1. Introduction
Even if all feed production requirements are met, there are risks of xenobiotics of various etiologies entering the feed [
1]. Once in animals, they cause serious health problems, can biotransform and accumulate in animal products, and thereby cause harm, not only to animals, but also to humans [
2,
3].
Meanwhile, animals consuming feeds containing various groups of xenobiotics suffer from damage to the limbs, liver, and digestive organs, metabolic disorders, postnatal complications, and reproductive disorders [
4]. Often, contaminants of fungal (mycotoxins), bacterial (exo- and endotoxins), and chemical origin (pesticides and heavy metals) are present simultaneously in the feed, which has a synergistic negative effect on the animal organism [
5,
6,
7].
The accumulation of xenobiotics determines changes in the intracellular content of dissolved substances, which can lead to volume changes, called isoosmotic volume changes, since the plasma membrane is easily permeable to water, and under normal conditions the body is able to conduct a rapid, multidirectional water flow to establish equilibrium, which is often disturbed by various pollutants [
8,
9]. This disturbance in the body can reach a state of oxidative stress when the balance between oxidants and antioxidants is disturbed. Stresses, including oxidative stress, are a special case of metabolic disorders, in which improper feeding and housing contribute to higher levels of endo- and exotoxins entering the bloodstream, causing inflammation in the animals, with a significant decrease in productivity [
10].
For many years, it was believed that ruminants have absolute resistance to mycotoxins. However, the metabolism of mycotoxins in the rumen is not equivalent to their complete detoxification. Furthermore, their potential degradation depends to a large extent on the stability of the rumen microbial community and the pH, which varies depending on the feed composition [
11]. It is known that high-yielding cows, transition cows, and calves are the most demanding in terms of optimal feeding and housing conditions. As a result of selection solely for milk productivity, low resistance, increased sensitivity to stresses, and pathological reaction to even insignificant changes in cattle maintenance and feeding are often observed in high-productivity cows. The presence of toxigens in feed is a factor leading to subclinical pathological conditions with a negative dynamic which, in the absence of a precise diagnosis and qualified help, is highly likely to lead to a fatal outcome. A typical example of this trend is the large number of high-yielding cattle that are culled immediately after calving or in the first months of lactation [
12]. According to Hadley et al. [
13], up to 80% of culled dairy cows have health problems.
The presence of toxins in feed primarily affects the rumen microflora, as many secondary metabolites are produced during digestion, negatively affecting numerous groups of microorganisms living in the rumen [
14]. It is therefore a proven fact that ruminants are more affected by natural and natural pollutants than other species [
15].
Many studies have been conducted on the effects of chemical toxins [
16] and mycotoxins [
17] on the ruminant organism. Mechanisms have also been developed to mitigate their effects on the animal organism, primarily related to measures to reduce emissions into the environment, to improve protocols for forage harvesting and storage, and to neutralize and remove them from the animal organism by feeding them targeted feed additives [
18,
19,
20].
Bacterial toxins are no less dangerous [
21]. It is well known that the secretion of proteins that damage host tissue is an integral part of the infectious processes of many bacterial pathogens [
22,
23]. In addition, toxins can be produced by many bacteria that live harmlessly in the animal’s body—representatives of normal and opportunistic microflora:
Escherichia coli,
Campylobacter spp.,
Shigella spp.,
Staphylococcus spp.,
Bacillus spp.,
Clostridium spp., etc. Individual species of these can cause infection, either by reaching a specific organ or under the influence of certain conditions [
24].
Much work has been undertaken to eliminate and/or reduce the incidence of anaerobic infections in veterinary medicine. Even today, despite the availability of specific prophylactics, chemotherapeutic agents, and antibiotics against most infections, clostridia remains a serious problem and causes considerable economic damage [
25,
26].
The pathogenic anaerobic microorganisms that are the causative agents of clostridia are widespread in nature [
27]. Their main reservoir is soil, and they also live in the intestines of animals and humans. Most clostridia are toxigenic infections. Exposure occurs through the formation of highly active toxins during growth, but other pathogenicity factors (hyaluronidase, lecithinase, collagenase, haemolysins, etc.) are also present. Clinically relevant clostridium species include the causative agents of independent diseases: tetanus, botulism, and emphysema, for example. However, in most cases the disease is caused by an association of species, as well as other anaerobes and aerobes. Although the clinical features of anaerobic diseases caused by different clostridial species differ greatly, there are several features that allow them to be grouped under the common name of Clostridiosis. These features include the ability to form spores, the anaerobic nature of the metabolic processes, the formation of highly active exotoxins, their habitation within animal intestines, and the fact that they often affect young animals with well-developed musculature [
28].
In humans and livestock,
C. perfringens causes intestinal infections, such as enteritis, enterocolitis, or enterotoxaemia. Enterotoxaemia in cattle is an acute or subacute syndrome with a mortality rate close to 100%, and is associated with the uncontrolled multiplication of
C. perfringens in the small intestine, with excessive production of toxins [
29]. These toxins act both locally and systemically, and can cause death within minutes to hours [
30].
Often, the problem of clostridial disease, as well as the development of mycotoxicoses, is associated with a failure to follow the technology of canned feed production, which results in secondary fermentation in the feed and proliferation of proteolytic bacteria, mainly clostridia. Cattle are most at risk during the grazing season, which means that disease control and prevention should be particularly targeted at this time of year [
31].
As all vaccines against clostridial disease are inactivated, two doses must be administered to heifers. In practice, however, the vaccinations are often administered too late, according to expert observations, and they are not always successful. In addition, spore-forming clostridia are resistant to broad-spectrum antibiotics, so their uncontrolled use in livestock further increases the risk of clostridial infections [
32].
One way of combating clostridial disease is to develop an integrated animal bioprotection function. The bioprotection function is based on a scientifically proven combination of sorbents and toxin degraders, with carefully selected biologically active substances to improve the functioning of the digestive system and restore the function of the immune system in animals [
33].
Anaerobic infections are a major problem for animal production worldwide. The damage consists of losses, not only from animal deaths, but also from unfavorable conditions in areas contaminated with pathogen spores and losses from quarantine measures and restrictions [
21]. The economic damage caused by reduced productivity can only be estimated approximately. Therefore, in addition to proper vaccination, additional means are needed to provide full protection against and prevention of clostridia, including the use of feed additives that neutralize not only the toxins, but also the clostridia themselves, and their spores [
34].
Research on the efficacy of feed additives for the prevention of clostridia in animals is still insufficient. It is of interest to study approaches to feeding animals complexes of substances that neutralize clostridia and/or their toxins. A new complex feed additive for dairy cows, which combines the functions of a sorbent, an immunomodulator, and a neutralizer, has been developed. Considering the specific character of the question under study, we have set an objective to investigate, on the one hand, the background values of clostridia presence in a modern Russian farm with a high level of fodder production and cattle breeding. On the other hand, we intend to reveal the possibility of preventing and increasing the immune status of cows with high productivity using a targeted bioactive feed additive, which can play an important role in the formation of new approaches in the integrated control of clostridia and its sequelae.
2. Materials and Methods
2.1. Study Site
The research was carried out on the grounds of Avangard LLC, located in the Ryazan region, Russia; in the laboratories of the feeding department, physiology and biochemistry department, and microbiology laboratory of farm animals at L.K. Ernst Federal Research Center for Animal Husbandry (FRCAH), located in Podolsk, Russia; in the laboratory of molecular and genetic studies of Biotroph Ltd., located in St. Petersburg, Russia.
