Abstract
Pequi oil (PO) is a natural feed additive rich in bioactive compounds, which can modulate antioxidant and immunological systems. Thus, the aim of this study was to evaluate the proteomic profile of laying hens supplemented with PO under heat stress conditions. Ninety-six 26-week-old Lohmann White hens were housed in a completely randomized design with a 2 × 2 factorial arrangement, with two climate chambers (cyclic heat stress and thermoneutral) and two diets (control and 0.6% PO). At 38 weeks old, liver samples were collected for protein extraction and digestion, and were submitted to liquid chromatography–tandem mass spectrometry (LC-MS/MS). A total of 996 differentially expressed proteins were identified in the liver proteome of laying hens fed with 0.6% PO under heat stress. These upregulated proteins (0.95 ≤ p ≤ 1.00) are associated with lipid metabolism (apolipoprotein B; vitellogenin-1; ovotransferrin), the antioxidant system (protein disulfide-isomerase A4; superoxide dismutase 1_ soluble; catalase), the immune system (Ig-like domain-containing protein) and chaperones (HSP 90; HSP 70). PO positively modulates a network of heat shock proteins and antioxidant enzymes, and the unique proteins identified can contribute to the discovery of new biomarkers related to heat stress reduction by phytogenic additives.
1. Introduction
The increase in temperature due to global warming is concerning because it leads to heat stress (HS), a condition characterized by an imbalance between the heat released by an animal into the environment and the internal heat generated by the animal’s metabolism [1]. When laying hens experience HS, they alter their behavior and physiological homeostasis in an attempt to return to their optimal thermal comfort. This results in lower egg weight due to reduced feed intake and nutrient absorption. Additionally, the acid–base imbalance caused by respiratory alkalosis reduces the availability of calcium (Ca), leading to poor eggshell mineralization in laying hens [2].
At the molecular level, HS is a factor that triggers the overproduction of reactive oxygen species. When this occurs, it overwhelms the neutralizing capacity of the antioxidant system, leading to oxidative stress. Oxidative stress initially promotes lipid peroxidation and damages unsaturated fatty acids in the cell membrane, as well as proteins and DNA, resulting in impaired cell and membrane integrity. To prevent such protein damage, the synthesis of heat shock proteins (HSPs) occurs, representing the last line of antioxidant defense. These HSPs assist in the formation, folding, and renaturation of misfolded proteins, prevent protein aggregation, and degrade unstable proteins [3]. Therefore, natural feed additives with antioxidant and HSP activation properties are considered the most important protective agents against stress in poultry [4].
The pequi (Caryocar Brasiliense Camb.) is a fruit found in the Brazilian Cerrado region, and its pulp can be used to extract an oil rich in polyunsaturated fatty acids. Pequi oil (PO) also contains carotenoids (β-carotene, lycopene, and lutein), vitamins (A, C, and E), and polyphenols, which are natural compounds with antioxidant and anti-inflammatory properties. These bioactive compounds and biomolecules can upregulate HSPs and enhance antioxidant molecule activity, which enables an improved response to heat stress. Thus, PO addition to poultry feed has the potential to strengthen antioxidant defenses and regulate HSP expression, preventing damage to vital biomolecules, when exposed to heat stress conditions [5,6].
Furthermore, a shotgun proteomic approach may facilitate a better understanding of protein function at the molecular level, making it a key tool for identifying proteins directly involved in heat stress, altering protein regulation, and discovering new biomarkers [7,8]. In this way, this study aimed to evaluate the abundance of proteins differentially expressed in the liver tissue of laying hens fed pequi oil under thermoneutral and cyclic heat stress conditions, using a proteomic approach. These results may contribute to a deeper understanding of the molecular mechanisms underlying the physiological response of laying hens to pequi oil.
2. Materials and Methods
The experiment was conducted from October 2022 to January 2023 at São Paulo State University (UNESP), School of Veterinary Medicine and Animal Science (FMVZ), Botucatu, in the Laboratory of Poultry Nutrition. All procedures performed in the present study were approved by the Ethics Committee on Animal Use from the same institution, under protocol number 221/2022. The hens were vaccinated upon hatching against Marek’s disease, avian poxvirus, and infectious bursal disease. The vaccination program followed up to the 26th week of age was adapted from regional practices, including ocular vaccine administration against infectious bursal disease (Poulvac® Bursa F, Zoetis, Parsippany–Troy Hills, NJ, USA), infectious bronchitis (CEVAC BR 120L, CEVA, Libourne, France), and Newcastle disease (CEVAC BRON+HB1, CEVA). The avian poxvirus (VECT FP MG AE, CEVA) vaccine was administered in the wing membrane, while the vaccines for Egg Drop Syndrome and infectious coryza (Poulvac® Mix6, Zoetis, Parsippany–Troy Hills, NJ, USA) were administered intramuscularly using an oily solution.
2.1. Experimental Design, Diets, and Treatments
Ninety-six 26-week-old Lohmann White hens with similar body weights (1544.58 g) and laying rates (98.44%) were housed in a climatic chamber equipped with 24 galvanized wire cages at a 1.500 cm2/hen density (0.5 m height × 0.6 m width × 1.0 m length), with trough-type feeders and nipple-type drinkers, with four birds per cage. The hens were distributed in a completely randomized design with a 2 × 2 factorial arrangement, including two climate chambers and two diets, resulting in four treatments with six replicates.
The hens were fed with a control (CO) diet without pequi oil (PO) supplementation and a diet supplemented with 0.6% PO. All diets were formulated according to the hens’ nutritional requirements, based on the values provided by Rostagno and collaborators [9]. The ingredients and calculated nutritional levels of the diet are presented in Silva et al. [10]. Pequi oil was added to the diet to replace soybean oil, with an adjustment for the apparent metabolizable energy value of 7370 kcal/kg for PO [11]. The hens were provided with water and feed ad libitum. A lighting program of 16 h of daily artificial light was applied according to the breeder’s recommendations. Pequi oil was purchased from Grande Sertão©, a cooperative located in Montes Claros (MG/Brazil), where the oil was extracted from the pulp using manual processes.
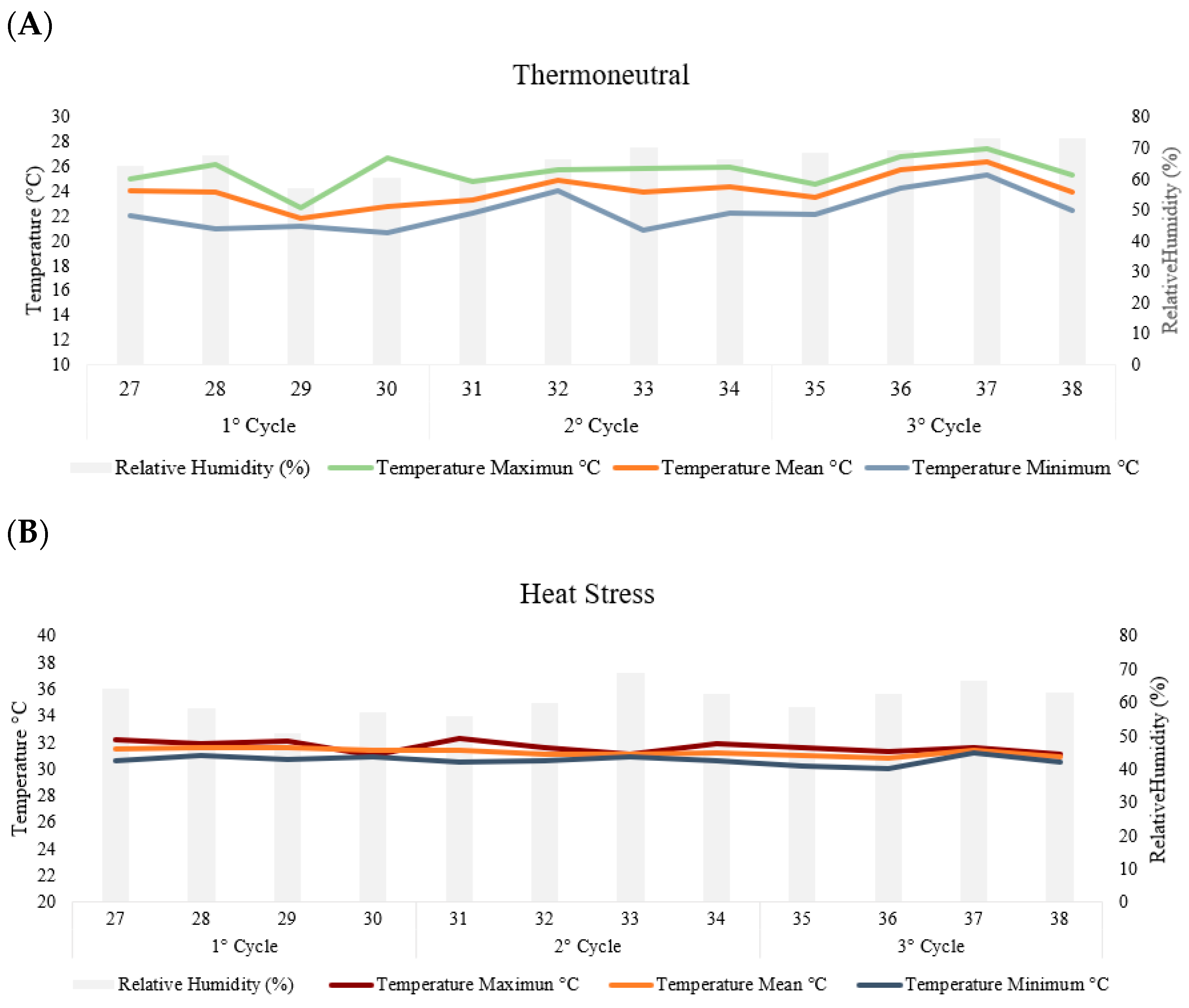
In the thermoneutral chamber, the hens were maintained at a thermal comfort temperature of 24 ± 1 °C and relative humidity of 66 ± 3, while in the cyclic heat stress chamber, the hens were exposed to elevated temperatures 31 ± 1° and relative humidity of 60 ± 2, 8 h daily (from 8:30 a.m. to 4:30 p.m.) (Figure 1). Daily monitoring of air temperature and relative humidity was conducted using a set of maximum and minimum thermometers placed at the height of the hens and using HOBO-type electronic sensors (Onset Data Loggers).
