
Using Bi-Temporal Lidar to Evaluate Canopy Structure and Ecotone Influence on Landsat Vegetation Index Trends Within a Boreal Wetland Complex





Abstract
1. Introduction
- 1.
- To evaluate the hypothesis that Landsat-derived VIs are correlated with spatio-temporally coincident lidar-derived Canopy Height Models (CHMs) for at least some landcovers—specifically woody wetlands (e.g., shrub swamp, treed swamps) and upland forest areas—where VI values are expected to better reflect vertical vegetation structure, given that VIs are proxies for green leafy foliage cover and biomass tends to increase with mean canopy height [61]. This is evaluated through two sub-hypotheses:
- (a)
- Based on the assumption that EVI is more sensitive to variations in canopy structure and density, and typically yields higher values in areas of taller canopy [62], the first sub-hypothesis tests for a stronger positive correlation between CHM and EVI than for NDVI;
- (b)
- Given that foliage cover and vegetation height are not the same quantity, the second sub-hypothesis tests for significant differences in the relationships between CHM and VI (and their changes/trends) across distinct landcover classes [63].
- 2.
- To examine how trends in NDVI and EVI correspond with changes in canopy height over time and characterize spatial correspondences between regions of contiguous ecosystem or landcover expansion (or shrinkage) and regions of VI trend increase (or decrease). It is not expected that NDVI or EVI increasing (greening) or decreasing (browning) trends will consistently or strongly correlate with increases or decreases in lidar-derived CHM magnitudes at the grid cell level. Specifically, it is expected that any spatial correspondence between aggregate regions of VI trend- and CHM-based change will be most apparent where ecosystem height growth and foliage infilling are most pronounced, i.e., those associated with woody (shrub or tree) ecotonal expansion.
- 3.
- To explore differences in the EVI and NDVI trend response to changing wetland conditions that are associated with long term changes in soil surface saturation inferred from trends in LST, particularly in non-woody wetland types.
