
Akkermansia muciniphila Promotes Bone Development and Improves Eggshell Quality during the Sexual Maturity Period of Laying Hens by Increasing Osteogenesis



Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Akkermansia muciniphila Suspensions and Lyophilized Powder
2.2. Ethics Statement
2.3. Animals
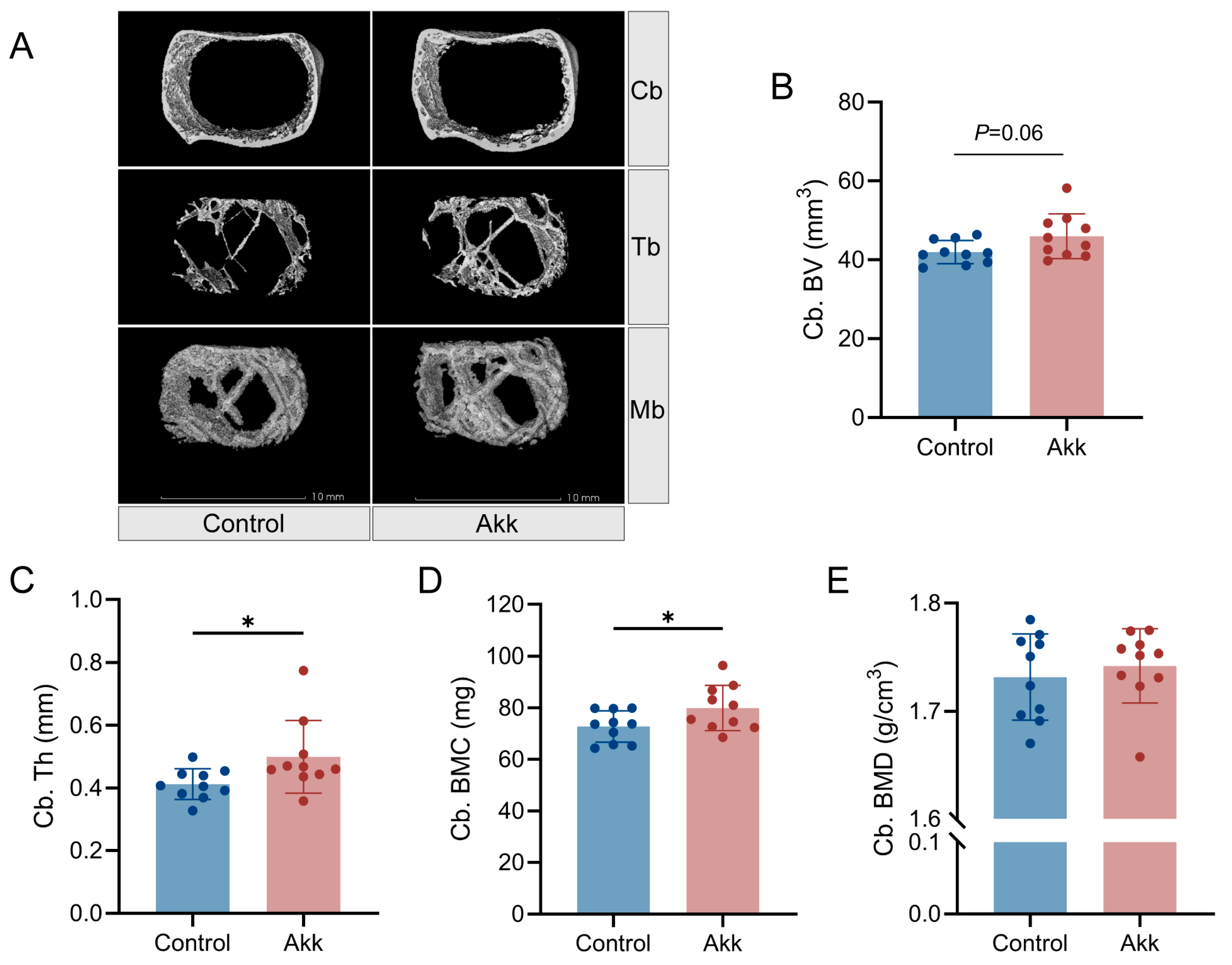
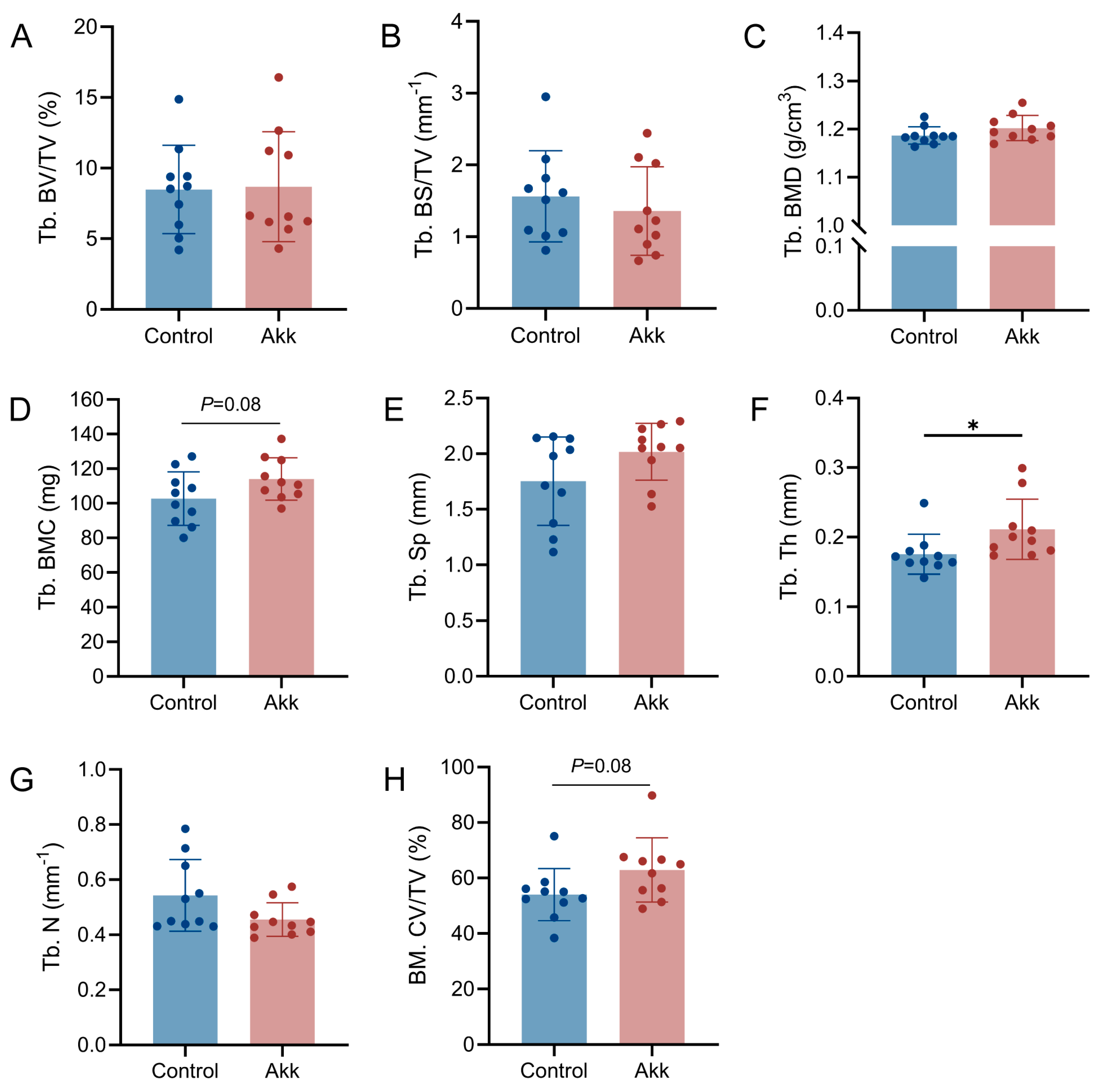
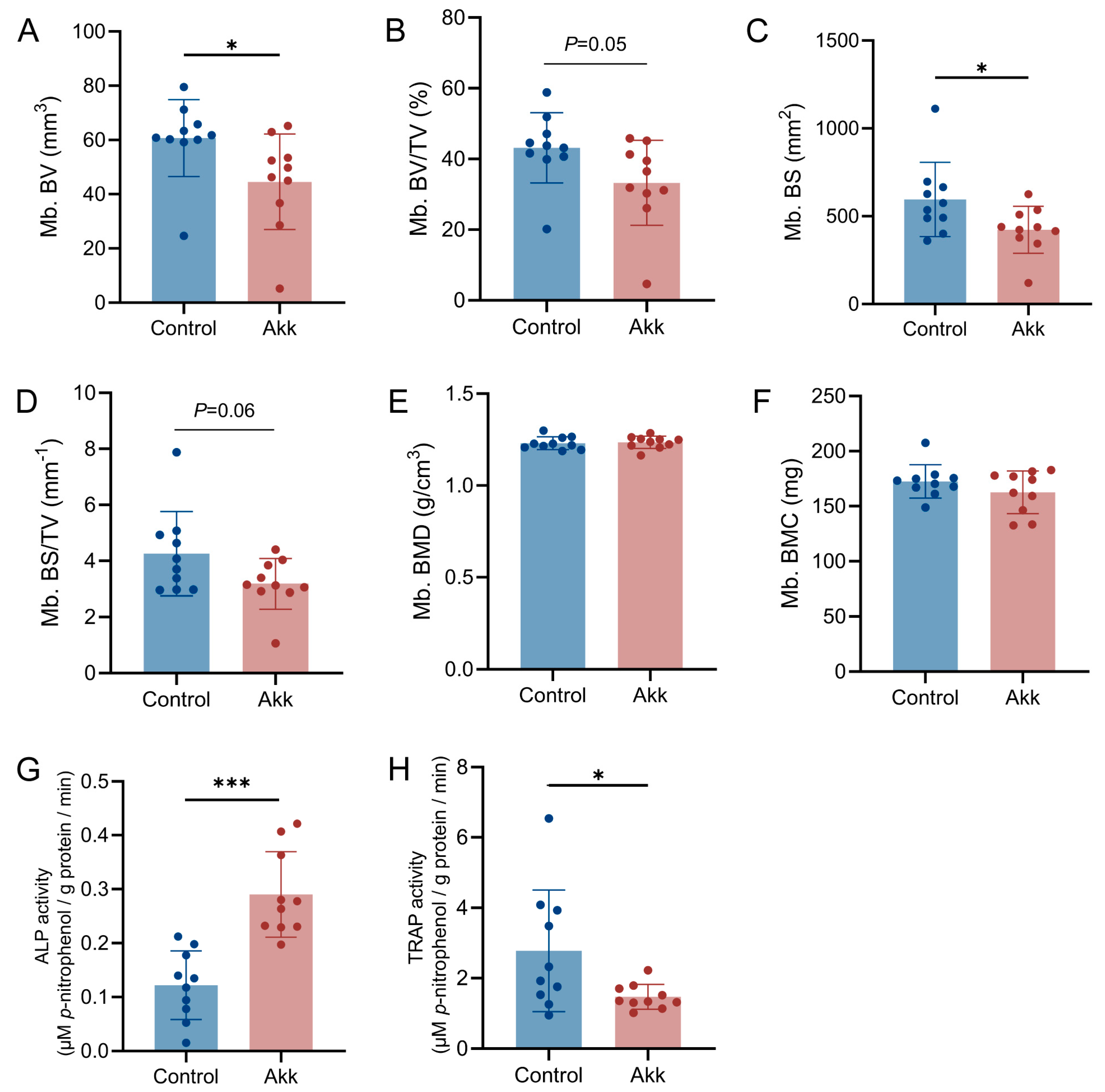
2.4. Micro-Computed Tomography Analysis
2.5. Determination of Alkaline Phosphatase Activity
2.6. Determination of Tartrate-Resistant Acid Phosphatase Activity
2.7. 16S rRNA Gene Sequencing and Analysis
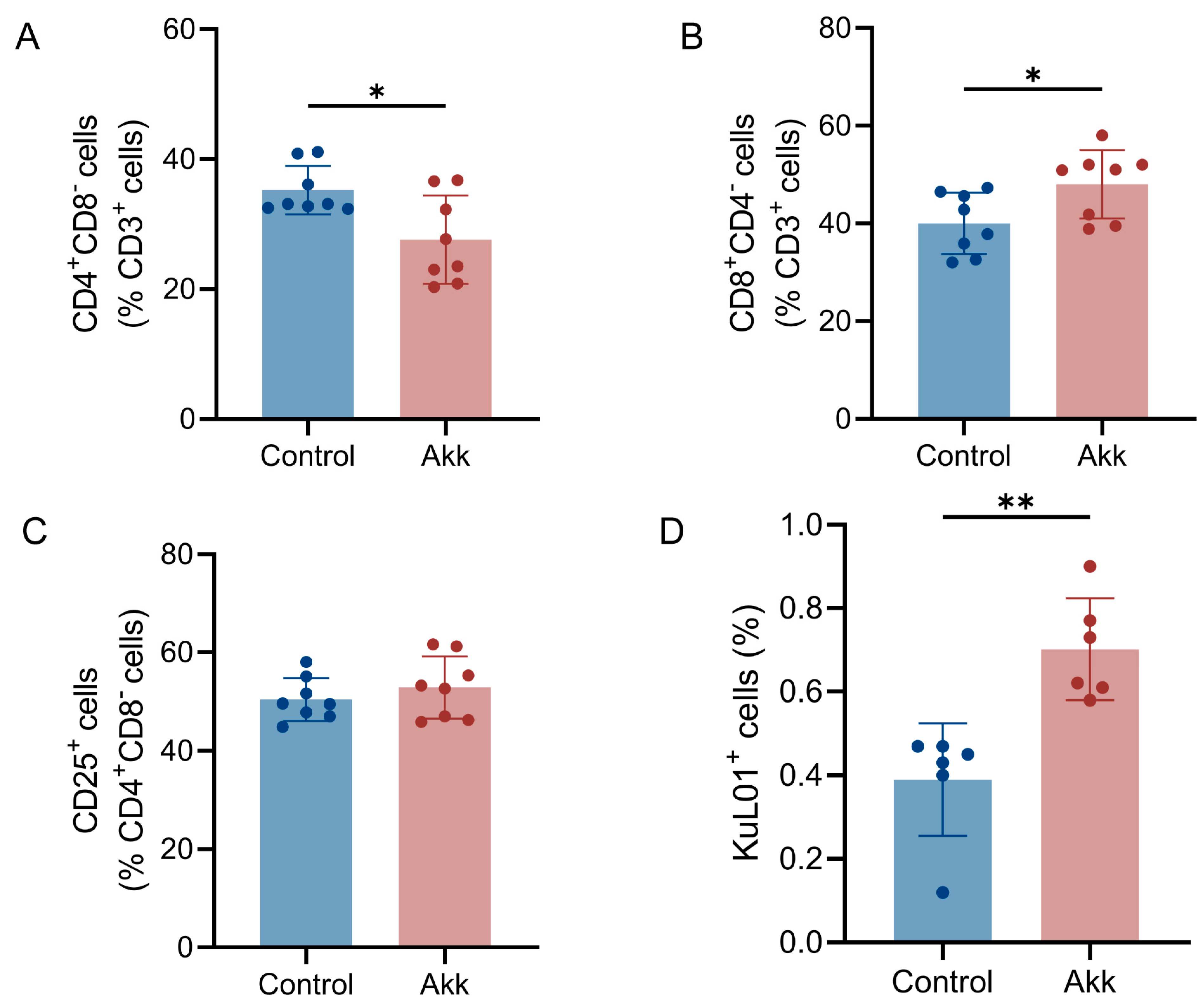
2.8. Preparation of Lymphocytes and Flow Cytometry
2.9. Measurement of Egg Quality
2.10. Statistical Analysis
3. Results
3.1. Effect of Akkermansia muciniphila Powder on Growth Performance of Laying Hens
3.2. Dietary Supplementation of Akk Powder Promotes Bone Development
3.3. Effect of Akk on Immune Cells Associated with Bone Remodeling in Bone Marrow
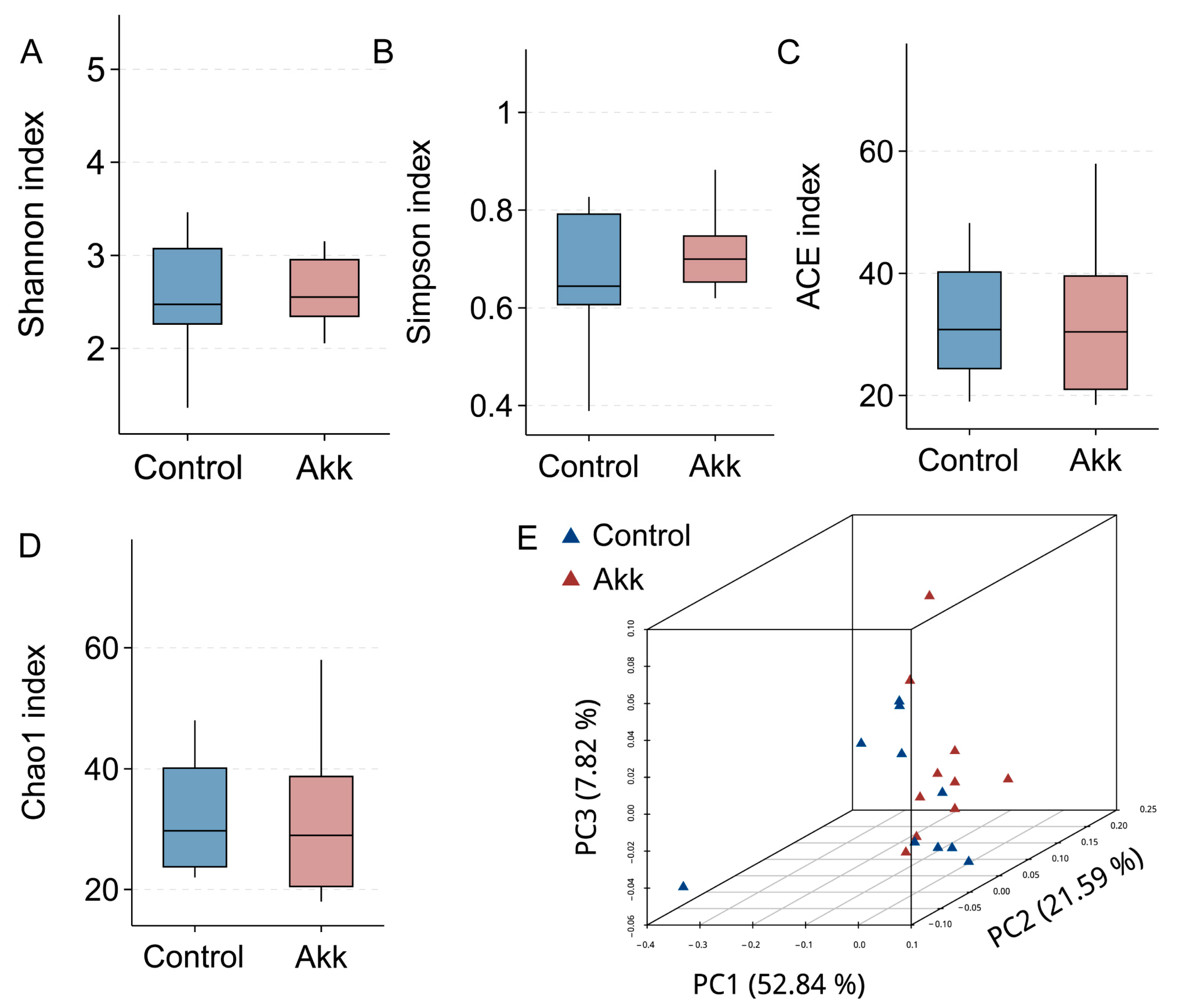
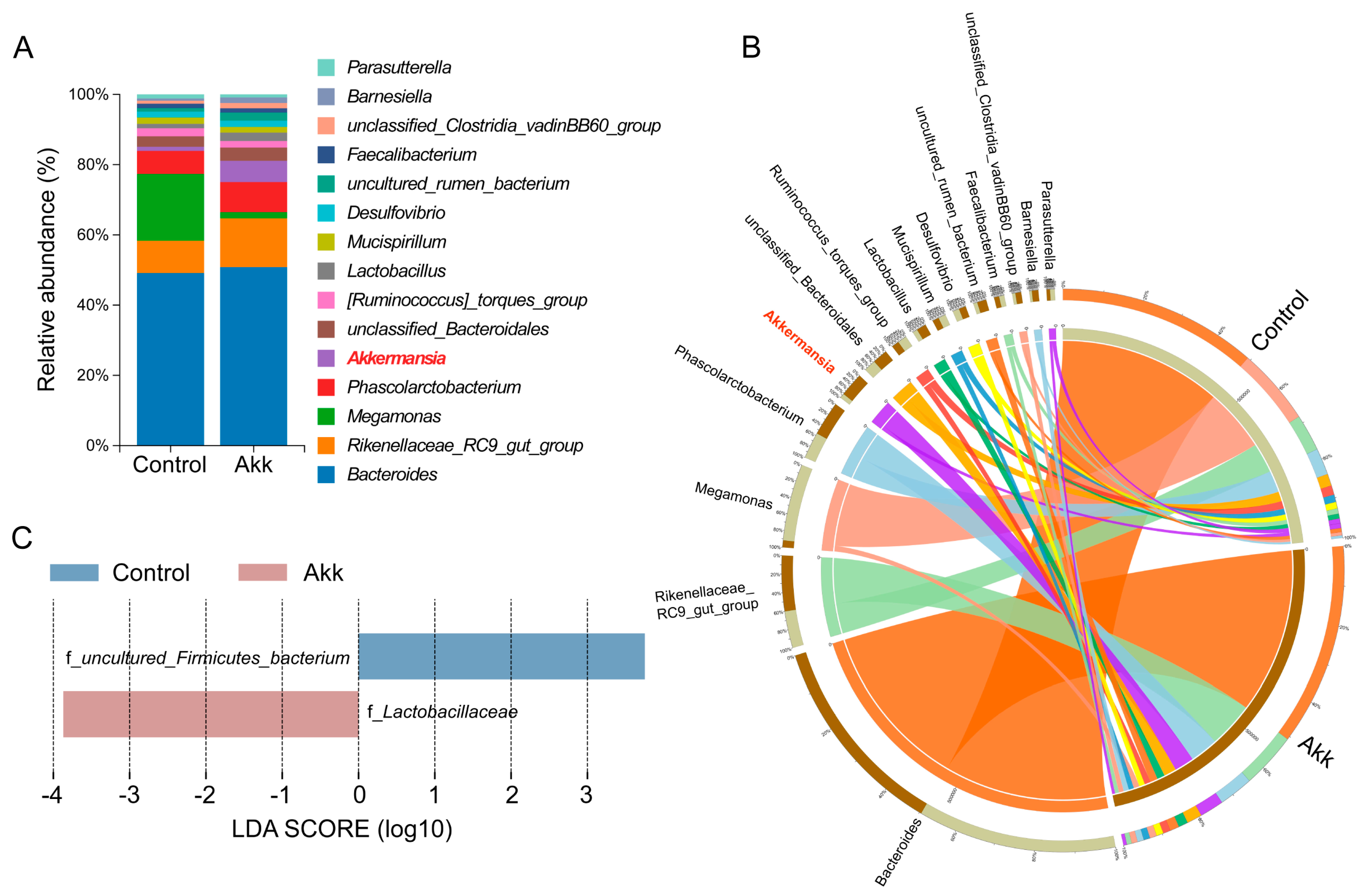
3.4. Effect of Akk on the Abundance and Diversity of Cecal Microbiota
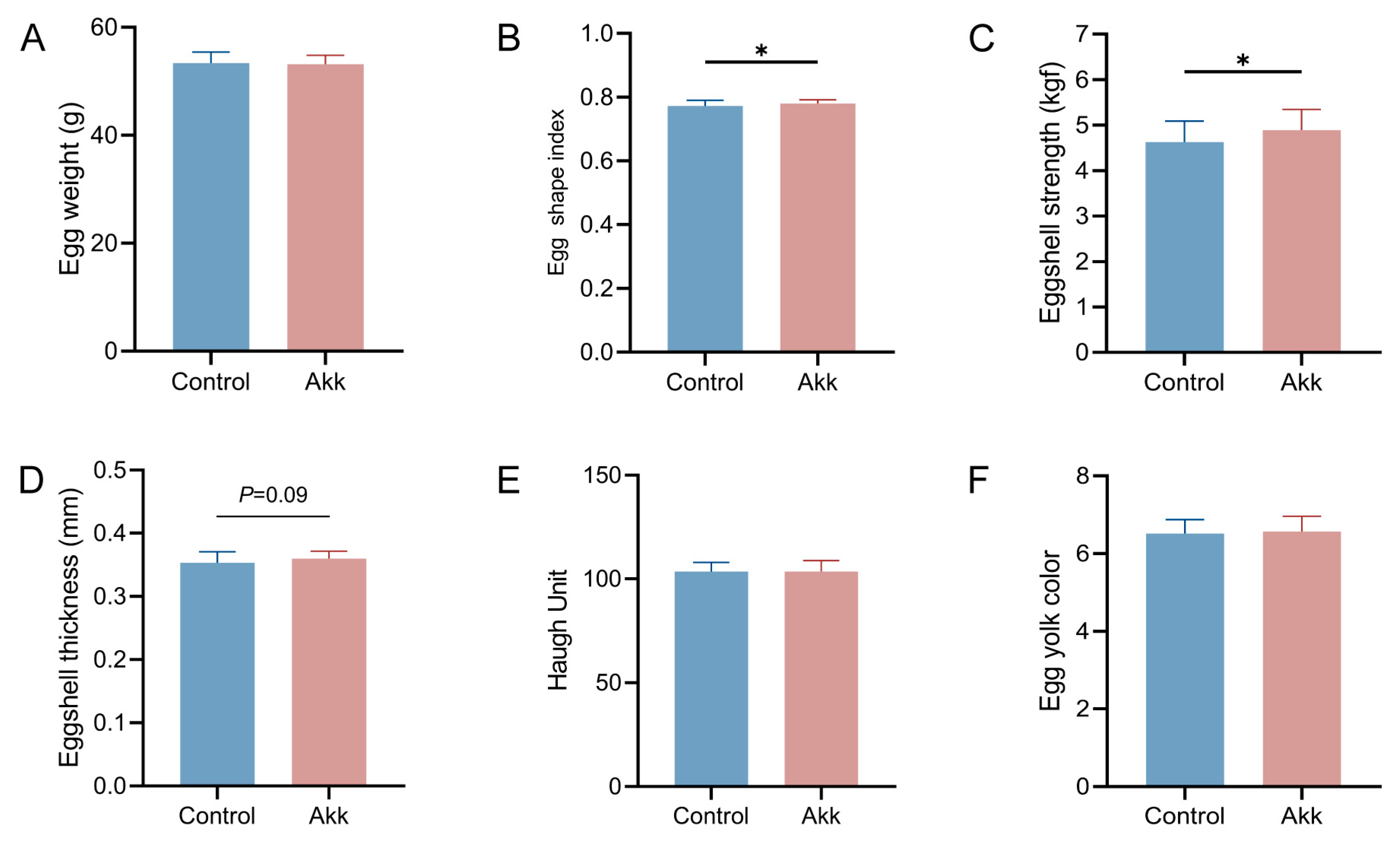
3.5. Dietary Supplementation of Akk Powder Increases Eggshell Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Akk | Akkermansia muciniphila |
| Cb | Cortical bone |
| Tb | Trabecular bone |
| Mb | Medullary bone |
| OVX | Ovariectomized |
| GAM | Gifu Anaerobic Medium |
| ALP | Alkaline phosphatase |
| TRAP | Tartrate-resistant acid phosphatase |
| LEfSe | Linear discriminant analysis effect size |
| ADFI | Average daily feed intake |
| RANKL | Receptor activator of NF-κB ligand |
| PCoA | Principal coordinate analysis |
| Cb. BV | Cortical bone volume |
| Cb. Th | Cortical bone thickness |
| Cb. BMC | Cortical bone mineral content |
| Cb. BMD | Cortical bone mineral density |
| FMT | Fecal microbiota transplantation |
| RT | Room temperature |
| Tb. BMC | Trabecular bone mineral content |
| Tb. Th | Trabecular bone thickness |
| Tb. BV/TV | The ratio of trabecular bone volume to total femur volume |
| Tb. BS/TV | The ratio of trabecular bone surface area to total femur volume |
| Tb. BMD | Trabecular bone mineral density |
| Tb. Sp | Trabecular bone spacing |
| Tb. N | Trabecular bone number |
| BM. CV/TV | The bone marrow cavity volume to the total femur volume |
| Mb. BV | Medullary bone volume |
| Mb. BV/TV | The ratio of medullary bone volume to total femur volume |
| Mb. BS | The medullary bone surface area |
| Mb. BS/TV | The ratio of Mb. BS to total femur volume |