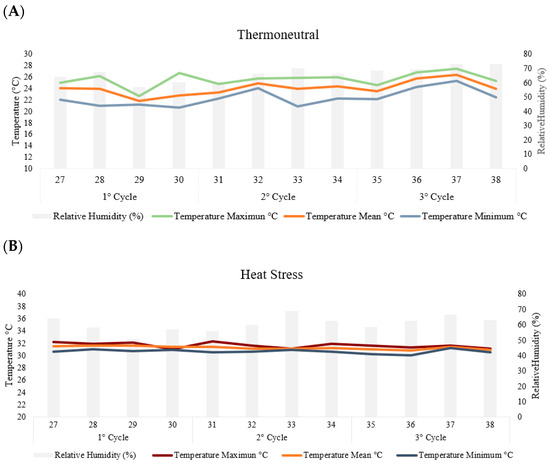
Figure 1.
The temperatures and relative humidity per week in the thermoneutral (A) and heat stress (B) chambers during the three production cycles (84 days).
2.2. Protein Relative Abundance Variance Analysis by Shotgun LC–MS/MS
2.2.1. Liver Tissue Analysis
At the end of the experimental period (84 days), the laying hens were individually identified and fasted for 8 h. One hen from each experimental unit, weighing close to the group’s average weight (thermoneutral: PO 1727 g and CO 1661 g; heat stress: PO 1727 g and CO 1765 g), was sacrificed by cervical dislocation to collect 5 g of the left liver lobe for protein relative abundance variance analysis using shotgun LC-MS/MS in Bioanalytical and Metalloproteomics Laboratory (LBM) at the Department of Chemistry and Biochemistry, IBB-UNESP-Botucatu. Proteomic analysis was conducted considering each liver sample from experimental groups (six per treatment) as biological replicates.
2.2.2. Liver Tissue Protein Preparation for Shotgun LC–MS/MS Analysis
To obtain the proteome of the studied species, the shotgun LC-MS/MS strategy was applied to liver tissue samples from the CO and PO groups. Approximately 150 mg of liver tissue was weighed, and 200 µL of extraction solution (7 mol/L Urea, 2 mol/L Thiourea, 40 mmol/L Dithiothreitol (DTT) solubilized in 50 mmol/L ammonium bicarbonate (AMBIC)) was added. The samples were processed using an OMNI/Analytic cell disruptor [12]. After centrifugation at 12,000× g for 30 min at 4 °C, the clear protein extracts were collected, and the total protein concentration was determined using the Bradford method [13]. The protein extract concentration was adjusted to 4 µg/µL, ensuring that 100 µL of extract contained 400 µg of protein. In each microtube containing the separate aliquots for each shotgun analysis, 10 µL of 0.05 mol/L ammonium bicarbonate and 25 µL of 0.2% (v/v) RapiGestTM (Waters, UK, USA) were added, and the samples were incubated at 37 °C for 30 min.
Subsequently, the samples underwent reduction and alkylation steps. The reduction was achieved by adding 9.5 µL of 100 mmol/L DTT solution (BIO-RAD, Hercules, CA, USA) to the microtubes, vortexing, and incubating again for 40 min at 37 °C. The alkylation process was conducted by adding 6.3 µL of 300 mmol/L iodoacetamide solution (GE, Boston, MA, USA), homogenizing, and incubating for 30 min at room temperature in the absence of light. For protein cleavage (tryptic digestion), 80 µL of 0.05 µg/µL trypsin (PRO-MEGA, cat #V5280, Madison, WI, USA) was added to each sample, and the samples were incubated for 16 h at 37 °C. At the end of the cleavage process, 10 µL aliquots of 5% (v/v) trifluoroacetic acid (Sigma-Aldrich, LC-MS, St. Louis, MO, USA) were added to stop the trypsin activity. For sample purification and desalting, C18 columns (Agilent, Santa Clara, CA, USA) were used. The solution containing the peptides of interest was concentrated in a vacuum concentrator (Christ RVC 2-18 CDplus, Osterode am Harz, Germany) to a volume of approximately 2 µL. The microtubes containing the peptides were stored in freezers until analysis by LC-MS/MS.
Trypsin was the enzyme used in the cleavage process. Cysteine carbamidomethylation was defined as a fixed modification and methionine oxidation as a variable modification. A precursor mass tolerance of 10 ppm and a fragment ion tolerance of 0.01 Da were applied, and a false discovery rate of 1% was considered.
To carry out the analyses, the samples were resuspended in 200 μL of 3% (v/v) acetonitrile (Sigma-Aldrich—UHPLC, USA) and 3% (v/v) formic acid (Thermo Fisher, Waltham, MA, USA) and subjected to analysis by LC-MS/MS. Peptide identification was performed on a nanoACQUITY UPLC-Xevo QT MS system (Waters, Manchester, UK, USA). The nanoACQUITY UPLC® was equipped with a nanoACQUITY HSS T3 reversed-phase analytical column (75 µm × 150 mm, 1.8 µm particle size). The column was equilibrated with 93% mobile phase A (0.1% (v/v) formic acid in water) and 7% mobile phase B (0.1% (v/v) formic acid in acetonitrile). Then, the peptides were separated with a 7–85% linear mobile phase gradient (0.1% formic acid in acetonitrile) for 70 min at a flow rate of 0.35 μL/min. The column temperature was maintained at 35 °C.
The Xevo® G2 Q-TOF mass spectrometer was operated in positive ion nanoelectrospray mode, and the data were collected using the MSE method at high energy (19–45 V), which allows the acquisition of precursor ion data and fragments in a single injection. The source conditions included a capillary voltage of 2.5 kV, a sample cone voltage of 30.0 V, an extraction cone voltage of 5.0 V, and a temperature of 80 °C. Data were acquired over 70 min, scanning in the range of 50 to 2000 Da. To ensure the accuracy and reproducibility of the analyses, a standard fibrinopeptide solution, 1 pmol/μL (Glu1), was used between sample injections.
The raw mass spectrometry data files were loaded into Progenesis QI for Proteomics v4.0 (Nonlinear Dynamics, Waters, Newcastle upon Tyne, UK) to perform quantitative analysis. For alignment, a sample result was automatically selected as a reference. The remaining results were aligned with the reference with an alignment score greater than 80%, and peak collection was performed using the automatic sensitivity method. Peak intensities were then normalized by calculating the abundance ratios relative to a reference. The peak list was searched in the UniProtKB/Swiss-Prot database using Gallus gallus species data and Mascot 2.6 software (Matrix Science, London, UK).
In the expression analyses, the sample size was six hens per treatment, and p < 0.05 was considered significant for negatively regulated proteins and p > 0.95 for positively regulated proteins. After compiling the results, functional analysis was performed using String 12.0 software (EMBL, BW, DE) to determine the categories of functional proteins based on gene ontology annotations of biological processes, molecular function, and cellular component [14].