2. Materials and Methods
2.1. Study Area
2.2. Data Sources
2.2.1. Land Cover Data
2.2.2. Landsat Imagery
2.2.3. Airborne Lidar and Photography
2.3. Analysis
2.3.1. Landsat Time Series
2.3.2. Lidar-Derived Canopy Structure
2.3.3. Statistics
3. Results
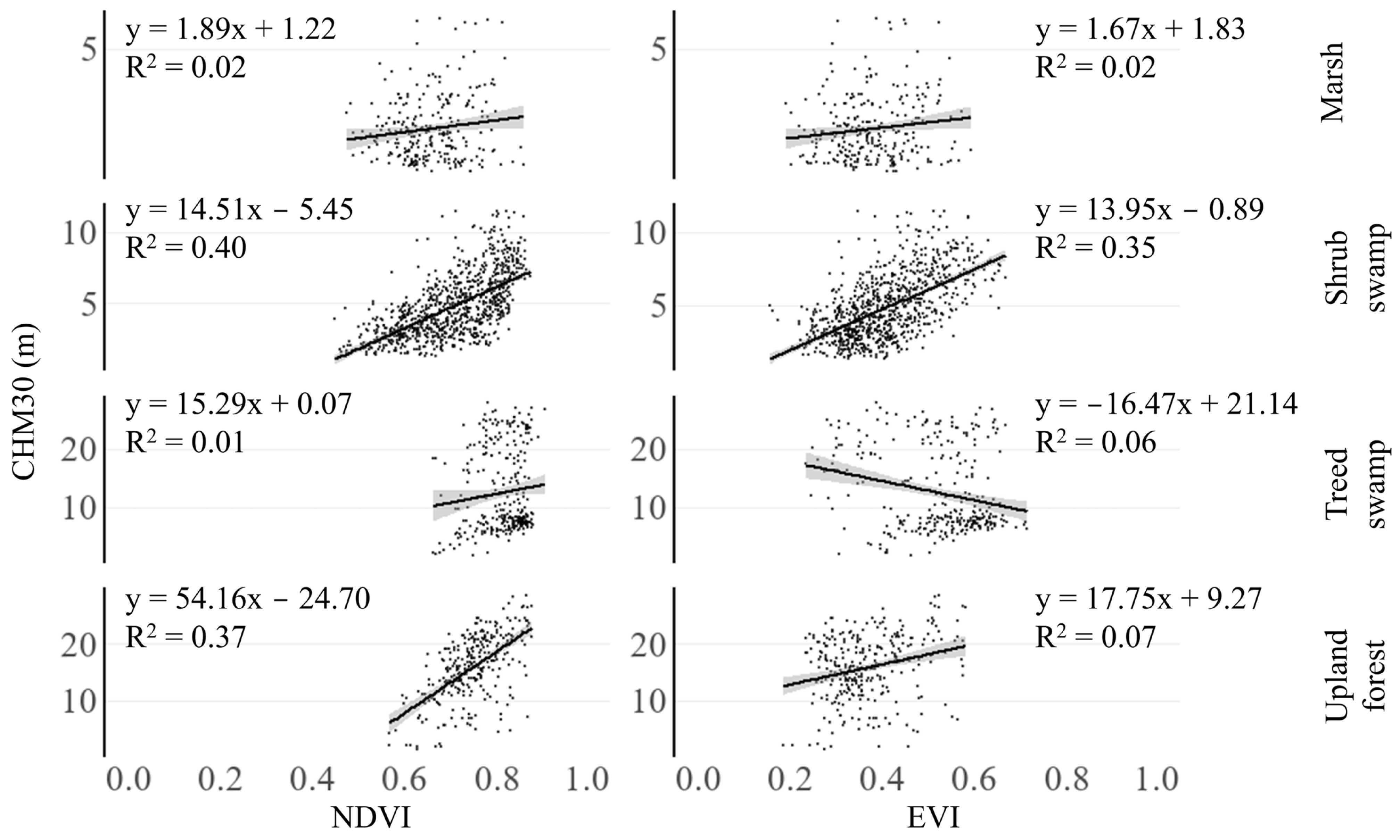
3.1. Correspondence Between Canopy Height and VIs
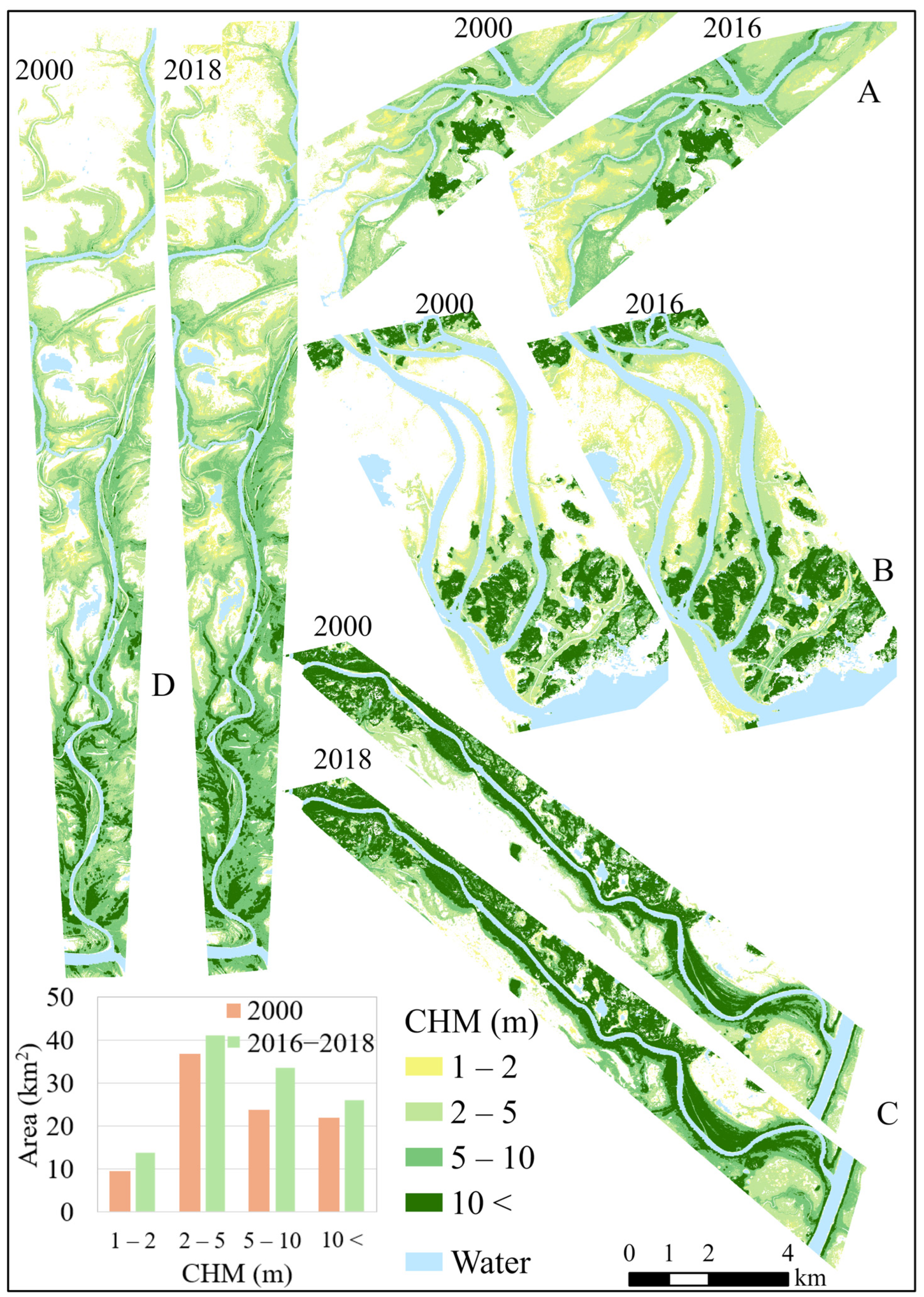
3.2. Changes in Canopy Height and Cover Using Bi-Temporal Lidar Data
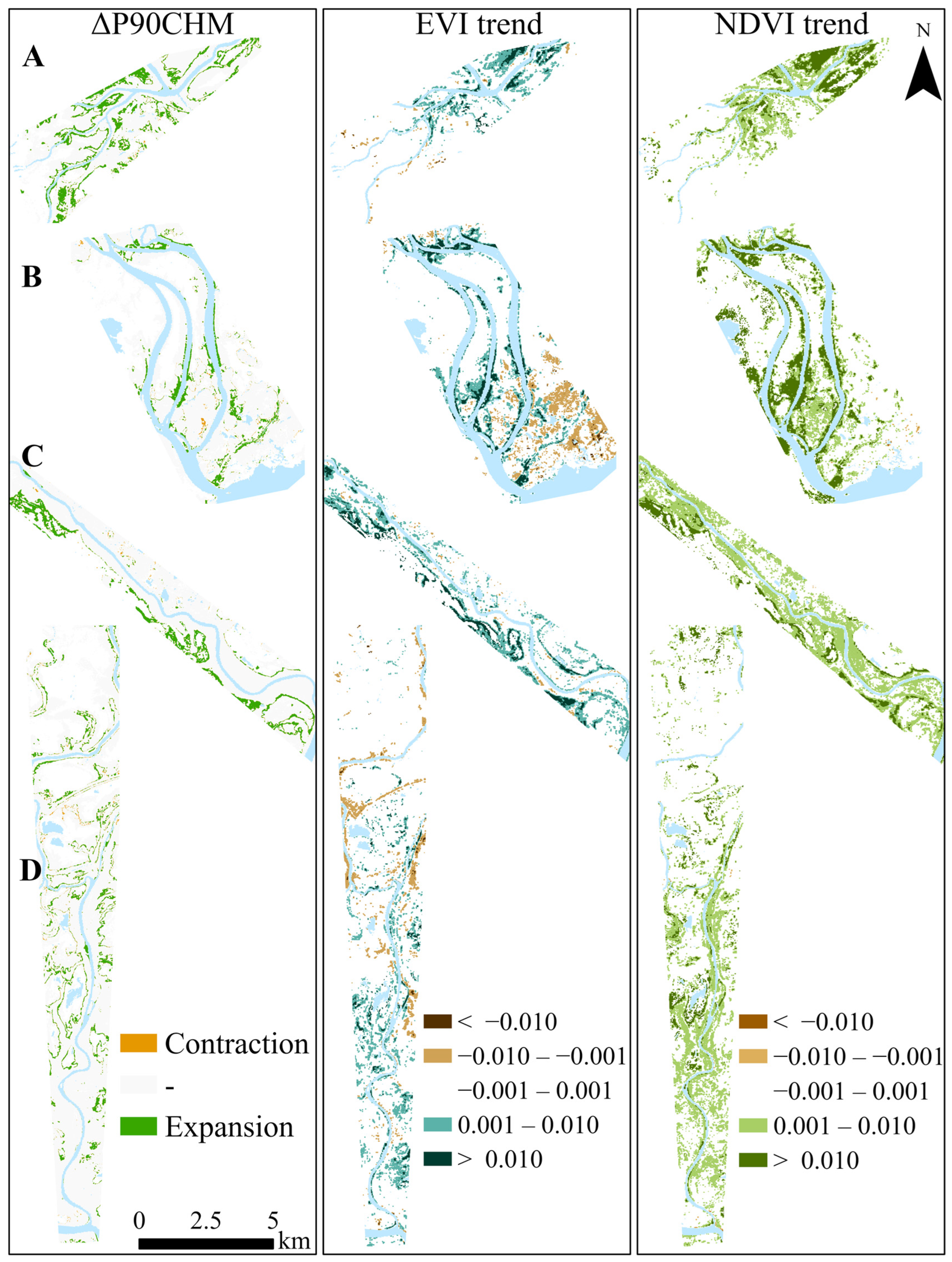
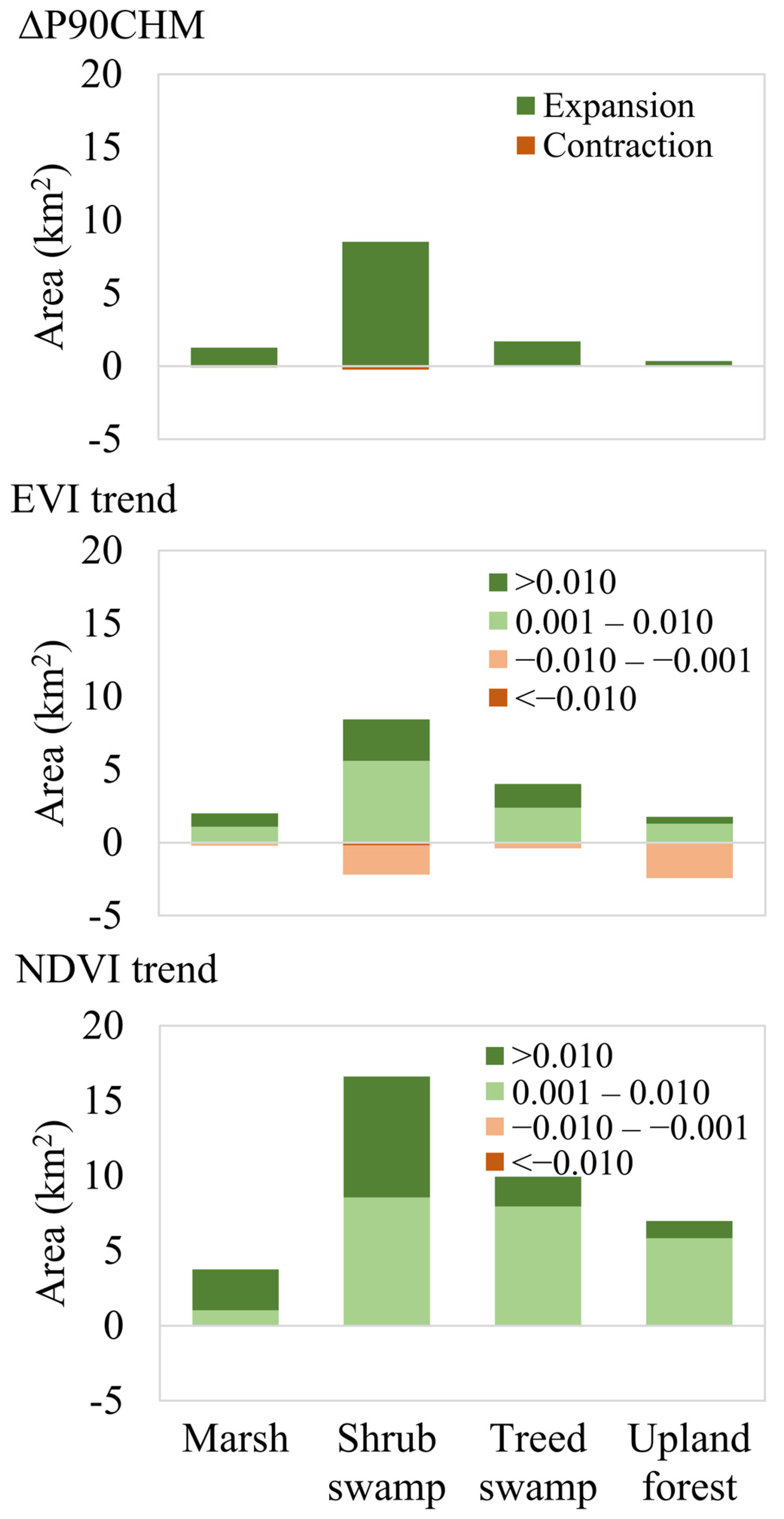
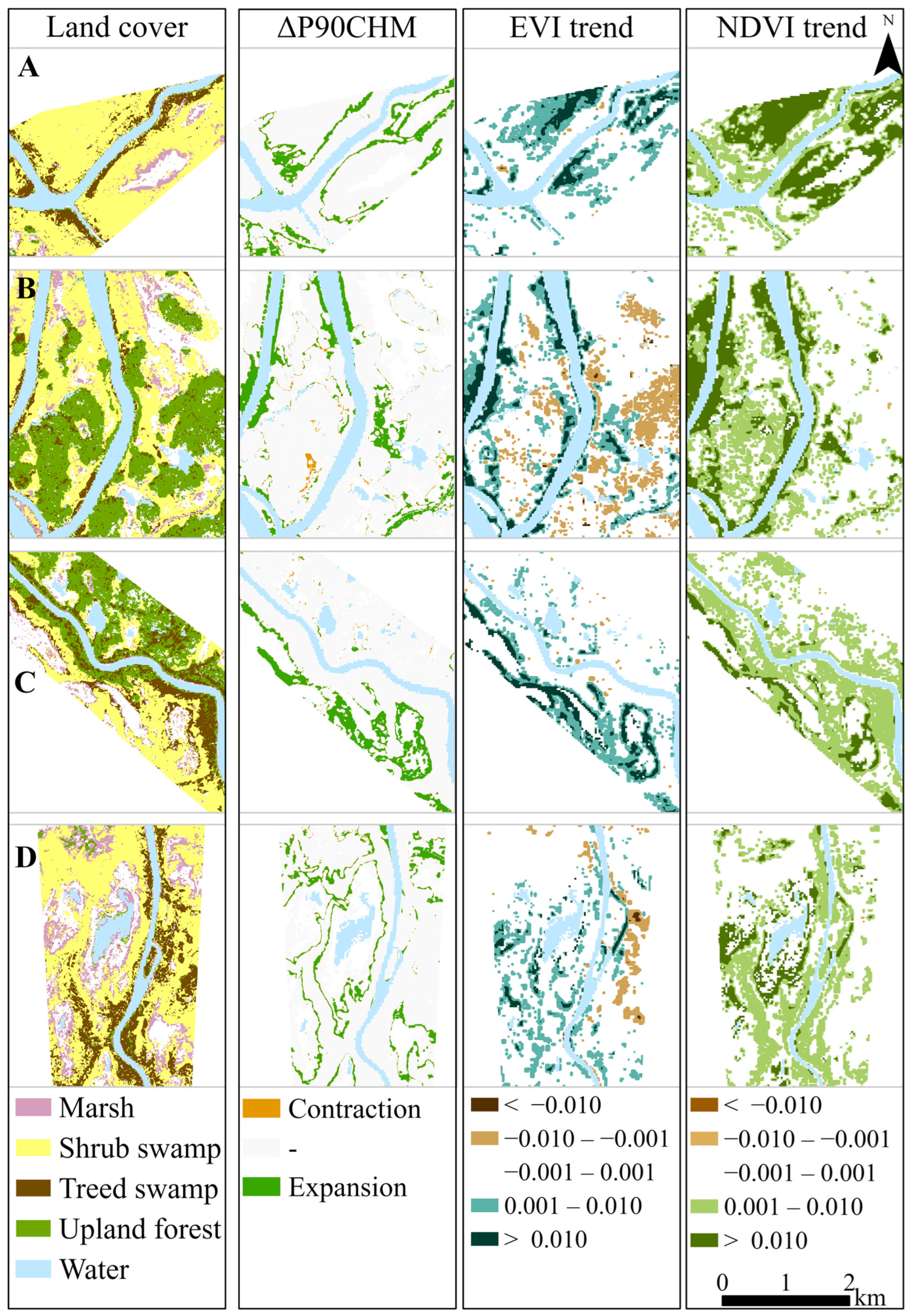
3.3. Changes in Ecotone Extent Inferred from CHM Change and VI Trend
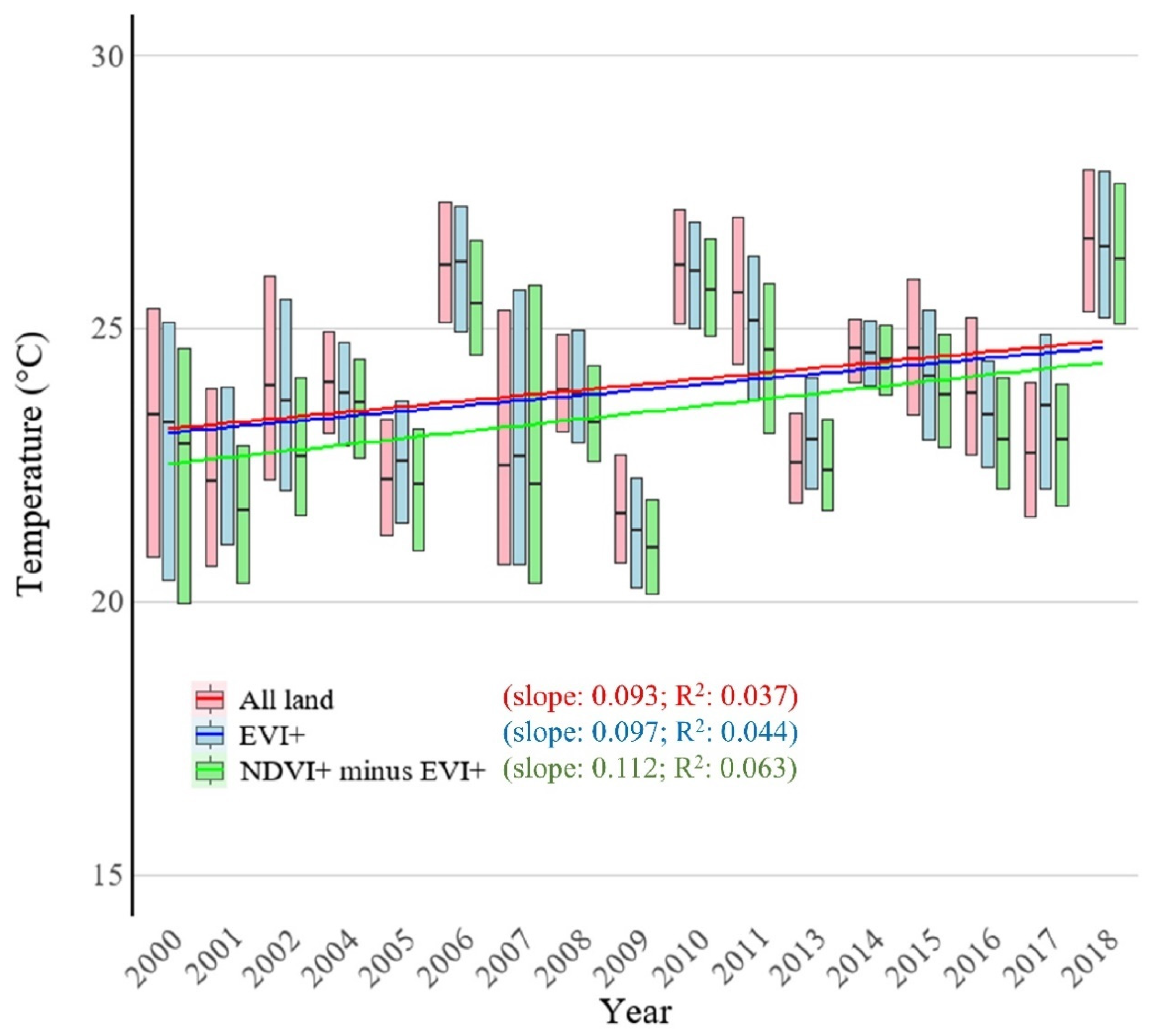
3.4. Changes in LST with VIs
4. Discussion
4.1. Do EVI and NDVI Correlate with Canopy Height?
4.2. Do Temporal Trends in EVI and NDVI Correlate with Canopy Growth?
4.3. How Do Trends in LST Differ Between EVI and NDVI Trends?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Timoney, K.; La Roi, G.; Zoltai, S.; Robinson, A. The high subarctic forest-tundra of northwestern Canada: Position, width, and vegetation gradients in relation to climate. Arctic 1992, 45, 1–9. [Google Scholar] [CrossRef]
- Ecosystem Classification Group. Ecological Regions of the Northwest Territories–Taiga Plains; Department of Environment and Natural Resources, Government of Northwest Territories: Yellowknife, NT, Canada, 2007.
- Fu, L.; Xie, R.; Ma, D.; Zhang, M.; Liu, L. Variations in soil microbial community structure and extracellular enzymatic activities along a forest–wetland ecotone in high-latitude permafrost regions. Ecol. Evol. 2023, 13, e10205. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, D.D.; Bhatti, J.S.; Grant, R.F. The transition zones (ecotone) between boreal forests and peatlands: Ecological controls on ecosystem productivity along a transition zone between upland black spruce forest and a poor forested fen in central Saskatchewan. Ecol. Model. 2014, 291, 96–108. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Rothwell, R.L. Rooting of peatland black spruce and tamarack in relation to depth of water table. Can. J. Bot. 1987, 65, 817–821. [Google Scholar] [CrossRef]
- Grant, R.F. Modeling topographic effects on net ecosystem productivity of boreal black spruce forests. Tree Physiol. 2004, 24, 1–18. [Google Scholar] [CrossRef]
- Gower, S.T.; Vogel, J.G.; Norman, J.M.; Kucharik, C.J.; Steele, S.J.; Stow, T.K. Carbon distribution and aboveground net primary production in aspen, jack pine, and black spruce stands in Saskatchewan and Manitoba, Canada. J. Geophys. Res. Atmos. 1997, 102, 29029–29041. [Google Scholar] [CrossRef]
- Timoney, K.P. The Peace-Athabasca Delta: Portrait of a Dynamic Ecosystem; University of Alberta: Edmonton, AB, Canada, 2013. [Google Scholar]
- Enayetullah, H.; Chasmer, L.; Hopkinson, C.; Thompson, D.; Cobbaert, D. Identifying Conifer Tree vs. Deciduous Shrub and Tree Regeneration Trajectories in a Space-for-Time Boreal Peatland Fire Chronosequence Using Multispectral Lidar. Atmosphere 2022, 13, 112. [Google Scholar] [CrossRef]
- Jean, S.A.; Pinno, B.D.; Nielsen, S.E. Early regeneration dynamics of pure black spruce and aspen forests after wildfire in boreal Alberta, Canada. Forests 2020, 11, 333. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Carpino, O.A.; Berg, A.A.; Quinton, W.L.; Adams, J.R. Climate change and permafrost thaw-induced boreal forest loss in northwestern Canada. Environ. Res. Lett. 2018, 13, 084018. [Google Scholar] [CrossRef]
