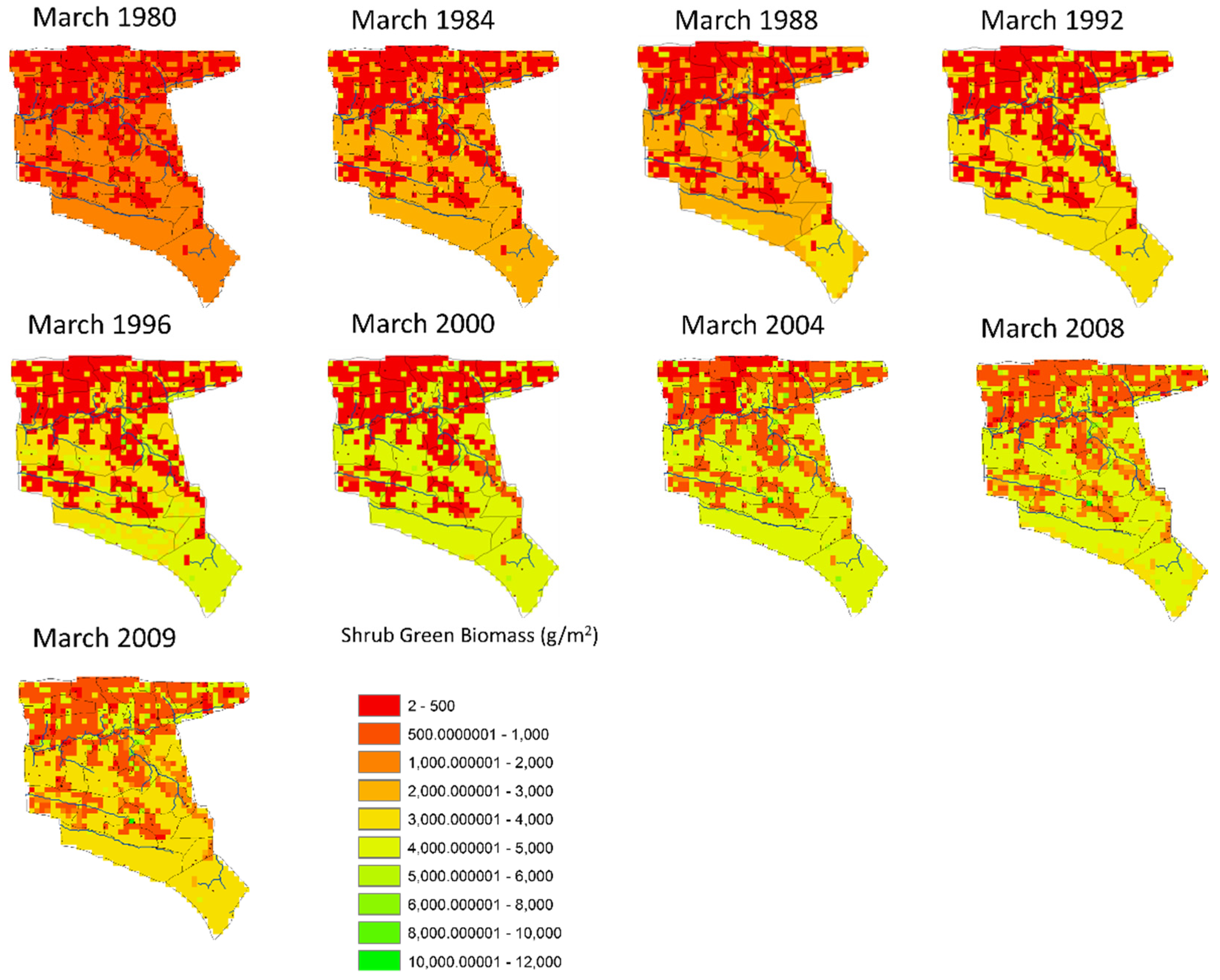
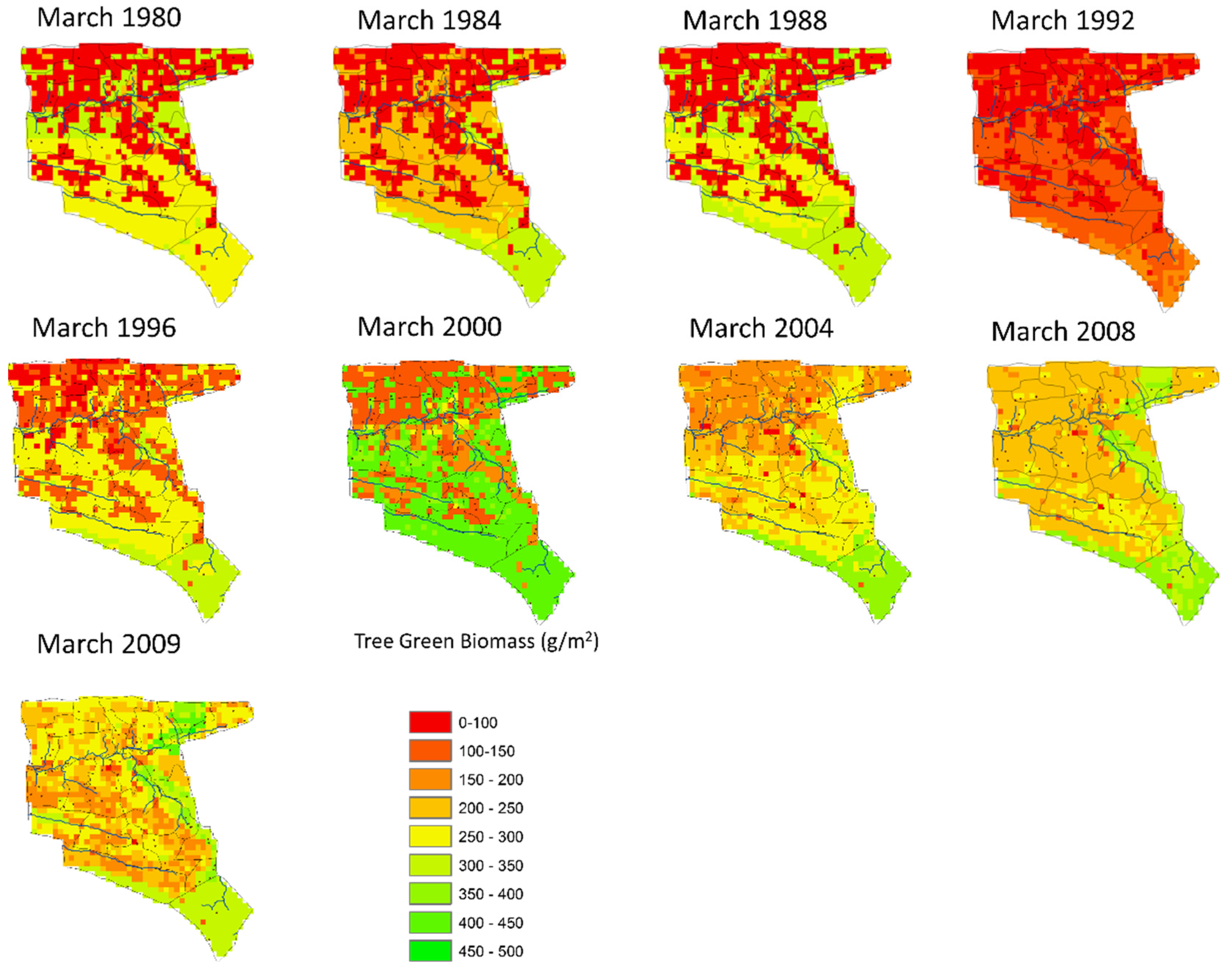
3.3. Historical Scenario: The Spatial Distribution of Shrub and Tree Cover
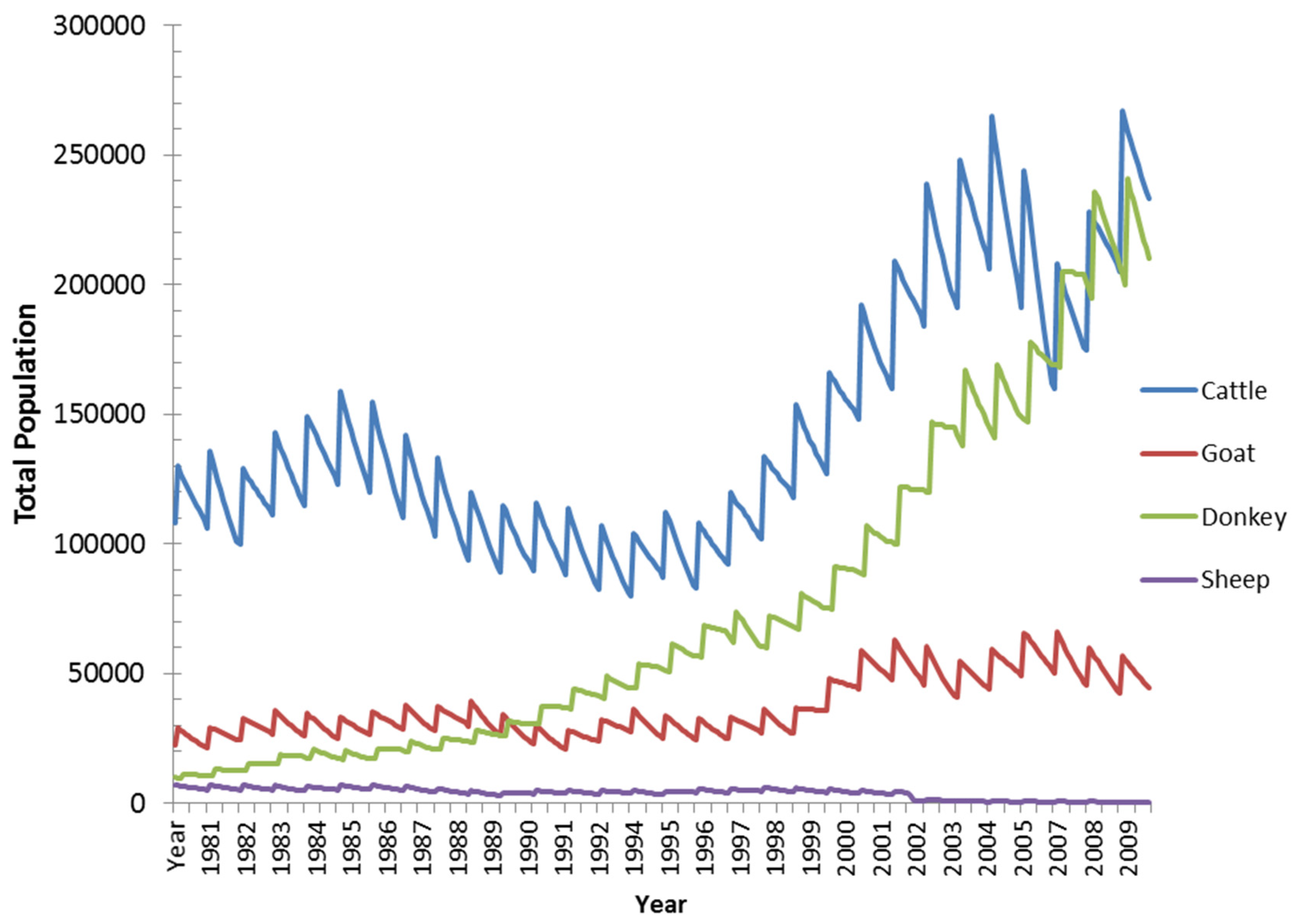
3.4. Historical Scenario: Livestock Population Dynamics
3.5. Climate Change Scenarios: Simulated Grass Biomass
3.6. Climate Change Scenarios: Simulated Shrub Biomass
3.7. Climate Change Scenarios: Simulated Tree Biomass
3.8. Climate Change Scenarios: Simulated Livestock Dynamics
4. Discussion
4.1. Response of Herbaceous Biomass to Yearly Rainfall Totals
4.2. Shrub Biomass and Cover Increase in All Historical and Climate Change Scenarios
4.3. Woody Biomass and Cover Increase Differently with Respect to Assumptions on Temperature, Precipitation and CO2 Effects
4.4. Simulated Livestock Populations Are Not Particularly Sensitive to Climate Change Conditions
5. Conclusions and Recommendations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turner, M.D.; McPeak, J.G.; Ayantunde, A.A. The Role of Livestock Mobility in the Livelihood Strategies of Rural Peoples in Semi-Arid West Africa. Hum. Ecol. 2014, 42, 231–247. [Google Scholar] [CrossRef]
- Seguin, B. The Consequences of Global Warming for Agriculture and Food Production; Rowlinson, P., Steele, M., Nefzaoui, A., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 9–11. [Google Scholar]
- Senda, T.S.; Peden, D.; Tui, S.H.-K.; Sisito, G.; Van Rooyen, A.F.; Sikosana, J.L.N. Gendered livelihood implications for improvements of livestock Water productivity In Zimbabwe. Exp. Agric. 2011, 47, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Goat Production and Marketing: Baseline Information for Semi-Arid Zimbabwe. Available online: http://oar.icrisat.org/397/1/CO_200704.pdf (accessed on 25 March 2020).