3. Results
The mass spectrometry results were imported into the database, identifying a total of 996 proteins (Supplementary Table S1). The proteins identified in the livers of laying hens fed with pequi oil (PO) and the control (CO) under heat stress were compared with those of laying hens fed with PO and CO at thermoneutral temperatures. Among the identified proteins, 146 were upregulated and 10 were downregulated in the PO group, while 12 were upregulated and 70 were downregulated in the CO group. These proteins were classified and discussed according to the research aims and are presented in Table 1. Additionally, it was possible to identify proteins uniquely regulated in the PO and CO groups, with 239 and 240 identified at thermoneutral temperature, and 62 and 217 identified under heat stress conditions, respectively. Of these proteins, 15 were selected based on research interest and are presented in Table 2.

Table 1.
Upregulated and downregulated proteins in the hepatic proteome of laying hens fed the diet supplemented with 0.6% pequi oil (PO) under heat stress compared with the group fed the control diet.
Table 2.
Proteins expressed uniquely in the pequi oil group under heat stress.
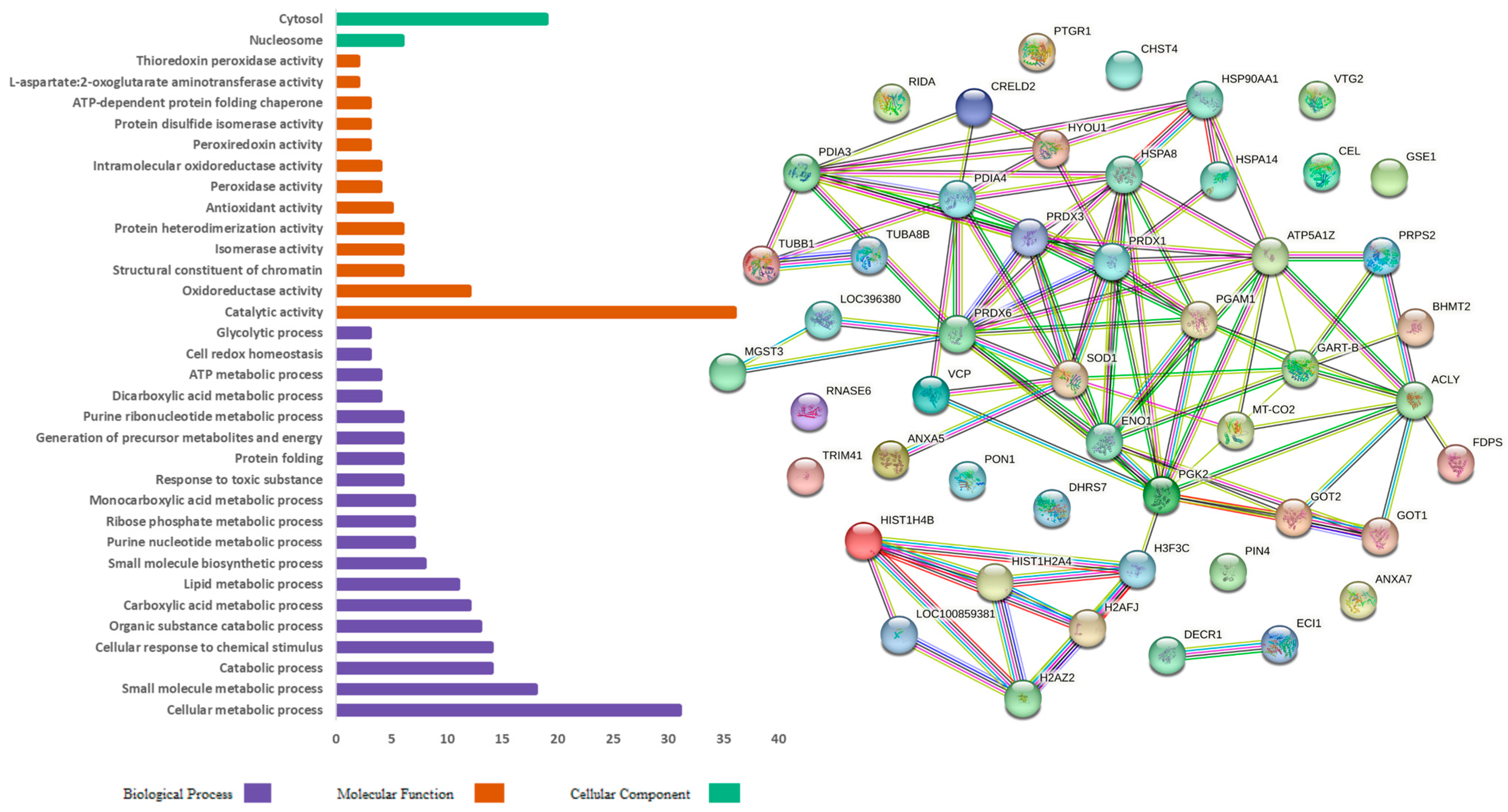
The gene ontology analysis classified the protein sequences into biological process (BP), molecular function (MF), and cellular component (CC). Figure 2 shows the gene ontology functional analysis of the proteins identified in the CO diet group. The identified proteins from the BP category were mainly involved in cellular metabolic processes (31), small-molecule metabolic processes (18), catabolic processes (14), and cellular responses to chemical stimuli (14). In the MF category, the proteins were primarily involved in catalytic activity (36), in oxidoreductase activity (12), as structural constituents of chromatin (6), and in isomerase activity (6). The CC category proteins were mainly involved in the cytosol (19) and nucleosome (6).
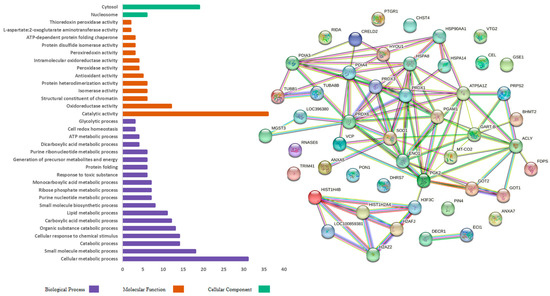
Figure 2.
Gene ontology functional analysis and protein interaction in the control group characterized in the hepatic proteome of laying hens.
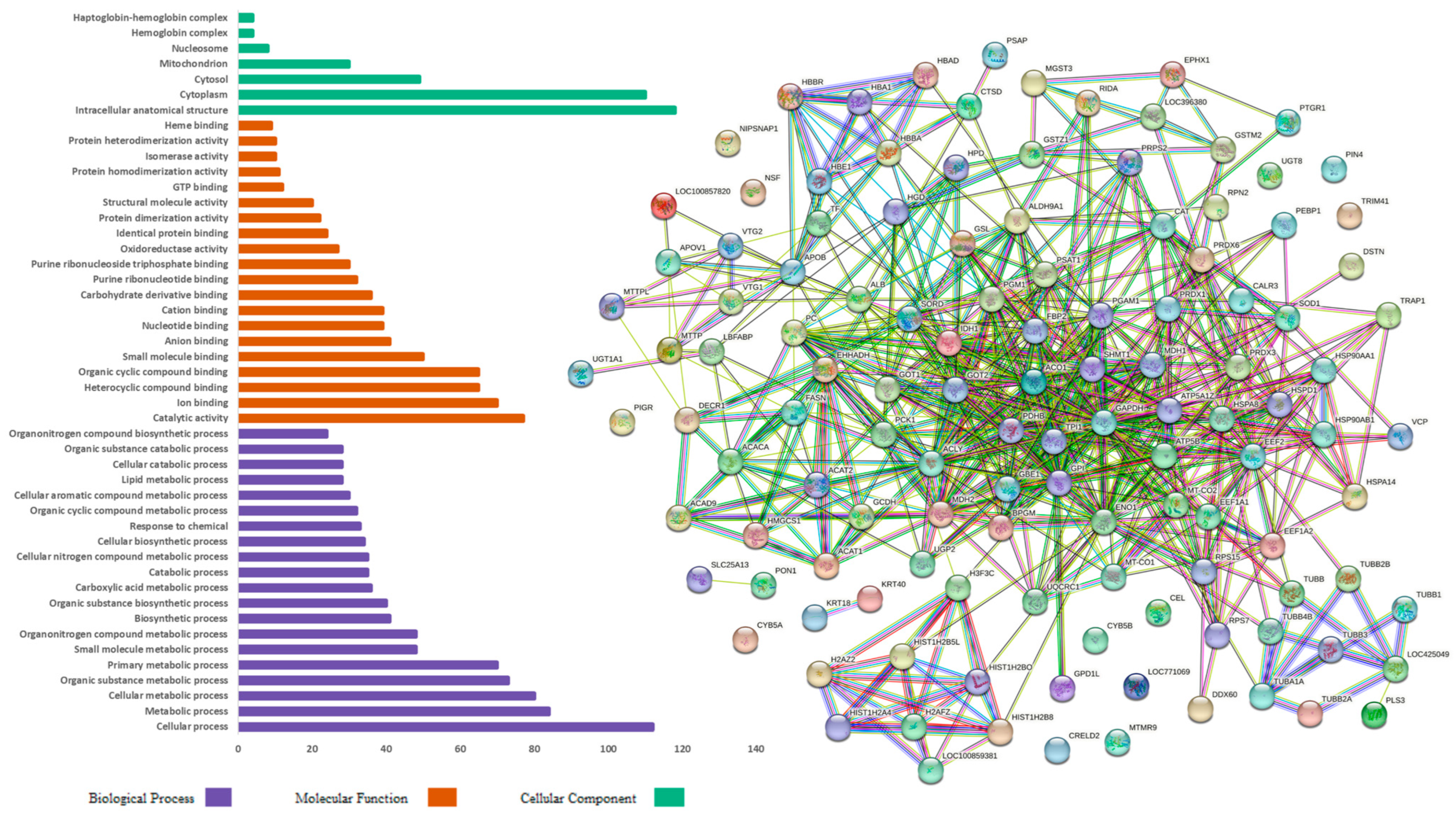
Figure 3 demonstrates the gene ontology functional analysis for the proteins identified in the hens fed the PO diet. The BP category proteins were mainly involved in metabolic processes (65), cellular metabolic processes (62), organic substance metabolic processes (60), and primary metabolic processes (56). In the MF category, the proteins were mainly involved in catalytic activity (64), ion binding (56), the binding of heterocyclic compounds (45), and the binding of organic cyclic compounds (45). The CC category proteins were primarily involved in intracellular anatomical structure (89), cytoplasm (86), cytosol (38), and extracellular space (17).