| Mb. BMD | Medullary bone mineral density |
| Mb. BMC | Medullary bone mineral content |
References
- Bain, M.M.; Nys, Y.; Dunn, I.C. Increasing persistency in lay and stabilising egg quality in longer laying cycles. What are the challenges? Br. Poult. Sci. 2016, 57, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Korver, D.R. Review: Current challenges in poultry nutrition, health, and welfare. Anim. Int. J. Anim. Biosci. 2023, 17 (Suppl. 2), 100755. [Google Scholar] [CrossRef] [PubMed]
- Sandilands, V. The laying hen and bone fractures. Vet. Rec. 2011, 169, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.C.; Fleming, R.H. Osteoporosis in cage layers. Poult. Sci. 2000, 79, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- van de Velde, J.P.; Vermeiden, J.P.; Bloot, A.M. Medullary bone matrix formation, mineralization, and remodeling related to the daily egg-laying cycle of Japanese quail: A histological and radiological study. Bone 1985, 6, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Navarro, A.B.; McCormack, H.M.; Fleming, R.H.; Alvarez-Lloret, P.; Romero-Pastor, J.; Dominguez-Gasca, N.; Prozorov, T.; Dunn, I.C. Influence of physical activity on tibial bone material properties in laying hens. J. Struct. Biol. 2018, 201, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, C.; Kieser, S.; Çolakoğlu, M.; Hadadi, N.; Brun, J.; Rigo, D.; Suárez-Zamorano, N.; Spiljar, M.; Fabbiano, S.; Busse, B.; et al. Warmth Prevents Bone Loss Through the Gut Microbiota. Cell Metab. 2020, 32, 575–590.e577. [Google Scholar] [CrossRef] [PubMed]
- Rauch, F. Bone growth in length and width: The Yin and Yang of bone stability. J. Musculoskelet. Neuronal Interact. 2005, 5, 194–201. [Google Scholar] [PubMed]
- Whitehead, C.C. Overview of bone biology in the egg-laying hen. Poult. Sci. 2004, 83, 193–199. [Google Scholar] [CrossRef]
- Squire, M.E.; Veglia, M.K.; Drucker, K.A.; Brazeal, K.R.; Hahn, T.P.; Watts, H.E. Estrogen levels influence medullary bone quantity and density in female house finches and pine siskins. Gen. Comp. Endocrinol. 2017, 246, 249–257. [Google Scholar] [CrossRef]
- Neijat, M.; Casey-Trott, T.M.; Robinson, S.; Widowski, T.M.; Kiarie, E. Effects of rearing and adult laying housing systems on medullary, pneumatic and radius bone attributes in 73-wk old Lohmann LSL lite hens1. Poult. Sci. 2019, 98, 2840–2845. [Google Scholar] [CrossRef] [PubMed]