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupa, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Gelats, F.L.; Fraser, E.D.; Morton, J.F.; Rivera-Ferre, M.G. What drives the vulnerability of pastoralists to global environmental change? A qualitative meta-analysis. Glob. Environ. Chang. 2016, 39, 258–274. [Google Scholar] [CrossRef]
- Telesetsky, A. Organisation for Economic Co-operation and Development (OECD). Int. Environ. Law 2012, 23, 578–582. [Google Scholar] [CrossRef]
- FAO and Platform for Agrobiodiversity Research. Biodivers. Food Agric. 2011, 53, 78.
- Herrero, M.; Grace, D.; Njuki, J.; Johnson, N.; Enahoro, D.; Silvestri, S.; Rufino, M.C. The roles of livestock in developing countries. Animal 2012, 7, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Illius, A.W.; Gordon, I.J.; Derry, J.F.; Magadzire, Z.; Mukungurutse, E. Environmental variability and productivity of semi-arid grazing systems. In Proceedings of the Third Workshop on Livestock Production Programme Projects, Matobo, Zimbabwe, 28 September 2000; pp. 125–136. [Google Scholar]
- Sullivan, S.; Rohde, R. On non-equilibrium in arid and semi-arid grazing systems. J. Biogeogr. 2002, 29, 1595–1618. [Google Scholar] [CrossRef] [Green Version]
- Verburg, P.H.; Van De Steeg, J.; Veldkamp, A.; Willemen, L.; Veldkamp, T. From land cover change to land function dynamics: A major challenge to improve land characterization. J. Environ. Manag. 2009, 90, 1327–1335. [Google Scholar] [CrossRef]
- Vetter, S. Rangelands at equilibrium and non-equilibrium: Recent developments in the debate. J. Arid. Environ. 2005, 62, 321–341. [Google Scholar] [CrossRef]
- Boone, R.; Wang, G. Cattle dynamics in African grazing systems under variable climates. J. Arid. Environ. 2007, 70, 495–513. [Google Scholar] [CrossRef]
- Puig, C.J.; Greiner, R.; Huchery, C.; Perkins, I.; Bowen, L.; Collier, N.; Garnett, S.T. Beyond cattle: Potential futures of the pastoral industry in the Northern Territory. Rangel. J. 2011, 33, 181–194. [Google Scholar] [CrossRef]
- Masikati, P.; Tui, S.H.-K.; Descheemaeker, K.; Sisito, G.; Senda, T.S.; Crespo, O.; Nhamo, N. Integrated Assessment of Crop–Livestock Production Systems Beyond Biophysical Methods. In Smart Technologies for Sustainable Smallholder Agriculture; Elsevier BV: Amsterdam, The Netherlands, 2017; pp. 257–278. [Google Scholar]
- Coughenour, M.B. Savanna-Landscape and Regional Ecosystem Model: User Manual. 1993. Available online: https://www.nrel.colostate.edu/assets/nrel_files/labs/coughenour-lab/pubs/Manual_1993/user_manual.pdf (accessed on 25 March 2020).
