

Simulated Digestion of Red Sea Cucumber (Parastichopus tremulus): A Study of Protein Quality and Antioxidant Activity

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
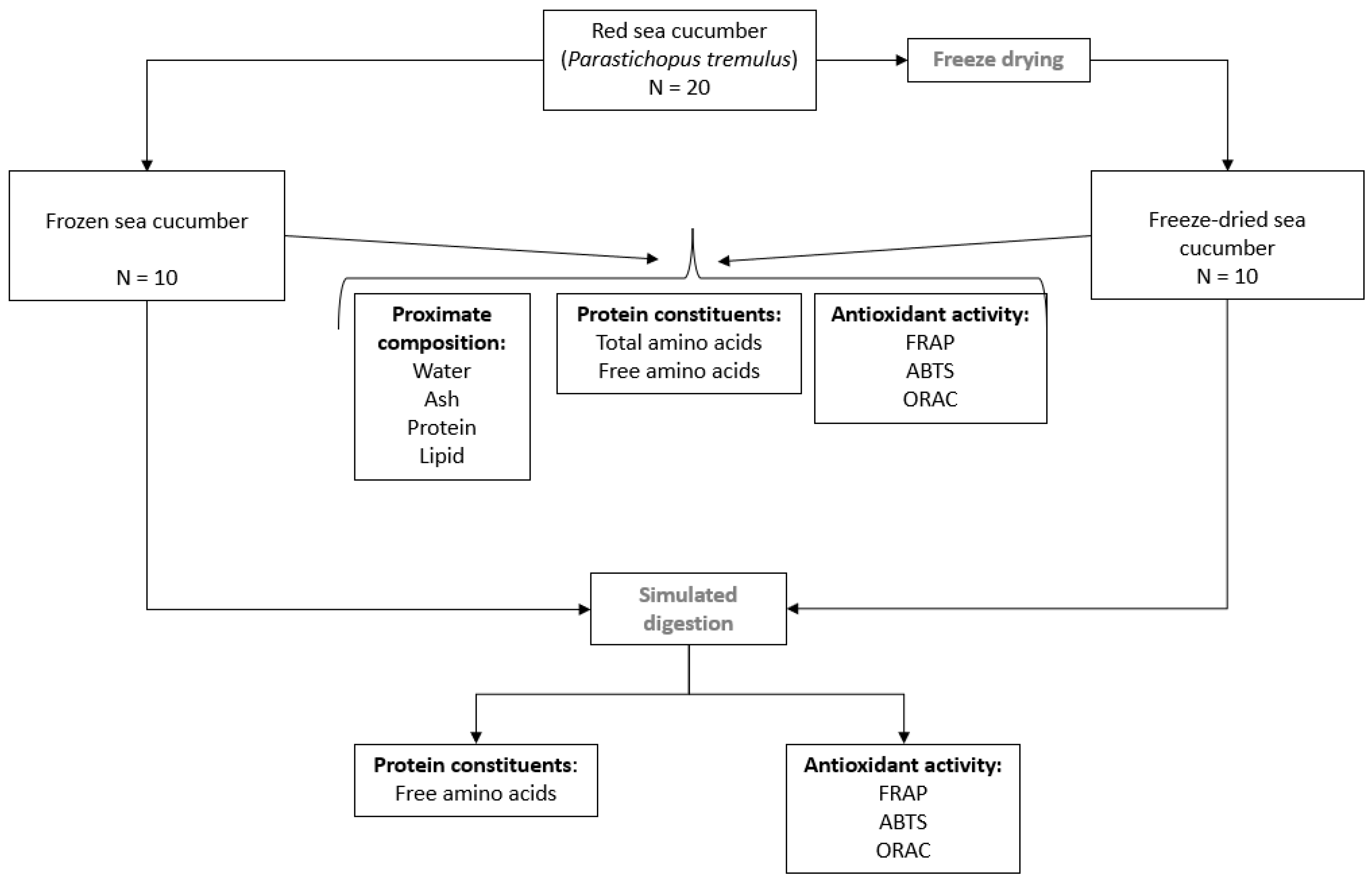
2.1. Study Design
2.2. Proximate Composition of Frozen and Freeze-Dried Sea Cucumbers
2.3. Analyses of Total and Free Amino Acids
2.4. Calculation of True Retention
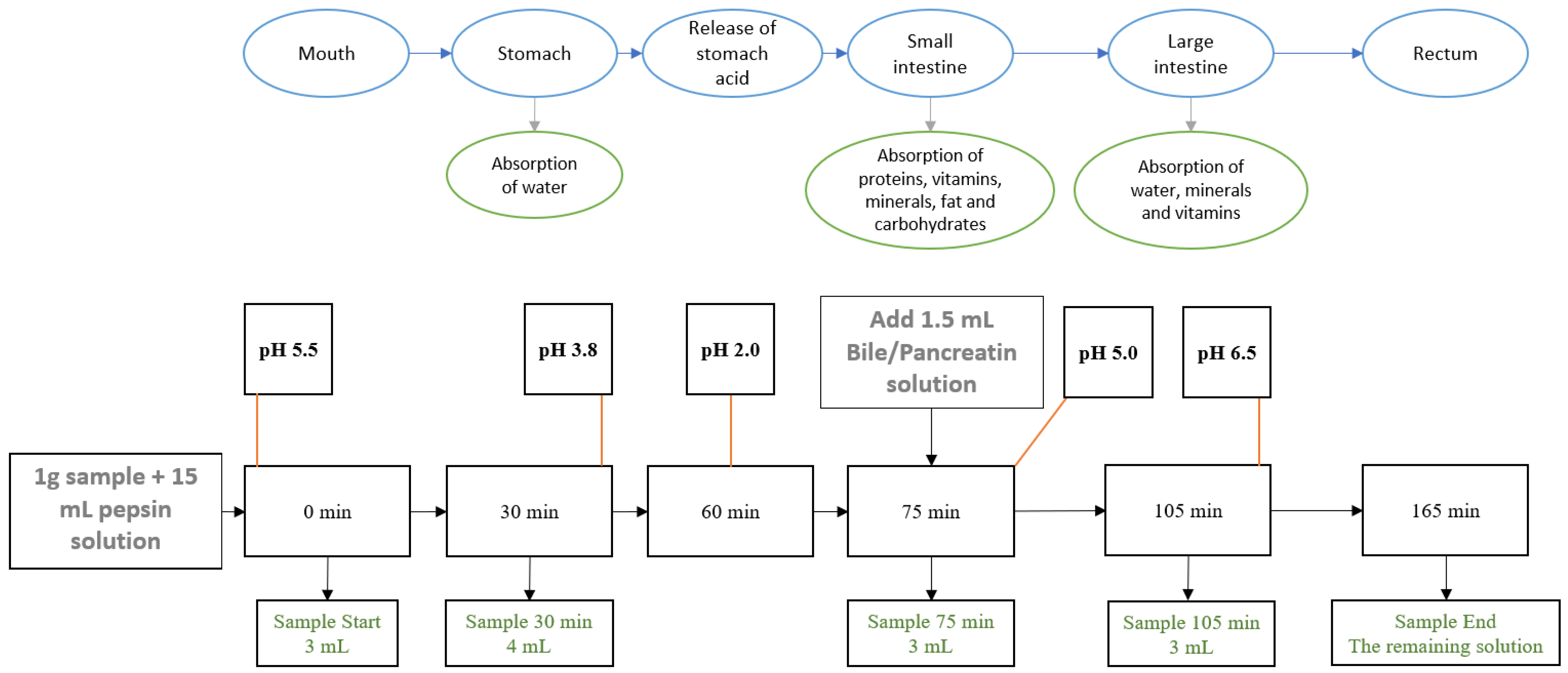
2.5. Simulated Gastrointestinal Digestion
2.6. Antioxidant Assays
2.7. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition of Frozen and Freeze-Dried Materials
3.2. Total and Free Amino Acid Composition in Frozen and Freeze-Dried Materials
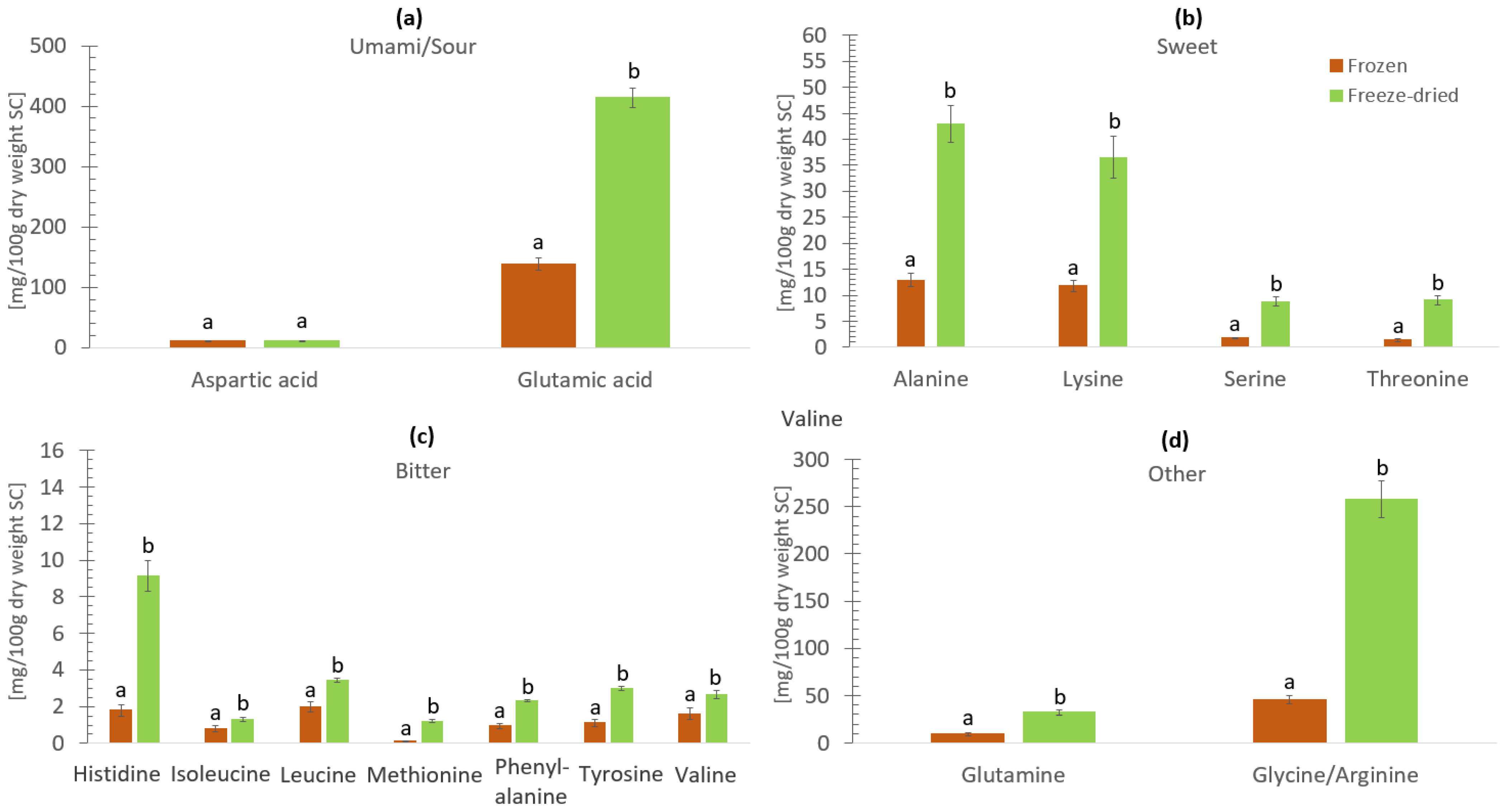
3.3. Free Amino Acids and Umami Flavor in Frozen and Freeze-Dried Materials
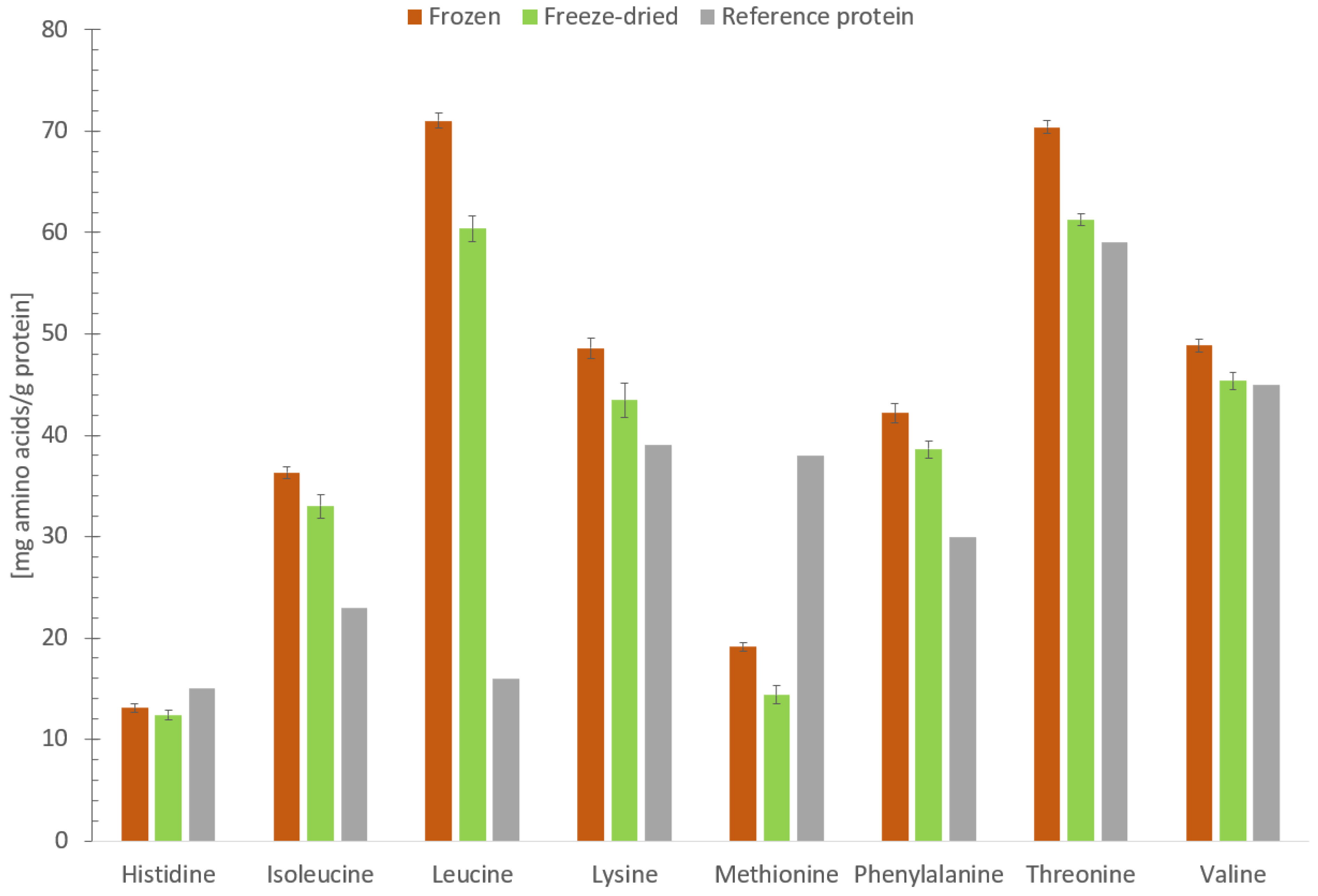
3.4. Protein Quality in Frozen and Freeze-Dried Materials
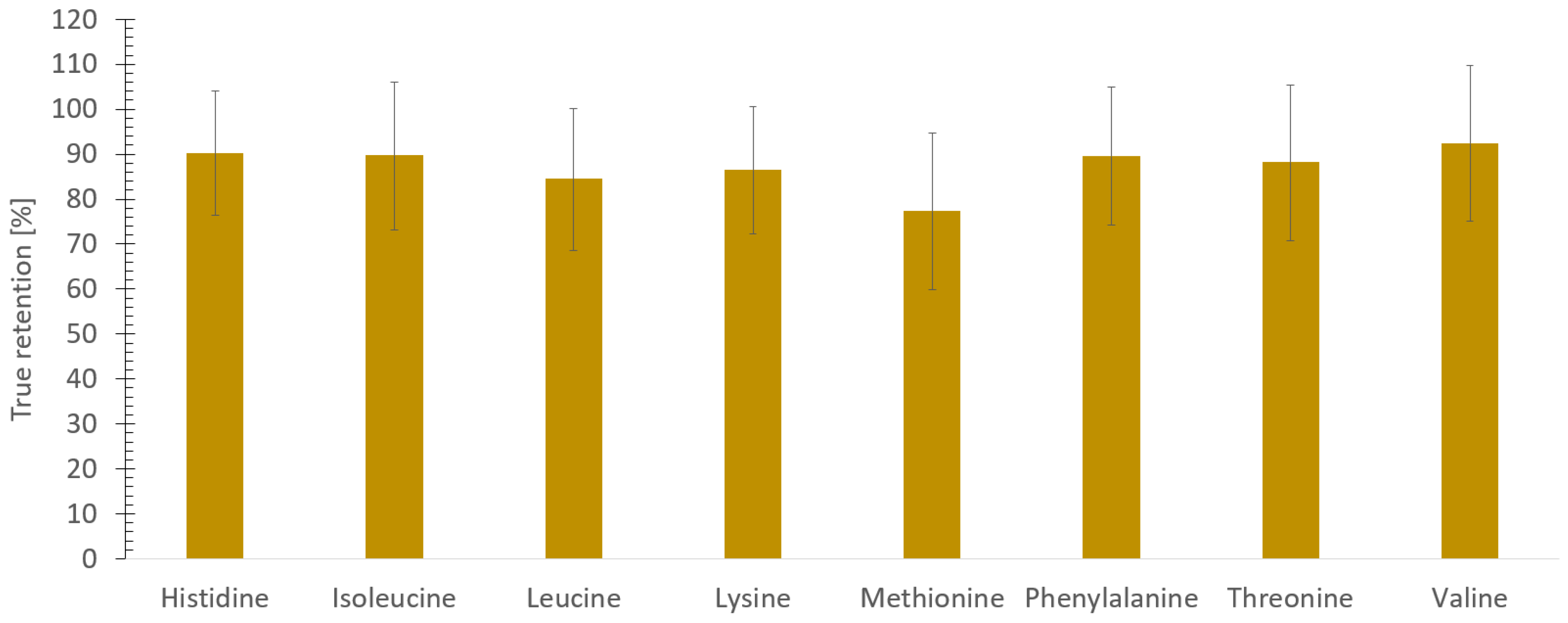
3.5. True Retention of Amino Acids during Freeze-Drying of Sea Cucumber
3.6. Antioxidant Activity in Frozen and Freeze-Dried Materials
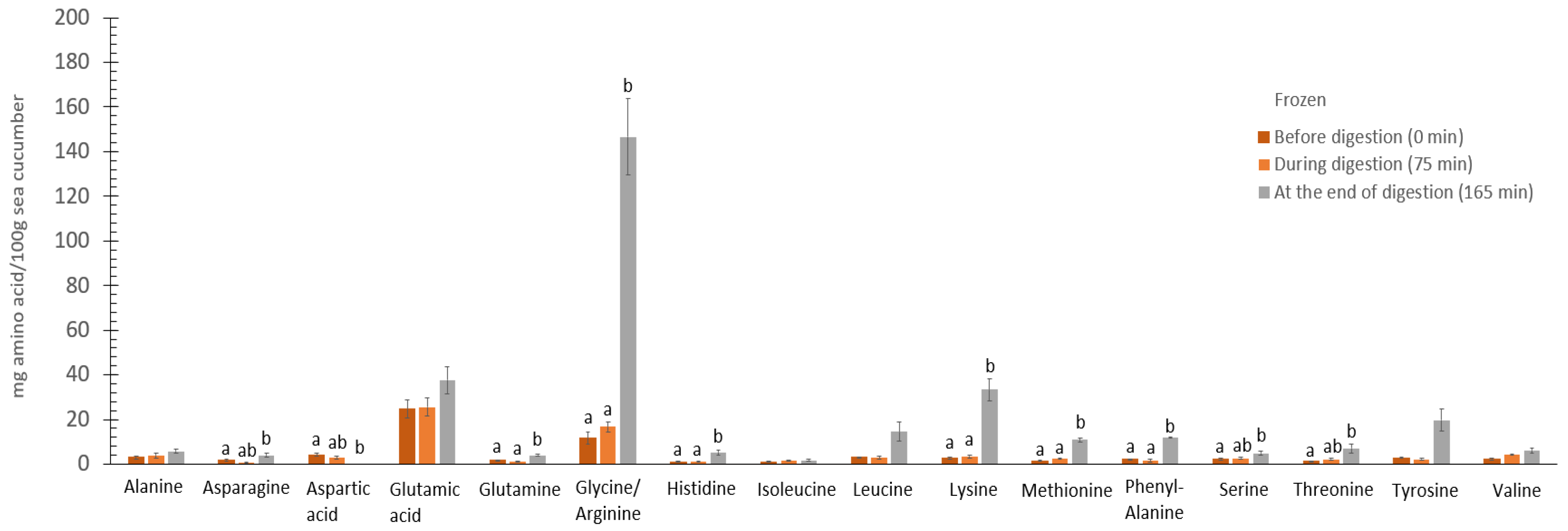
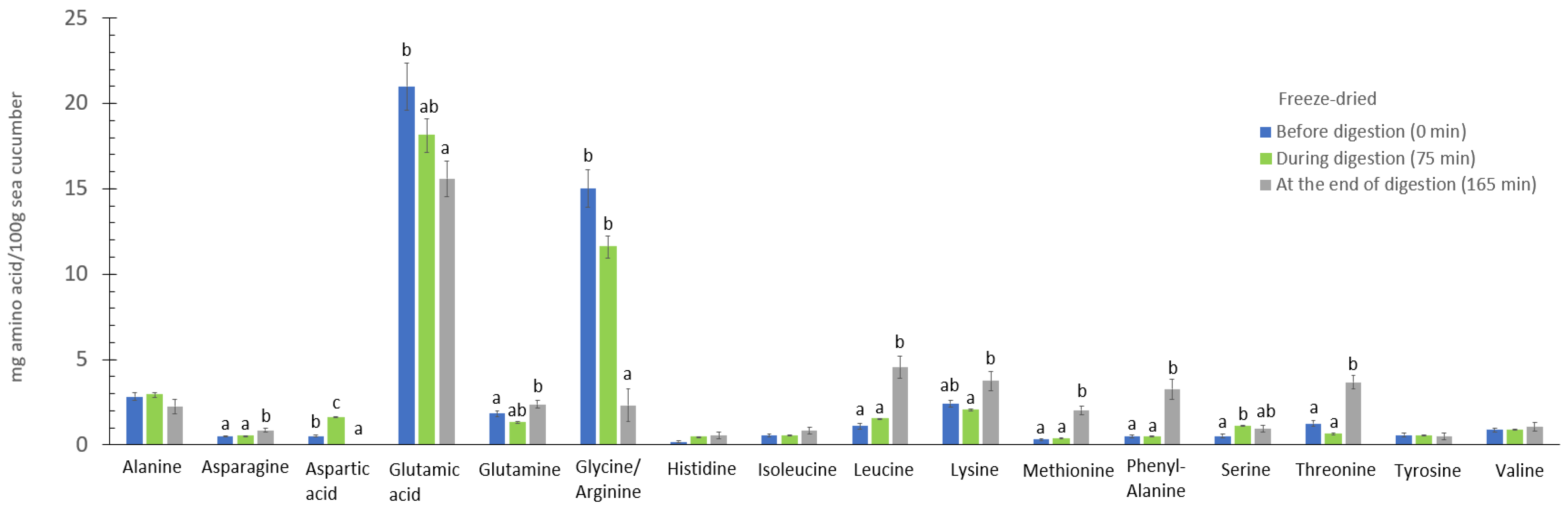
3.7. The Digestive Effect on Amino Acids
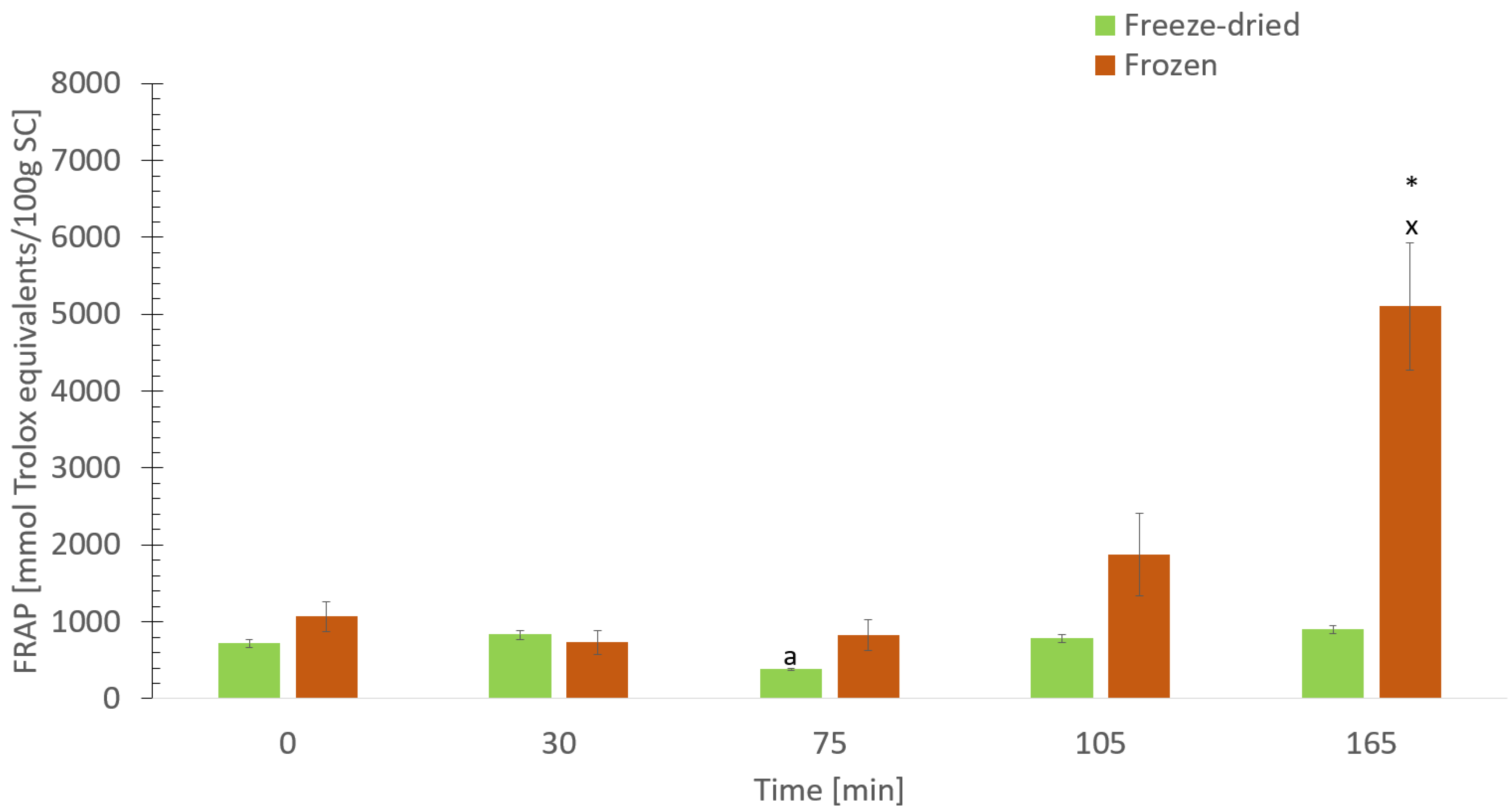
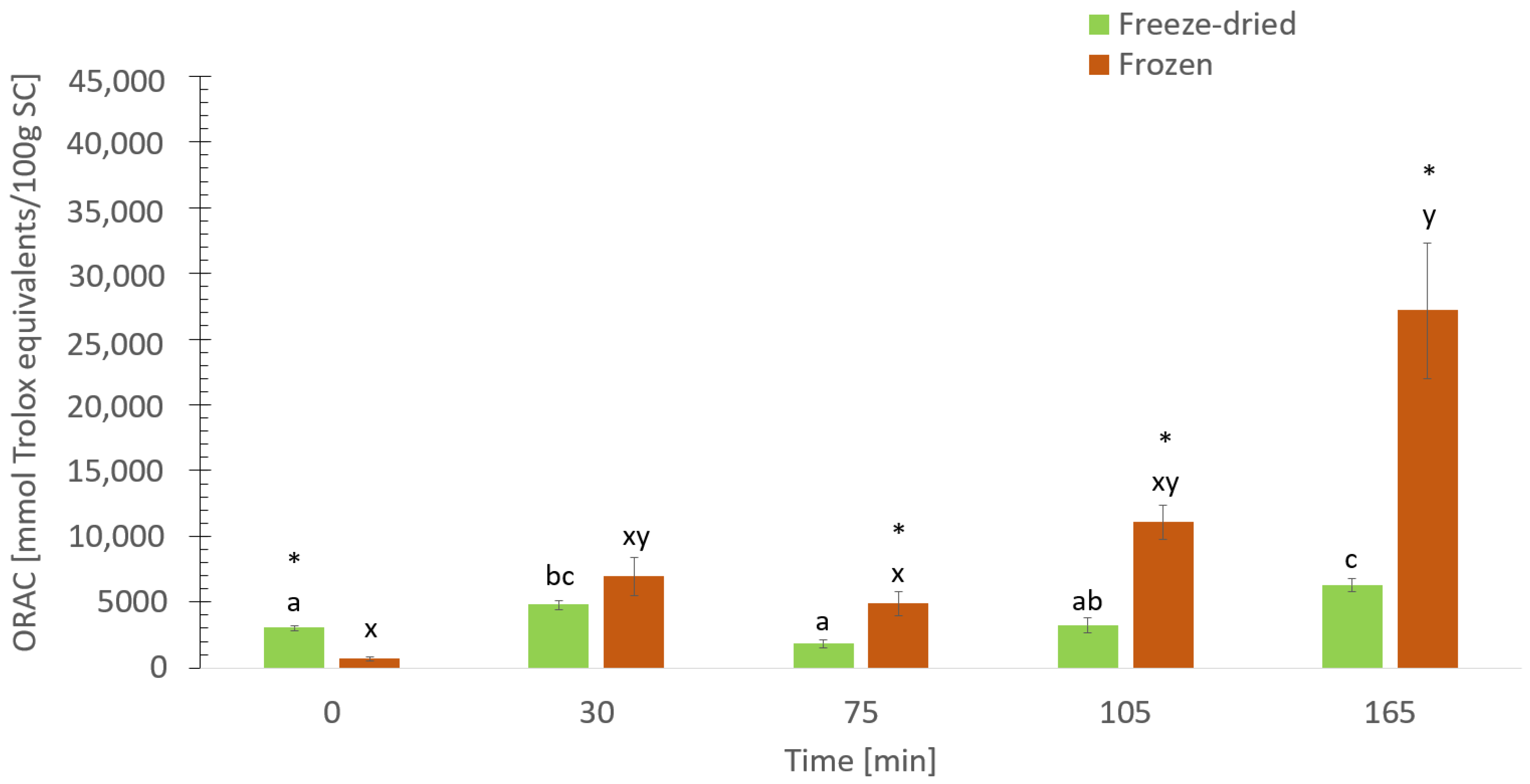
3.8. The Digestive Effect on Antioxidant Activity
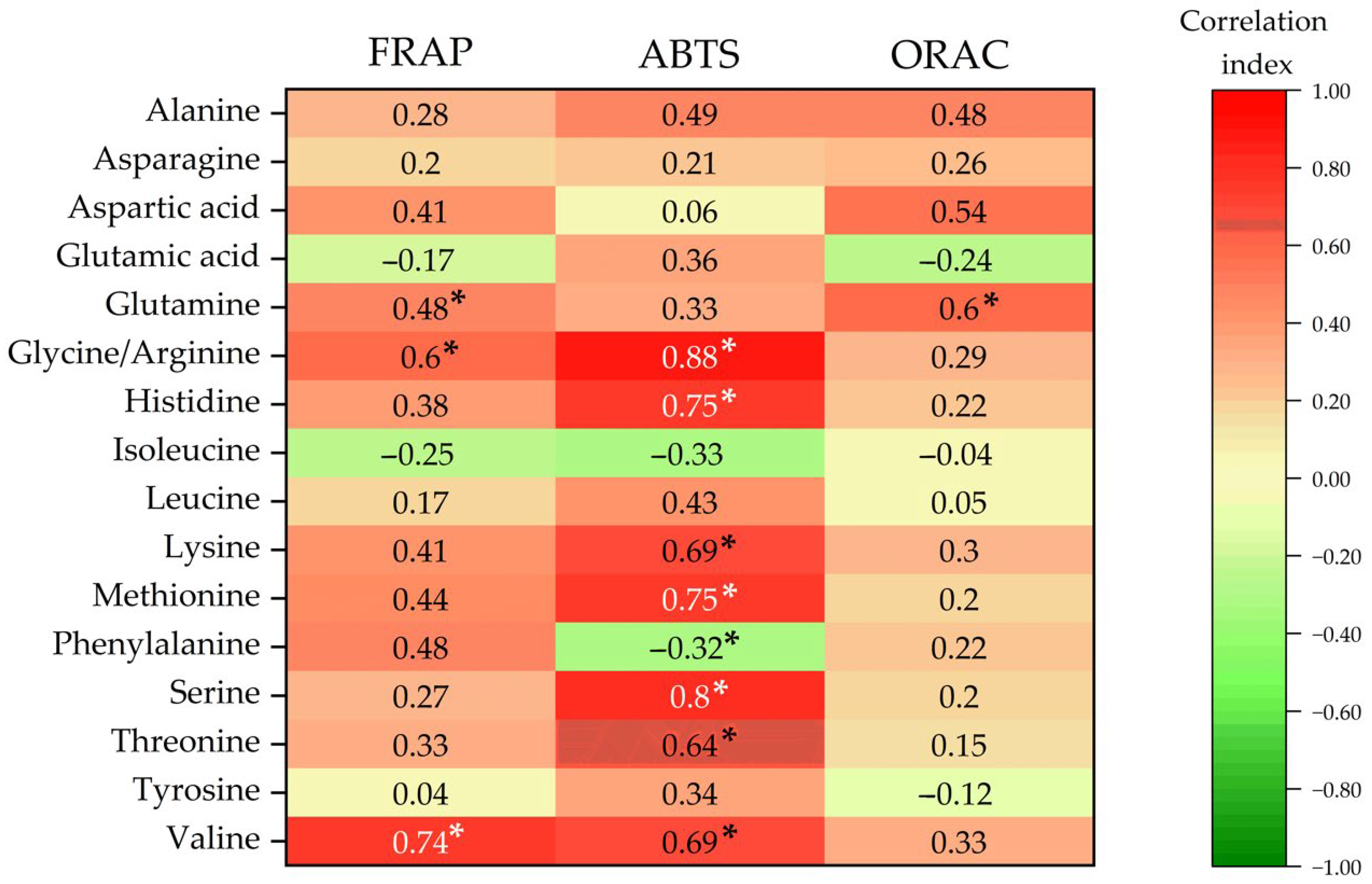
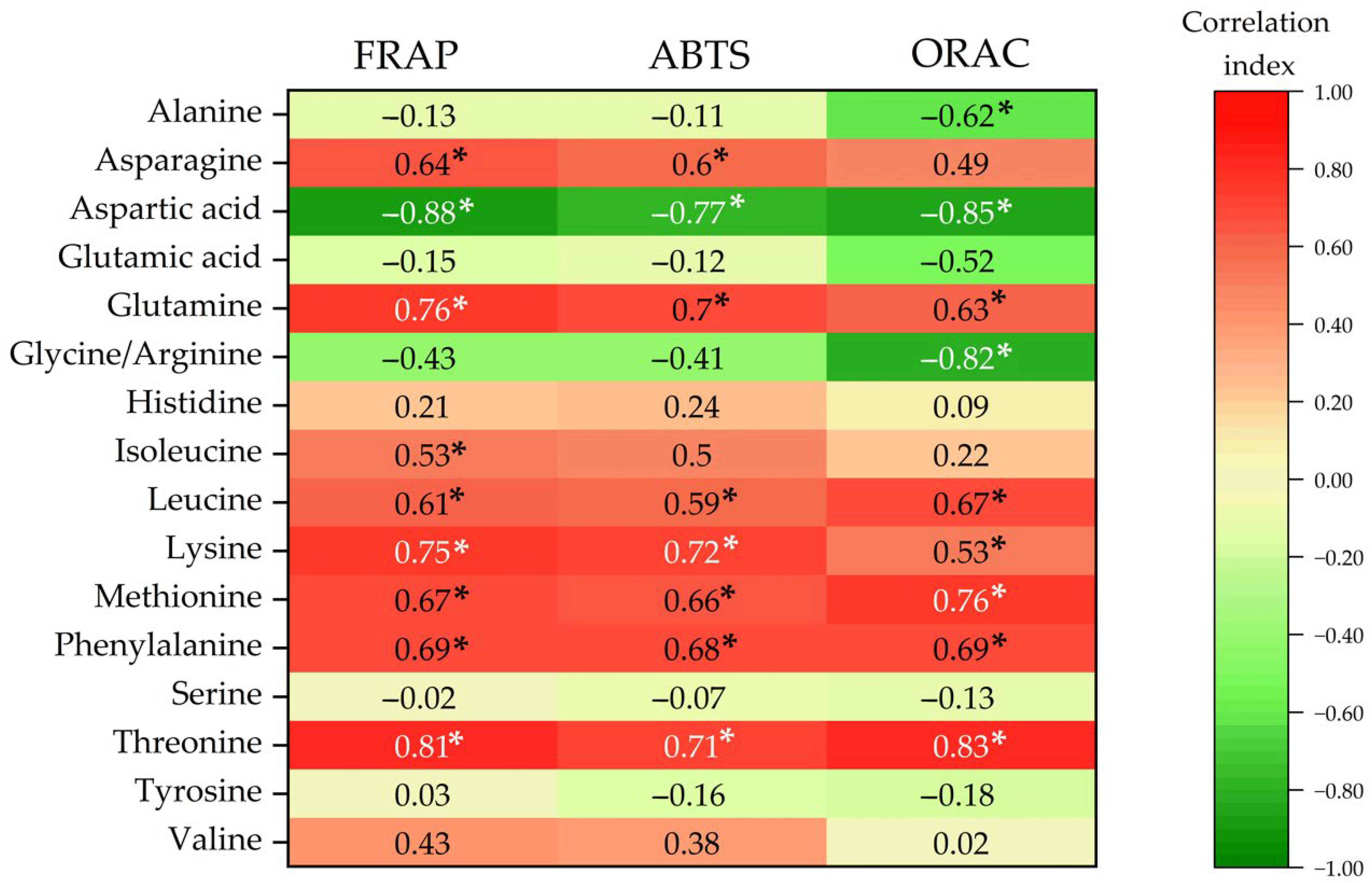
3.9. Release of Free Amino Acids Contributing to Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lobine, D.; Rengasamy, K.R.; Mahomoodally, M.F. Functional foods and bioactive ingredients harnessed from the ocean: Current status and future perspectives. Crit. Rev. Food Sci. Nutr. 2022, 62, 5794–5823. [Google Scholar] [CrossRef] [PubMed]
- Admassu, H.; Zhao, W.; Yang, R.; Gasmalla, M.; Alsir, E. Development of functional foods: Seaweeds (algae) untouched potential and alternative resource—A review. Int. J. Sci. Technol. Res. 2015, 4, 108–115. [Google Scholar]
- Golden, C.D.; Koehn, J.Z.; Shepon, A.; Passarelli, S.; Free, C.M.; Viana, D.F.; Matthey, H.; Eurich, J.G.; Gephart, J.A.; Fluet-Chouinard, E. Aquatic foods to nourish nations. Nature 2021, 598, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L. Ranching sea cucumbers to repair the oceans—And improve human health. Nature 2023, 617, 856. [Google Scholar] [CrossRef]
- Wirawati, I.; Pratiwi, R.; Widyastuti, E.; Ibrahim, P.S. Commercial sea cucumber trading status in Indonesia. Aquac. Aquar. Conserv. Legis. 2021, 14, 3204–3216. [Google Scholar]