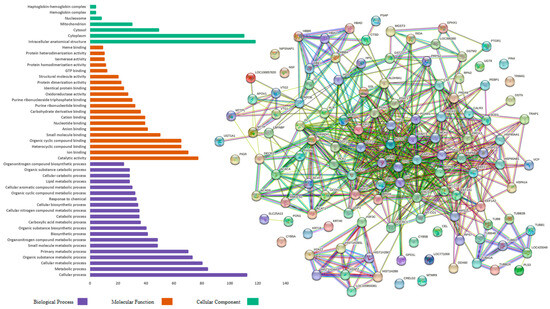
Figure 3.
Gene ontology functional analysis and protein interaction in the pequi oil group characterized in the hepatic proteome of laying hens.
Figure 2 and Figure 3 also present the protein–protein interaction networks, which were assembled using the String 12.0 software (EMBL, BW, DE) and the Gallus gallus genome, showing the upregulated and downregulated proteins identified in the hepatic proteome of laying hens fed CO and PO diets, respectively. In these interaction networks, each circle represents a protein, and the lines represent the interactions between them; the thicker the line, the greater the interaction between proteins.
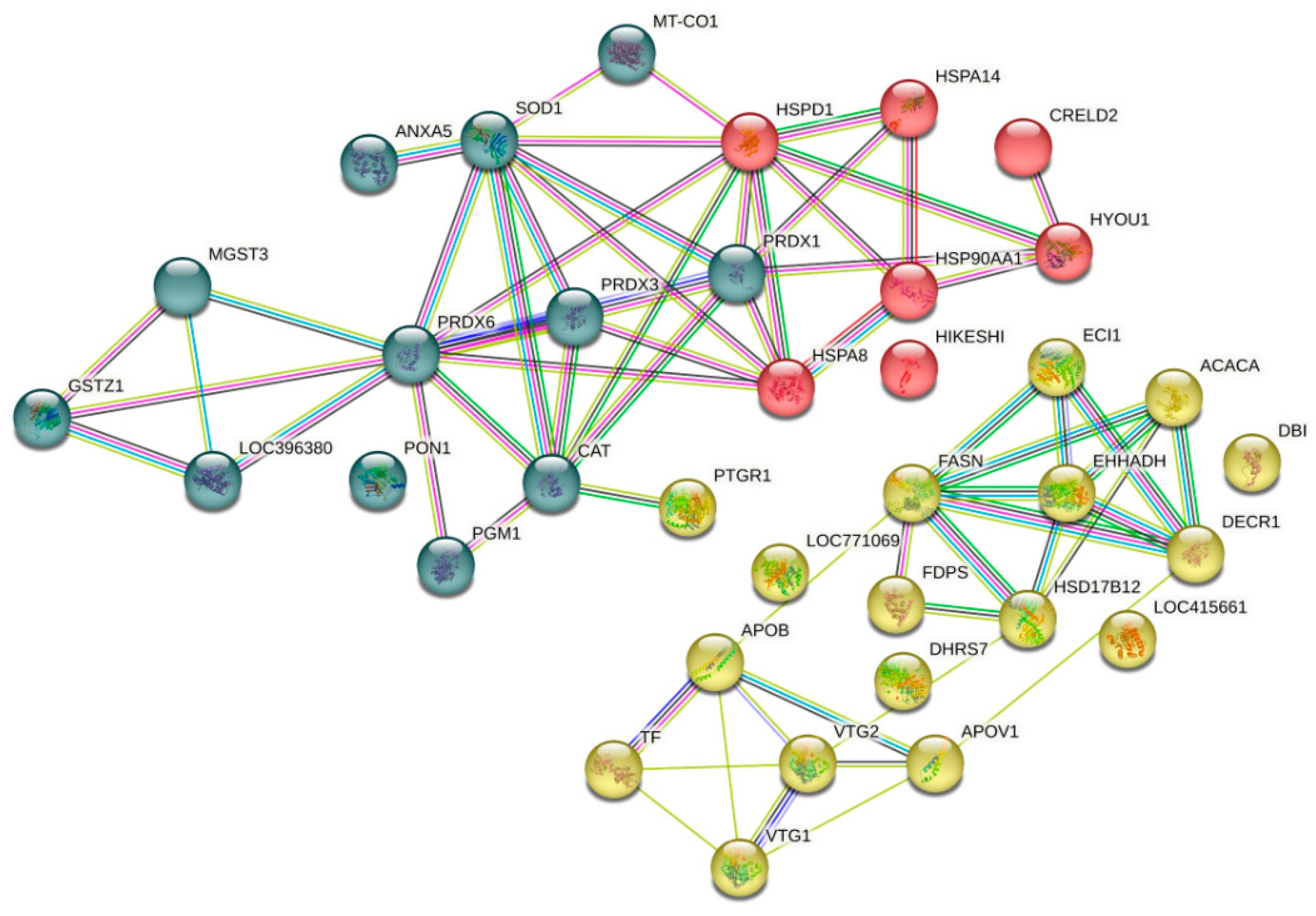
The String 12.0 software (EMBL, BW, DE) and the Gallus gallus genome were used to construct the protein–protein interaction network of the liver of laying hens fed the PO diet and indicate how they contribute to a shared function (Figure 4). The network was grouped into three clusters, highlighting the interactions between proteins involved in antioxidant activity (blue cluster), heat shock proteins (red cluster), and lipid metabolism (yellow cluster).
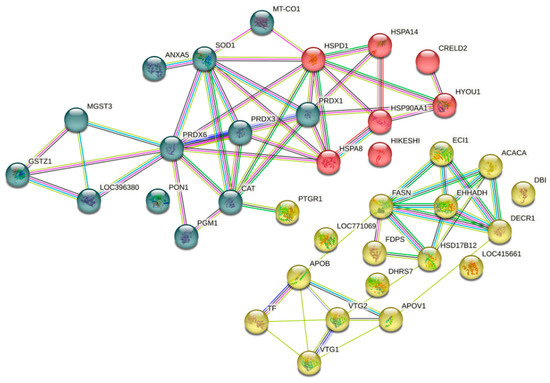
Figure 4.
Protein–protein interaction network clustered from the liver proteome of laying hens fed pequi oil under heat stress.
4. Discussion
In poultry, several nutritional strategies are already used to mitigate the effects of heat stress, including the supplementation of vitamins [15,16], carotenoids [17], phenolic compounds [18,19], probiotics [20,21] and essential oil [22]. However, the use of pequi oil (PO) in the nutrition of laying hens under heat stress, particularly its effect on the regulation of proteins in the hepatic proteome, is underexplored. Therefore, the present study included 0.6% PO in the diet of laying hens subjected to heat stress and observed an increase in the abundance of upregulated proteins linked to the antioxidant system, immune system, and lipid metabolism.