- Reich, T.; Gefen, A. Effect of trabecular bone loss on cortical strain rate during impact in an in vitro model of avian femur. Biomed. Eng. Online 2006, 5, 45. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Casey-Trott, T.M.; Korver, D.R.; Guerin, M.T.; Sandilands, V.; Torrey, S.; Widowski, T.M. Opportunities for exercise during pullet rearing, Part II: Long-term effects on bone characteristics of adult laying hens at the end-of-lay. Poult. Sci. 2017, 96, 2518–2527. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, J.; Chen, J.; Shi, H.; Liao, X.; Pan, C.; Liu, Y.; Yang, X.; Ren, Z.; Yang, X. Adjusting phosphate feeding regimen according to daily rhythm increases eggshell quality via enhancing medullary bone remodeling in laying hens. J. Anim. Sci. Biotechnol. 2023, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Kerschnitzki, M.; Zander, T.; Zaslansky, P.; Fratzl, P.; Shahar, R.; Wagermaier, W. Rapid alterations of avian medullary bone material during the daily egg-laying cycle. Bone 2014, 69, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Cransberg, P.H.; Parkinson, G.B.; Wilson, S.; Thorp, B.H. Sequential studies of skeletal calcium reserves and structural bone volume in a commercial layer flock. Br. Poult. Sci. 2001, 42, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Casey-Trott, T.M.; Korver, D.R.; Guerin, M.T.; Sandilands, V.; Torrey, S.; Widowski, T.M. Opportunities for exercise during pullet rearing, Part I: Effect on the musculoskeletal characteristics of pullets. Poult. Sci. 2017, 96, 2509–2517. [Google Scholar] [CrossRef] [PubMed]
- Fleming, R.H. Nutritional factors affecting poultry bone health. Proc. Nutr. Soc. 2008, 67, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Gautron, J.; Réhault-Godbert, S.; Van de Braak, T.G.H.; Dunn, I.C. Review: What are the challenges facing the table egg industry in the next decades and what can be done to address them? Anim. Int. J. Anim. Biosci. 2021, 15 (Suppl. 1), 100282. [Google Scholar] [CrossRef]
- Papadopoulou, S.K.; Papadimitriou, K.; Voulgaridou, G.; Georgaki, E.; Tsotidou, E.; Zantidou, O.; Papandreou, D. Exercise and Nutrition Impact on Osteoporosis and Sarcopenia-The Incidence of Osteosarcopenia: A Narrative Review. Nutrients. 2021, 13, 4499. [Google Scholar] [CrossRef]
- Li, J.Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex steroid deficiency-associated bone loss is microbiota dependent and prevented by probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.C.; Bilezikian, J.P. Parathyroid hormone: Anabolic and catabolic actions on the skeleton. Curr. Opin. Pharmacol. 2015, 22, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Nakashima, T.; Shinohara, M.; Negishi-Koga, T.; Komatsu, N.; Terashima, A.; Sawa, S.; Nitta, T.; Takayanagi, H. Osteoimmunology: The Conceptual Framework Unifying the Immune and Skeletal Systems. Physiol. Rev. 2017, 97, 1295–1349. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, M.M.; Jones, R.M.; Schett, G.; Pacifici, R. The gut-bone axis: How bacterial metabolites bridge the distance. J. Clin. Investig. 2019, 129, 3018–3028. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Hu, Y.; Guo, Y.; Liu, D. Modulation of bone remodeling by the gut microbiota: A new therapy for osteoporosis. Bone. Res. 2023, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R. Probiotic L. reuteri treatment prevents bone loss in a menopausal ovariectomized mouse model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Parvaneh, K.; Ebrahimi, M.; Sabran, M.R.; Karimi, G.; Hwei, A.N.; Abdul-Majeed, S.; Ahmad, Z.; Ibrahim, Z.; Jamaluddin, R. Probiotics (Bifidobacterium longum) Increase Bone Mass Density and Upregulate Sparc and Bmp-2 Genes in Rats with Bone Loss Resulting from Ovariectomy. Biomed. Res. Int. 2015, 2015, 897639. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.L.; Liu, G.; Darby, T.M.; Fernandes, L.M.; Diaz-Hernandez, M.E.; Jones, R.M.; Drissi, H. Bifidobacterium adolescentis supplementation attenuates fracture-induced systemic sequelae. Biomed. Pharmacother. 2020, 132, 110831. [Google Scholar] [CrossRef]
- Liu, J.H.; Chen, C.Y.; Liu, Z.Z.; Luo, Z.W.; Rao, S.S.; Jin, L.; Wan, T.F.; Yue, T.; Tan, Y.J.; Yin, H.; et al. Extracellular Vesicles from Child Gut Microbiota Enter into Bone to Preserve Bone Mass and Strength. Adv. Sci. 2021, 8, 2004831. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Naito, Y.; Uchiyama, K.; Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr. 2018, 63, 33–35. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Everard, A.; Gómez-Valadés, A.G.; Matamoros, S.; Ramírez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Feng, S.; Arjan, N.; Chen, W. A next generation probiotic, Akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 2019, 59, 3227–3236. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Mulhall, H.; DiChiara, J.M.; Deragon, M.; Iyer, R.; Huck, O.; Amar, S. Akkermansia muciniphila and Its Pili-Like Protein Amuc_1100 Modulate Macrophage Polarization in Experimental Periodontitis. Infect. Immun. 2020, 89, e00500-20. [Google Scholar] [CrossRef]
- Huck, O.; Mulhall, H.; Rubin, G.; Kizelnik, Z.; Iyer, R.; Perpich, J.D.; Haque, N.; Cani, P.D.; de Vos, W.M.; Amar, S. Akkermansia muciniphila reduces Porphyromonas gingivalis-induced inflammation and periodontal bone destruction. J. Clin. Periodontol. 2020, 47, 202–212. [Google Scholar] [CrossRef]