- Kjerstad, M.; Bjørkevoll, I.; Christophersen, G. Sesonal Variation in Quality of Red Sea Cucumber—A Novel Seafood Product in Møre and Romsdal; Report No. MA 20-02; Møreforskning AS: Ålesund, Norway, 2020; pp. 1–51. [Google Scholar]
- Rasyid, A.; Yasman, Y.; Putra, M.Y. Current prospects of nutraceutical and pharmaceutical use of sea cucumbers. Pharmacia 2021, 68, 561. [Google Scholar]
- Purcell, S.W.; Mercier, A.; Conand, C.; Hamel, J.F.; Toral-Granda, M.V.; Lovatelli, A.; Uthicke, S. Sea cucumber fisheries: Global analysis of stocks, management measures and drivers of overfishing. Fish Fish. 2013, 14, 34–59. [Google Scholar] [CrossRef]
- Uthicke, S.; Welch, D.; Benzie, J. Slow growth and lack of recovery in overfished holothurians on the Great Barrier Reef: Evidence from DNA fingerprints and repeated large-scale surveys. Conserv. Biol. 2004, 18, 1395–1404. [Google Scholar] [CrossRef]
- The State of Queensland, Department of Primary Industries and Fisheries. Annual Status Report: East Coast beche-de-mer Fishery 2007. Brisbane. 2007. Available online: https://www.daf.qld.gov.au/__data/assets/pdf_file/0016/70270/AnnualStatusReport-EastCoastBeche-de-mer2007-10Jan08.pdf (accessed on 15 March 2024).
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Northern sea cucumber (Cucumaria frondosa): A potential candidate for functional food, nutraceutical, and pharmaceutical sector. Mar. Drugs 2020, 18, 274. [Google Scholar] [CrossRef]
- Vu, D.T.; Falch, E.; Elvevoll, E.O.; Jensen, I.-J. Enzymatic Hydrolysis of Orange-Footed Sea Cucumber (Cucumaria frondosa)—Effect of Different Enzymes on Protein Yield and Bioactivity. Foods 2023, 12, 3685. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.L.; Ljubin, P.; Skjoldal, H.R.; Ingvaldsen, R.B.; Anisimova, N.; Manushin, I. Distribution of benthic megafauna in the Barents Sea: Baseline for an ecosystem approach to management. ICES J. Mar. Sci. 2015, 72, 595–613. [Google Scholar] [CrossRef]
- Hernández, J.C.; Clemente, S.; Tuya, F.; Pérez-Ruzafa, A.; Sangil, C.; Moro-Abad, L.; Bacallado-Aránega, J.J. Echinoderms of the Canary Islands, Spain. In Echinoderm Research and Diversity in Latin America; Alvarado, J., Solis-Marin, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 471–510. [Google Scholar]
- Purcell, S.W.; Lovatelli, A.; González-Wangüemert, M.; Solís-Marín, F.A.; Samyn, Y.; Conand, C. Commercially Important Sea Cucumbers of the World; FAO: Rome, Italy, 2023. [Google Scholar]
- Nakai, K.; Tsuruta, D. What are reactive oxygen species, free radicals, and oxidative stress in skin diseases? Int. J. Mol. Sci. 2021, 22, 10799. [Google Scholar] [CrossRef]
