
Geongangbuja-Tang Decoction and Its Active Ingredient, Aconiti Lateralis Radix Preparata, Exerts Inhibitory Effects on Heat Stress-Induced Inflammation in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Animals
2.3. Open-Field Test
2.4. Measurement of Cortisol Level in Serum
2.5. Western Blot Analysis
2.6. Measurement of IL-9, IL-13, PGE1 and PGE2 Levels
2.7. Statistical Analysis
3. Results
3.1. Effects of GBT on Heat-Induced Sickness Behaviors and C-Fos Expression in Mice
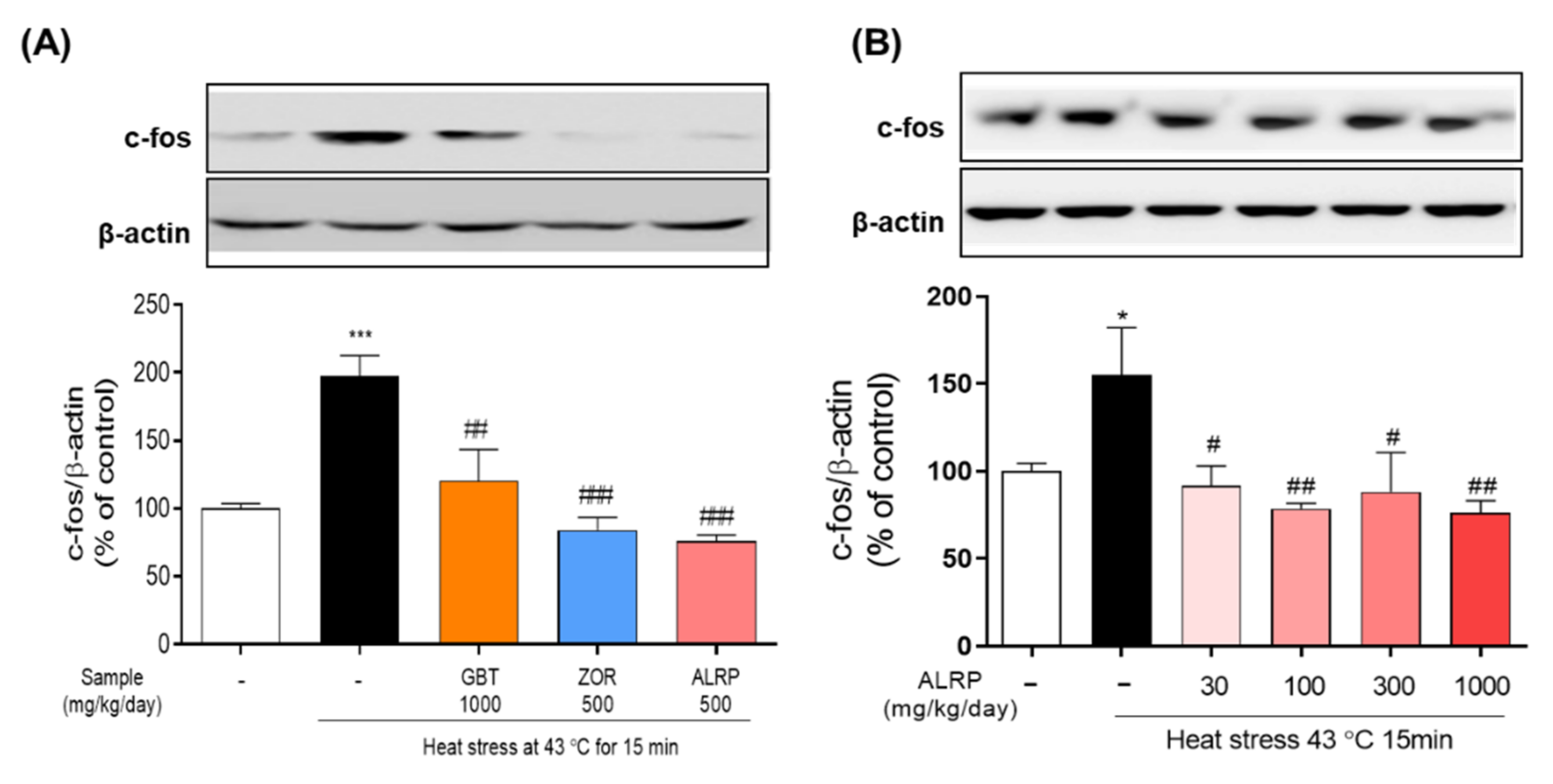
3.2. Effects of GBT and Its Components, ALRP and ZOR, on C-Fos Expression in the Mouse Hypothalamus
3.3. Effects of ALRP on Heat Stress-Induced Production of Inflammatory Mediators in the Mouse Hypothalamus
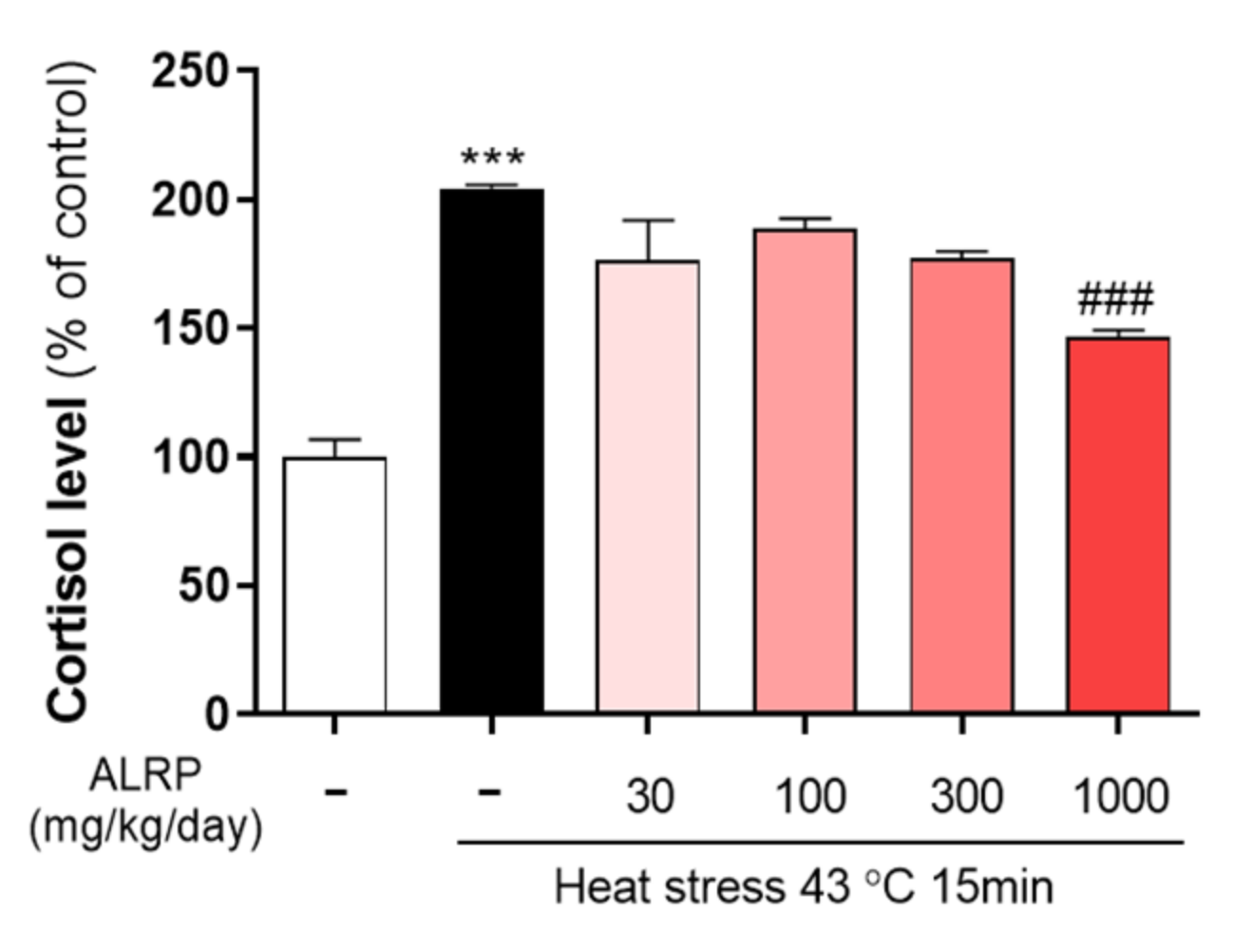
3.4. Effects of ALRP on Heat Stress-Indzuced Secretion of Cortisol
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Hoffman-Goetz, L.; Keir, R. Body temperature responses of aged mice to ambient temperature and humidity stress. J. Gerontol. 1984, 39, 547–551. [Google Scholar] [CrossRef]
- Lee, W.; Moon, M.; Kim, H.G.; Lee, T.H.; Oh, M.S. Heat stress-induced memory impairment is associated with neuroinflammation in mice. J. Neuroinflamm. 2015, 12, 102. [Google Scholar] [CrossRef] [Green Version]
- Wright, H.E.; Selkirk, G.A.; McLellan, T.M. HPA and SAS responses to increasing core temperature during uncompensable exertional heat stress in trained and untrained males. Eur. J. Appl. Physiol. 2010, 108, 987–997. [Google Scholar] [CrossRef]
- Jasnic, N.; Djordjevic, J.; Vujovic, P.; Lakic, I.; Djurasevic, S.; Cvijic, G. The effect of vasopressin 1b receptor (V1bR) blockade on HPA axis activity in rats exposed to acute heat stress. J. Exp. Biol. 2013, 216, 2302–2307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fee, C.; Prevot, T.; Misquitta, K.; Banasr, M.; Sibille, E. Chronic Stress-induced Behaviors Correlate with Exacerbated Acute Stress-induced Cingulate Cortex and Ventral Hippocampus Activation. Neuroscience 2020, 440, 113–129. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, Y.L.; Liu, S.X.; Zheng, Q.F.; Liu, T.T.; Ma, X.; Zhang, P.; Li, R.S.; Wang, L.F.; Li, Y.G.; et al. Study on hot property differences of Aconiti Lateralis Radix Praeparata and its compatibility with different ginger processed products based on bio-thermodynamics. J. Therm. Anal. Calorim. 2015, 120, 1043–1051. [Google Scholar] [CrossRef]
- Wen, J.X.; Zou, W.J.; Wang, R.L.; Liu, H.H.; Yang, Y.X.; Li, H.T.; Wei, S.Z.; Li, R.S.; Cai, H.D.; Wang, J.; et al. Cardioprotective effects of Aconiti Lateralis Radix Praeparata combined with Zingiberis Rhizoma on doxorubicin-induced chronic heart failure in rats and potential mechanisms. J. Ethnopharmacol. 2019, 238, 111880. [Google Scholar] [CrossRef]
- Ibtisham, F.; Nawab, A.; Niu, Y.F.; Wang, Z.; Wu, J.; Xiao, M.; An, L.L. The effect of ginger powder and Chinese herbal medicine on production performance, serum metabolites and antioxidant status of laying hens under heat-stress condition. J. Therm. Biol. 2019, 81, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Safiullah; Chand, N.; Khan, R.U.; Naz, S.; Ahmad, M.; Gul, S. Effect of ginger (Zingiber officinale Roscoe) and organic selenium on growth dynamics, blood melanodialdehyde and paraoxonase in broilers exposed to heat stress. J. Appl. Anim. Res. 2019, 47, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.Y.; Kim, M.H.; Hong, J.; Kim, S.H.; Yang, W.M. Dried Ginger (Zingiber officinalis) Inhibits Inflammation in a Lipopolysaccharide-Induced Mouse Model. Evid. Based Complement. Altern. Med. 2013, 2013, 914563. [Google Scholar] [CrossRef] [Green Version]
- Waggas, A.M. Neuroprotective evaluation of extract of ginger (Zingiber officinale) root in monosodium glutamate-induced toxicity in different brain areas male albino rats. Pak. J. Biol. Sci. 2009, 12, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, L.; Dong, L.; Hu, X.S.; Feng, F.Q.; Chen, F. 6-Gingerol, a Functional Polyphenol of Ginger, Promotes Browning through an AMPK-Dependent Pathway in 3T3-L1 Adipocytes. J. Agric. Food Chem. 2019, 67, 14056–14065. [Google Scholar] [CrossRef]