2.2. Characteristics of the Compound Feed Additive
2.2.1. Composition of the Feed Additive
Bioactive feed additive (BFA) contains a toxin neutralizer, digestion modifiers, and natural substances of antibacterial nature (diatomite, enzyme–probiotic preparation, etc.).
For the experiment, experimental batches of multifunctional BFA were prepared. Composition of the additive, in 100 g:
- (1)
enzyme–probiotic component
- -
B. amyloliquefaciens159—not less than 6 × 107 CFU/g–0.1 g;
- (2)
antibacterial component
- -
lauric acid—4.2 g;
- -
pine bark extract (aqueous)—8.3 g;
- -
essential oil—0.064 g;
- (3)
sorbent constituent
- -
diatomite—40 g;
- -
wheat bran (as a filler)—55.8 g.;
The strain B. amyloliquefaciens159 was obtained from the collection of BIOTROF LLC (St. Petersburg, Russia). The original strain was extracted from the chyme of cow rumen. It is not a genetically modified organism.
The taxonomic identity of strain
B. amyloliquefaciens159 was previously confirmed by whole-genome sequencing using the MiSeq platform (Illumina, Inc., San Diego, CA, USA). Application has been made (SUB12907853; Taxonomy ID: 3029762; BioProject ID: PRJNA937299) to deposit the genomic sequence in the BioProjects collection (NCBI,
https://www.ncbi.nlm.nih.gov/bioproject/ (accessed on 21 Febuary 2023)).
According to the data obtained using conventional methods [
35,
36,
37] strain
B. amyloliquefaciens159 has no properties of toxigenicity and virulence, cell components of the strain are not toxic to laboratory animals.
Strain B. amyloliquefaciens159 was cultured at 37 °C on GRM nutrient broth medium of the following composition (g/L): enzymatic peptone, pancreatic hydrolysate of fish meal, sodium chloride, pH = 7.2 ± 0.2. (Siana LLC, Izhevsk, Russia).
2.2.2. Component Properties of the Feed Additive
In the laboratory of molecular genetic studies of Biotroph Ltd.:
- -
Anticlostridial activity of
B. amyloliquefaciens159 strains was determined. A daily culture of
B. amyloliquefaciens159 strain at a concentration of 1 × 10
8 CFU/mL, (grown on GRM nutrient broth at 37 °C for 24 h) was seeded with one linear stroke (strip) on the surface of dense RCB and blood agar media (BIOMEDIA LLC, Russia) along the diameter of a Petri dish, using a sterile swab. The cultures were incubated at 37 °C for 24 h.
C. perfringens indicator strains from the American Type Culture Collection (ATCC) [
38,
39] (13124, 10543, 12916) were seeded in tubes with thioglyco-left medium and incubated at 37 °C for 24 h under anaerobic conditions using Gaz Pak Anaerobe Container System (BD) (Thermo Fisher Scientific Inc., Waltham, USA). The results were evaluated according to GPhM. 1.7.2.0009.15 (Determination of the specific activity of probiotics) [
40]. The size of the no-growth zone of the test strain was considered.
C. perfringens belonged to different toxinotypes [
41]. Strain
C. perfringens 13124 belongs to type A, because it produces alpha toxin (CPA), and perfringolysin O (PfoA),
C. perfringens 12916 (type F) synthesizes CPA, and enterotoxin (CPE),
C. perfringens 10543 (type C) simultaneously produces 4 toxins, CPA, CPE, beta2 toxin (CPB2), Pfoa. This suggested a high level of virulence [
30,
42]. In addition, the antimicrobial activity of
B. mucilaginsous159 to other bacterial species, such as
E. coli K-12 F + Str. R (KS-507),
Salmonella typhimurium LT2,
Staphylococcus aureus ATCC 29213,
Pseudomonas aeruginosa TO (obtained from the National Bioresource Center All-Russian Collection of Industrial Microorganisms of the Kurchatov Institute (VKPM)), may also be associated with the occurrence of intestinal enteritis in animals [
43,
44]. Petri dishes with Mueller–Hinton agar medium (BIOLITE SPB, Russia) were pre-seeded with test strains. Then, holes with a diameter of 8 mm were made with sterile metal cylinders. 100 μL of daily culture of strain
B. amyloliquefaciens159 at a concentration of 1 × 10
8 CFU/mL, (grown on nutrient broth of GRM at 37 °C for 24 h) was placed in culture wells. Sterile GRM-bouillon nutrient medium was used as a control. Petri dishes were cultured in an aerostat at 38 °C. The level of antimicrobial activity was assessed after 24 h of growth.
- -
The bactericidal properties of lauric acid, a component of BFA, were determined. Lauric acid was introduced in concentrations (0.31; 0.16; 0.07 g/L, control without LA) into RCB agar culture medium (MERCK MILLIPORE, USA) to study growth zones of reference C. perfringens strains from ATCC collection (13124, 10543, 12916). Incubated under anaerobic conditions for 48 h.
- -
The true sorption capacity of diatomite with respect to aflatoxins (sum), ochratoxin A, T-2 toxin, zearalenone, and deoxynivalenol was determined [
45]. «True» sorption capacity of this BFA component was determined by the difference between adsorption and desorption of mycotoxins from solutions. «True» sorption capacity of the sorbent was calculated by the following formula:
where: C—true sorption capacity, %; A—adsorption of toxins in an acid medium simulating the pH level in the stomach/choke of animals %; D—desorption of toxins in the conditions that simulate the alkaline medium in the intestines of animals, %.
Adsorption meant the total amount of mycotoxins bound by the sorbent during the time the feed was in an acidic environment (pH = 2.5) simulating the conditions of animal stomach/intestine. To determine mycotoxin adsorption, a standard solution of mycotoxin was prepared with a concentration of 1000 µg/cm3. An exact weigh of 5.0 µg of dry mycotoxin standard (Romer Labs Inc., Getzersdorf, Austria) was dissolved in 5 cm3 of methanol or distilled water (in the case of deoxynivalenol). Diatomite was placed into the solution at a concentration of 0.02 g/mL. The sorption process was carried out for 1 h at 37 °C in the thermostat with constant stirring using a V-1 plus personal vortex (BIOSAN Ltd., Riga, Latvia). The resulting suspension was precipitated by centrifugation for 5 min at 10,000 rpm.
The supernatant and the initial solution were analyzed for mycotoxin content by competitive enzyme immunoassay using Agra Quant test systems (Romer Labs Inc., Austria) according to the manufacturer’s recommendations. Optical density was measured at λ = 450 nm using a Stat Fax 303+ microstrip photometer (Awareness Technology, Inc., Palm City, FL, USA).
Adsorption was calculated using the formula:
where
A—adsorption, %;
Ainitial—value of mycotoxin concentration in the initial working solution, μg/cm3;
Aresidual—value of mycotoxin concentration in the working solution after the sorption process, µg/cm3.
To determine desorption, the total amount of mycotoxin removed from the adsorbent surface was measured in an alkaline environment close in pH to the conditions in the ruminant intestine. For this purpose, the obtained suspension containing the adsorbent particles with mycotoxin bound to them was alkalized to pH = 7.0. The suspension was incubated in the thermostat for 1 h at 37 °C with constant stirring using a V-1 plus vortex (Biosan Ltd., Riga, Latvia). The obtained suspension was centrifuged for 5 min at 10,000 rpm.