- Aruoma, O.I. Free radicals, oxidative stress, and antioxidants in human health and disease. J. Am. Oil Chem. Soc. 1998, 75, 199–212. [Google Scholar] [CrossRef]
- Senadheera, T.R.; Hossain, A.; Dave, D.; Shahidi, F. Antioxidant and ACE-Inhibitory Activity of Protein Hydrolysates Produced from Atlantic Sea Cucumber (Cucumaria frondosa). Molecules 2023, 28, 5263. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, C.; Coscueta, E.R.; Fernandes, J.; Pereira, R.N.; Teixeira, J.A.; Rodrigues, A.S.; Pintado, M.E. Bioactivity and bioaccessibility of bioactive compounds in gastrointestinal digestion of tomato bagasse extracts. Foods 2022, 11, 1064. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC, 18th ed.; Association of Official Analytical Chemist: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Latimer, G. Association of Official Analytical Chemists International, Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2019. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, S. Amino Acid Determination: Methods and Techniques, 2nd ed.; Marcel Dekker: New York, NY, USA, 1978. [Google Scholar]
- Osnes, K.K.; Mohr, V. Peptide hydrolases of Antartic krill, Euphausia superba. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1985, 82, 599–606. [Google Scholar] [CrossRef]
- Murphy, E.W.; Criner, P.E.; Gray, B.C. Comparisons of methods for calculating retentions of nutrients in cooked foods. J. Agric. Food Chem. 1975, 23, 1153–1157. [Google Scholar] [CrossRef]
- Jensen, I.-J.; Abrahamsen, H.; Maehre, H.K.; Elvevoll, E.O. Changes in antioxidative capacity of saithe (Pollachius virens) and shrimp (Pandalus borealis) during in vitro digestion. J. Agric. Food Chem. 2009, 57, 10928–10932. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Nenadis, N.; Lazaridou, O.; Tsimidou, M.Z. Use of reference compounds in antioxidant activity assessment. J. Agric. Food Chem. 2007, 55, 5452–5460. [Google Scholar] [CrossRef] [PubMed]
- Nenadis, N.; Wang, L.-F.; Tsimidou, M.; Zhang, H.-Y. Estimation of scavenging activity of phenolic compounds using the ABTS•+ assay. J. Agric. Food Chem. 2004, 52, 4669–4674. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC− fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Elvevoll, E.O.; James, D.; Toppe, J.; Gamarro, E.G.; Jensen, I.-J. Food Safety Risks Posed by Heavy Metals and Persistent Organic Pollutants (POPs) related to Consumption of Sea Cucumbers. Foods 2022, 11, 3992. [Google Scholar] [CrossRef] [PubMed]
- Ringvold, H.; Kjerstad, M. Norwegian red sea cucumber, Parastichopus tremulus (Gunnerus, 1767) (Holothuroidea, Echinodermata): Chemical and nutritional analysis. In Blue Growth: Aquaculture, Fisheries, Market and Health Perspectives Orkana; Orkana Academy: Stamsund, Norway, 2018. [Google Scholar]
