

Untargeted Metabolomic Profiling of Fructus Chebulae and Fructus Terminaliae Billericae



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Instruments and Equipment
2.3. Experimental Methods
2.3.1. Extraction of Metabolites
2.3.2. Sample Analysis
2.3.3. Statistical Analysis
2.4. Data Processing
3. Results
3.1. Phenotypic Characterization
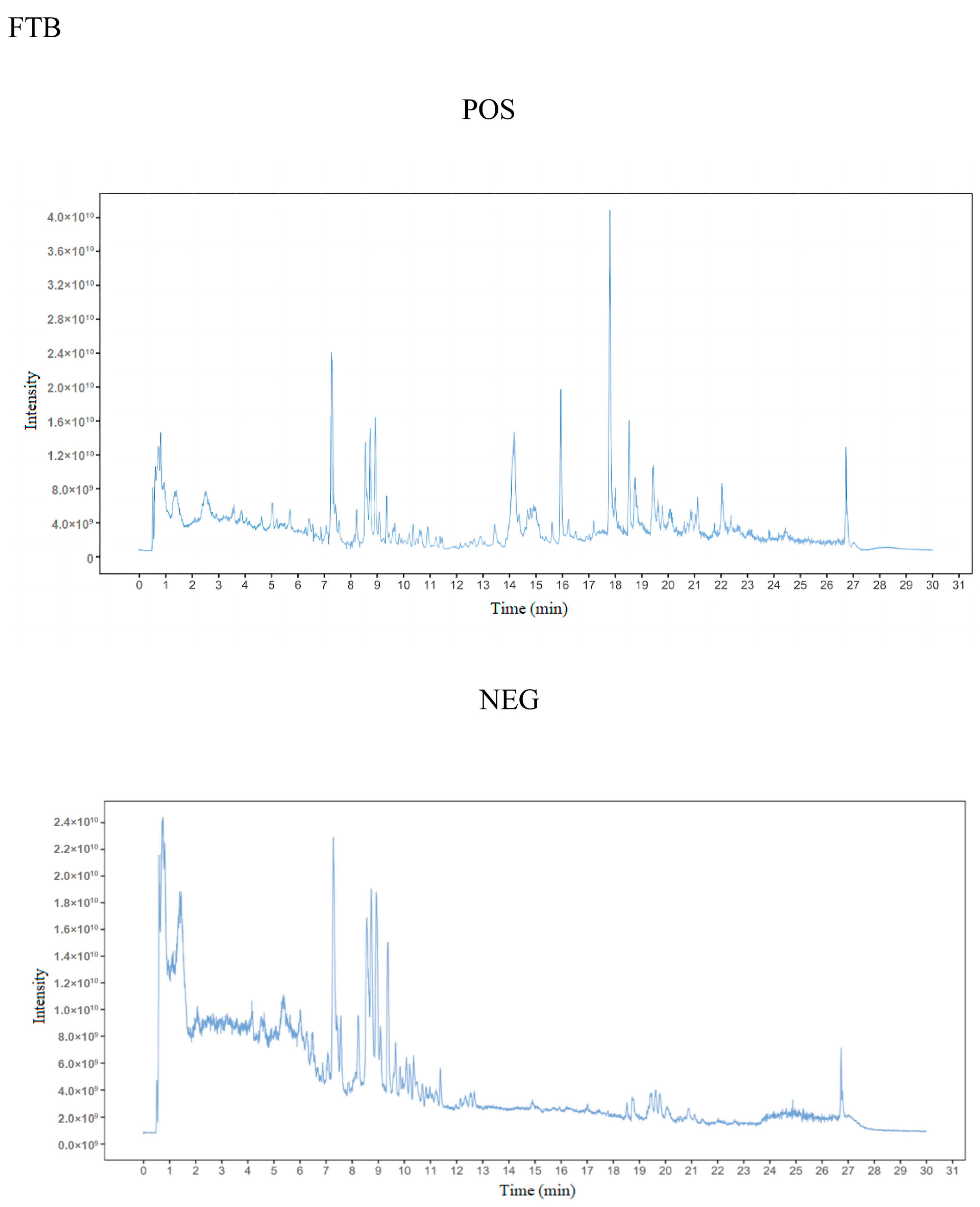
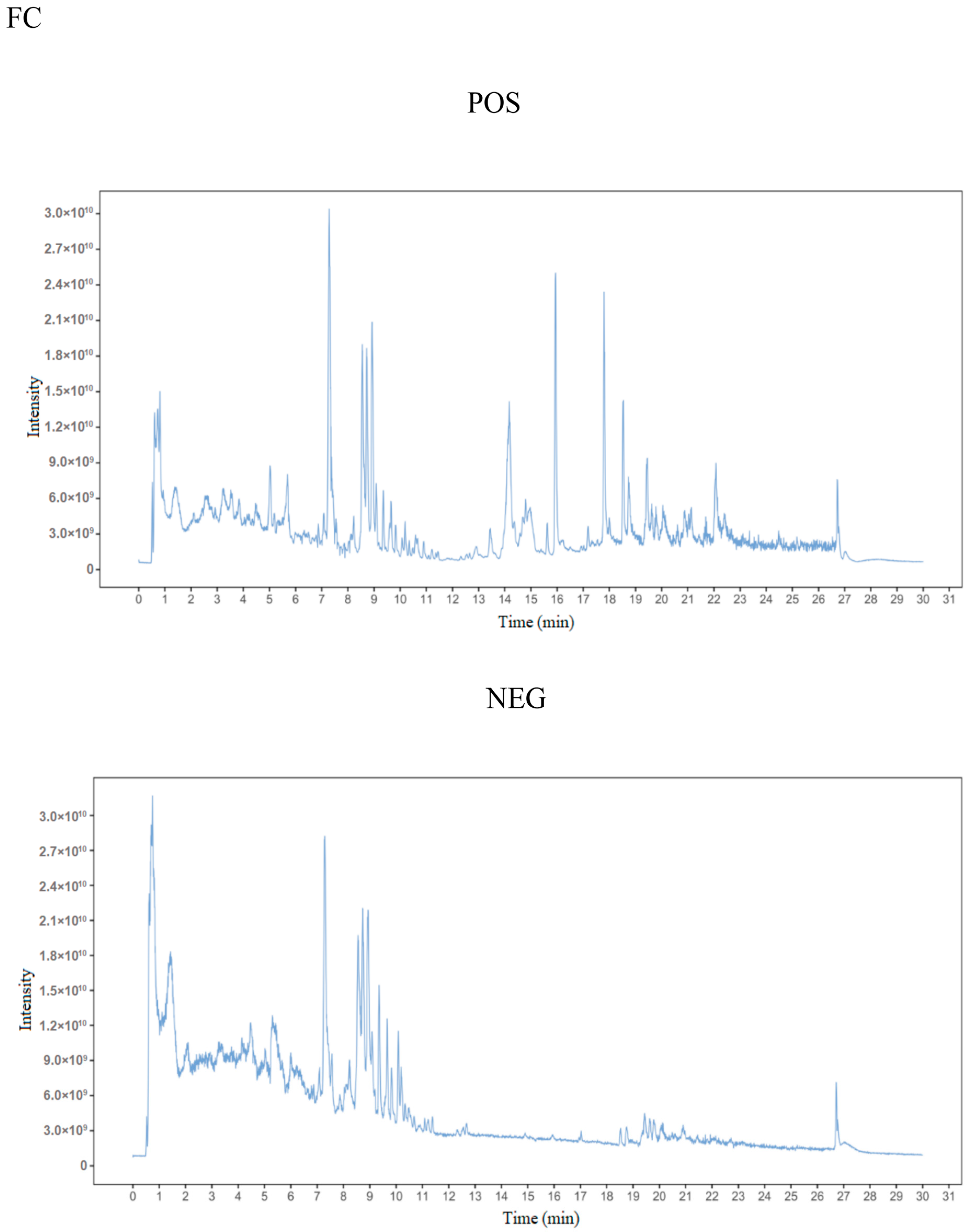
3.2. Metabolite Detection
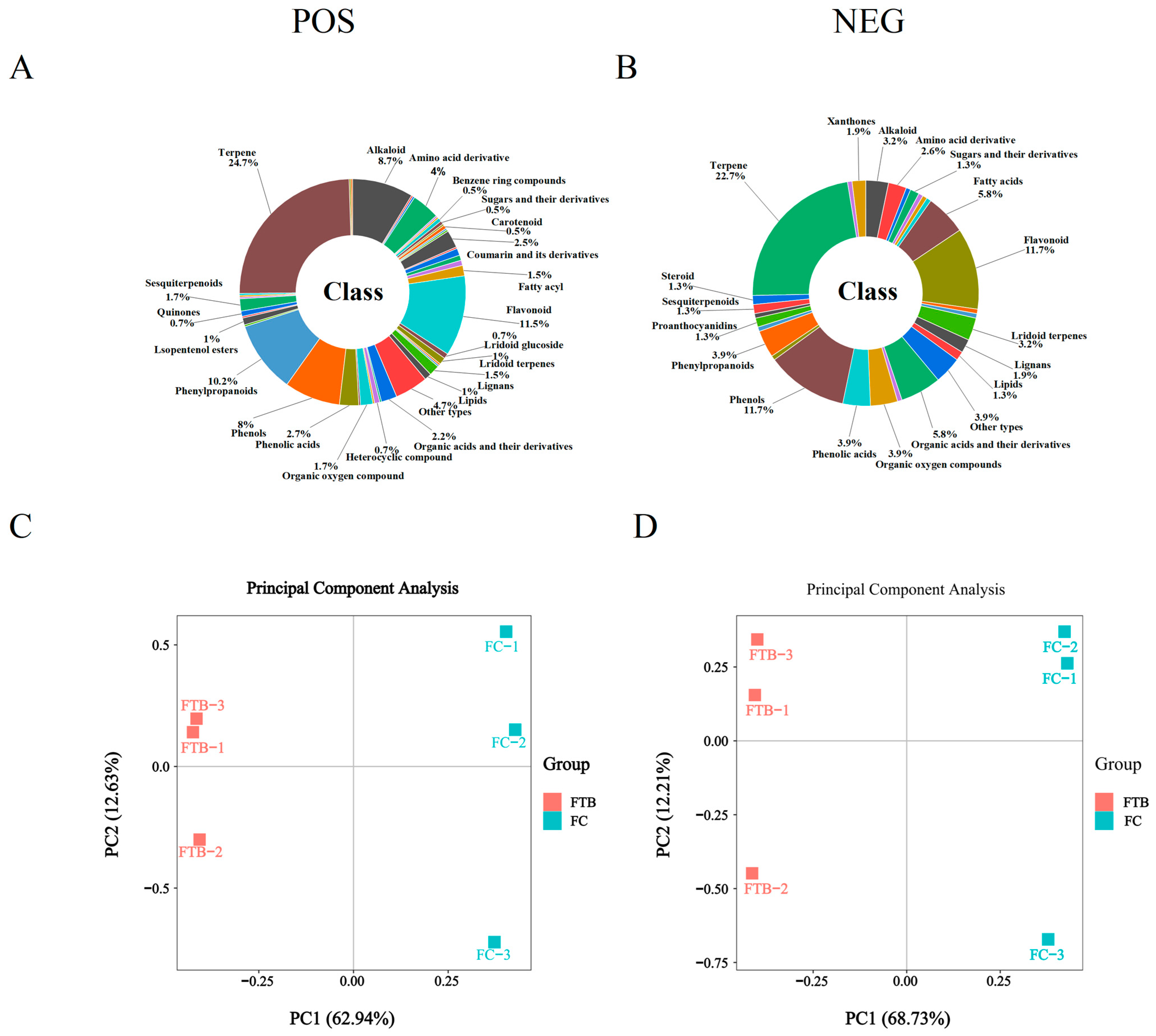
Metabolite Identification and PCA Analysis
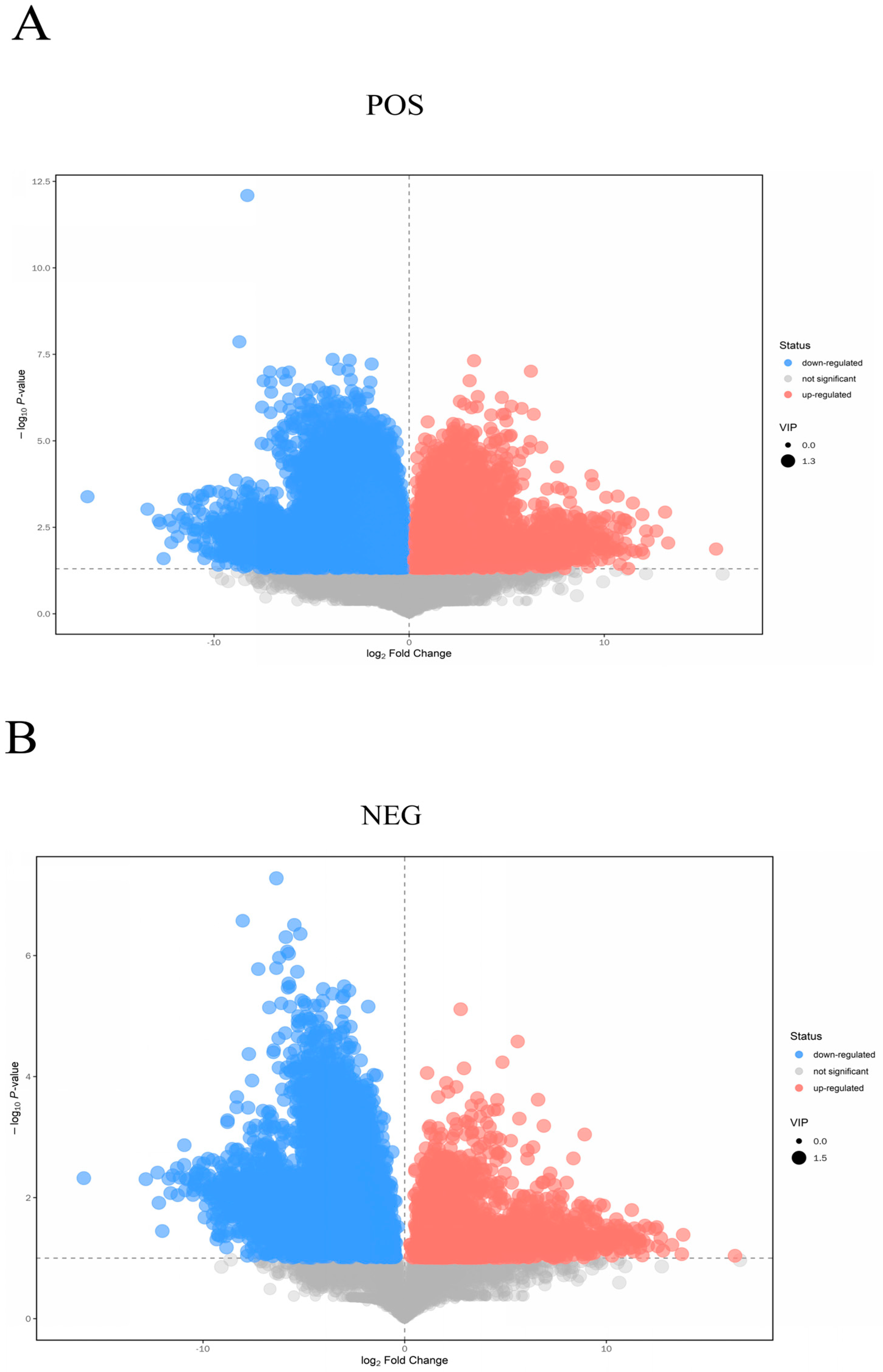
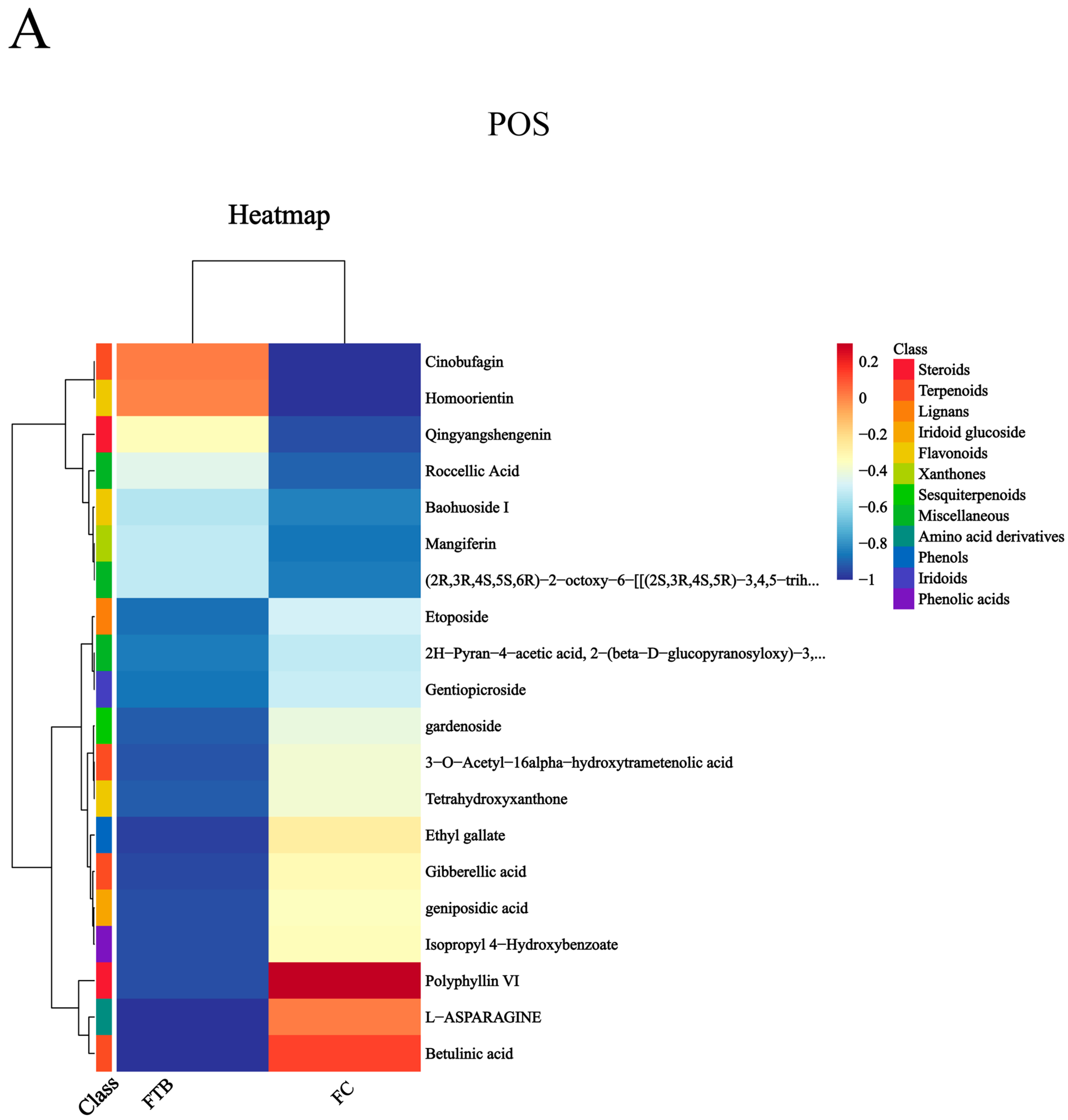
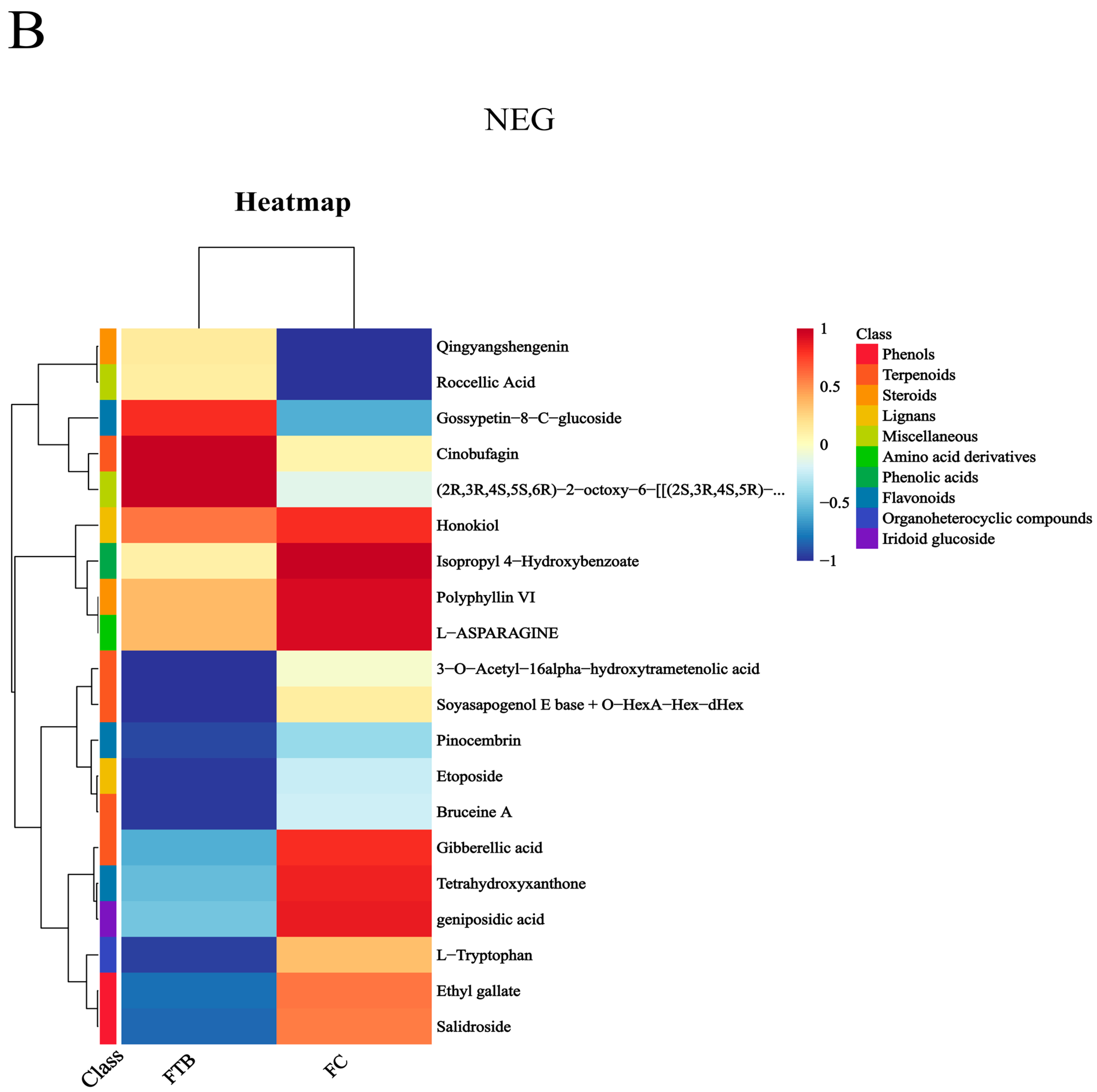
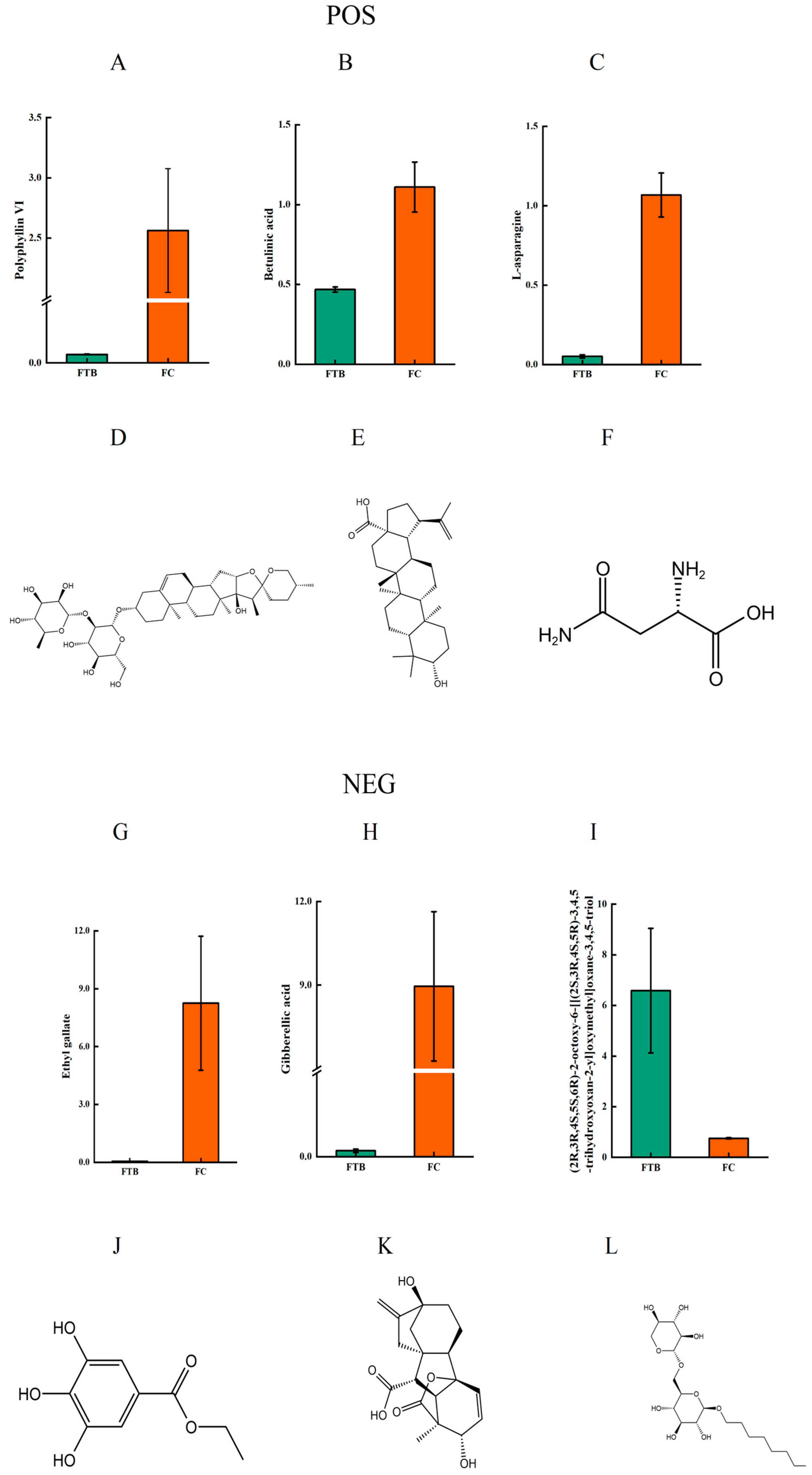
3.3. Analysis of Differential Metabolites
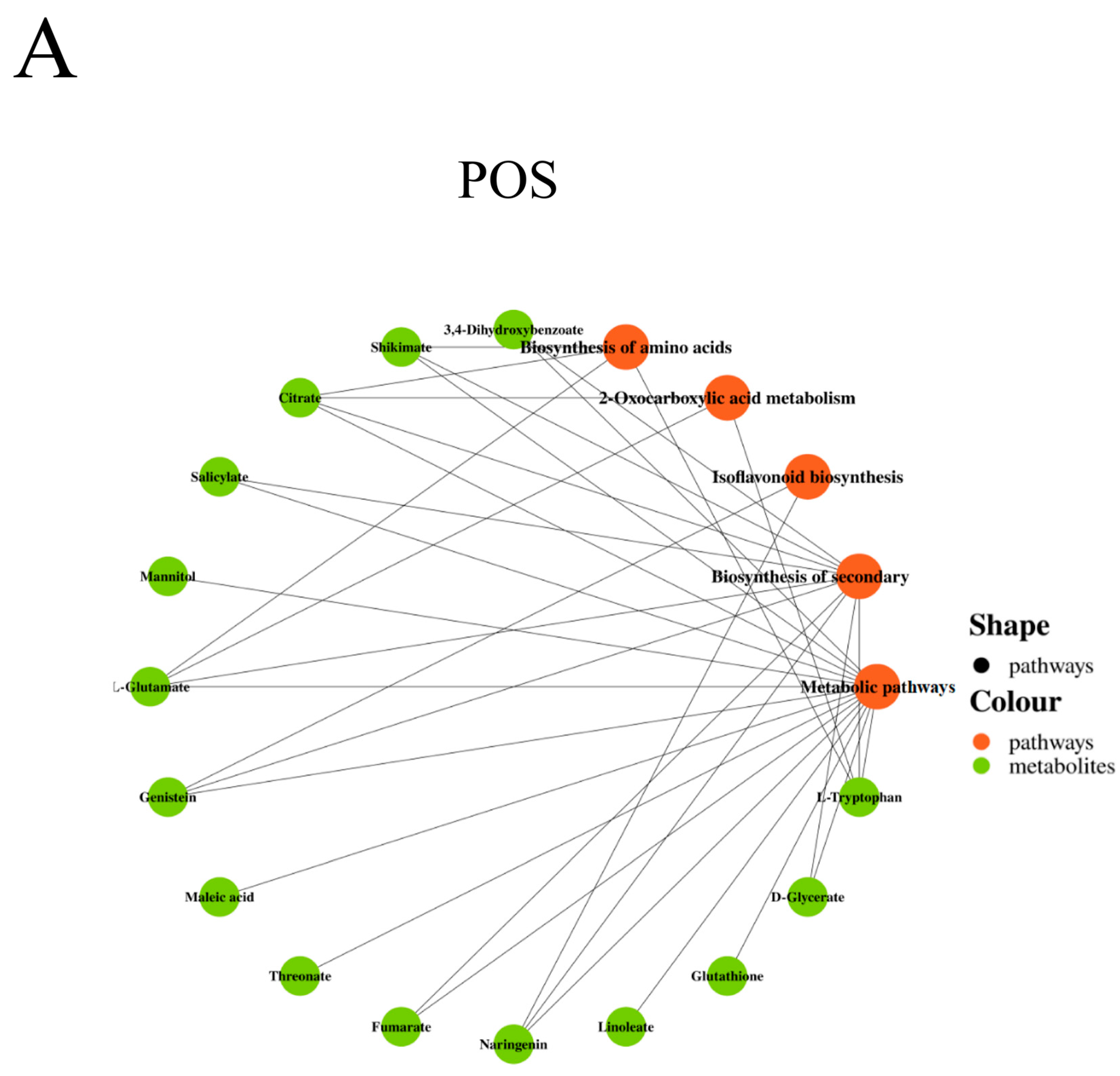
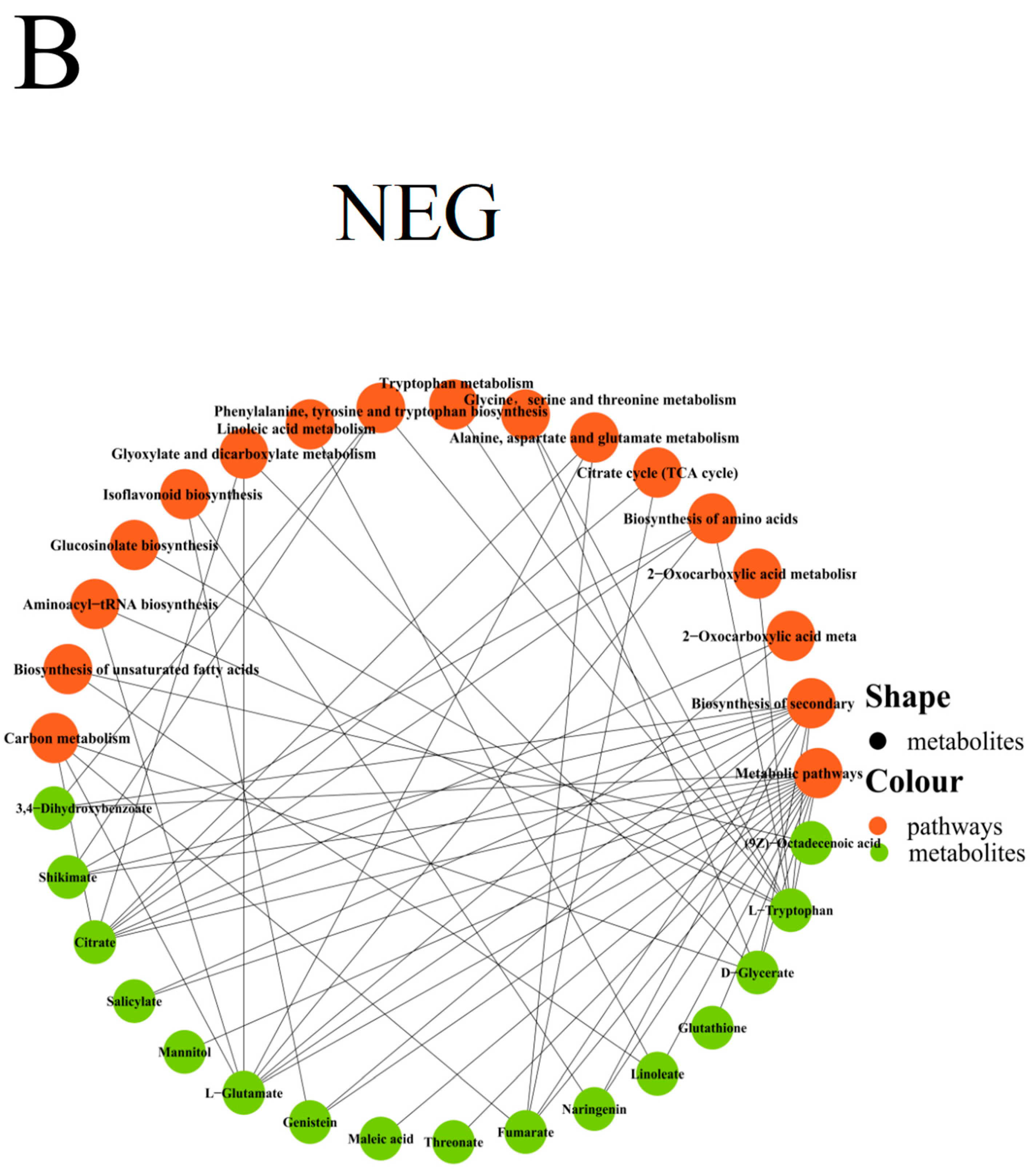