The supernatant and the initial solution were analyzed for mycotoxin content by ELISA.
2.3. Animals and Housing
The object of the study were Holstein cows (importing country: Hungary). The total number of cattle in Avangard LLC at the time of the study was 1968 milking cows, 274 dry cows and 1467 calves.
For the given experiment, we selected 60 lactating Holstein cows of the second lactation with the average milk yield of 34.16 ± 0.45 kg, days in milking (DIM) = 39.5 ± 1.4 days, fat content = 3.53 ± 0.01% and protein content in milk = 3.05 ± 0.01%. Cows were previously ranked in descending order by DIM, milk yield, fat and protein content in milk, and allocated to control (C, n = 15) and experimental (E48, n = 15; E80, n = 15; E112, n = 15) groups to obtain 48, 80, 112 g/head/day BFA, respectively. This allocation procedure was adopted to ensure that all experimental groups had the same DIM, milk yield and milk parameters at d 0. Feeding of the supplement (specially prepared capsules) was carried out with cows of groups E48–E80–E112 daily in the morning hours after milking with an individual food probe.
Cows were kept in group stalls with unlimited access to water and total mixed ration (TMR; 1.5 m linear cow space per cow) during the whole period of the experiment. Cows were milked twice a day using a carousel system (05-00 and 16-00 h).
2.4. Experimental Diets of Cows
The diet for dairy cows met the energy and nutritional value requirements for high-yielding animals (32–36 kg milk yield) [
46,
47]. Calculation of feeding rations was carried out by using the software complex KormOptimaExpert (Version 2016, KormoResurs LLC, Voronezh, Russia) (
Table 1).
The chemical composition of feeds was determined at the Department of Animal Physiology and Biochemistry of the L.K. Ernst Federal Research Center for Animal Husbandry [
48]. Feeds were grounded at a Laboratory mill (LMC-1M, NV-Lab, Moscow, Russia) and then analyzed for dry matter (DM) and crude ash (CA) (ISO 6496-83 and ISO 5984, respectively). Initial moisture content was determined by drying the samples at 65 ± 2 °C to constant weight, and drying at 103 ± 2 °C for 3 h to constant sample weight for two successive readings that determined the hygroscopic moisture content of the samples. In dry samples, the raw ash content was determined by burning the sample in a muffle furnace at 550 °C. The nitrogen content was analyzed with ISO 5983-2-2016, from which crude protein (CP) content was calculated with formula (Nitrogen × 6.25), crude fiber with ISO 6865-2015, crude fat ISO 6492:1999, calcium (Ca) ISO 6490-1:1985, phosphorus (P) ISO 6491-2016; ME—calculation method [
48,
49].
The total duration of the study was 98 days. Cows were fed 48, 80, and 112 g/head/day of BFA during the whole experiment period.
2.5. Milk Performance and Analyses in Milk
Analysis of milk productivity during the experiment and during lactation was conducted according to Dairy Plan C21 (Version 5.285.093). The average daily milk yield was calculated from the results of control milkings before the experiment (d 0), on days 30, 60, 98 within the main period of the experiment (d 30; d 60; d 98). Average daily milk samples on these days were taken from each cow (n = 60) and in them, on an infraspectrometric analyzer (in the Department of Population Genetics and Genetic Bases of Animal Breeding at the L.K. Ernst Federal Research Center for Animal Husbandry) CombiFoss 7 DCC (FOSS, Denmark), fat and protein were determined by standard methods (ISO 9622/IDF 141:2013; AOAC 972. 16). Individual plastic flasks containing bronpol as a preservative were used for milk sampling. Based on the results obtained, the mean values for each indicator were calculated for the animal group before the start of the experiment and at subsequent sampling control points.
2.6. Vaccination and Level of Specific Immunity
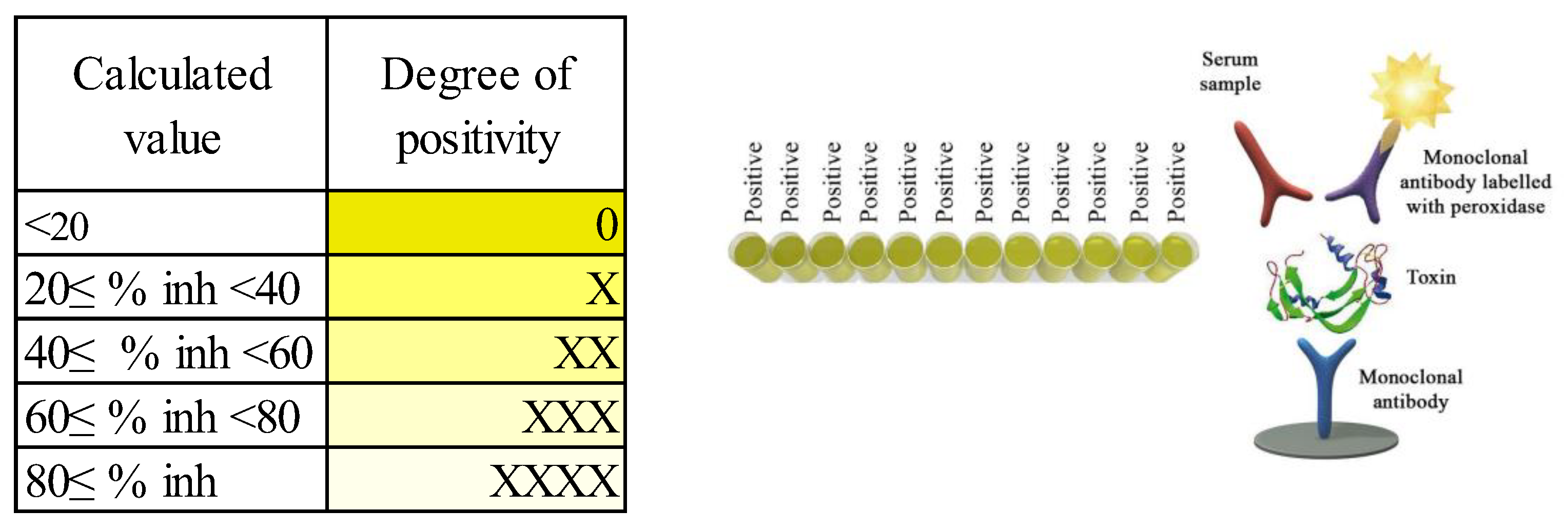
All cows were vaccinated with “Coglavax” (vaccine against bovine and sheep clostridial disease, Ceva-Phylaxia VeterinaryBiologicals, Hungary), reinjected two weeks before the experiment. This induces the formation of immune responses in cattle and sheep against alpha, beta, and epsilon anatoxins of C. perfringens types A, B, C, D, and others, from 2–3 weeks after reintroduction, which lasts for 10–12 months. Blood was drawn from animals of the experimental groups at the beginning of the experiment (n = 60, d0) and at the end of the experiment (n = 55, 5 animals dropped out of the experiment, d98). Blood samples from the subcostal vein were collected in vacuum containers (Vacuette, Greiner bioone, Kremsmünster, Austria), with blood clotting activator. Blood samples were stored on ice immediately after extraction and centrifuged 2 h after delivery. All samples were centrifuged at 4 °C at 4500 rpm for 10 min (Tagler centrifuge machine, table model SM-12 with rotor RU-06, NV-LAB, Moscow, Russia). Thereafter, serum samples were stored at −20 °C until analysis. In the Department of Feeding of Farm Animals L.K. Ernst Federal Research Center for Animal Husbandry, the content of antibodies to C. perfringens beta toxin in the blood serum were studied. The studies were performed on an automatic microplate photometer Immunochem-2100 (High Technology Inc, North Attleboro, MA, USA) by solid-phase enzyme immunoassay, using a Monoscreen AbELISA C. perfringens beta toxin reagent kit (BIO K 317/2, Bio-X Diagnostics, Rochefort, Belgique). Blood samples without hemolysis were collected (n = 40 at the beginning of the experiment, n = 50 at the end of the experiment). We also used 2 additional serum samples from animals diagnosed with C. difficile disease as positive controls.