- Wen, J.; Hu, C.; Fan, S. Chemical composition and nutritional quality of sea cucumbers. J. Sci. Food Agric. 2010, 90, 2469–2474. [Google Scholar] [CrossRef] [PubMed]
- Abuzaytoun, R.; Budge, S.M.; Xia, W.; MacKinnon, S. Unusual ether lipids and branched chain fatty acids in sea cucumber (Cucumaria frondosa) viscera and their seasonal variation. Mar. Drugs 2022, 20, 435. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, J.; Shimizu, A.; Kimizuka, A.; Ninomiya, T.; Katsuya, N. Contribution of peptides and amino acids to the taste of foods. J. Agric. Food Chem. 1969, 17, 689–695. [Google Scholar] [CrossRef]
- Sarower, M.G.; Hasanuzzaman, A.F.M.; Biswas, B.; Abe, H. Taste producing components in fish and fisheries products: A review. Int. J. Food Ferment. Technol. 2012, 2, 113–121. [Google Scholar]
- Fuke, S.; Konosu, S. Taste-active components in some foods: A review of Japanese research. Physiol. Behav. 1991, 49, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Kendler, S.; Thornes, F.W.; Jakobsen, A.N.; Lerfall, J. Nutritional profiling and contaminant levels of five underutilized fish species in Norway. Front. Nutr. 2023, 10, 1118094. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, X.; Liu, Y. Characterization and evaluation of umami taste: A review. Trends Anal. Chem. 2020, 127, 115876. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Styrbæk, K. Design and ‘umamification’ of vegetable dishes for sustainable eating. Int. J. Food Des. 2020, 5, 9–42. [Google Scholar] [CrossRef]
- Takagi, S.; Sato, Y.; Murata, Y.; Kokubun, A.; Touhata, K.; Ishida, N.; Agatsuma, Y. Quantification of the Flavor and Taste of Gonads from the Sea Urchin Mesocentrotus nudus Using GC–MS and a Taste-Sensing System. J. Sens. 2020, 20, 7008. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.-Y.; Jung, M.-J.; Nam, J.-W.; Han, A.-R.; Kim, B.-M.; Jun, J.-Y. Preparation and Taste Profiling of the Enzymatic Protein Hydrolysate from a by-Product of Red Snow Crab Processing as a Natural Seasoning Compound. Foods 2022, 11, 3911. [Google Scholar] [CrossRef]
- Schmidt, C.V.; Olsen, K.; Mouritsen, O.G. Umami synergy as the scientific principle behind taste-pairing champagne and oysters. Sci. Rep. 2020, 10, 20077. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.V.; Mouritsen, O.G. The solution to sustainable eating is not a one-way street. Front. Psychol. 2020, 11, 531. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Ninomiya, K. Umami and food palatability. J. Nutr. 2000, 130, 921S–926S. [Google Scholar] [CrossRef]