Metabolic Pathway Analysis of Differential Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Liu, X.; Gao, S.; Qian, K.; Liu, Q.; Yin, X. Research on the neuro-protective compounds in Terminalia chebula retz extracts in-vivo by UPLC–QTOF-MS. Acta Chromatogr. 2017, 30, 169–174. [Google Scholar] [CrossRef]
- Singamaneni, V.; Dokuparthi, S.K.; Banerjee, N.; Kumar, A.; Chakrabarti, T. Phytochemical Investigation and Antimutagenic Potential of Ethanolic Extracts of Emblica officinalis, Terminalia chebula and Terminalia bellirica. Nat. Prod. J. 2020, 10, 488–494. [Google Scholar] [CrossRef]
- Muneer, A.; Alhowail, A.; Aldubayan, M.; Rabbani, S.I. The activity of Terminalia chebula Retz. extract on doxorubicin-induced renal damage in rats. J. Pharm. Pharmacogn. Res. 2020, 8, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Zhao, J.; Muhammad, I.; Zhang, Y. Optimization of total phenolic content from Terminalia chebula Retz. fruits using response surface methodology and evaluation of their antioxidant activities. PLoS ONE 2018, 13, e0202368. [Google Scholar] [CrossRef] [PubMed]
- Ekambaram, S.P.; Babu, K.B.; Perumal, S.S.; Rajendran, D. Repeated oral dose toxicity study on hydrolysable tannin rich fraction isolated from fruit pericarps of Terminalia chebula Retz in Wistar albino rats. Regul. Toxicol. Pharmacol. 2018, 92182–92188. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gesang, S.; Wu, J. Review of plant classification and pharmaceutical characteristics of Quixote, Quixote and Gansu. Appl. Med. Biotechnol. China 2004, 01, 14–28. [Google Scholar]