- Birch, J.D.; Lutz, J.A.; Hogg, E.H.; Simard, S.W.; Pelletier, R.; LaRoi, G.H.; Karst, J. Decline of an ecotone forest: 50 years of demography in the southern boreal forest. Ecosphere 2019, 10, e02698. [Google Scholar] [CrossRef]
- Tremblay, B.; Lévesque, E.; Boudreau, S. Recent expansion of erect shrubs in the Low Arctic: Evidence from Eastern Nunavik. Environ. Res. Lett. 2012, 7, 035501. [Google Scholar] [CrossRef]
- Frost, G.V.; Epstein, H.E. Tall shrub and tree expansion in Siberian tundra ecotones since the 1960s. Glob. Change Biol. 2014, 20, 1264–1277. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.M.; Topp, E.; Gergel, S.E.; Martin, K.; Pirotti, F.; Sitzia, T. Expansion of subalpine woody vegetation over 40 years on Vancouver Island, British Columbia, Canada. Can. J. For. Res. 2016, 46, 437–443. [Google Scholar] [CrossRef]
- Chasmer, L.; Cobbaert, D.; Mahoney, C.; Millard, K.; Peters, D.; Devito, K.; Brisco, B.; Hopkinson, C.; Merchant, M.; Montgomery, J.; et al. Remote Sensing of Boreal Wetlands 1: Data Use for Policy and Management. Remote Sens. 2020, 12, 1320. [Google Scholar] [CrossRef]
- Töyrä, J.; Pietroniro, A. Towards operational monitoring of a northern wetland using geomatics-based techniques. Remote Sens. Environ. 2005, 97, 174–191. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Huete, A.; Liu, H.; Batchily, K.; Van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Huete, A.; Justice, C.; Liu, H. Development of vegetation and soil indices for MODIS-EOS. Remote Sens. Environ. 1994, 49, 224–234. [Google Scholar] [CrossRef]
- Chen, A.; Lantz, T.C.; Hermosilla, T.; Wulder, M.A. Biophysical controls of increased tundra productivity in the western Canadian Arctic. Remote Sens. Environ. 2021, 258, 112358. [Google Scholar] [CrossRef]
- Seider, J.H.; Lantz, T.C.; Hermosilla, T.; Wulder, M.A.; Wang, J.A. Biophysical determinants of shifting tundra vegetation productivity in the Beaufort Delta region of Canada. Ecosystems 2022, 25, 1435–1454. [Google Scholar] [CrossRef]
- Guay, K.C.; Beck, P.S.A.; Berner, L.T.; Goetz, S.J.; Baccini, A.; Buermann, W. Vegetation productivity patterns at high northern latitudes: A multi-sensor satellite data assessment. Glob. Change Biol. 2014, 20, 3147–3158. [Google Scholar] [CrossRef] [PubMed]
- Lebre, D. Differences in Vegetation Composition and Structure at Greening and Non-Greening Sites in the Northwest Territories. Ph.D. Thesis, Queen’s University, Ontario, CA, Canada, 2019. [Google Scholar]
- Morawitz, D.F.; Blewett, T.M.; Cohen, A.; Alberti, M. Using NDVI to Assess Vegetative Land Cover Change in Central Puget Sound. Environ. Monit. Assess. 2006, 114, 85–106. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.D.l.I.; Labib, S.M. Demystifying normalized difference vegetation index (NDVI) for greenness exposure assessments and policy interventions in urban greening. Environ. Res. 2023, 220, 115155. [Google Scholar] [CrossRef]
- Leahy, M.G.; Jollineau, M.Y.; Howarth, P.J.; Gillespie, A.R. The use of Landsat data for investigating the long-term trends in wetland change at Long Point, Ontario. Can. J. Remote Sens. 2005, 31, 240–254. [Google Scholar] [CrossRef]
- Olthof, I.; Pouliot, D.; Latifovic, R.; Chen, W. Recent (1986–2006) vegetation-specific NDVI trends in northern Canada from satellite data. Arctic 2008, 61, 381–394. [Google Scholar] [CrossRef]