- Sroyraya, M.; Hanna, P.J.; Siangcham, T.; Tinikul, R.; Jattujan, P.; Poomtong, T.; Sobhon, P. Nutritional components of the sea cucumber Holothuria scabra. Funct. Foods Health Dis. 2017, 7, 168–181. [Google Scholar] [CrossRef]
- Yin, M.; Matsuoka, R.; Yanagisawa, T.; Xi, Y.; Zhang, L.; Wang, X. Effect of different drying methods on free amino acid and flavor nucleotides of scallop (Patinopecten yessoensis) adductor muscle. Food Chem. 2022, 396, 133620. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Toydemir, G.; Boyacioglu, D.; Beekwilder, J.; Hall, R.D.; Capanoglu, E. A review on the effect of drying on antioxidant potential of fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2016, 56, S110–S129. [Google Scholar] [CrossRef] [PubMed]
- Chong, N.V.W.; Pindi, W.; Chye, F.-Y.; Shaarani, S.M.; Lee, J.-S. Effects of drying methods on the quality of dried sea cucumbers from Sabah—A review. Int. J. Nov. Res. Life Sci. 2015, 2, 49–64. [Google Scholar]
- Waghmare, R.B.; Perumal, A.B.; Moses, J.A.; Anandharamakrishnan, C. 3.05—Recent Developments in Freeze Drying of Foods. In Innovative Food Processing Technologies; Knoerzer, K., Muthukumarappan, K., Eds.; Elsevier: Oxford, UK, 2021; pp. 82–99. [Google Scholar] [CrossRef]
- Duan, X.; Zhang, M.; Mujumdar, A.S.; Wang, S. Microwave freeze drying of sea cucumber (Stichopus japonicus). J. Food Eng. 2010, 96, 491–497. [Google Scholar] [CrossRef]
- Hoehl, K.; Schoenberger, G.; Busch-Stockfisch, M. Stimulus and recognition thresholds for the basic tastes in deionized water. Are the recommendations for citric acid too high. Ernähr. Umsch. 2014, 61, 130–136. [Google Scholar]
- Guennoun, Y.; Benajiba, N.; Elkari, K.; Bouziani, A.; Elammari, L.; Al-Jawaldeh, A.; Elhaloui, N.; Barkat, A.; Benkirane, H.; Aguenaou, H. The threshold of sweet taste recognition among a sample of Moroccan population. Food Sci. Nutr. 2022, 52, 45–60. [Google Scholar] [CrossRef]
- WHO. Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation. WHO Technical Report Series; No. 935. 2007. Available online: https://apps.who.int/iris/handle/10665/43411 (accessed on 20 September 2022).
- Ardiansyah, A.; Rasyid, A.; Siahaan, E.A.; Pangetistu, R.; Murniasih, T. Nutritional value and heavy metals content of sea cucumber Holothuria scabra commercially harvested in Indonesia. Curr. Res. Nutr. Food Sci. 2020, 8, 765–773. [Google Scholar] [CrossRef]
- Li, M.; Gao, Y.; Qi, Y.-X.; Song, Z.-Y.; Li, Z.-B.; Lin, Y.-T.; Zhao, Q.-C. Assessment of the nutritional value of cultured sea cucumber Apostichopus japonicus. J. Aquat. Food Prod. Technol. 2021, 30, 868–879. [Google Scholar] [CrossRef]