The degree of positivity of each sample was evaluated using a scale (
Figure 1).
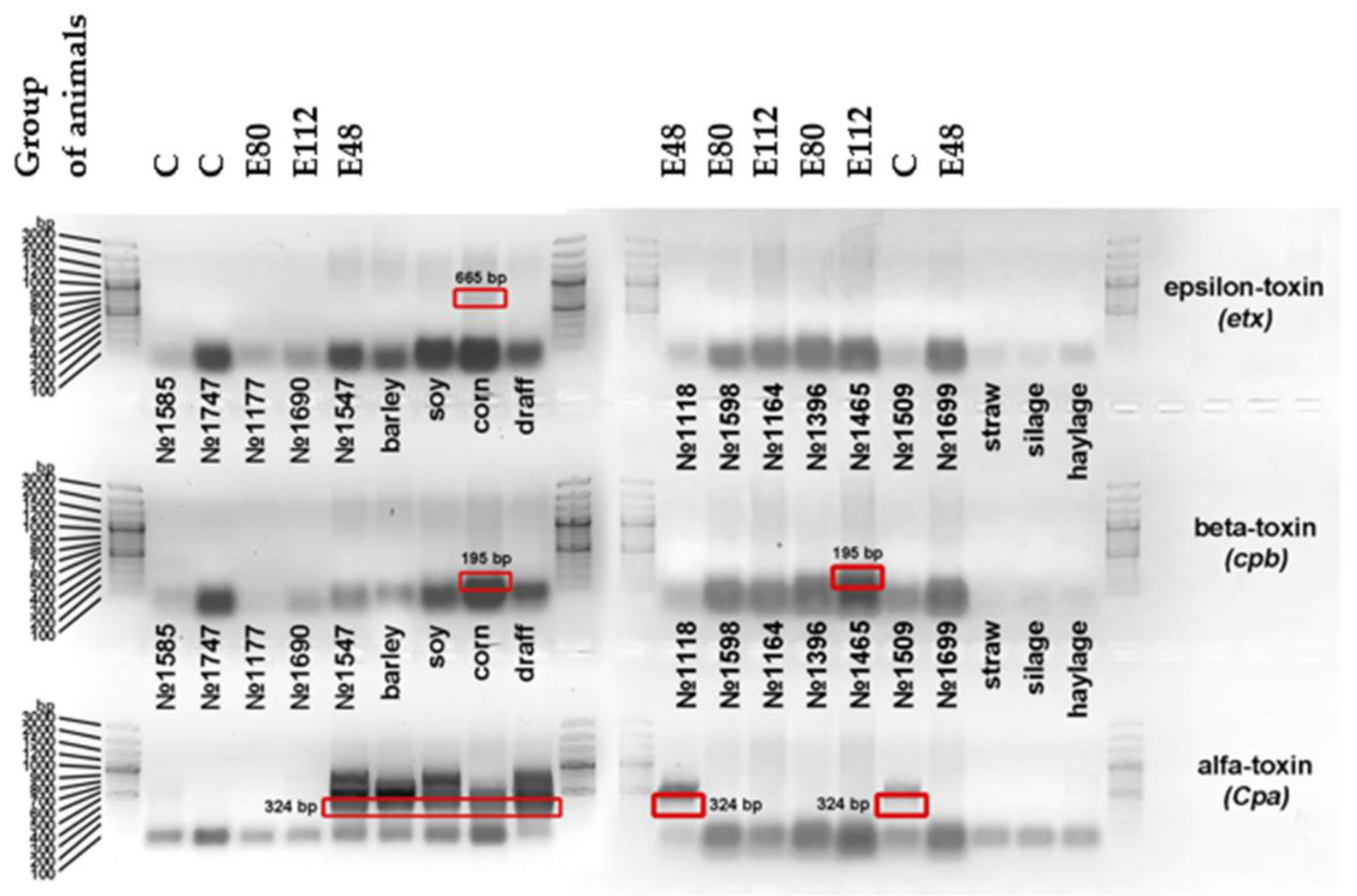
2.7. Molecular Genetic Studies to Detect Some Species of C. perfringens and Their Toxins in the Rumen Contents
Molecular and genetic studies were carried out by the laboratory of Biotroph Ltd. Samples of rumen contents sampled at the end of the experiment (n = 12, 3 animals from each group) and dietary feed (soybean meal, corn grains, DDGS, straw, corn silage; alfalfa haylage) were tested for the presence of toxin genes produced by microorganism C. perfringens: alpha-toxins 1 and 2 (Cpa); beta-toxin (cpb) and epsilon-toxin (ext). The analysis was performed using the PCR method. DNA was isolated using standard methods using the Genomic DNA Purification Kit (Thermo Fisher Scientific, Inc., Waltham, USA) according to the attached instructions. The following specific primers (5′→3′) were chosen for analysis: for the alpha-toxin Cpa gene, F: GCTAATGTTACTGCCGCCGTTGA, R: CCTCTGATACATCGTGTAAGAATC, for the cpb beta-toxin gene—F: GCGAATGCTGAATCATCTA, R: GCAGGAACATTAGTATATCTTC, for the etx epsilon-toxin gene—F: GCGGTGATATCCATCTATTC, R: CCACTTACTTGTCCTACTAAC.
Amplification reactions were performed using a DTLight amplifier (DNA-Technology, Moscow, Russia) and the Tersus Plus PCR kit (Eurogen, Moscow, Russia) according to the manufacturer’s protocol. Mode and conditions of amplification were as follows: 3 min at 94 °C (prewarming); 40 s at 94 °C, 60 s at 55 °C, 90 s at 72 °C (34 cycles).
2.8. Bacterial Strains and Culture Conditions
In the Laboratory of Microbiology of L.K. Ernst Federal Research Center for Animal Husbandry, in selected blood samples, by conventional methods, the indices of nonspecific resistance of experimental animals were determined: bactericidal, lysozyme activity of serum, and phagocytic activity of blood cells.
Strains E. coli ATCC 25922 and M. luteus (lysodeicticus) ATCC 4698 were obtained from Federal Budget Institution of Science «State Research Center for Applied Microbiology & Biotechnology». These bacteria were cultured in Tryptic Soy Agar (TSA) (Merck, Rahway, Germany) at 37 °C for 24 h. The cultures were suspended in phosphate-buffered saline and adjusted by Densi-La-Meret (PLIVA-Lachema Diagnostika, Brno, Czech Republic) for phagocytosis assay to 4.5 McF (E. coli), for bactericidal activity assay to 1.9 McF (E. coli), for lysozyme activity assay to 0.6 McF (M. luteus), and used within 15 min.