- Li, D.; Zhu, W.; Duan, S.; Lai, Y. Determination of half-lethal amount of bark alcohol extract and its effect on intestinal muscle. J. Dali Univ. 2014, 13, 10–13. [Google Scholar]
- Zhou, K.; Jian, P.; Liang, W.; Liang, L.; Ye, T.; Chang, Z.; Zhang, Q.; Zhang, L. Analysis of the chemical composition of the Tibetan herbs and the Tibetan Quixote based on UPLC-Q-Exactive Orbitrap-MS. J. Chin. Mass Spectrom. 2020, 41, 254–267. [Google Scholar]
- Lee, H.S.; Jung, S.H.; Yun, B.S.; Lee, K.W. lsolation of chebulic acid from Terminalia chebula Retz and its antioxidant effect in isolated rat hepatocytes. Arch Toxicol 2007, 81, 211–218. [Google Scholar] [CrossRef]
- Lin, M.C.; Yin, M.C. Preventive effects of ellagic acid against doxorubicin-induced cardio-toxicity in mice. Cardiovasc. Toxicol. 2013, 13, 185–193. [Google Scholar] [CrossRef]
- Lee, J.; Nho, Y.H.; Yun, S.K.; Hwang, Y.S. Use of ethanol extracts of Terminalia chebula to prevent periodontal disease induced by dental plaque bacterial. BMC Complement. Altern. Med. 2017, 17, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.S.; Ankola, A.V.; Metgud, S.C.; Bolmal, U.K. An in vitro study to determine the effect of Terminalia chebula extract and its formulation on Streptococcus mutans. J. Contemp. Dent. Pract. 2014, 15, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Shankara, B.R.; Ramachandra, Y.L.; Rajan, S.S.; Ganapathy, P.S.; Yarla, N.S.; Richard, S.A.; Dhananjaya, B.L. Evaluating the anticancer potential of ethanolic gall extract of Terminalia chebula (Gaertn.) Retz. (combretaceae). Pharmacogn. Res. 2016, 8, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Messeha, S.S.; Zarmouh, N.O.; Taka, E.; Gendy, S.G.; Shokry, G.R.; Kolta, M.G.; Soliman, K.F.A. The