- Okuno, T.; Morimoto, S.; Nishikawa, H.; Haraguchi, T.; Kojima, H.; Tsujino, H.; Arisawa, M.; Yamashita, T.; Nishikawa, J.; Yoshida, M. Bitterness-Suppressing Effect of Umami Dipeptides and Their Constituent Amino Acids on Diphenhydramine: Evaluation by Gustatory Sensation and Taste Sensor Testing. Chem. Pharm. Bull. 2020, 68, 234–243. [Google Scholar] [CrossRef]
- Böhm, V.; Kühnert, S.; Rohm, H.; Scholze, G. Improving the nutritional quality of microwave-vacuum dried strawberries: A preliminary study. Food Sci. Technol. Int. 2006, 12, 67–75. [Google Scholar] [CrossRef]
- Valentina, V.; Pratiwi, R.A.; Hsiao, P.; Tseng, H.; Hsieh, J.; Chen, C. Sensorial characterization of foods before and after freeze-drying. Austin Food Sci. 2016, 1, 1027. [Google Scholar]
- Nursid, M.; Hadiati, D.; Winanto, T. Antioxidant capacity of dry sea cucumber Holothuria edulis, Pearsonothuria graeffei, and Stichopus herrmanni from Boalemo waters, Gorontalo, Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2022, 967, 012027. [Google Scholar] [CrossRef]
- Korczek, K.R.; Tkaczewska, J.; Duda, I.; Migdał, W. Effect of Heat Treatment on the Antioxidant Activity as Well as In vitro Digestion Stability of Herring (Clupea harengus) Protein Hydrolysates. J. Aquat. Food Prod. 2021, 30, 806–825. [Google Scholar] [CrossRef]
- Hjellnes, V.; Rustad, T.; Jensen, I.-J.; Eiken, E.; Pettersen, S.M.; Falch, E. Ultrafiltration of Saithe (Pollachius virens) Protein Hydrolysates and Its Effect on Antioxidative Activity. Catalysts 2021, 11, 1053. [Google Scholar] [CrossRef]
- Mildenberger, J.; Remm, M.; Atanassova, M. Self-assembly potential of bioactive peptides from Norwegian sea cucumber Parastichopus tremulus for development of functional hydrogels. LWT 2021, 148, 111678. [Google Scholar] [CrossRef]
- Jahan-Mihan, A.; Luhovyy, B.L.; Khoury, D.E.; Anderson, G.H. Dietary proteins as determinants of metabolic and physiologic functions of the gastrointestinal tract. Nutrients 2011, 3, 574–603. [Google Scholar] [CrossRef] [PubMed]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Hayes, K.D.; Shahidi, F. Comparative study on antioxidative activity of yellow stripe trevally protein hydrolysate produced from Alcalase and Flavourzyme. Int. J. Food Sci. Technol. 2008, 43, 1019–1026. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Arranz, S.; Tabernero, M.; Díaz-Rubio, M.E.; Serrano, J.; Goñi, I.; Saura-Calixto, F. Updated methodology to determine antioxidant capacity in plant foods, oils and beverages: Extraction, measurement and expression of results. Food Res. Int. 2008, 41, 274–285. [Google Scholar] [CrossRef]