- Kim, W.; Lee, W.; Choi, J.G.; Ju, I.G.; Kim, Y.K.; Lee, T.H.; Oh, M.S. Inhibitory effects of Aconiti Lateralis Radix Preparata on chronic intermittent cold-induced inflammation in the mouse hypothalamus. J. Ethnopharmacol. 2018, 215, 27–33. [Google Scholar] [CrossRef]
- Makino, T.; Kato, K.; Mizukami, H. Processed aconite root prevents cold-stress-induced hypothermia and immuno-suppression in mice. Biol. Pharm. Bull. 2009, 32, 1741–1748. [Google Scholar] [CrossRef] [Green Version]
- Moon, M.; Huh, E.; Lee, W.; Song, E.J.; Hwang, D.S.; Lee, T.H.; Oh, M.S. Coptidis Rhizoma Prevents Heat Stress-Induced Brain Damage and Cognitive Impairment in Mice. Nutrients 2017, 9, 1057. [Google Scholar] [CrossRef] [Green Version]
- Gavrilov, Y.V.; Perekrest, S.V.; Novikova, N.S.; Korneva, E.A. Stress-induced changes in cellular responses in hypothalamic structures to administration of an antigen (lipopolysaccharide) (in terms of c-Fos protein expression). Neurosci. Behav. Physiol. 2008, 38, 189–194. [Google Scholar] [CrossRef]
- De Medeiros, M.A.; Reis, L.R.; Mello, L.E. Stress-induced c-Fos expression is differentially modulated by dexamethasone, diazepam and imipramine. Neuropsychopharmacology 2005, 30, 1246–1256. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Bueno, B.; Madrigal, J.L.; Perez-Nievas, B.G.; Leza, J.C. Stress mediators regulate brain prostaglandin synthesis and peroxisome proliferator-activated receptor-gamma activation after stress in rats. Endocrinology 2008, 149, 1969–1978. [Google Scholar] [CrossRef] [Green Version]
- Varghese, F.P.; Brown, E.S. The Hypothalamic-Pituitary-Adrenal Axis in Major Depressive Disorder: A Brief Primer for Primary Care Physicians. Prim. Care Companion J. Clin. Psychiatry 2001, 3, 151–155. [Google Scholar] [CrossRef]