4.1. Heat Shock Proteins
Oxidative stress can lead to protein oxidation, resulting in their aggregation and loss of function. If this condition persists, protein peroxides can attack other proteins [23]. In response to this process, heat shock proteins (HSPs) are expressed to repair the affected proteins [1]. The upregulation of HSPs may be related to the enhanced tolerance of hens to heat stress by improving adaptive responses and increasing proteostasis. HSP90 promotes conformational changes in oxidized proteins related to important regulatory processes, preventing protein aggregation and non-specific interactions. HSP70 correctly folds newly synthesized polypeptides, degrades unstable proteins, and translocates precursor proteins to mitochondria [24,25]. Both groups of chaperones were upregulated in the PO group compared to the control group, suggesting that the use of this oil helped to maintain cellular metabolism and the structural integrity of proteins—reducing the effects of oxidative stress in laying hens—and corroborating with the results of Tu et al. [26], who noted upregulation of HSP90AA1 in the hypothalamus of laying hens under heat stress. Additionally, in fish, the use of functional additives that positively regulate HSPs is already considered a nutritional strategy to enhance stress tolerance [27]. This effect has been observed in recent studies, such as the inclusion of thymol and yeast in rainbow trout (Oncorhynchus mykiss) [28], rosemary oil in thinlip mullet (Chelon ramada) [29], and the use of Bacillus mojavensis B191 and Bacillus subtilis MRS11 in Nile tilapia (Oreochromis niloticus) [30]. In the present study, this response can be attributed to the carotenoids present in the composition of PO, mainly β-carotene, a pigment involved in the stimulation of chaperones such as HSP70 and HSP90, which may have contributed to the increased expression of HSPs [31,32].
Furthermore, chaperones can work together to maximize their action. The binding immunoglobulin protein (BiP) chaperone can bind to HSP90 or HSP70 and act as a cochaperone. This association enhances conformational changes in proteins and improves the capture of proteins that require repair. In the present study, the PO group showed an increase in the expression of this cochaperone in hens under heat stress, promoting greater action by HSPs [33]. Both the control and PO groups positively regulated the expression of protein disulfide-isomerase (PDI), which has chaperone activity. These findings indicate that the protective network against oxidative stress is more responsive to an oxidative challenge, as PDI contributes to protein repair through synthesis, folding, and degradation via its oxidoreductase activity [34].
Another protein with chaperone activity that showed increased expression in the PO group was peptidyl-prolyl cis/trans isomerase (PPIase). PPIase catalyzes the cis/trans isomerization of the peptidyl-prolyl peptide bond in proteins, accelerating the refolding process. This action is essential because some proteins, after undergoing damage, are unable to quickly refold and resume their function [35]. Therefore, the action of PPIase is crucial for proteins to return to their normal functioning.
4.2. Antioxidant System
Endogenous antioxidant enzymes combat free radicals. Among them, superoxide dismutase (SOD) reduces the superoxide anion, transforming it into hydrogen peroxide, which is less toxic but can still damage cells. The enzyme catalase (CAT) then decomposes hydrogen peroxide. Glutathione S-transferase transfers the thiol group from reduced glutathione, thereby reducing free radical damage [36]. All these enzymes were upregulated in the PO group in the present study, which may be related to the carotenoids present in this oil. Carotenoids are pigments with antioxidant properties; in addition to this capacity, they can modulate transcription factors, such as regulating Nrf-2 (nuclear factor erythroid 2-related factor 2) and downregulating NF-kB (nuclear factor kappa B) [37,38]. The activation of Nrf-2 is associated with greater expression of antioxidant enzymes, such as catalase and superoxide dismutase [39]. Considering that PO contains β-carotene, ζ-carotene, crypto flavin, β-cryptoxanthin, antheraxanthin, zeaxanthin, and mutatoxanthin, as well as phenolic compounds such as ellagic acid, gallic acid, 4-hydroxybenzoic acid, and p-coumaric acid, these compounds may have contributed to the increased expression of antioxidant enzymes [40].
Ellagic acid and gallic acid also showed promise in enhancing the activity of SOD and CAT, demonstrating significant antioxidant capacity through the elimination of DPPH radicals and inhibition of lipid peroxidation, as observed in both in vitro and in vivo studies [41,42]. When copper and iron ions interact with free radicals, lipid hydroperoxide decomposition is accelerated. However, proteins such as transferrin, ferritin, albumin, myoglobin, metallothionein, and ceruloplasmin bind to these metals, preventing Fenton and Haber–Weiss reactions [1]. The present study found that albumin was upregulated, providing more proteins to bind copper and prevent cellular damage caused by oxidative stress. Furthermore, ellagic acid can also act by complexing ionic iron and inhibiting peroxidation [41].
Antioxidant enzymes are present in various locations, including cell membranes. Paraoxonase was also upregulated by PO; this enzyme is a component of HDL lipoproteins, which allows it to hydrolyze LDL and oxidized phospholipids, thus preventing damage to the phospholipid membrane [43]. Peroxiredoxins (PRXs) regulate peroxide levels, providing greater protection against hydrogen peroxide in cells. In the present study, the control diet resulted in the downregulation of these enzymes [44], and this low expression may be related to the lack of activation of chaperones and antioxidant enzymes, making cells more susceptible to oxidation. High levels of hydrogen peroxide have been reported to promote PRX hyperoxidation, leading to inactivation and dysfunction of the enzyme [45,46].
4.3. Lipid Metabolism
Egg yolk contains 33% lipids; therefore, for the correct synthesis and deposition of lipids in the yolk, the laying hen must have a well-functioning lipid metabolism [47]. In hens’ livers, there is continuous synthesis of vitellogenin-1 and vitellogenin-2, which are specialized in transporting nutrients to follicles. However, under heat stress conditions, animals have a reduction in lipid mobilization [48]. In the present study, vitellogenin-1 was positively regulated by PO, demonstrating an improvement in the transport of lipids to the yolk, thus avoiding possible reduction in egg weight [2]. In contrast, the control diet showed downregulation of vitellogenin-2, which may be related to the ineffective management of the negative effects of heat stress in the control group.
Very low-density lipoproteins play a crucial role in transporting lipids to the yolk, where apolipoprotein B can be found. Ovotransferrin is present in the albumen and provides antimicrobial and antioxidant protection for the egg [49,50]. The PO group stimulated the upregulation of apolipoprotein B and ovotransferrin, thereby improving the mechanisms involved in egg production and its shelf life, due to reduced lipid oxidation [51].
Laying hens under heat stress undergo a series of physiological changes to maintain homeostasis, such as the release of corticosteroids by the adrenal glands, which aim to increase energy production through gluconeogenesis. On the other hand, follicular growth is inhibited, and the production of lipoproteins is reduced [52,53,54]. In the present study, PO promoted the upregulation of these proteins linked to egg production. This result may reflect the activation of all antioxidant system levels, which contributed to mitigating the negative effects of hyperthermia and, thus, minimizing the inhibitory action of corticosteroids on lipoproteins, allowing greater productive efficiency.
Heat stress exacerbates lipogenesis, which can lead to ectopic deposition and result in hepatic steatosis [55]. The PO diet downregulated enoyl-CoA hydratase, an enzyme that catalyzes the second stage of the β-oxidation of fatty acids. Therefore, its inhibition may be related to the activation of acetyl-CoA carboxylase, which promotes the conversion of acetyl-CoA into malonyl-CoA. Excess malonyl-CoA can induce negative feedback in lipid degradation through the inhibition of carnitine [56]. Thus, the downregulation of this enzyme may be a response mechanism to control lipid degradation during heat stress.
4.4. Immune System
Laying hens submitted to heat stress conditions present a suppressed immune system [57]. In the present study, the control group showed downregulation of immunoglobulin lambda-like polypeptide 1 (IGLL1) and the Ig lambda chain C region. This result may be associated with the downregulation of chaperones, likely due to the low concentration of carotenoids and phenolic compounds in the control diet. Considering that these molecules have immunomodulatory activity, their absence may have failed to mitigate the effects of heat stress on the immune system. The PO group exhibited upregulation of Ig-like domain-containing protein (IG-like), demonstrating the immunomodulatory capacity of PO, corroborating with Santos et al. [10], who observed the upregulation of IGLL1 and IG-like in the plasma of laying hens supplemented with pequi oil under heat stress conditions. This result is likely due to the increase in chaperones, which act through protective mechanisms to improve immunological responses. The upregulation of HSPs during heat stress improves immune system protection in laying hens by activating immune cells, regulating cytokine production, and enhancing antigen presentation. This modulation strengthens immune defense, making it more effective in responding to challenges [58].
Another hypothesis to this response is the presence of oleic acid (omega-9) in PO, as this monounsaturated fatty acid is involved in anti-inflammatory processes by reducing tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), interleukin-12 (IL-12), and NF-κB. Oleic acid also promotes the antioxidant system by increasing heme oxygenase-1 (HO-1), glutathione peroxidase, superoxide dismutase (SOD), and interleukin-10 (IL-10); these effects have been observed in phagocytic cells [59]. Furthermore, ellagic acid, a phenolic compound present in PO, has antioxidant activity and contributes to the normalization of lipid metabolism and the lipid profile, along with exhibiting immunomodulatory action, which may have contributed to the observed response [60]. Therefore, these factors may have contributed to the PO group’s better immune response to heat stress.