role of monocarboxylate transporters and their chaperone CD147 in lactate efflux inhibition and the anticancer effects of Terminalia chebula in neuroblastoma cell line N2-A. Eur. J. Med. Plants 2016, 12, EJMP.23992. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Saha, A.K.; Das, S.K.; Kar, N.B. Antijuvenoid Action of Terpenoid Imidazole Compound on Larval–Pupal–Adult Development of Silkworm, Bombyx mori L. Int. J. Ind. Entomol. 2007, 14, 127–135. [Google Scholar]
- Al-Maharik, N.; Jaradat, N.; Bassalat, N.; Hawash, M.; Zaid, H. Isolation, Identification and Pharmacological Effects of Mandragora autumnalis Fruit Flavonoids Fraction. Molecules 2022, 27, 1046. [Google Scholar] [CrossRef] [PubMed]
- Razola-Díaz, M.D.C.; Aznar-Ramos, M.J.; Benítez, G.; Gómez-Caravaca, A.M.; Verardo, V. Exploring the potential of phenolic and antioxidant compounds in new Rosaceae fruits. J. Sci. Food Agric. 2024, 104, 3705–3718. [Google Scholar] [CrossRef] [PubMed]
- Niu, F.; Du, Y.; Huang, Y.; Zhou, H.; Liu, J. Progress in the synthesis of phenylpropanoid acid compounds and its derivatives by engineered microorganisms. Synth. Biol. 2020, 1, 337–357. [Google Scholar]
- Meng, Q.; Liang, J.; Wu, G.; Lu, H. Progress in the pharmacological effects of alkaloid compounds. Shizhen Natl. Med. 2003, 11, 700–702. [Google Scholar]
- Saccenti, E.; Hoefsloot, H.C.; Smilde, A.K.; Westerhuis, J.A.; Hendriks, M.M. Reflections on univariate and multivariate analysis of metabolomics data. Metabolomics 2014, 10, 361–374. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps, R Package version 061; The Comprehensive R Archive Network (CRAN): Vienna, Austria, 2015.