2.8.1. Phagocytosis Assay
E. coli culture (0.5 mL) was added to 0.5 mL of blood and incubated on shaker at 37 °C for 30 min. The sediment of mixture was smeared, fixed with 96% methanol, stained with Romanowsky–Giemsa method, and viewed under microscope (90×).
E. coli-engulfed neutrophils were counted as positive cells. We analyzed 100 neutrophils per slide. The following parameters were determined:
2.8.2. Lysozyme Activity Assay
Lysozyme was measured by turbidimetric method in a spectrophotometer USAMultiskan FC (ThermoFisher Scientific Inc., Vantaa, Finland). Tubes of blood serum (0.1 mL) were heated (56 °C) in a water bath for 30 min, then 1.4 mL of standard M. luteus culture was added and incubated at 37 °C for 3 h. The following parameters were determined: lysozyme activity of blood serum (LA), concentration of serum lysozyme (lysozyme, µg/mL), activity unit (AU) per 1 mg protein (AU/TP).
Lysozyme activity (LA) of a blood serum is calculated using the following formula:
Do is the difference in the optical density of the prototype,
Dk is the difference in the optical density of the control,
Do1 is the optical density of the prototype immediately,
Dk1 is the optical density of the control.
The concentration of lysozyme in serum was calculated based on calibration with dilutions of chicken egg-white lysozyme (L6876, Sigma-Aldrich, St. Louis, MO, USA) ranging from 0.1 to 51.2 µg/mL.
Due to variations in protein content in the blood serum of animals, the level of lysozyme activity was converted and expressed in arbitrary units of activity per 1 mg of protein (activity units per 1 mg of TP or AU/TP).
2.8.3. Bactericidal Activity of Blood Serum
Bactericidal activity (BA) of blood serum was measured by turbidimetric method in a spectrophotometer UNICO-2100 (United products & instruments, Ins., Suite E Dayton, NJ, USA) at OD540. E. coli culture (0.005 mL) was mixed with 4.5 mL of Tryptic Soy Broth (TSB) (Merck, Rahway, Germany) and 0.5 mL of blood serum in sterile tubes. Control was 0.5 mL of physiological saline with phosphate buffer instead of serum. All tubes were cultured at 37 °C for 5 h.
Percentage of BA was calculated from the following formula:
2.9. Blood Sampling and Analyses
Blood samples from the jugular vein were collected at the start (d 0) and at the end (d 98) of the main experimental BFA-feeding period at 09-00 h. Blood was collected in vacuum containers (Vacuette, Greiner bio-one, Kremsmünster, Austria), with clotting activator. At the same time, blood was drawn from each cow into vacuum containers (Vacuette, Greiner bio-one, Kremsmünster, Austria) with K3 EDTA. Blood samples were delivered to the Department of Physiology and Biochemistry of Farm Animals of L.K. Ernst Federal Research Center for Animal Husbandry within 2 h. To determine erythrocytes (RDC), leukocytes (WBC), hemoglobin (HGB), and hematocrit (HCT), a hematological analyzer ABC VET analyzer (Horiba ABZ, France) with Uni-Gem reagent kits (ReaMed, Russia) was used. To obtain serum, the samples were centrifuged (3000 rpm) for 20 min, with further storage of the separated serum at −20 °C until analysis. An automatic biochemical analyzer Erba Mannheim automatic XL-640 (Lachema s.r.o., Brno, Czech Republic) was used for analysis with determination of: AST, ALT by UV kinetic method without pyridoxal phosphate, IFCC; ALP—IFCC AM buffer; TP—biuret method; ALB—BKZ method; CREA—Jaffe kinetic method; UREA—Uricase GLDG, kin. method; TBIL—quantification by Walters and Gerarde method; electrolytes: Ca—ARSENAZO III method, P—Ammonium molybdate, Mg—Xylidine blue.
To assess the antioxidant status (AOS), the total amount of water-soluble antioxidants (TAWSA) in blood serum of experimental animals was determined by amperometric method (chromatograph, TsvetYauza 01-AA, SPA Himavtomatika PLC, Moscow, Russia) in samples. The concentrations of TBK-active products were determined using “TBK-Agat” kits (Agat-Med, Russia), and ceruloplasmin (CP) activity was determined using the Revin method. To characterize the prooxidant–antioxidant status of cows, the ratio of TBK-AP to ceruloplasmin (CP) levels was calculated.
2.10. Statistical Analyses
Statistical analyses of the data were performed with STATISTICA software (version 13RU, StatSoft, Inc., 2011;
www.statsoft.com (accessed on 27 December 2022)) by using a general linear model. Each group was considered as an experimental unit in measuring milk performance. For analyzing blood characteristic, the experimental units were individual animals. The relationship between the factor and the parameters was revealed using animal sampling, single-factor and two-factor analysis of variance (ANOVA), and Dunnett’s test, with testing of individual measures by Tukey’s multiple comparison method. Statistical differences were considered highly significant at
p < 0.01, significant at
p < 0.05 and values between
p ≥ 0.05 and
p ≤ 0.1.
2.11. Ethical Approval
Ethical approval for the study was provided by the bioethical commission of the L.K. Ernst Federal Research Center for Animal Husbandry (protocol #2022-02/1, dated 14 February 2022).
4. Discussion
In today’s livestock farming conditions, vaccination is a reliable way to control clostridium disease [
51]. A vaccine helps the body’s immune system identify and eradicate life-threatening pathogens such as
Clostridium. However, due to the widespread distribution of pathogens in the environment, the acute or hyperacute course of the disease, and the severity of tissue damage in the body, treatment of animals is almost 100% ineffective compared to specific prophylaxis [
26]. Earlier research [
26] had shown that clostridiosis in productive cattle mainly occurs in the form of malignant oedema, with the following prevalence:
C. septicum—found in 34.5% of cases,
C. perfringens type A—23.25%,
C. perfringens type C—14.25%,
C. perfringens type D—6.5%,
C. oedematiens—2.5%,
C. sordellii—6.5%. According to [
52]
C. perfringens type A (56.66%), type D (26.66%) and type B (16.66%) were, respectively, the most common bacteria in gut samples from 30 cattle and calves with enterotoxemia. On the other hand, there is evidence that dairy cows are not a major source of the potentially pathogenic enterotoxin gene for humans [
53]. The type diversity of
C. perfringens was low because primarily type A was cultivated. The enterotoxin gene was present in only 0.8% of all isolates. Nevertheless, it poses a serious risk and causes considerable economic damage to the industry [
34].
C. perfringens causes several diseases in animals and humans, including histotoxic and intestinal infections.
C. perfringens toxins can be divided into major, minor and enterotoxins. Strains of
C. perfringens are classified into five toxinotypes (A, B, C, D and E) based on the production of four major toxins. Chromosome-encoded CP toxins type A (CPA) are causative agents of histotoxic infection in humans and animals, and are the leading pathogenetic factors in the development of gas gangrene. The structural genes encoding these toxins,
cpa (or
plc), are located on the chromosome. CPA activates the extracellular signal-regulated kinase pathway, and thereby induces oxidative stress in the affected cells and the production of interleukin-8 by stimulating the action of kinase and mitogene-activated protein kinase. Beta-toxin SR (CPB), a pore-forming toxin, has a similarity to the amino acid sequence of the pore-forming toxins of
Staphylococcus aureus in 20–28% of strains. This toxin is extremely sensitive to trypsin.