4.5. Unique Proteins
Oxidative stress leads to protein malformation, which, if not repaired, results in degradation. This mechanism is fundamental for preventing the oxidation of other proteins and cellular malfunction [1]. The HUWE1-associated protein, which modifies stress responses, is a ubiquitin ligase that performs cell proliferation, stress signaling, and DNA repair through ubiquitylation [61]. Ubiquitin-40S ribosomal protein S27a, polyubiquitin-B, ubiquitin B, ubiquitin-like domain-containing protein, and HUWE1-associated protein, which modify stress responses, were uniquely regulated by the PO group under heat stress. These proteins are associated with protein degradation processes, and their greater abundance in the PO group is suggested to be linked to the oxidation of target proteins, thereby preventing cellular damage.
The 14-3-3 family of proteins, in addition to their roles in neurotransmitter biosynthesis, signal transduction, and apoptosis, can act as chaperones under heat stress. This protein was positively regulated through heat stress factors, with the 14-3-3 zeta isoform providing protection against protein aggregation and facilitating refolding [62]. Its expression exclusively in the PO group indicates a greater response to stress compared to the control group. This protein may also act indirectly by preventing cell death during oxidative stress through its interaction with FOXO (Forkhead box O) transcription factors, thus inhibiting the upregulation of apoptosis-related genes [63].
Another crucial protein during oxidative stress is ATP-binding cassette subfamily E member 1 (ABCG1). It mobilizes substrates such as cholesterol and antioxidants from inside to outside cells, and negative regulation of this protein is associated with increased production of reactive oxygen species [64]. In the present study, PO promoted the regulation of this protein, contributing to a greater availability of antioxidants to neutralize free radicals. A study with human macrophages reported that palmitic acid promoted upregulation of the ABCG1 gene [65]. Therefore, the regulation of the ABCG1 protein in the PO group may be related to the higher palmitic acid content in this oil, which is 39.06%, compared to only 15.56% in the soybean oil present in the control diet [5,66]. Thus, it is suggested that this factor may have led to the expression of ABCG1 by PO, providing greater protection against oxidative stress.
It was possible to reinforce the hypothesis that PO reduces oxidative stress and stimulates the antioxidant and immune systems. Therefore, this work opens new opportunities for future studies by varying the PO inclusion levels in different stress conditions and identifying specific biomarkers. Furthermore, having observed molecular-level benefits, studies assessing the effects of this supplementation should be conducted through performance evaluations and egg quality assessments.
5. Conclusions
Pequi oil’s inclusion in laying hens’ nutrition strengthens their antioxidant and immune systems, helping to reduce oxidative stress and modulate immune responses. These characteristics are essential for increasing poultry tolerance and minimizing the negative effects of heat on egg production. The liver proteome analysis of laying hens fed pequi oil under heat stress confirms the oil’s potential as an upregulator (p ≥ 0.95 and 1.00) of chaperones (HSP 90; HSP 70), a potentiator of antioxidant defense mechanisms at three levels (protein disulfide-isomerase A4; Su-peroxide dismutase 1_ soluble; catalase), a regulator of lipid metabolism (apolipoprotein B; vitellogenin-1; ovotransferrin), and an immunomodulator (Ig-like domain-containing protein). Additionally, the 15 identified proteins uniquely regulated by pequi oil under heat stress conditions were linked to the maintenance of proteostasis and support increased antioxidant action, and can contribute to the discovery of new biomarkers related to heat stress reduction by phytogenic additives.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture15080867/s1, Supplementary Table S1.
Author Contributions
Conceptualization, P.A.D.R. and J.A.d.S.; methodology, P.A.D.R., J.C.S.V., M.A.R.B., and P.d.M.P., software, M.A.R.B.; validation, P.A.D.R. and P.d.M.P.; formal analysis, P.d.M.P. and M.A.R.B.; investigation, P.A.D.R., J.A.d.S., J.C.S.V., G.A.d.L., L.G.C., E.O.F.C., J.d.L.L., and J.R.S.; data curation, J.C.S.V., G.A.d.L., and P.A.D.R.; writing—original draft preparation, P.A.D.R. and J.R.S.; writing—review and editing, P.A.D.R., J.A.d.S., L.G.C., J.R.S., and P.d.M.P.; visualization, P.A.D.R. and P.d.M.P.; supervision, J.C.S.V., P.d.M.P., and J.R.S.; project administration, J.R.S. and P.d.M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the following Brazilian research-funding agencies: National Council for Scientific and Technological Development—CNPq, Process: 163519/2021-8 (Paola Aparecida Damázio Rodrigues); Coordination of Superior Level Staff Improvement—CAPES-Print AUX-PE, Process: 88881.31074332018-0.1; and Financier of Studies and Projects—FINEP, Process: 0282/19.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee on Animal Use (protocol n° 9 221/2022) in a meeting on 10 October 2022.
Data Availability Statement
The data are contained within the article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
The following abbreviations are used in this manuscript:
| ABCG1 | Binding cassette subfamily E member 1 |
| AMBIC | Bicarbonate |
| BD | Basal diet |
| BiP | Binding immunoglobulin protein |
| BP | Biological process |
| CAT | Catalase |
| CC | Cellular component |
| CEUA | Ethics Committee on the Use of Animals |
| CO | Control diet |
| DTT | Dithiothreitol |
| FMVZ | School of Veterinary Medicine and Animal Science |
| FOXO | Forkhead box O |
| HO | 1- Heme oxygenase-1 |
| HS | Heat stress |
| HSPs | Heat shock proteins |
| IBB | Department of Chemistry and Biochemistry |
| IL-10 | Interleukin-10 |
| IL-12 | Interleukin-12 |
| IL-6 | Interleukin-6 |
| LBM | Bioanalytical and Metalloproteomics Laboratory |
| LC-MS/MS- | Liquid chromatography–tandem mass spectrometry |
| MF | Molecular function |
| NF-kB | Nuclear factor kappa B |
| Nrf-2 | Nuclear factor erythroid 2-related factor 2 |
| PDI | Protein disulfide-isomerase |
| PO | Pequi oil |
| PPIase | Peptidyl-prolyl cis/trans isomerase |
| PRXs | Peroxiredoxins |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| TNF-α | Tumor necrosis factor-alpha |
| UNESP | São Paulo State University |
References
- Surai, P.F. Vitagenes in Avian Biology and Poultry Health; Wageningen Academic Publishers: Wageningen, The Netherlands, 2020. [Google Scholar]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Belal, S.; Kang, D.; Cho, E.; Park, G.; Shim, K. Taurine Reduces Heat Stress by Regulating the Expression of Heat Shock Proteins in Broilers Exposed to Chronic Heat. Braz. J. Poult. Sci. 2018, 20, 479–486. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Chen, M.; Sun, Y.; Safdar, A.; Pasha, R.H.; Munir, M.; Ding, C. Oxidative Stress in Poultry: Lessons from the Viral Infections. Oxid. Med. Cell. Longev. 2018, 14, 5123147. [Google Scholar] [CrossRef]
- Malacrida, C.R.; Freitas, C.; da Vera, E.; Chiea, A. Effect of the application of an enzymatic pretreatment on bioactive compounds of Caryocar brasiliense Camb pulp oil. J. Food Process. Preserv. 2018, 42, 13828. [Google Scholar] [CrossRef]