- Liu, J.H.; Yue, T.; Luo, Z.W.; Cao, J.; Yan, Z.Q.; Jin, L.; Wan, T.F.; Shuai, C.J.; Wang, Z.G.; Zhou, Y.; et al. Akkermansia muciniphila promotes type H vessel formation and bone fracture healing by reducing gut permeability and inflammation. Dis. Model. Mech. 2020, 13, dmm043620. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.K.; Liu, G.; White, D.L.; Tompkins, Y.H.; Kim, W.K. Graded levels of Eimeria challenge altered the microstructural architecture and reduced the cortical bone growth of femur of Hy-Line W-36 pullets at early stage of growth (0–6 wk of age). Poult. Sci. 2023, 102, 102888. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. The gut microbiota regulates bone mass in mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Yang, X.; Zhang, M.; Xu, C.; Hu, Y.; Liu, D. Akkermansia muciniphila Enhances Egg Quality and the Lipid Profile of Egg Yolk by Improving Lipid Metabolism. Front. Microbiol. 2022, 13, 927245. [Google Scholar] [CrossRef] [PubMed]
- Passi, N.; Gefen, A. Trabecular bone contributes to strength of the proximal femur under mediolateral impact in the avian. J. Biomech. Eng. 2005, 127, 198–203. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, C.; Kim, W.K. The application of micro-CT in egg-laying hen bone analysis: Introducing an automated bone separation algorithm. Poult. Sci. 2020, 99, 5175–5183. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hu, Y.; Wu, Y.; Liu, Y.; Liu, G.; Yan, Z.; Li, Q.; Zhou, Z.; Li, Z. Influences of bioapatite mineral and fibril structure on the mechanical properties of chicken bone during the laying period. Poult. Sci. 2019, 98, 6393–6399. [Google Scholar] [CrossRef] [PubMed]
- Schepper, J.D.; Collins, F.L.; Rios-Arce, N.D.; Raehtz, S.; Schaefer, L.; Gardinier, J.D.; Britton, R.A.; Parameswaran, N.; McCabe, L.R. Probiotic Lactobacillus reuteri Prevents Postantibiotic Bone Loss by Reducing Intestinal Dysbiosis and Preventing Barrier Disruption. J. Bone Miner. Res. 2019, 34, 681–698. [Google Scholar] [CrossRef]
- Tompkins, Y.H.; Choi, J.; Teng, P.Y.; Yamada, M.; Sugiyama, T.; Kim, W.K. Reduced bone formation and increased bone resorption drive bone loss in Eimeria infected broilers. Sci. Rep. 2023, 13, 616. [Google Scholar] [CrossRef]
- Lv, Z.; Fan, H.; Gao, M.; Zhang, X.; Li, G.; Fan, Y.; Ning, Z.; Guo, Y. The accessible chromatin landscape of lipopolysaccharide-induced systemic inflammatory response identifying epigenome signatures and transcription regulatory networks in chickens. Int. J. Biol. Macromol. 2024, 266, 131136. [Google Scholar] [CrossRef]