- Matsushita, B.; Yang, W.; Chen, J.; Onda, Y.; Qiu, G. Sensitivity of the Enhanced Vegetation Index (EVI) and Normalized Difference Vegetation Index (NDVI) to Topographic Effects: A Case Study in High-density Cypress Forest. Sensors 2007, 7, 2636–2651. [Google Scholar] [CrossRef]
- Garroutte, E.L.; Hansen, A.J.; Lawrence, R.L. Using NDVI and EVI to Map Spatiotemporal Variation in the Biomass and Quality of Forage for Migratory Elk in the Greater Yellowstone Ecosystem. Remote Sens. 2016, 8, 404. [Google Scholar] [CrossRef]
- Birky, A.K. NDVI and a simple model of deciduous forest seasonal dynamics. Ecol. Model. 2001, 143, 43–58. [Google Scholar] [CrossRef]
- Sellers, P.J. Canopy reflectance, photosynthesis and transpiration. Int. J. Remote Sens. 1985, 6, 1335–1372. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Liu, H.Q.; Huete, A. A feedback based modification of the NDVI to minimize canopy background and atmospheric noise. IEEE Trans. Geosci. Remote Sens. 1995, 33, 457–465. [Google Scholar] [CrossRef]
- Sesnie, S.E.; Dickson, B.G.; Rosenstock, S.S.; Rundall, J.M. A comparison of Landsat TM and MODIS vegetation indices for estimating forage phenology in desert bighorn sheep (Ovis canadensis nelsoni) habitat in the Sonoran Desert, USA. Int. J. Remote Sens. 2012, 33, 276–286. [Google Scholar] [CrossRef]
- Berner, L.T.; Goetz, S.J. Satellite observations document trends consistent with a boreal forest biome shift. Glob. Change Biol. 2022, 28, 3275–3292. [Google Scholar] [CrossRef] [PubMed]
- Eastman, J.R.; Sangermano, F.; Machado, E.A.; Rogan, J.; Anyamba, A. Global trends in seasonality of normalized difference vegetation index (NDVI), 1982–2011. Remote Sens. 2013, 5, 4799–4818. [Google Scholar] [CrossRef]
- Han, Y.; Wang, Y.; Zhao, Y. Estimating soil moisture conditions of the greater Changbai Mountains by land surface temperature and NDVI. IEEE Trans. Geosci. Remote Sens. 2010, 48, 2509–2515. [Google Scholar]
- Naga Rajesh, A.; Abinaya, S.; Purna Durga, G.; Lakshmi Kumar, T.V. Long-term relationships of MODIS NDVI with rainfall, land surface temperature, surface soil moisture and groundwater storage over monsoon core region of India. Arid Land Res. Manag. 2023, 37, 51–70. [Google Scholar] [CrossRef]
- Thiebault, K.; Young, S. Snow cover change and its relationship with land surface temperature and vegetation in northeastern North America from 2000 to 2017. Int. J. Remote Sens. 2020, 41, 8453–8474. [Google Scholar] [CrossRef]
- Sharma, M.; Bangotra, P.; Gautam, A.S.; Gautam, S. Sensitivity of normalized difference vegetation index (NDVI) to land surface temperature, soil moisture and precipitation over district Gautam Buddh Nagar, UP, India. Stoch. Environ. Res. Risk Assess. 2022, 36, 1779–1789. [Google Scholar] [CrossRef]
- Raynolds, M.K.; Comiso, J.C.; Walker, D.A.; Verbyla, D. Relationship between satellite-derived land surface temperatures, arctic vegetation types, and NDVI. Remote Sens. Environ. 2008, 112, 1884–1894. [Google Scholar] [CrossRef]
- Kim, J.; Hogue, T.S. Improving spatial soil moisture representation through integration of AMSR-E and MODIS products. IEEE Trans. Geosci. Remote Sens. 2011, 50, 446–460. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, L.-W.; Shi, J.-J.; Huang, J.-F. Soil Moisture Monitoring Based on Land Surface Temperature-Vegetation Index Space Derived from MODIS Data. Pedosphere 2014, 24, 450–460. [Google Scholar] [CrossRef]
- Kornelsen, K.C.; Coulibaly, P. Advances in soil moisture retrieval from synthetic aperture radar and hydrological applications. J. Hydrol. 2013, 476, 460–489. [Google Scholar] [CrossRef]