- Batista, F.O.; Sousa, R.S. Compostos bioativos em frutos pequi (Caryocar brasiliense Camb.) E baru (Dipteryx alata vogel) e seus usos potenciais: Uma revisão. Braz. J. Dev. 2019, 5, 9259–9270. [Google Scholar] [CrossRef]
- Guglielmetti, C.; Manfredi, M.; Brusadore, S.; Sciuto, S.; Esposito, G.P.G.U.; Magnani, L.; Gili, S.; Marengo, E.; Acutis, P.L.; Mazza, M. Two-dimensional gel and shotgun proteomics approaches to distinguish fresh and frozen-thawed curled octopus (Eledone cirrhosa). J. Proteom. 2018, 186, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Kang, D.A.; Shim, K.S. Proteomic changes in broiler liver by body weight differences under chronic heat stress. Poult. Sci. 2022, 101, 101794. [Google Scholar] [CrossRef]
- Rostagno, H.S.; Albino, L.F.T.; Hannas, M.I.; Donzele, J.L.; Sakomura, N.K.; Perazzo, F.G.; Saraiva, A.; Teixeira, M.L.; Rodrigues, P.B.; Oliveira, R.F.; et al. Tabelas brasileiras para aves e suínos: Composição de Alimentos e Exigências Nutricionais. Univ. Fed. Viçosa Dep. Zootec. 2017, 4, 451–488. [Google Scholar]
- Da Silva, J.; Andrade, L.G.; Rodrigues, P.A.D.; Cordeiro, L.G.; Lima, G.A.; Lopes, J.L.; Castillo, E.O.F.; Martins, R.A.; Assunção, A.S.A.; Vieira, J.C.S.; et al. Plasma Proteome Alterations of Laying Hens Subjected to Heat Stress and Fed a Diet Supplemented with Pequi Oil (Caryocar brasiliense Camb.): New Insights in the Identification of Heat Stress Biomarkers. Biomolecules 2024, 14, 1424. [Google Scholar] [CrossRef]
- Cruvinel, J.M.; Urayama, P.M.G.; Oura, C.Y.; Kaiser, F.K.L.; Santos, T.S.; Alves, B.A.; Kadri, S.M.; Corrêa, C.R.; Sartori, J.R.; Pezzato, A.C. Pequi Oil (Caryocar brasiliense Camb.) Attenuates the Adverse Effects of Cyclical Heat Stress and Modulates the Oxidative Stress-Related Genes in Broiler Chickens. Animals 2023, 13, 1896. [Google Scholar] [CrossRef]
- Almeida, E.C.; Faria, V.D.; Cirinêu, F.D.; Santiago, M.G.A.; Miotto, B.; Vieira, J.C.S.; Braga, C.P.; Adamec, J.; Fernandes, A.A.H.; Buzalaf, M.A.R.; et al. Metalloproteomic Investigation of Hg-Binding Proteins in Renal Tissue of Rats Exposed to Mercury Chloride. Int. J. Mol. Sci. 2024, 25, 164. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, L.O.; Chemelo, V.S.; Aragão, W.A.B.; Puty, B.; Dionizio, A.; Teixeira, F.B.; Fernandes, M.S.; Silva, M.C.F.; Fernandes, L.M.P.; de Oliveira, E.H.C.; et al. From Molecules to Behavior in Long-Term Inorganic Mercury Intoxication: Unraveling Proteomic Features in Cerebellar Neurodegeneration of Rats. Int. J. Mol. Sci. 2021, 23, 111. [Google Scholar] [CrossRef]
- Bollengier-lee, S.; Mitchell, M.A.; Utomo, D.B.; Williams, P.E.; Whitehead, C.C. Influence of high dietary vitamin E supplementation on egg production and plasma characteristics in hens subjected to heat stress. Br. Poult. Sci. 1998, 39, 106–112. [Google Scholar] [CrossRef]
- Pasri, P.; Pukkung, C.; Mermillod, P.; Gérard, N.; Jantasaeng, O.; Sirisopapong, M.; Okrathok, S.; Thiabching, T.; Rakngam, S.; Kamkajon, K.; et al. Alleviating heat stress on broiler breeder hens: Effect of dietary antioxidant supplementation on reproductive performance, egg quality, offspring growth, and antioxidant capacity. J. Appl. Poult. Res. 2025, 34. [Google Scholar] [CrossRef]
- Gouda, A.; El-Monairy, M.M.A.; Hassan, H.M.A.; Hamouda, Y.M.A.; Youssef, A.M.A. Lycopene Supplementation Enhances Growth Performance, Antioxidant Enzymes, Heat Shock Protein 70, and Some Biochemical and Immune Parameters in Broiler Chickens Exposed to Heat Stress. Poult. Sci. J. 2025, 13, 127–134. [Google Scholar] [CrossRef]
- Yuan, J.; Li, Y.; Miao, J.; Zhang, X.; Xiong, Y.; Ma, F.; Ding, J.; He, S. Bamboo leaf flavonoids ameliorate cyclic heat stress-induced oxidative damage in broiler liver through activation of Keap1-Nrf2 signaling pathway. Poul. Sci. 2025, 104, 104952. [Google Scholar] [CrossRef]
- Fathi, M.; Zarrinkavyani, K.; Biranvand, Z.; Mustafa, Y. The Effect of Silymarin on Antioxidant, Performance, Immunoglobulin Protein Levels, Cecal Microbiota, and Hemobiochemical Indicators in Heat Stressd Broilers. Poult. Sci. J. 2025, 13, 115–126. [Google Scholar] [CrossRef]
- Xu, L.; Gao, P.; Wu, H.; Gao, Y.; Ji, H.; Huang, X.; Zhang, S.; Fan, W.; Song, S. Lactobacillus plantarum 4-2 alleviates cyclic heat stress-induced oxidative stress and damage in the ileum of laying hens via Keap1-Nrf2 pathway. J. Therm. Biol. 2025, 127, 104072. [Google Scholar] [CrossRef]
- Hashemitabar, S.H.; Hosseinian, S.A. The comparative effects of probiotics on growth, antioxidant indices and intestinal histomorphology of broilers under heat stress condition. Sci. Rep. 2024, 14, 23471. [Google Scholar] [CrossRef]
- Yilmaz, E.; Gul, M. Effects of essential oils on heat-stressed poultry: A review. J. Anim. Physiol. Anim. Nutr. 2024, 108, 1481–1497. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef]
- Herring, G.; Gawlik, D.E. The Role of Stress Proteins in the Study of Allostatic Overload in Birds: Use and Applicability to Current Studies in Avian Ecology. Sci. World J. 2007, 7, 1596–1602. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Nielsen, M.; Carcione, T.; Li, S.; Shi, J. Apolipoprotein E regulates mitochondrial function through the PGC-1α-sirtuin 3 pathway. Aging 2019, 11, 11148–11156. [Google Scholar] [CrossRef]
- Tu, W.; Cheng, C.; Chen, C.; Chan, H.; Wang, S.; Tang, P.; Chen, C.; Lee, Y.; Chen, S.; Huang, S. Annotation of differential protein expression in the hypothalami of layer-type Taiwan country chickens in response to acute heat stress. J. Therm. Biol 2018, 77, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Cara, J.; Aluru, N.; Moyano, F.; Vijayan, M. Food-deprivation induces HSP70 and HSP90 protein expression in larval gilthead sea bream and rainbow trout. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 142, 426–431. [Google Scholar] [CrossRef]
- Yousefi, M.; Adineh, H.; Taheri Mirghaed, A.; Hoseini, S.M. Co-Supplementation of Diet with Saccharomyces cerevisiae and Thymol: Effects on Growth Performance, Antioxidant and Immunological Responses of Rainbow Trout, Oncorhynchus mykiss. Animals 2025, 15, 302. [Google Scholar] [CrossRef]
- Ashraf, E.; Ahmed, M.F.; Shahin, S.A.; Omar, A.A.; Zayed, M.M.; Abdel-Rahim, M.M. Dietary rosemary oil with/without zymogen forte improves water quality, growth hormones, immune-physiological response, stress resilience, and health status of Chelon ramada grown in groundwater. BMC Vet. Res. 2025, 21, 27. [Google Scholar] [CrossRef]
- Büyükdeveci, M.E.; Cengizler, İ.; Balcázar, J.L.; Demirkale, İ. Effects of two host-associated probiotics Bacillus mojavensis B191 and Bacillus subtilis MRS11 on growth performance, intestinal morphology, expression of immune-related genes and disease resistance of Nile tilapia (Oreochromis niloticus) against Streptococcus iniae. Dev. Comp. Immunol. 2023, 138, 104553. [Google Scholar] [CrossRef]
- Schwartz, J.L.; Singh, R.; Teicher, B.A.; Wright, J.E.; Trites, D.H.; Shklar, G. Induction of a 70kD protein associated with the selective cytotoxicity of beta-carotene in human epidermal carcinoma. Biochem. Biophys. Res. Commun. 1990, 169, 941–946. [Google Scholar] [CrossRef]
- Toba, T.; Shidoji, Y.; Fujii, J.; Moriwaki, H.; Muto, Y.; Suzuki, T.; Ohishi, N.; Yagi, K. Growth suppression and induction of heat-shock protein-70 by 9-cis β-carotene in cervical dysplasia-derived cells. Life Sci. 1997, 61, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Sun, M.; Hoxie, R.; Kotler, J.L.M.; Friedman, L.J.; Gelles, J.; Street, T.O. The endoplasmic reticulum chaperone BiP is a closure-accelerating cochaperone of Grp94. Proc. Natl. Acad. Sci. USA 2022, 119, e2118793119. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Wang, C. Protein disulfide–isomerase, a folding catalyst and a redox-regulated chaperone. Free Radic. Biol. Med. 2015, 83, 305–313. [Google Scholar] [CrossRef]