- Suwaluk, R.; Chansuwan, W.; Sirinupong, N.; Chinachoti, P. Biological properties of peptide released by in-vitro stimulated digestion of cooked eats. J. Food Nutr. Res. 2021, 9, 87–95. [Google Scholar]
- Ucak, I.; Afreen, M.; Montesano, D.; Carrillo, C.; Tomasevic, I.; Simal-Gandara, J.; Barba, F.J. Functional and bioactive properties of peptides derived from marine side streams. Mar. Drugs 2021, 19, 71. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G. Chapter 12—Digestion—Absorption. In Medical Biochemistry; Academic Press: London, UK, 2017; pp. 251–273. [Google Scholar]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Zhang, Y.; He, S.; Bonneil, É.; Simpson, B.K. Generation of antioxidative peptides from Atlantic sea cucumber using alcalase versus trypsin: In vitro activity, de novo sequencing, and in silico docking for in vivo function prediction. Food Chem. 2020, 306, 125581. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-H.; Liu, B.-Q.; Cui, B.; Wen, L.; Xu, Z.; Chen, M.-L.; Wu, H. Alanine Substitution to Determine the Effect of LR5 and YR6 Rice Peptide Structure on Antioxidant and Anti-Inflammatory Activity. Nutrients 2023, 15, 2373. [Google Scholar] [CrossRef] [PubMed]
- Sichanova, M.; Geneva, M.; Petrova, M.; Miladinova-Georgieva, K.; Kirova, E.; Nedev, T.; Tsekova, D.; Ivanova, V.; Trendafilova, A. Influence of the Abiotic Elicitors Ag Salts of Aspartic Acid Derivatives, Self-Organized in Nanofibers with Monomeric and Dimeric Molecular Structures, on the Antioxidant Activity and Stevioside Content in Micropropagated Stevia rebaudiana Bert. Plants 2023, 12, 3574. [Google Scholar] [CrossRef] [PubMed]
- Fardus, J.; Hossain, S.; Rob, M.M.; Fujita, M. l-glutamic acid modulates antioxidant defense systems and nutrient homeostasis in lentil (Lens culinaris Medik.) under copper toxicity. Environ. Sci. Pollut. Res. 2023, 30, 78507–78520. [Google Scholar] [CrossRef]
- Sun, Y.; Miao, F.; Wang, Y.; Liu, H.; Wang, X.; Wang, H.; Guo, J.; Shao, R.; Yang, Q. L-Arginine alleviates the reduction in photosynthesis and antioxidant activity induced by drought stress in maize seedlings. J. Antioxid. 2023, 12, 482. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Frozen [%] | Freeze-Dried [%] |
|---|---|---|
| Water | 89.7 ± 0.8 | 3.4 ± 1.0 |
| Protein | 4.4 ± 0.7 | 43.1 ± 2.4 |
| Ash | 4.0 ± 0.4 | 36.3 ± 2.8 |
| Lipid | 1.3 ± 0.4 | 8.5 ± 2.2 |
| Total Amino Acids | Free Amino Acids | |||
|---|---|---|---|---|
| Amino Acids | g/100 g Dry Weight Frozen | g/100 g Dry Weight Freeze-Dried | mg/100 g Dry Weight Frozen | mg/100 g Dry Weight Freeze-Dried |