- Joshi, N.; Baumann, M.; Ehammer, A.; Fensholt, R.; Grogan, K.; Hostert, P.; Jepsen, M.R.; Kuemmerle, T.; Meyfroidt, P.; Mitchard, E.T.A.; et al. A Review of the Application of Optical and Radar Remote Sensing Data Fusion to Land Use Mapping and Monitoring. Remote Sens. 2016, 8, 70. [Google Scholar] [CrossRef]
- Hopkinson, C.; Chasmer, L.E.; Sass, G.; Creed, I.F.; Sitar, M.; Kalbfleisch, W.; Treitz, P. Vegetation class dependent errors in lidar ground elevation and canopy height estimates in a boreal wetland environment. Can. J. Remote Sens. 2005, 31, 191–206. [Google Scholar] [CrossRef]
- Wulder, M.A.; White, J.C.; Bater, C.W.; Coops, N.C.; Hopkinson, C.; Chen, G. Lidar plots—A new large-area data collection option: Context, concepts, and case study. Can. J. Remote Sens. 2012, 38, 600–618. [Google Scholar] [CrossRef]
- Montgomery, J.; Brisco, B.; Chasmer, L.; Devito, K.; Cobbaert, D.; Hopkinson, C. SAR and LiDAR temporal data fusion approaches to boreal wetland ecosystem monitoring. Remote Sens. 2019, 11, 161. [Google Scholar] [CrossRef]
- Rood, S.B.; Kaluthota, S.; Philipsen, L.J.; Slaney, J.; Jones, E.; Chasmer, L.; Hopkinson, C. Camo-maps: An efficient method to assess and project riparian vegetation colonization after a major river flood. Ecol. Eng. 2019, 141, 105610. [Google Scholar] [CrossRef]
- Chasmer, L.; Mahoney, C.; Millard, K.; Nelson, K.; Peters, D.; Merchant, M.; Hopkinson, C.; Brisco, B.; Niemann, O.; Montgomery, J. Remote sensing of boreal wetlands 2: Methods for evaluating boreal wetland ecosystem state and drivers of change. Remote Sens. 2020, 12, 1321. [Google Scholar] [CrossRef]
- Bolton, D.K.; Coops, N.C.; Hermosilla, T.; Wulder, M.A.; White, J.C. Evidence of vegetation greening at alpine treeline ecotones: Three decades of Landsat spectral trends informed by lidar-derived vertical structure. Environ. Res. Lett. 2018, 13, 084022. [Google Scholar] [CrossRef]
- Hopkinson, C.; Sitar, M.; Chasmer, L.; Treitz, P. Mapping snowpack depth beneath forest canopies using airborne lidar. Photogramm. Eng. Remote Sens. 2004, 70, 323–330. [Google Scholar] [CrossRef]
- Jones, E.A.; Chasmer, L.E.; Devito, K.J.; Hopkinson, C.D. Shortening fire return interval predisposes west-central Canadian boreal peatlands to more rapid vegetation growth and transition to forest cover. Glob. Change Biol. 2024, 30, e17185. [Google Scholar] [CrossRef] [PubMed]
- Hopkinson, C.; Chasmer, L.; Barr, A.; Kljun, N.; Black, T.; McCaughey, J. Monitoring boreal forest biomass and carbon storage change by integrating airborne laser scanning, biometry and eddy covariance data. Remote Sens. Environ. 2016, 181, 82–95. [Google Scholar] [CrossRef]
- Ji, P.; Su, R.; Wu, G.; Xue, L.; Zhang, Z.; Fang, H.; Gao, R.; Zhang, W.; Zhang, D. Projecting Future Wetland Dynamics Under Climate Change and Land Use Pressure: A Machine Learning Approach Using Remote Sensing and Markov Chain Modeling. Remote Sens. 2025, 17, 1089. [Google Scholar] [CrossRef]
- Gxokwe, S.; Dube, T.; Mazvimavi, D. Multispectral Remote Sensing of Wetlands in Semi-Arid and Arid Areas: A Review on Applications, Challenges and Possible Future Research Directions. Remote Sens. 2020, 12, 4190. [Google Scholar] [CrossRef]
- Pan, F.; Xie, J.; Lin, J.; Zhao, T.; Ji, Y.; Hu, Q.; Pan, X.; Wang, C.; Xi, X. Evaluation of Climate Change Impacts on Wetland Vegetation in the Dunhuang Yangguan National Nature Reserve in Northwest China Using Landsat Derived NDVI. Remote Sens. 2018, 10, 735. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, W.; Xiao, A.; Zhang, R. The Susceptibility of Wetland Areas in the Yangtze River Basin to Temperature and Vegetation Changes. Remote Sens. 2023, 15, 4534. [Google Scholar] [CrossRef]