- Gavini, N.; Tungtur, S.; Pulakat, L. Peptidyl-Prolyl cis/trans Isomerase-Independent Functional NifH Mutant of Azotobacter vinelandii. J. Bacteriol. 2006, 188, 6020–6025. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Rattan, S.I.S. Primary Stress Response Pathways for Preconditioning and Physiological Hormesis. In The Science of Hormesis in Health and Longevity; Rattan, S.I.S., Kyriazi, M., Eds.; Academic Press: London, UK, 2019; Volume 39, pp. 35–51. [Google Scholar] [CrossRef]
- Yang, J.; Huang, J.; Shen, C.; Cheng, W.; Yu, P.; Wang, L.; Tang, F.; Guo, S.; Yang, Q.; Zhang, J. Resveratrol Treatment in Different Time-Attenuated Neuronal Apoptosis After Oxygen and Glucose Deprivation/Reoxygenation via Enhancing the Activation of Nrf-2 Signaling Pathway In Vitro. Cell Transplant. 2018, 27, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T. Carotenoids and Markers of Oxidative Stress in Human Observational Stud-ies and Intervention Trials: Implications for Chronic Diseases. Antioxidants 2019, 8, 179. [Google Scholar] [CrossRef]
- Linnewiel, K.; Ernst, H.; Caris-Veyrat, C.; Ben-Dor, A.; Kampf, A.; Salman, H.; Danilenko, M.; Levy, J.; Sharoni, Y. Structure activity relationship of carotenoid derivatives in activation of the electrophile/antioxidant response element transcription system. Free Radic. Biol. Med. 2009, 47, 659–667. [Google Scholar] [CrossRef]
- Brito, R.M.; Barcia, M.T.; Almeida Farias, C.A.; Zambiazi, R.C.; Fujimori, M.; Honorio-França, A.C.; França, E.L.; Pertuzatti, P.B. Bioactive compounds of pequi pulp and oil extracts modulate antioxidant activity and antiproliferative activity in cocultured blood mononuclear cells and breast cancer cells. Food Nutr. Res. 2022, 66, 10-29219. [Google Scholar] [CrossRef]
- Han, D.S.; Lee, M.J.; Kim, J.H. Antioxidant and apoptosis-inducing activities of ellagic acid. Anticancer Res. 2006, 26, 3601–3606. [Google Scholar]
- Ray, S.D.; Krmic, M.; Hussain, A.; Marvilli, C.; Fabian, R.; Niha, A.; Danai, M.; Smith, Z.; Jalshgari, A.; Malick Alhariri, A.; et al. Toxicity of Natural Products, 4th ed.; Academic Press: Oxford, UK, 2023; Volume 4. [Google Scholar] [CrossRef]
- Pinheiro, D.S.; Jesuíno, R.S.A. O Gene da Paraoxonase 1 (PON1) no Contexto Doença Arterial Coronariana. Arq. Bras. Cardiol. 2022, 119, 602–603. [Google Scholar] [CrossRef]
- Perkins, A.; Nelson, K.J.; Parsonage, D.; Poole, L.B.; Karplus, P.A. Peroxiredoxins: Guardians against oxidative stress and modulators of peroxide signaling. Trends Biochem. Sci. 2015, 40, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Wood, Z.A. Peroxiredoxin Evolution and the Regulation of Hydrogen Peroxide Signaling. Science 2003, 300, 650–653. [Google Scholar] [CrossRef]
- Sobotta, M.C.; Liou, W.; Stöcker, S.; Talwar, D.; Oehler, M.; Ruppert, T.; Scharf, A.N.D.; Dick, T.P. Peroxiredoxin-2 and STAT3 form a redox relay for H2O2 signaling. Nat. Chem. Biol. 2015, 11, 64–70. [Google Scholar] [CrossRef]
- Burley, R.W.; Evans, A.J.; Pearson, J.A. Molecular Aspects of the Synthesis and Deposition of Hens’ Egg Yolk with Special Reference to Low Density Lipoprotein. Poul. Sci. 1993, 72, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Baumgard, L.H.; Rhoads, R.P. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed]
- Valenti, P.; Antonini, G.; von Hunolstein, C.; Visca, P.; Orsi, N.; Antonini, E. Studies of the antimicrobial activity of ovotransferrin. PubMed 1983, 5, 97–105. [Google Scholar]
- Ibrahim, H.R.; Hoq, M.d.I.; Aoki, T. Ovotransferrin possesses SOD-like superox-ide anion scavenging activity that is promoted by copper and manganese binding. Int. J. Biol. Macromol. 2007, 41, 631–640. [Google Scholar] [CrossRef]
- Rathnapala, E.C.N.; Ahn, D.U.; Abeyrathne, S. Functional properties of ovotransferrin from chicken egg white and its derived peptides: A review. Food Sci. Biotechnol. 2021, 30, 619–663. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef]
- Wang, X.J.; Li, Y.; Song, Q.Q.; Guo, Y.Y.; Jiao, H.C.; Song, Z.G.; Lin, H. Corticosterone regulation of ovarian follicular development is dependent on the energy status of laying hens. J. Lipid Res. 2013, 54, 1860–1876. [Google Scholar] [CrossRef]
- Kumari, K.N.R.; Nath, X.J. Ameliorative measures to counter heat stress in poultry. Worlds Poult. Sci. J. 2018, 74, 117–130. [Google Scholar] [CrossRef]
- Lu, Z.T.; He, X.T.; Ma, B.; Zhang, L.; Li, J.C.; Jiang, Y.; Zhou, G.J.; Gao, F. Increased fat synthesis and limited apolipoprotein B cause lipid accumulation in the liver of broiler chickens exposed to chronic heat stress. Poul. Sci. 2019, 98, 3695–3704. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, G.; Liu, H. Enoyl-CoA hydratase. reaction, mechanism, and inhibition. Bioorg. Med. Chem. 2003, 11, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, R.; Wei, H.; Wang, Y.; Chen, Y.; Zhang, H.; Li, X.; Liu, H.; Bao, J. Enriched environment housing improved the laying hen’s resistance to transport stress via modulating the heat shock protective response and inflammation. Poul. Sci. 2021, 100, 100939. [Google Scholar] [CrossRef]
- Abare, M.Y.; Rahayu, S.; Tugiyanti, E. Review: The role of heat shock proteins in chicken: Insights into stress adaptation and health. Res. Vet. Sci. 2023, 165, 105057. [Google Scholar] [CrossRef] [PubMed]
- Santa-María, C.; Enríquez, S.L.; la Paz, S.M.; Geniz, I.; Quiroz, M.E.R.; Moreno, M.; Palomares, F.; Sobrino, F.; Alba, G. Update on Anti-Inflammatory Molecular Mechanisms Induced by Oleic Acid. Nutrients 2023, 15, 224. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, K.; Kumar, R.; Jamieson, S.; Pandey, A.K.; Bishayee, A. Neuroprotective Potential of Ellagic Acid: A Critical Review. Adv. Nutr. 2021, 12, 1211–1238. [Google Scholar] [CrossRef]
- Monda, J.K.; Ge, X.; Hunkeler, M.; Donovan, K.A.; Ma, M.W.; Jin, C.Y.; Leonard, M.; Fischer, E.S.; Bennett, E.J. HAPSTR1 localizes HUWE1 to the nucleus to limit stress signaling pathways. Cell Rep. 2023, 42, 112496. [Google Scholar] [CrossRef]
- Yano, M.; Nakamuta, S.; Wu, X.; Okumura, Y.; Kido, H. A Novel Function of 14-3-3 Protein: 14-3-3ζ Is a Heat-Shock–related Molecular Chaperone That Dissolves Thermal-aggregated Proteins. Mol. Biol. Cell. 2006, 17, 4769–4779. [Google Scholar] [CrossRef]
- Pennington, K.; Chan, T.; Torres, M.; Andersen, J. The dynamic and stress-adaptive signaling hub of 14-3-3: Emerging mechanisms of regulation and context-dependent protein–protein interactions. Oncogene 2018, 37, 5587–5604. [Google Scholar] [CrossRef]
- Xue, J.; Fan, J.; Li, Y.; Wu, W.; Yan, Q.; Zheng, Q. ABCG1 Attenuates Oxidative Stress Induced by H2O2 through the Inhibition of NADPH Oxidase and the Upregulation of Nrf2-Mediated Antioxidant Defense in Endothelial Cells. Anal. Cell. Pathol. 2020, 2020, 2095645. [Google Scholar] [CrossRef] [PubMed]
- Kardassis, D.; Gafencu, A.; Zannis, V.I.; Davalos, A. Regulation of HDL Genes: Transcriptional, Posttranscriptional, and Posttranslational. Handb. Exp. Pharmacol. 2014, 224, 113–179. [Google Scholar] [CrossRef]
- Roll, A.A.P.; Forgiarini, J.; Xavier, E.G.; Lopes, D.C.N.; Roll, V.F.B.; Rutz, F. Replacing soybean oil with increasing levels of soybean acid oil in diets of coturnix quails (Coturnix coturnix coturnix) and the effects on egg quality. An. Acad. Bras. Cienc. 2017, 90, 529–539. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).