- O’Connor, T.; Kiker, G.A. Collapse of the Mapungubwe Society: Vulnerability of Pastoralism to Increasing Aridity. Clim. Chang. 2004, 66, 49–66. [Google Scholar] [CrossRef]
- Weisberg, P.J.; Coughenour, M.B.; Bugmann, H. Large Herbivore Ecology, Ecosystem Dynamics and Conservation; Cambridge University Press: Cambridge, UK, 2006; pp. 348–382. [Google Scholar]
- Bunting, E.L.; Fullman, T.; Kiker, G.; Southworth, J. Utilization of the SAVANNA model to analyze future patterns of vegetation cover in Kruger National Park under changing climate. Ecol. Model. 2016, 342, 147–160. [Google Scholar] [CrossRef]
- Hilbers, J.P.; Van Langevelde, F.; Prins, H.H.T.; Grant, C.C.; Peel, M.J.S.; Coughenour, M.B.; De Knegt, H.; Slotow, R.; Smit, I.P.J.; Kiker, G.A.; et al. Modeling elephant-mediated cascading effects of water point closure. Ecol. Appl. 2015, 25, 402–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fullman, T.J.; Bunting, E.L.; Kiker, G.A.; Southworth, J. Predicting shifts in large herbivore distributions under climate change and management using a spatially-explicit ecosystem model. Ecol. Model. 2017, 352, 1–18. [Google Scholar] [CrossRef]
- Boone, R.; Galvin, K.A.; Coughenour, M.B.; Hudson, J.W.; Weisberg, P.J.; Vogel, C.H.; Ellis, J.E. Ecosystem Modeling Adds Value to a South African Climate Forecast. Clim. Chang. 2004, 64, 317–340. [Google Scholar] [CrossRef]
- Thornton, P.; Fawcett, R.; Galvin, K.; Boone, R.; Hudson, J.; Vogel, C. Evaluating management options that use climate forecasts: Modelling livestock production systems in the semi-arid zone of South Africa. Clim. Res. 2004, 26, 33–42. [Google Scholar] [CrossRef]
- Thornton, P.; BurnSilver, S.; Boone, R.; Galvin, K. Modelling the impacts of group ranch subdivision on agro-pastoral households in Kajiado, Kenya. Agric. Syst. 2006, 87, 331–356. [Google Scholar] [CrossRef]
- Boone, R.; Galvin, K.A.; BurnSilver, S.B.; Thornton, P.; Ojima, D.S.; Jawson, J.R. Using Coupled Simulation Models to Link Pastoral Decision Making and Ecosystem Services. Ecol. Soc. 2011, 16. [Google Scholar] [CrossRef]
- Boone, R.; Conant, R.T.; Sircely, J.; Thornton, P.; Herrero, M. Climate change impacts on selected global rangeland ecosystem services. Glob. Chang. Boil. 2017, 24, 1382–1393. [Google Scholar] [CrossRef]
- Higgins, S.I.; Scheiter, S. Atmospheric CO2 forces abrupt vegetation shifts locally, but not globally. Nature. 2012, 488, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Weber, U.; Jung, M.; Reichstein, M.; Beer, C.; Braakhekke, M.C.; Lehsten, V.; Ghent, D.; Kaduk, J.; Viovy, N.; Ciais, P.; et al. The interannual variability of Africa’s ecosystem productivity: A multi-model analysis. Biogeosciences 2009, 6, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Kiker, G.A. Development and Comparison of Savanna Ecosystem Models to Explore the Concept of Carrying Capacity. Ph.D. Thesis, Cornell University, New York, NY, USA, August 1998. [Google Scholar]
- Second Round Crop and Livestock Assessment Report: 2014/15 Season. 2015. Available online: https://documents.wfp.org/stellent/groups/public/documents/newsroom/wfp275316.pdf (accessed on 25 March 2020).