In vivo, CPB causes necrotizing enteritis in sheep, cattle and horses [
28]. Epsilon-toxin CP (ETX) is considered to be the most potent clostridial pore-forming toxin after botulinum and tetanus toxins, and is the cause of necrotizing enteritis and enterotoxemia in sheep, cattle and horses. Thus, the role of each
C. perfringens toxin has been established in animal disease, and the mechanism of action of these toxins in vivo has been studied, which makes it possible to develop more effective vaccines against diseases caused by these microorganisms [
30].
It is now generally accepted that the unjustified use of antibiotics to prevent and treat infectious diseases causes selection pressure on bacteria, leading to increased antimicrobial resistance and loss of microbial diversity in the environment [
54]. Feed additives should be an additional prevention strategy. Therefore, in developing an effective natural feed additive for the prevention of clostridial diseases, we relied on modern approaches to improve the specific and non-specific resistance of livestock. Antimicrobial peptides, immune regulators, probiotics and bacterial toxin binding mannan oligosaccharides [
55,
56,
57] are known to be used as preventive agents. The frequent lack of significant treatment effects, as well as the low reproducibility of the results obtained, leads to the conclusion that, although various feed additives may contribute to the health and immune status of animals, no conclusions can be drawn from these results regarding their effect on the prevention of clostridiosis. According to some researchers [
58], high efficacy of treatment and prevention of bovine diseases such as abomasitis, and enteritis caused by
C. perfringens, can be achieved by using a comprehensive approach. This should include prevention of
C. perfringens proliferation, reduction of exotoxin production, restoration of normal gastrointestinal microflora and gut architecture. The use of probiotic strains selected for the biodegradation of toxic compounds in feed additives is promising [
59]. The use of probiotic strains and immunostimulants has great potential as therapy, but requires further development [
56,
60]. Such bacteria, in addition to their powerful antimicrobial effect, are capable of exerting versatile effects on the host organism. Probiotics, used for prevention or treatment of clostridia, act through multiple possible mechanisms, including the restoration of normobiotic resistance, antagonistic activity, and synthesis of a wide range of metabolites, which have a protective effect on the main targets of
Clostridia [
61,
62,
63]. They protect cells from damage and reduce the level of expression (activity) of apoptosis (cell death) and inflammation genes [
64,
65].
The choice of
B. amyloliquefaciens159 as the basis of BFA was based on numerous data [
66,
67] that suggest that
Bacillus bacteria can effectively restore the composition of the host microbiota, increase its resistance to infectious diseases, and contribute to improved animal performance. Currently, various commercial probiotics contain bacilli as active ingredients, due to their ability to colonize the digestive system, their high stability in relation to endosporogenesis, which allows them to survive in stressful situations, such as the aggressive conditions of the internal environment of the body, and their longer shelf life [
68]. Representatives of this taxon are known to be capable of synthesis of a wide range of metabolites with antimicrobial properties, including bacteriocins (subtilin, bacillocin 22, sublancin 168 and others) that have an antagonistic effect against a wide range of pathogens [
69,
70]. In an in vitro experiment, we observed pronounced antimicrobial, particularly anticlostridial, activity of
B. amyloliquefaciens159, which was probably due to the diffusion of antimicrobial metabolites into agar. Neutralizing toxins, antioxidants, and increasing the nonspecific resistance of animals can serve as important additions for the complex prophylaxis of clostridiosis. Lauric acid has strong antimicrobial activity due to its function of disordering lipid membranes of microorganisms [
66,
71]. Pine extract (pycnogenol) has antioxidant activity due to its high content of procyanidides, catechins, polyphenols, as well as antimicrobial and anti-inflammatory properties [
72]. We used diatomite as an enterosorbent in BFA. The unique firing technology, at temperatures of 800 °C, significantly increases the specific surface area of the sorbent (up to 40 ha/kg), which makes it the most effective among other sorbents [
46,