- Peek, C.T.; Ford, C.A.; Eichelberger, K.R.; Jacobse, J.; Torres, T.P.; Maseda, D.; Latour, Y.L.; Piazuelo, M.B.; Johnson, J.R.; Byndloss, M.X.; et al. Intestinal Inflammation Promotes MDL-1(+) Osteoclast Precursor Expansion to Trigger Osteoclastogenesis and Bone Loss. Cell. Mol. Gastroenterol. Hepatol. 2022, 14, 731–750. [Google Scholar] [CrossRef] [PubMed]
- Arron, J.R.; Choi, Y. Bone versus immune system. Nature 2000, 408, 535–536. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.K.; Regmi, P.; Applegate, T.; Chai, L.; Kim, W.K. Osteoimmunology: A Link between Gastrointestinal Diseases and Skeletal Health in Chickens. Animals 2023, 13, 1816. [Google Scholar] [CrossRef] [PubMed]
- Ciucci, T.; Ibáñez, L.; Boucoiran, A.; Birgy-Barelli, E.; Pène, J.; Abou-Ezzi, G.; Arab, N.; Rouleau, M.; Hébuterne, X.; Yssel, H.; et al. Bone marrow Th17 TNFα cells induce osteoclast differentiation, and link bone destruction to IBD. Gut 2015, 64, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.M.; Yu, M.; Darby, T.M.; Vaccaro, C.; Li, J.Y.; Owens, J.A.; Hsu, E.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; et al. The Microbial Metabolite Butyrate Stimulates Bone Formation via T Regulatory Cell-Mediated Regulation of WNT10B Expression. Immunity 2018, 49, 1116–1131.e1117. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Ventura, M.; Buttó, L.F.; Duranti, S.; O’Toole, P.W.; Motherway, M.O.; van Sinderen, D. Molecular dialogue between the human gut microbiota and the host: A Lactobacillus and Bifidobacterium perspective. Cell. Mol. Life Sci. 2014, 71, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.S.; Pan, T.M. Antiosteoporotic effects of Lactobacillus -fermented soy skim milk on bone mineral density and the microstructure of femoral bone in ovariectomized mice. J. Agric. Food Chem. 2011, 59, 7734–7742. [Google Scholar] [CrossRef] [PubMed]
- Ong, A.M.; Kang, K.; Weiler, H.A.; Morin, S.N. Fermented Milk Products and Bone Health in Postmenopausal Women: A Systematic Review of Randomized Controlled Trials, Prospective Cohorts, and Case-Control Studies. Adv. Nutr. 2020, 11, 251–265. [Google Scholar] [CrossRef]
- Tu, M.Y.; Han, K.Y.; Chang, G.R.; Lai, G.D.; Chang, K.Y.; Chen, C.F.; Lai, J.C.; Lai, C.Y.; Chen, H.L.; Chen, C.M. Kefir Peptides Prevent Estrogen Deficiency-Induced Bone Loss and Modulate the Structure of the Gut Microbiota in Ovariectomized Mice. Nutrients 2020, 12, 3432. [Google Scholar] [CrossRef]
- Lee, C.S.; Kim, J.Y.; Kim, B.K.; Lee, I.O.; Park, N.H.; Kim, S.H. Lactobacillus-fermented milk products attenuate bone loss in an experimental rat model of ovariectomy-induced post-menopausal primary osteoporosis. J. Appl. Microbiol. 2021, 130, 2041–2062. [Google Scholar] [CrossRef]