- Stuart-Hill, G.; Scholes, R.J.; Walker, B.H. An African Savanna: Synthesis of the Nylsvley Study. J. Appl. Ecol. 1994, 31, 791. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, G.F. A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas. Glob. Chang. Boil. 2000, 6, 865–869. [Google Scholar] [CrossRef]
- Michael, B.C. Natural Resource Ecology Laboratory, Colorado State University. Unpublished work. 2014. [Google Scholar]
- Warf, B.; Poore, B.S. United States Geological Survey (USGS) (2020) Earth Explorer. Available online: https://earthexplorer.usgs.gov/ (accessed on 25 March 2020).
- Chirima, A.; Vanrooyen, A.; Homann, S.; Mundy, P.; Ncube, N.; Sibanda, N. Land degradation: Evaluating land cover change of mixed crop livestock systems in the semi-arid Zimbabwe from 1990 to 2009. Midlands State Univ. J. Sci. Agric. Technol. 2012, 3. [Google Scholar]
- Whitlow, R. Potential versus actual erosion in Zimbabwe. Appl. Geogr. 1988, 8, 87–100. [Google Scholar] [CrossRef]
- Pike, A.; Schulze, R.E. AUTOSOIL Version 3: A Soils Decision Support System for South African Soils; Department of Agricultural Engineering, University of Natal: Pietermaritzburg, South Africa, 1995. [Google Scholar]
- Soils: Agrohydrological Information Needs, Information Sources and Decision Support. Available online: http://sarva2.dirisa.org/ (accessed on 25 March 2020).
- Senda, T.S. University of Nairobi, LARMAT, Kenya. Unpublished work. 2020. [Google Scholar]
- Antle, J.; Valdivia, R. Modelling the supply of ecosystem services from agriculture: A minimum-data approach. Aust. J. Agric. Resour. Econ. 2006, 50, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gama, A.C.; Mapemba, L.D.; Masikati, P.; Tui, S.H.K.; Crespo, O.; Bandason, E. Modeling Potential Impacts of Future Climate Change in Mzimba District, Malawi, 2040–2070: An Integrated Biophysical and Economic Modeling Approach; International Food Policy Research Institute: Washington, DC, USA, 2014; Volume 8. [Google Scholar]
- Coughenour, M. The Ellis paradigm—humans, herbivores and rangeland systems. Afr. J. Range Forage Sci. 2004, 21, 191–200. [Google Scholar] [CrossRef]
- Gordon, I.J. Browsing and grazing ruminants: Are they different beasts? For. Ecol. Manag. 2003, 181, 13–21. [Google Scholar] [CrossRef]
- Forests, Rangelands and Climate Change in Southern Africa. Available online: https://www.uncclearn.org/sites/default/files/inventory/fao190.pdf (accessed on 25 March 2020).
- Mokgosi, R.O. Effects of bush encroachment control in a communal managed area in the Taung region, North West Province, South Africa. Ph.D. Thesis, North-West University, Potchefstroom, South Africa, May 2018. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Core Questions | GCM Scenarios | |
|---|---|---|
| Historic Scenario (1980–2010) | ||
| 1. What are the temporal and spatial dynamics of biomass production from the three layers? (tree, shrub, herbaceous) 2. How do the livestock populations respond to changes in the vegetation? |
|
|
| Hot and wet scenario | ||
|
| |
| Hot and dry scenario | ||
|
| |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senda, T.S.; Kiker, G.A.; Masikati, P.; Chirima, A.; van Niekerk, J. Modeling Climate Change Impacts on Rangeland Productivity and Livestock Population Dynamics in Nkayi District, Zimbabwe. Appl. Sci. 2020, 10, 2330. https://doi.org/10.3390/app10072330
Senda TS, Kiker GA, Masikati P, Chirima A, van Niekerk J. Modeling Climate Change Impacts on Rangeland Productivity and Livestock Population Dynamics in Nkayi District, Zimbabwe. Applied Sciences. 2020; 10(7):2330. https://doi.org/10.3390/app10072330
Chicago/Turabian StyleSenda, Trinity S., Gregory A. Kiker, Patricia Masikati, Albert Chirima, and Johan van Niekerk. 2020. "Modeling Climate Change Impacts on Rangeland Productivity and Livestock Population Dynamics in Nkayi District, Zimbabwe" Applied Sciences 10, no. 7: 2330. https://doi.org/10.3390/app10072330