| Alanine | 2.76 ± 1.36 | 1.94 ± 0.35 | 13.0 ± 4.2 | 42.9 ± 10.6 * |
| Asparagine | <0.01 | <0.01 | 1.0 ± 0.6 | 3.5 ± 3.0 * |
| Aspartic acid | 5.08 ± 2.21 | 3.60 ± 0.73 | 10.8 ± 1.6 | 11.2 ± 3.4 |
| Glutamic acid | 6.79 ± 3.14 | 4.50 ± 1.08 | 139.2 ± 31.4 | 414.2 ± 48.1 * |
| Glutamine | 0.04 ± 0.02 * | <0.01 | 9.5 ± 4.4 | 32.2 ± 7.1 * |
| Glycine/Arginine | 6.07 ± 3.02 | 8.45 ± 2.04 | 45.8 ± 12.4 | 257.6 ± 57.7 * |
| Histidine | 0.48 ± 0.21 | 0.37 ± 0.07 | 1.8 ± 1.0 | 9.1 ± 2.5 * |
| Isoleucine | 1.35 ± 0.58 | 1.00 ± 0.25 | <1 | 1.3 ± 0.3 * |
| Leucine | 2.65 ± 1.16 | 1.83 ± 0.41 | 2.0 ± 0.9 | 3.4 ± 0.3 * |
| Lysine | 1.81 ± 0.80 | 1.31 ± 0.30 | 11.8 ± 3.3 | 36.5 ± 12.2 * |
| Methionine | 0.72 ± 0.34 * | 0.45 ± 0.16 | <1 | 1.2 ± 0.2 * |
| Phenylalanine | 1.55 ± 0.65 | 1.17 ± 0.24 | 1.0 ± 0.5 | 2.3 ± 0.2 * |
| Serine | 2.57 ± 1.21 * | 1.58 ± 0.46 | 1.8 ± 0.5 | 8.8 ± 2.5 * |
| Threonine | 2.63 ± 1.16 | 1.86 ± 0.42 | 1.4 ± 1.2 | 9.1 ± 2.5 * |
| Tyrosine | 1.33 ± 0.56 | 0.94 ± 0.19 | 1.1 ± 0.6 | 3.0 ± 0.3 * |
| Valine | 1.82 ± 0.78 | 1.38 ± 0.31 | 1.6 ± 0.9 | 2.6 ± 0.6 * |
| ∑TAAs ∑FAAs | 37.64 ± 17.15 | 30.39 ± 6.67 | 242.7 ± 53.1 | 839.2 ± 110.3 * |
| Samples | FRAP (mmol TE/100 g Dry Weight) | ABTS (mmol PGE/100 g Dry Weight) | ORAC (mmol TE/100 g Dry Weight) |
|---|---|---|---|
| Frozen | 3.2 ± 1.7 * | 0.1 ± 0.0 | 229.6 ± 107.9 * |
| Freeze-dried | 1.2 ± 0.2 | 0.3 ± 0.1 * | 130.7 ± 37.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, D.T.; Kletthagen, M.C.; Elvevoll, E.O.; Falch, E.; Jensen, I.-J. Simulated Digestion of Red Sea Cucumber (Parastichopus tremulus): A Study of Protein Quality and Antioxidant Activity. Appl. Sci. 2024, 14, 3267. https://doi.org/10.3390/app14083267
Vu DT, Kletthagen MC, Elvevoll EO, Falch E, Jensen I-J. Simulated Digestion of Red Sea Cucumber (Parastichopus tremulus): A Study of Protein Quality and Antioxidant Activity. Applied Sciences. 2024; 14(8):3267. https://doi.org/10.3390/app14083267
Chicago/Turabian StyleVu, Dat Trong, Malin Christine Kletthagen, Edel O. Elvevoll, Eva Falch, and Ida-Johanne Jensen. 2024. "Simulated Digestion of Red Sea Cucumber (Parastichopus tremulus): A Study of Protein Quality and Antioxidant Activity" Applied Sciences 14, no. 8: 3267. https://doi.org/10.3390/app14083267
APA StyleVu, D. T., Kletthagen, M. C., Elvevoll, E. O., Falch, E., & Jensen, I.-J. (2024). Simulated Digestion of Red Sea Cucumber (Parastichopus tremulus): A Study of Protein Quality and Antioxidant Activity. Applied Sciences, 14(8), 3267. https://doi.org/10.3390/app14083267