- Tu, M.Y.; Chen, H.L.; Tung, Y.T.; Kao, C.C.; Hu, F.C.; Chen, C.M. Short-Term Effects of Kefir-Fermented Milk Consumption on Bone Mineral Density and Bone Metabolism in a Randomized Clinical Trial of Osteoporotic Patients. PLoS ONE 2015, 10, e0144231. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.G.; Sundh, D.; Bäckhed, F.; Lorentzon, M. Lactobacillus reuteri reduces bone loss in older women with low bone mineral density: A randomized, placebo-controlled, double-blind, clinical trial. J. Intern. Med. 2018, 284, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, C.; Engdahl, C.; Fåk, F.; Andersson, A.; Windahl, S.H.; Farman, H.H.; Movérare-Skrtic, S.; Islander, U.; Sjögren, K. Probiotics protect mice from ovariectomy-induced cortical bone loss. PLoS ONE 2014, 9, e92368. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Cui, L.Y.; Hou, J.F.; Shi, C.; Ke, X.; Yang, L.C.; Ma, X.P. Effects of age and dietary soybean oil level on eggshell quality, bone strength and blood biochemistry in laying hens. Br. Poult. Sci. 2014, 55, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Gervais, O.; Nirasawa, K.; Vincenot, C.E.; Nagamine, Y.; Moriya, K. Effect of Long-Term Selection for Non-Destructive Deformation on Egg Shape in White Leghorns. J. Poult. Sci. 2016, 53, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Sirri, F.; Zampiga, M.; Berardinelli, A.; Meluzzi, A. Variability and interaction of some egg physical and eggshell quality attributes during the entire laying hen cycle. Poult. Sci. 2018, 97, 1818–1823. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content, % | ||||
|---|---|---|---|---|---|
| Week | 1–2 | 3–6 | 7–10 | 11–14 | 15–20 |
| Corn (CP 7.8%) | 60.307 | 63.893 | 66.446 | 67.878 | 66.370 |
| Soybean meal (CP 44%) | 34.104 | 31.386 | 26.298 | 18.179 | 22.543 |
| Soybean oil | 1.200 | 0.339 | 0.000 | 0.000 | 0.000 |
| Corn gluten meal (60%) | 0.000 | 0.000 | 0.000 | 1.606 | 1.393 |
| Wheat bran (15.7%) | 0.000 | 0.000 | 2.931 | 7.575 | 1.475 |
| Calcium hydrogen phosphate | 1.900 | 2.131 | 2.180 | 2.096 | 1.906 |
| Limestone | 1.300 | 1.165 | 1.182 | 1.865 | 5.150 |
| Sodium chloride | 0.389 | 0.388 | 0.256 | 0.218 | 0.352 |
| Mineral premix a | 0.300 | 0.300 | 0.300 | 0.300 | 0.300 |
| DL-Methionine, 98% | 0.220 | 0.219 | 0.179 | 0.103 | 0.228 |
| L-Lysine sulphate, 78% | 0.100 | 0.000 | 0.047 | 0.000 | 0.104 |
| Choline chloride, 50% | 0.100 | 0.100 | 0.100 | 0.100 | 0.100 |
| Vitamin premix b | 0.040 | 0.040 | 0.040 | 0.040 | 0.040 |
| Ethoxyquinoline, 33% | 0.020 | 0.020 | 0.020 | 0.020 | 0.020 |
| Phytase 10,000 c | 0.020 | 0.020 | 0.020 | 0.020 | 0.020 |
| Total | 100 | 100 | 100 | 100 | 100 |