- Asner, G.P.; Clark, J.K.; Mascaro, J.; Vaudry, R.; Chadwick, K.D.; Vieilledent, G.; Rasamoelina, M.; Balaji, A.; Kennedy-Bowdoin, T.; Maatoug, L. Human and environmental controls over aboveground carbon storage in Madagascar. Carbon Balance Manag. 2012, 7, 2. [Google Scholar] [CrossRef]
- Goetz, S.; Dubayah, R. Advances in remote sensing technology and implications for measuring and monitoring forest carbon stocks and change. Carbon Manag. 2011, 2, 231–244. [Google Scholar] [CrossRef]
- Zhou, G.; Yin, X. Relationship of cotton nitrogen and yield with Normalized Difference Vegetation Index and plant height. Nutr. Cycl. Agroecosyst. 2014, 100, 147–160. [Google Scholar] [CrossRef]
- Ramsar Convention. An Introduction to the Ramsar Convention on Wetlands, 7th ed. Available online: https://www.ramsar.org/sites/default/files/documents/library/handbook1_5ed_introductiontoconvention_final_e.pdf (accessed on 1 February 2024).
- Peters, D.L.; Prowse, T.D.; Pietroniro, A.; Leconte, R. Flood hydrology of the Peace—Athabasca Delta, northern Canada. Hydrol. Process. Int. J. 2006, 20, 4073–4096. [Google Scholar] [CrossRef]
- Peters, D.L.; Prowse, T.D.; Marsh, P.; Lafleur, P.M.; Buttle, J.M. Persistence of water within perched basins of the Peace-Athabasca Delta, Northern Canada. Wetl. Ecol. Manag. 2006, 14, 221–243. [Google Scholar] [CrossRef]
- Newton, B.W.; Taube, N. Regional variability and changing water distributions drive large-scale water resource availability in Alberta, Canada. Can. Water Resour. J. Rev. Can. Ressour. Hydr. 2023, 48, 300–326. [Google Scholar] [CrossRef]
- Ducks Unlimited Canada. Wood Buffalo National Park Enhanced Wetland Classification User’s Guide; Ducks Unlimited Canada: Edmonton, AB, Canada, 2020; p. 50. [Google Scholar]
- Mishra, N.; Helder, D.; Barsi, J.; Markham, B. Continuous calibration improvement in solar reflective bands: Landsat 5 through Landsat 8. Remote Sens. Environ. 2016, 185, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Potapov, P.; Turubanova, S.; Hansen, M.C. Regional-scale boreal forest cover and change mapping using Landsat data composites for European Russia. Remote Sens. Environ. 2011, 115, 548–561. [Google Scholar] [CrossRef]
- Roy, D.P.; Kovalskyy, V.; Zhang, H.; Vermote, E.F.; Yan, L.; Kumar, S.; Egorov, A. Characterization of Landsat-7 to Landsat-8 reflective wavelength and normalized difference vegetation index continuity. Remote Sens. Environ. 2016, 185, 57–70. [Google Scholar] [CrossRef]
- Qiu, S.; Lin, Y.; Shang, R.; Zhang, J.; Ma, L.; Zhu, Z. Making Landsat time series consistent: Evaluating and improving Landsat analysis ready data. Remote Sens. 2018, 11, 51. [Google Scholar] [CrossRef]
- Chen, S.; Woodcock, C.E.; Bullock, E.L.; Arévalo, P.; Torchinava, P.; Peng, S.; Olofsson, P. Monitoring temperate forest degradation on Google Earth Engine using Landsat time series analysis. Remote Sens. Environ. 2021, 265, 112648. [Google Scholar] [CrossRef]
- Masek, J.G.; Wulder, M.A.; Markham, B.; McCorkel, J.; Crawford, C.J.; Storey, J.; Jenstrom, D.T. Landsat 9: Empowering open science and applications through continuity. Remote Sens. Environ. 2020, 248, 111968. [Google Scholar] [CrossRef]
- USGS. Landsat Collection 2. Available online: https://pubs.usgs.gov/fs/2021/3002/fs20213002.pdf (accessed on 1 April 2024).