- Zhao, X.; Li, Y.; Zhang, J. Effect of betulinic acid 51 on oxidative damage and apoptosis of PC12H2O. J. Med. Mol. Biol. 2021, 18, 428–433. [Google Scholar]
- Li, Y.R.; Cao, W.; Guo, J.; Miao, S.; Ding, G.R.; Li, K.C.; Wang, J.; Guo, G.Z. Comparative investigations on the protective effects of rhodioside, ciwujianoside-B and astragaloside IV on radiation injuries of the hematopoietic system in mice. Phytother. Res. 2011, 25, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Wang, M.; Yuan, J.; Liu, J. Effect of ethyl gallate on invasion abilities and its mechanism of breast cancer MDA-MB-231 cells. Yao Xue Xue Bao = Acta Pharm. Sin. 2015, 50, 45–49. [Google Scholar]
- Liu, Q.; Shi, Z.; Wu, J.; Huang, B.; Liang, X.; Cui, X.; Hu, S.; Niu, S.; Kong, W. Analysis of associated substances and major metabolic pathways for the development of pellagra lateral ear fruiting bodies based on non-target metabolomics. J. Bact. 2024, 43, 59–73. [Google Scholar] [CrossRef]
- Dai, M.; Jiang, Z.; Huang, X. Progress in the physiological function of tryptophan and metabolites and its role in disease. Zhongnan Pharm. 2021, 19, 909–915. [Google Scholar]
- Keranmu, A.; Pan, L.B.; Fu, J.; Han, P.; Yu, H.; Zhang, Z.W.; Xu, H.; Yang, X.-Y.; Hu, J.-C.; Zhang, H.-J.; et al. Biotransformation of Liquiritigenin into Characteristic Metabolites by the Gut Microbiota. Molecules 2022, 27, 3057. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.N.; Huang, F.Y.; Wang, S.P.; Kumar, R. Synergistic Antioxidant and Antibacterial Activity of Curcumin-c3 Encapsulated Chitosan Nanoparticles. Curr. Pharm. Des. 2020, 26, 5021–5029. [Google Scholar] [CrossRef] [PubMed]
- Luan, X.; Ou, W.; Hu, J.; Lu, J. Gallic acid alleviates lipopolysaccharide-induced renal injury in rats by inhibiting cell pro-death and inflammatory response and its mechanism. Cell. Mol. Biol. 2023, 69, 31–35. [Google Scholar] [CrossRef]
- Aleixandre, A.; Gil, J.V.; Sineiro, J.; Rosell, C.M. Understanding phenolic acids inhibition of a-amylase and c-glucosidase and influence of reaction conditions. Food Chem. 2022, 372, 1231. [Google Scholar] [CrossRef]
- Xu, H.; Lyu, X.; Guo, X.; Yang, H.; Duan, L.; Zhu, H.; Pan, H.; Gong, F.; Wang, L. Distinct AMPK-Mediated FAS/HSL Pathway Is Implicated in the Alleviating Effect of Nuciferine on Obesity and Hepatic Steatosis in HFD-Fed Mice. Nutrients 2022, 14, 1898. [Google Scholar] [CrossRef]
- Bolik, S.; Schlaich, A.; Mukhina, T.; Amato, A.; Bastien, O.; Schneck, E.; Demé, B.; Jouhet, J. Lipid bilayer properties potentially contributed to the evolutionary disappearance of betaine lipids in seed plants. BMC Biol. 2023, 21, 275. [Google Scholar] [CrossRef] [PubMed]
- Whaun, M.J.; Brown, D.N. Treatment of chloroquine-resistant malaria with esters of cephalotaxine: Homoharringtonine. Ann. Trop. Med. Parasitol. 2016, 84, 229–237. [Google Scholar] [CrossRef]
- Cui, W.; Bai, X.; Wang, J.; Jin, H. Progress in the regulation of abscisic acid and the biosynthesis of secondary metabolites related to plant disease resistance. Bot. Med. 2022, 1, 1–11. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Fresh Leaf Mass (g) | Dry Leaf Mass (g) | Fresh Fruit Mass (g) | Dry Fruit Mass (g) |
|---|---|---|---|---|
| FTB | 1.27 ± 0.11 b | 0.78 ± 0.01 b | 13.25 ± 5.06 a | 5.17 ± 1.65 a |
| FC | 1.69 ± 0.07 a | 0.85 ± 0.02 a | 10.20 ± 1.58 a | 3.91 + 0.58 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Gong, H. Untargeted Metabolomic Profiling of Fructus Chebulae and Fructus Terminaliae Billericae. Appl. Sci. 2024, 14, 3123. https://doi.org/10.3390/app14073123
Song Y, Gong H. Untargeted Metabolomic Profiling of Fructus Chebulae and Fructus Terminaliae Billericae. Applied Sciences. 2024; 14(7):3123. https://doi.org/10.3390/app14073123
Chicago/Turabian StyleSong, Yuman, and Hede Gong. 2024. "Untargeted Metabolomic Profiling of Fructus Chebulae and Fructus Terminaliae Billericae" Applied Sciences 14, no. 7: 3123. https://doi.org/10.3390/app14073123
APA StyleSong, Y., & Gong, H. (2024). Untargeted Metabolomic Profiling of Fructus Chebulae and Fructus Terminaliae Billericae. Applied Sciences, 14(7), 3123. https://doi.org/10.3390/app14073123