73]. More research is needed to study the use of neutralizers of clostridial toxins, as well as the role of individual substances in the formation of additional animal bioprotection.
In a healthy animal or poultry organism, immunity must perform the task of resisting clostridia. However, antibiotics, often used in the treatment of
Clostridia, not only do not increase it, but also reduce it, making the organism more helpless and unable to defend itself [
74]. Our study was based on previous findings, that certain strains of the bacteria
Bacillus spp. Are biostimulants of non-specific immunity [
75]. Some strains are known to reduce the effects of oxidative stress, apoptosis and inflammatory gene expression levels [
76]. In addition, pine bark extract, chosen as an ingredient of BFA, is also known for its immunomodulatory properties [
77]. The combined action of these components of the feed additive resulted in a significant increase in the level of humoral immunity of cows, which is especially important at the beginning of lactation. The main integral indicators of natural resistance of cattle are the bactericidal (BA) and lysozyme activity (LA) of blood serum. Using the feed additive increased the concentration of lysozyme in the blood serum of cows of group E112 by 3,6 times compared to the control (
p < 0.001). BA levels increased by 10.7% (
Table 3). It is important to note that vaccines against clostridia are not always effective. In some cases, experts attribute their lack of effectiveness against clostridia to the low level of production in animals of specific antibodies in response to the introduction of the antigen [
78]. Prior to the experiment, the animals of the experimental groups were vaccinated with a preparation against bovine clostridiosis. The drug should cause formation of immune response in animals against α-, β- and ε-anatoxins of
C. perfringens types A, B, C, D and others, 2–3 weeks after repeated administration, and immunity in this case should be maintained for 10–12 months. The use of BFA in the feeding of dairy cows contributed to a better serological response after immunization with the vaccine and natural contact with
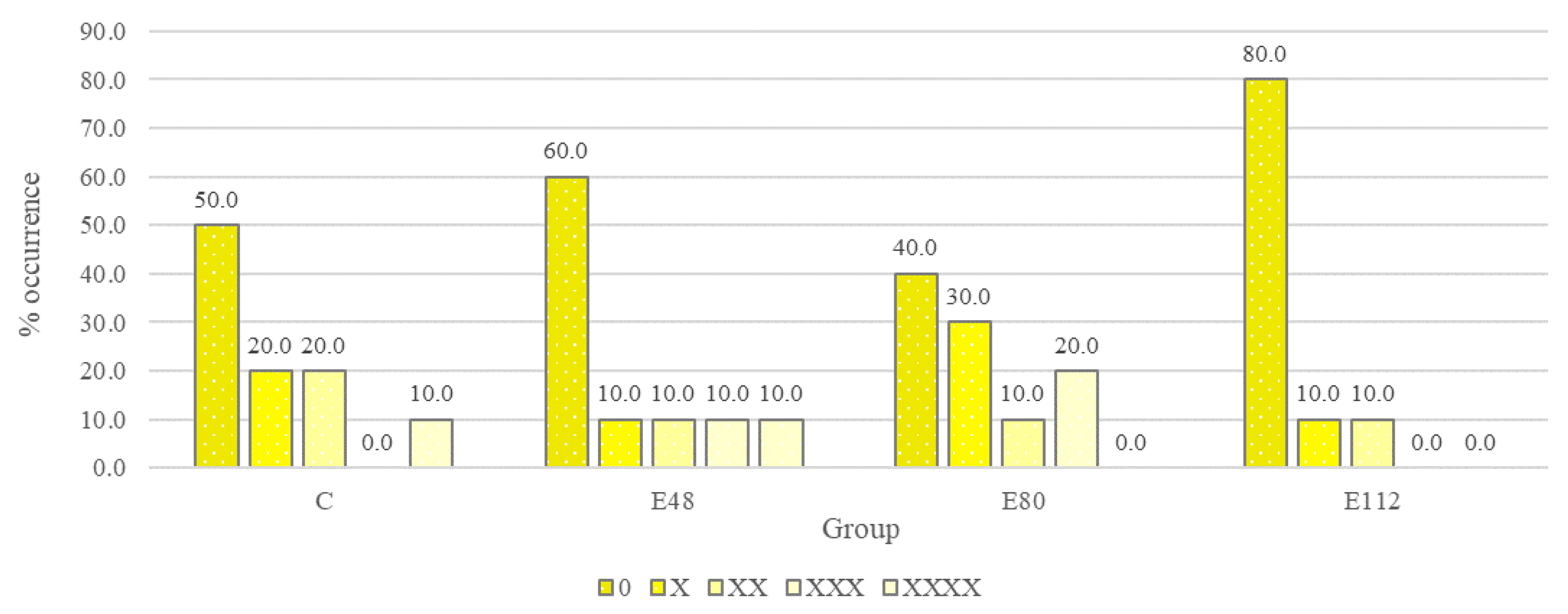
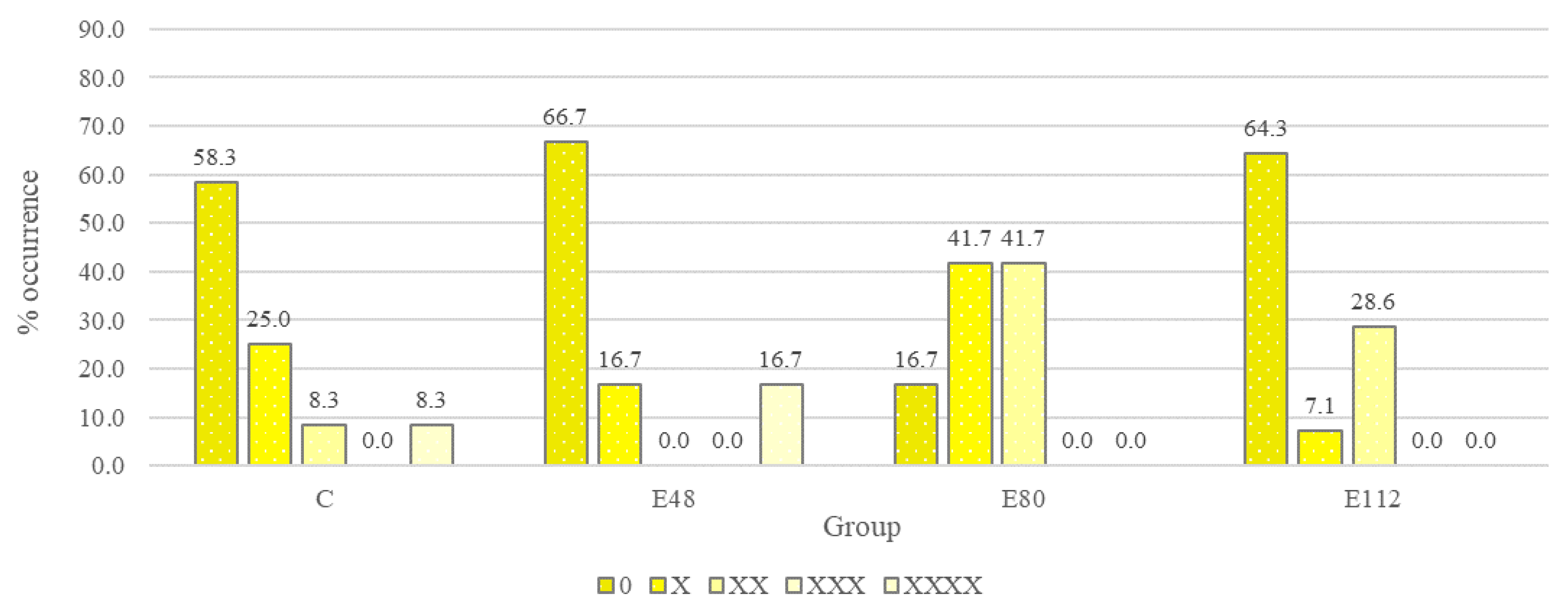
C. perfringens. The level of serological response in the control group decreased slightly after three months of experience. At the beginning of the experiment, a positive serological response was observed in 50% of the animals, and at the end of the experiment the serological response rate was 41.6% of the number of animals examined (
Figure 5). In contrast, the serological response rate in the experimental groups increased at the end of the experiment. The positive effect of the drug on the effectiveness of vaccination is explained by the fact that the intestinal tract of mammals is the largest immune organ and consists of cells of non-hematopoietic (epithelium, Paneth cells, goblet cells) and hematopoietic (macrophages, dendritic cells, T cells) origin, while simultaneously being a home to symbiotic microbiota, which largely modulate immune cell activity, including through regulation of gene expression [
79]. A disturbed microbiome, which is observed in most animals in intensive livestock farming conditions [
80], is capable of causing a reduction in vaccination efficacy. Intestinal dysbiosis contributes to systemic inflammation, immunosuppression and a lesser response to vaccines. It has previously been demonstrated in model organisms that dysbiosis reduced specific IgG responses to various vaccines, and the transplantation of fecal commensal microbiota helped to stimulate vaccine responses [
81]. Due to the healing of the microbiome and the manifestation of immunomodulatory activity in our experiment, the biopreparation had a positive effect on postvaccination immunity. It is known that the microbiome can influence the efficacy of vaccination through various mechanisms, including direct effects (e.g., through the structural component of the flagellin protein), as well as the synthesis of lipopolysaccharides, volatile fatty acids and other metabolites that can activate Toll-like receptors, and, in turn, stimulate the response of dendritic cells [
82]. Thus, Bron et al. [
83], after analyzing the available data on the adjuvant activity of three strains of lactobacilli, noted a role in improving vaccination efficacy in compounds such as lipoteichoic acid in
Lactiplantibacillus plantarum, the GG (also known as LGG) proteins p40 and p75 secreted by
Lacticaseibacillus rhamnosus, and surface layer protein A (SlpA) in
L. acidophilus strain NCFM. Earlier results in poultry have also shown that a number of probiotic bacteria increase efficacy against Newcastle disease [
84] and avian influenza [
85].