- Töyrä, J.; Pietroniro, A.; Hopkinson, C.; Kalbfleisch, W. Assessment of airborne scanning laser altimetry (lidar) in a deltaic wetland environment. Can. J. Remote Sens. 2003, 29, 718–728. [Google Scholar] [CrossRef]
- Hopkinson, C.; Chasmer, L.; Gynan, C.; Mahoney, C.; Sitar, M. Multisensor and Multispectral LiDAR Characterization and Classification of a Forest Environment. Can. J. Remote Sens. 2016, 42, 501–520. [Google Scholar] [CrossRef]
- Hopkinson, C.; Chasmer, L.; Hall, R. The uncertainty in conifer plantation growth prediction from multi-temporal lidar datasets. Remote Sens. Environ. 2008, 112, 1168–1180. [Google Scholar] [CrossRef]
- Lim, K.; Hopkinson, C.; Treitz, P. Examining the effects of sampling point densities on laser canopy height and density metrics. For. Chron. 2008, 84, 876–885. [Google Scholar] [CrossRef]
- Crego, R.D.; Stabach, J.A.; Connette, G. Implementation of species distribution models in Google Earth Engine. Divers. Distrib. 2022, 28, 904–916. [Google Scholar] [CrossRef]
- Hemati, M.; Hasanlou, M.; Mahdianpari, M.; Mohammadimanesh, F. A systematic review of landsat data for change detection applications: 50 years of monitoring the earth. Remote Sens. 2021, 13, 2869. [Google Scholar] [CrossRef]
- Teluguntla, P.; Thenkabail, P.S.; Oliphant, A.; Xiong, J.; Gumma, M.K.; Congalton, R.G.; Yadav, K.; Huete, A. A 30-m landsat-derived cropland extent product of Australia and China using random forest machine learning algorithm on Google Earth Engine cloud computing platform. ISPRS J. Photogramm. Remote Sens. 2018, 144, 325–340. [Google Scholar] [CrossRef]
- Gumma, M.K.; Thenkabail, P.S.; Teluguntla, P.G.; Oliphant, A.; Xiong, J.; Giri, C.; Pyla, V.; Dixit, S.; Whitbread, A.M. Agricultural cropland extent and areas of South Asia derived using Landsat satellite 30-m time-series big-data using random forest machine learning algorithms on the Google Earth Engine cloud. GIScience Remote Sens. 2020, 57, 302–322. [Google Scholar] [CrossRef]
- Oliphant, A.J.; Thenkabail, P.S.; Teluguntla, P.; Xiong, J.; Gumma, M.K.; Congalton, R.G.; Yadav, K. Mapping cropland extent of Southeast and Northeast Asia using multi-year time-series Landsat 30-m data using a random forest classifier on the Google Earth Engine Cloud. Int. J. Appl. Earth Obs. Geoinf. 2019, 81, 110–124. [Google Scholar] [CrossRef]
- Aslami, F. Lidar Derived Models and NDVI Trends Indicate Vegetation Threshold Response to Hydroclimatic Drivers Across the Peace Athabasca Delta. Master’s Thesis, University of Lethbridge, Lethbridge, AB, Canada, 2024. [Google Scholar]
- Ju, J.; Masek, J.G. The vegetation greenness trend in Canada and US Alaska from 1984–2012 Landsat data. Remote Sens. Environ. 2016, 176, 1–16. [Google Scholar] [CrossRef]
- Xu, H. Modification of normalised difference water index (NDWI) to enhance open water features in remotely sensed imagery. Int. J. Remote Sens. 2006, 27, 3025–3033. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Theil, H. A rank-invariant method of linear and polynomial regression analysis. Indag. Math. 1950, 12, 173. [Google Scholar]
- Sen, P.K. Estimates of the regression coefficient based on Kendall’s tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Kang, Y.; Guo, E.; Wang, Y.; Bao, Y.; Bao, Y.; Mandula, N. Monitoring vegetation change and its potential drivers in Inner Mongolia from 2000 to 2019. Remote Sens. 2021, 13, 3357. [Google Scholar] [CrossRef]
- Munir, S. Analysing temporal trends in the ratios of PM2. 5/PM10 in the UK. Aerosol Air Qual. Res. 2017, 17, 34–48. [Google Scholar] [CrossRef]
- Mancino, G.; Console, R.; Greco, M.; Iacovino, C.; Trivigno, M.L.; Falciano, A. Assessing vegetation decline due to pollution from solid waste management by a multitemporal remote sensing approach. Remote Sens. 2022, 14, 428. [Google Scholar] [CrossRef]
- Eastman, J.R. TerrSet, Geospatial Monitoring and Modeling Software; Clark Labs, Clark University: Worcester, MA, USA, 2015. [Google Scholar]
- Xu, Z.; Takeuchi, K.; Ishidaira, H. Monotonic trend and step changes in Japanese precipitation. J. Hydrol. 2003, 279, 144–150. [Google Scholar] [CrossRef]
- Tran, T.V.; Tran, D.X.; Myint, S.W.; Huang, C.-Y.; Pham, H.V.; Luu, T.H.; Vo, T.M.T. Examining spatiotemporal salinity dynamics in the Mekong River Delta using Landsat time series imagery and a spatial regression approach. Sci. Total Environ. 2019, 687, 1087–1097. [Google Scholar] [CrossRef]
- Kumar, R.; Nath, A.J.; Nath, A.; Sahu, N.; Pandey, R. Landsat-based multi-decadal spatio-temporal assessment of the vegetation greening and browning trend in the Eastern Indian Himalayan Region. Remote Sens. Appl. Soc. Environ. 2022, 25, 100695. [Google Scholar] [CrossRef]
- Schucknecht, A.; Erasmi, S.; Niemeyer, I.; Matschullat, J. Assessing vegetation variability and trends in north-eastern Brazil using AVHRR and MODIS NDVI time series. Eur. J. Remote Sens. 2013, 46, 40–59. [Google Scholar] [CrossRef]
- Li, L.; Zhan, W.; Du, H.; Lai, J.; Wang, C.; Fu, H.; Huang, F.; Liu, Z.; Wang, C.; Li, J. Long-Term and Fine-Scale Surface Urban Heat Island Dynamics Revealed by Landsat Data Since the 1980s: A Comparison of Four Megacities in China. J. Geophys. Res. Atmos. 2022, 127, e2021JD035598. [Google Scholar] [CrossRef]
- Andronis, V.; Karathanassi, V.; Tsalapati, V.; Kolokoussis, P.; Miltiadou, M.; Danezis, C. Time Series Analysis of Landsat Data for Investigating the Relationship between Land Surface Temperature and Forest Changes in Paphos Forest, Cyprus. Remote Sens. 2022, 14, 1010. [Google Scholar] [CrossRef]
- Srivastava, P.K.; Han, D.; Ramirez, M.R.; Islam, T. Machine Learning Techniques for Downscaling SMOS Satellite Soil Moisture Using MODIS Land Surface Temperature for Hydrological Application. Water Resour. Manag. 2013, 27, 3127–3144. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, L.; Wang, Y.; Chen, L.; Yao, F.; Wu, P.; Wang, B.; Li, Y.; Zhou, T.; Zhang, T. Research on the Contribution of Urban Land Surface Moisture to the Alleviation Effect of Urban Land Surface Heat Based on Landsat 8 Data. Remote Sens. 2015, 7, 10737–10762. [Google Scholar] [CrossRef]
- Nelson, K.; Chasmer, L.; Hopkinson, C. Quantifying Lidar Elevation Accuracy: Parameterization and Wavelength Selection for Optimal Ground Classifications Based on Time since Fire/Disturbance. Remote Sens. 2022, 14, 5080. [Google Scholar] [CrossRef]
- Wang, C.; Pavelsky, T.; Kyzivat, E.; Garcia-Tigreros, F.; Yao, F.; Yang, X.; Zhang, S.; Song, C.; Langhorst, T.; Dolan, W. ABoVE: Wetland Vegetation Classification for Peace-Athabasca Delta, Canada, 2019; ORNL DAAC: Oak Ridge, TN, USA, 2022. [Google Scholar]
- Royston, P. Approximating the Shapiro-Wilk W-test for non-normality. Stat. Comput. 1992, 2, 117–119. [Google Scholar] [CrossRef]
- Taddeo, S.; Dronova, I.; Depsky, N. Spectral vegetation indices of wetland greenness: Responses to vegetation structure, composition, and spatial distribution. Remote Sens. Environ. 2019, 234, 111467. [Google Scholar] [CrossRef]