| Nutrient levels | |||||
| Metabolizable energy, Mcal/kg | 2.887 | 2.860 | 2.830 | 2.800 | 2.750 |
| Lysine, % | 1.236 | 1.086 | 1.000 | 0.776 | 0.840 |
| Methionine, % | 0.541 | 0.529 | 0.470 | 0.384 | 0.490 |
| Methionine + cystine, % | 0.877 | 0.855 | 0.780 | 0.686 | 0.760 |
| Crude protein, % | 20.000 | 19.000 | 17.500 | 15.800 | 16.200 |
| Nonphytate phosphorus, % | 0.405 | 0.440 | 0.450 | 0.440 | 0.500 |
| Calcium, % | 1.010 | 1.000 | 1.000 | 1.200 | 2.500 |
| Phosphorus, % | 0.671 | 0.699 | 0.703 | 0.678 | 0.770 |
| Abbreviation | Description of Variables | Unit |
|---|---|---|
| BV | Bone volume of the bone segment | mm3 |
| TV | Total volume of the entire region of interest | mm3 |
| BV/TV | The ratio of bone volume to total volume | % |
| BS | Bone surface area of the bone segment | mm2 |
| BS/TV | The ratio of bone surface area to total volume | % |
| BMC | Bone mineral content | mg |
| BMD | Bone mineral density | g/cm3 |
| Th | Thickness of the microarchitecture | mm |
| Tb. Sp | Mean distance between trabeculae | mm |
| Tb. N | Trabecular bone number | mm−1 |
| BM. CV/TV | The volume of the medullary cavity versus total femur volume ratio | % |
| Items | Control | Akk | SEM | p Value |
|---|---|---|---|---|
| 0–6 weeks, g | 19.95 | 19.42 | 0.35 | 0.15 |
| 0–12 weeks, g | 36.23 | 35.55 | 0.32 | 0.05 |
| 0–18 weeks, g | 45.85 | 45.00 | 0.55 | 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, Z.; Zhang, Y.; Yuan, G.; Zhang, F.; Hu, Y.; Liu, D. Akkermansia muciniphila Promotes Bone Development and Improves Eggshell Quality during the Sexual Maturity Period of Laying Hens by Increasing Osteogenesis. Agriculture 2024, 14, 598. https://doi.org/10.3390/agriculture14040598
Lyu Z, Zhang Y, Yuan G, Zhang F, Hu Y, Liu D. Akkermansia muciniphila Promotes Bone Development and Improves Eggshell Quality during the Sexual Maturity Period of Laying Hens by Increasing Osteogenesis. Agriculture. 2024; 14(4):598. https://doi.org/10.3390/agriculture14040598
Chicago/Turabian StyleLyu, Zhengtian, Yuying Zhang, Gaoxiang Yuan, Fengwenhui Zhang, Yongfei Hu, and Dan Liu. 2024. "Akkermansia muciniphila Promotes Bone Development and Improves Eggshell Quality during the Sexual Maturity Period of Laying Hens by Increasing Osteogenesis" Agriculture 14, no. 4: 598. https://doi.org/10.3390/agriculture14040598
APA StyleLyu, Z., Zhang, Y., Yuan, G., Zhang, F., Hu, Y., & Liu, D. (2024). Akkermansia muciniphila Promotes Bone Development and Improves Eggshell Quality during the Sexual Maturity Period of Laying Hens by Increasing Osteogenesis. Agriculture, 14(4), 598. https://doi.org/10.3390/agriculture14040598