Active oxygen species (AOS) are produced during mitochondrial oxidative metabolism as well as in the cellular response to xenobiotics [
86] and changes in the rumen microbiome. When AOS suppress the cellular antioxidant defense system, whether by increasing AOS levels or decreasing cellular antioxidant capacity, oxidative stress occurs, affecting the rumen microbiome among others [
87]. In turn, it is known that the microbiota of the digestive system can affect the levels of enzymes that metabolize xenobiotics, both in the gut and in the liver [
88]. Glutathione (GSH) is present in all mammalian tissues, as the most abundant non-protein thiol that protects the body against oxidative stress [
89]. GSH is also a key determinant of redox signaling. Being vital for xenobiotic detoxification, it regulates cell proliferation, apoptosis, immune function and fibrogenesis. GSH biosynthesis occurs in the cytosol in a highly regulated manner. The interrelation of antioxidants and glutathione is caused by their overproduction of indirect biological activity, allowing them to realize their action through signal pathways, mediated by NF-κB and LPS/TLR-4 (proinflammatory cascades), STAT3 and MAR kinases, PI3K/Akt/mTOR (autophagy), and G-protein coupled receptors [
90,
91,
92]. Comparative analysis of the total amount of water-soluble antioxidants is directly related to the induction of ARE-dependent phase II xenobiotic detoxification enzymes and antioxidant enzymes directly involved in the induction of total GST activity in U937 myeloid cells, catalyzing reactions of chemical modification of xenobiotics with GSH participation (conjugation and nucleophilic substitution reactions of xenobiotics and GSH, and reduction of organic peroxides to alcohols with the formation of GSSG). Thus, the accumulation of antioxidants indicates a better level of stress resistance in productive animals, adaptive qualities, and mobility of biochemical pathways in the body under the conditions of intensive milk production. Our data indicate that the feeding of BFA led not only to an increase in TAWSA, but could also improve the antioxidant properties of the milk. The conjugacy of lipid peroxidation and the antioxidant system can be assessed by calculating the correlation between a number of components of these systems and the correlations between them (
Table 6). The TBA–AP / CP ratios of blood serum of cows we determined were in direct relation to the TBA–AP content, and decreased in the blood of Group E48 animals (
p < 0.01). The highest TBA–AP content was observed in the blood serum of the control group, which probably indicates the activation of free radical processes in cows against the background of high productivity at the beginning of lactation. Thus, the use of BFA significantly improved the antioxidant protection of the body of the experimental animals. We attribute some differences in blood values with physiological norms at the beginning of lactation (
Table 4), firstly to the physiological period (cows average 34th day of lactation), secondly to high productivity, and thirdly to feeding. There were intergroup differences (AST, UREA,
p < 0.05; WBC,
p < 0.01) but they were not systematic. Under the influence of the factor under study, important parameters, such as TP, GLB, A/G, and RDC, stabilized and normalized in the animals of groups E80 and E112, compared to the animals of groups C and E48. Therewith, the following intergroup differences were observed: under the influence of BFA feeding, the TP content in blood serum decreased to normal (
p < 0.001), ALB level increased (
p < 0.05), and GLB decreased to normal (
p < 0.001). This stabilized the A/G-ratio from 0.47–0.49 to 0.64–0.71 (
p < 0.001) in the E80 and E112 groups, against the background of urea and ALT increase (
p < 0.01). This indicates an optimization of protein metabolism when using BFA, and is also consistent with the higher milk productivity of the experimental groups. Better productivity and milk fat content were connected with the improvement in carbohydrate-lipid metabolism: the blood serum cholesterol concentration in the cows of the experimental groups increased (
p < 0.01). GLU in blood of group C animals was below the norm, while in animals of experimental groups its concentration corresponded to the norm (
p > 0.05). It is also necessary to note the tendency (
p < 0.10) of WBC to decrease in the blood serum of cows in group E112, which reflects the positive effect and characterizes the decrease in inflammatory processes in animals of experimental groups under the influence of BFA. It should be noted that during the three months of the experiment, the cows in the experimental groups maintained their productivity at a high level, unlike the animals in the control group. At the end of the experiment, the average daily milk yield was higher in the experimental groups (
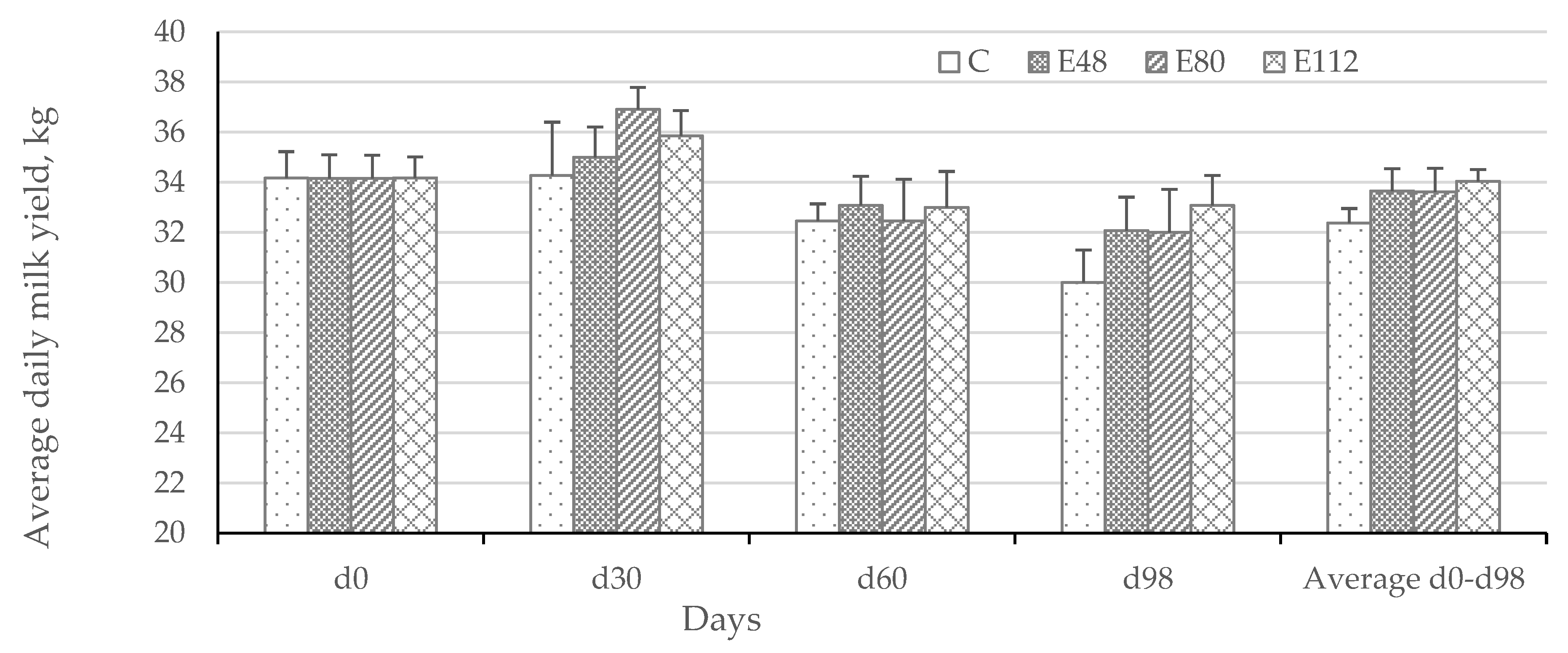
Table 7), which indicates not only the ability of the chosen strategy to prevent clostridiosis and improve cattle health and resistance, but also shows the short-term effect expressed in additional milk production.




 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 —below normal,
—below normal,  —above normal. 3 According to [50].
—above normal. 3 According to [50]. —below normal,
—below normal,  —above normal. 3 According to [50].
—above normal. 3 According to [50].