- Moura, Y.M.; Galvão, L.S.; dos Santos, J.R.; Roberts, D.A.; Breunig, F.M. Use of MISR/Terra data to study intra- and inter-annual EVI variations in the dry season of tropical forest. Remote Sens. Environ. 2012, 127, 260–270. [Google Scholar] [CrossRef]
- Asner, G.P. Biophysical and biochemical sources of variability in canopy reflectance. Remote Sens. Environ. 1998, 64, 234–253. [Google Scholar] [CrossRef]
- Chen, X.; Vierling, L.; Rowell, E.; DeFelice, T. Using lidar and effective LAI data to evaluate IKONOS and Landsat 7 ETM+ vegetation cover estimates in a ponderosa pine forest. Remote Sens. Environ. 2004, 91, 14–26. [Google Scholar] [CrossRef]
- Galvão, L.S.; Breunig, F.M.; Teles, T.S.; Gaida, W.; Balbinot, R. Investigation of terrain illumination effects on vegetation indices and VI-derived phenological metrics in subtropical deciduous forests. GIScience Remote Sens. 2016, 53, 360–381. [Google Scholar] [CrossRef]
- Morton, D.C.; Nagol, J.; Carabajal, C.C.; Rosette, J.; Palace, M.; Cook, B.D.; Vermote, E.F.; Harding, D.J.; North, P.R.J. Amazon forests maintain consistent canopy structure and greenness during the dry season. Nature 2014, 506, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Galvão, L.S.; dos Santos, J.R.; Roberts, D.A.; Breunig, F.M.; Toomey, M.; de Moura, Y.M. On intra-annual EVI variability in the dry season of tropical forest: A case study with MODIS and hyperspectral data. Remote Sens. Environ. 2011, 115, 2350–2359. [Google Scholar] [CrossRef]
- Mo, Y.; Kearney, M.S.; Riter, J.A.; Zhao, F.; Tilley, D.R. Assessing biomass of diverse coastal marsh ecosystems using statistical and machine learning models. Int. J. Appl. Earth Obs. Geoinf. 2018, 68, 189–201. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; Macias-Fauria, M.; Sass-Klaassen, U.; Lévesque, E. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef]
- Peters, D.; Buttle, J. The effects of flow regulation and climatic variability on obstructed drainage and reverse flow contribution in a Northern river–lake–Delta complex, Mackenzie basin headwaters. River Res. Appl. 2010, 26, 1065–1089. [Google Scholar] [CrossRef]
- Timoney, K.P. Has river regulation damaged the Peace-Athabasca Delta? Ecoscience 2024, 31, 118–148. [Google Scholar] [CrossRef]
- Nill, L.; Grünberg, I.; Ullmann, T.; Gessner, M.; Boike, J.; Hostert, P. Arctic shrub expansion revealed by Landsat-derived multitemporal vegetation cover fractions in the Western Canadian Arctic. Remote Sens. Environ. 2022, 281, 113228. [Google Scholar] [CrossRef]
- Gao, W.; Zheng, C.; Liu, X.; Lu, Y.; Chen, Y.; Wei, Y.; Ma, Y. NDVI-based vegetation dynamics and their responses to climate change and human activities from 1982 to 2020: A case study in the Mu Us Sandy Land, China. Ecol. Indic. 2022, 137, 108745. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Martín-Hernández, N.; Reig, F.; Azorin-Molina, C.; Zabalza, J.; Beguería, S.; Domínguez-Castro, F.; El Kenawy, A.; Peña-Gallardo, M.; Noguera, I.; et al. Vegetation greening in Spain detected from long term data (1981–2015). Int. J. Remote Sens. 2020, 41, 1709–1740. [Google Scholar] [CrossRef]
- Luo, H.; Zhang, L.; Zhang, X. Shifts in land-greening hotspots in the Yellow River Eco-Economic Belt during the last four decades and their connections to human activities. Remote Sens. Appl. Soc. Environ. 2022, 27, 100783. [Google Scholar] [CrossRef]
- Li, J.; Holmgren, M.; Xu, C. Greening vs browning? Surface water cover mediates how tundra and boreal ecosystems respond to climate warming. Environ. Res. Lett. 2021, 16, 104004. [Google Scholar] [CrossRef]
- Ghulam, A.; Qin, Q.; Zhan, Z. Designing of the perpendicular drought index. Environ. Geol. 2007, 52, 1045–1052. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; Price, D.T.; Kurz, W.A.; Wu, C. Land surface phenology from optical satellite measurement and CO2 eddy covariance technique. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Dourado, G.F.; Motta, J.S.; Filho, A.C.P.; Scott, D.F.; Gabas, S.G. The Use of Remote Sensing Indices for Land Cover Change Detection. Anuário Inst. Geociências 2019, 42, 72–85. [Google Scholar] [CrossRef]
- Lu, L.; Kuenzer, C.; Wang, C.; Guo, H.; Li, Q. Evaluation of three MODIS-derived vegetation index time series for dryland vegetation dynamics monitoring. Remote Sens. 2015, 7, 7597–7614. [Google Scholar] [CrossRef]
- Huete, A.; Tucker, C. Investigation of soil influences in AVHRR red and near-infrared vegetation index imagery. Int. J. Remote Sens. 1991, 12, 1223–1242. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landcover Class | N | Correlation with CHM30 | p-Value | ||
|---|---|---|---|---|---|
| NDVI | EVI | NDVI | EVI | ||
| Marsh | 261 | 0.05 | 0.11 | 0.425 | 0.090 |
| Shrub swamp | 881 | 0.65 ** | 0.59 ** | <0.001 | <0.001 |
| Treed swamp | 292 | 0.13 * | −0.14 * | 0.023 | 0.014 |
| Upland forest | 293 | 0.58 ** | 0.22 ** | <0.001 | <0.001 |
| All | 1727 | 0.63 ** | 0.31 ** | <0.001 | <0.001 |
| Survey Area | ΔP90CHM | NDVI Trend | EVI Trend | ΔP90CHM vs. NDVI Trend | ΔP90CHM vs. EVI Trend | ||
|---|---|---|---|---|---|---|---|
| Mean Rate of Expansion m/yr (SD) | Difference | p Value | Difference | p Value | |||
| A | 3.3 (3.1) | 5.5 (3.2) | 2.4 (3.3) | −2.2 | 0.00 | 0.9 | 0.24 |
| B | 2.6 (3.0) | 5.5 (5.0) | 1.9 (3.0) | −2.9 | 0.00 | 0.7 | 0.39 |
| C | 2.4 (0.3) | 7.2 (4.7) | 3.6 (3.8) | −4.8 | < 0.001 | −1.2 | 0.04 |
| D | 1.5 (1.2) | 4.1 (2.7) | 1.7 (3.3) | −2.6 | < 0.001 | −0.2 | 0.62 |
| All | 2.2 (2.3) | 5.4 (4.1) | 2.3 (3.8) | −3.2 | < 0.001 | −0.1 | 0.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslami, F.; Hopkinson, C.; Chasmer, L.; Mahoney, C.; Peters, D.L. Using Bi-Temporal Lidar to Evaluate Canopy Structure and Ecotone Influence on Landsat Vegetation Index Trends Within a Boreal Wetland Complex. Appl. Sci. 2025, 15, 4653. https://doi.org/10.3390/app15094653
Aslami F, Hopkinson C, Chasmer L, Mahoney C, Peters DL. Using Bi-Temporal Lidar to Evaluate Canopy Structure and Ecotone Influence on Landsat Vegetation Index Trends Within a Boreal Wetland Complex. Applied Sciences. 2025; 15(9):4653. https://doi.org/10.3390/app15094653
Chicago/Turabian StyleAslami, Farnoosh, Chris Hopkinson, Laura Chasmer, Craig Mahoney, and Daniel L. Peters. 2025. "Using Bi-Temporal Lidar to Evaluate Canopy Structure and Ecotone Influence on Landsat Vegetation Index Trends Within a Boreal Wetland Complex" Applied Sciences 15, no. 9: 4653. https://doi.org/10.3390/app15094653
APA StyleAslami, F., Hopkinson, C., Chasmer, L., Mahoney, C., & Peters, D. L. (2025). Using Bi-Temporal Lidar to Evaluate Canopy Structure and Ecotone Influence on Landsat Vegetation Index Trends Within a Boreal Wetland Complex. Applied Sciences, 15(9), 4653. https://doi.org/10.3390/app15094653