- Liu, J.; Tan, Y.; Ao, H.; Peng, C. Aqueous extracts of Aconite promote thermogenesis in rats with hypothermia via regulating gut microbiota and bile acid metabolism. Chin. Med. 2021, 16, 29. [Google Scholar] [CrossRef]
- Kvetnansky, R.; Sabban, E.L.; Palkovits, M. Catecholaminergic Systems in Stress: Structural and Molecular Genetic Approaches. Physiol. Rev. 2009, 89, 535–606. [Google Scholar] [CrossRef]
- Sato, H.; Yamada, C.; Konno, Y.; Ohzumi, Y.; Hikino, H. Pharmacological actions of aconitine alkaloids. Tohoku J. Exp. Med. 1979, 128, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Hu, J.; Zhang, X.; Zuo, Y.; Wang, Y.; Zhu, S. Research progress of aconitine toxicity and forensic analysis of aconitine poisoning. Forensic Sci. Res. 2020, 5, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Siyiti, M.; Zhang, J.; Yao, M.; Zhao, F. Anti-inflammatory and anti-rheumatic activities in vitro of alkaloids separated from Aconitum soongoricum Stapf. Exp. Ther. Med. 2021, 21, 493. [Google Scholar] [CrossRef]
- Li, X.; Gu, L.; Yang, L.; Zhang, D.; Shen, J. Aconitine: A potential novel treatment for systemic lupus erythematosus. J. Pharmacol. Sci. 2017, 133, 115–121. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huh, E.; Lee, W.; Choi, Y.; Lee, T.H.; Oh, M.S. Geongangbuja-Tang Decoction and Its Active Ingredient, Aconiti Lateralis Radix Preparata, Exerts Inhibitory Effects on Heat Stress-Induced Inflammation in Mice. Appl. Sci. 2021, 11, 6902. https://doi.org/10.3390/app11156902
Huh E, Lee W, Choi Y, Lee TH, Oh MS. Geongangbuja-Tang Decoction and Its Active Ingredient, Aconiti Lateralis Radix Preparata, Exerts Inhibitory Effects on Heat Stress-Induced Inflammation in Mice. Applied Sciences. 2021; 11(15):6902. https://doi.org/10.3390/app11156902
Chicago/Turabian StyleHuh, Eugene, Wonil Lee, Yujin Choi, Tae Hee Lee, and Myung Sook Oh. 2021. "Geongangbuja-Tang Decoction and Its Active Ingredient, Aconiti Lateralis Radix Preparata, Exerts Inhibitory Effects on Heat Stress-Induced Inflammation in Mice" Applied Sciences 11, no. 15: 6902. https://doi.org/10.3390/app11156902