
Corin Deficiency Diminishes Intestinal Sodium Excretion in Mice
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mouse Models
2.2. Tissue Collection and Preparation
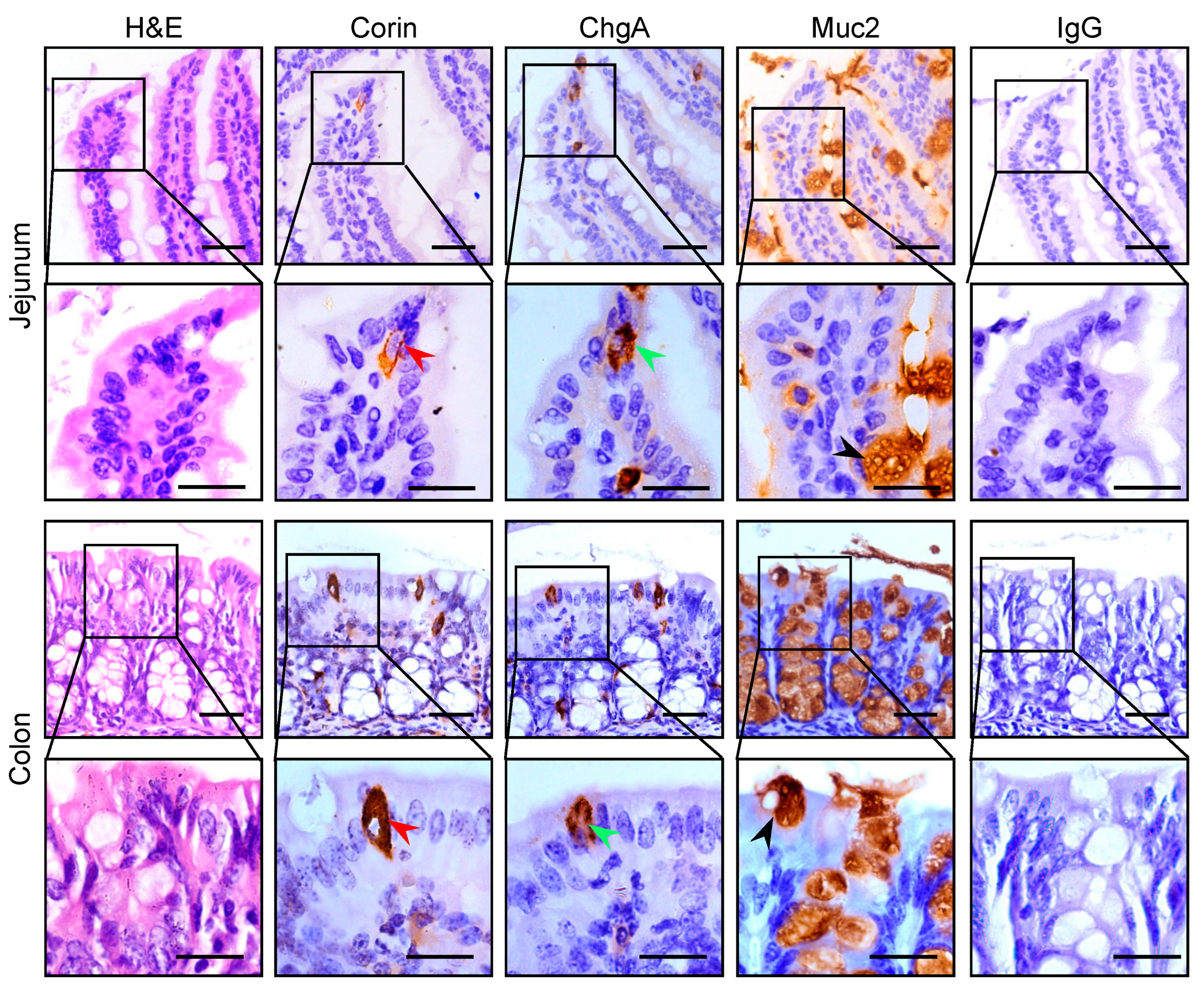
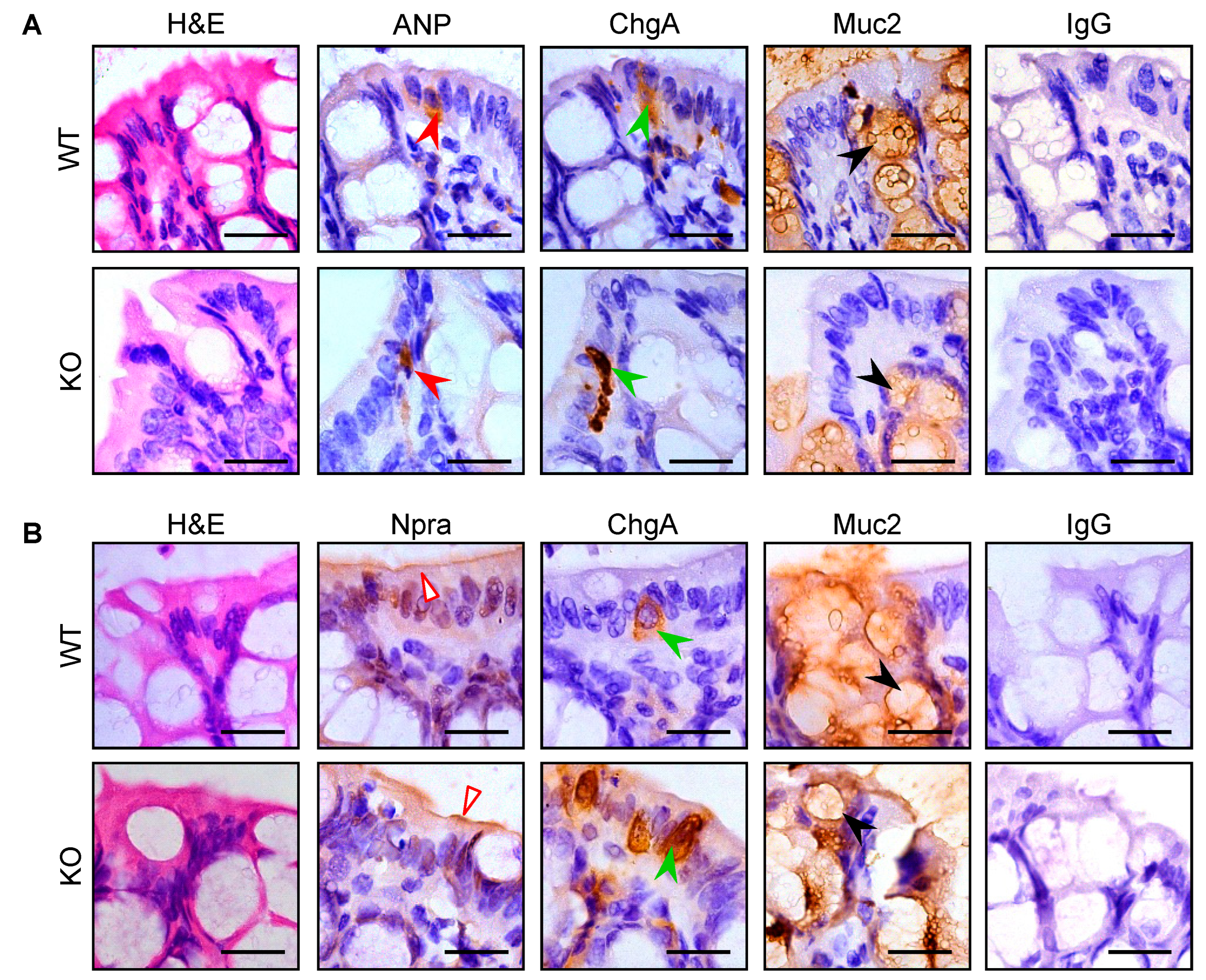
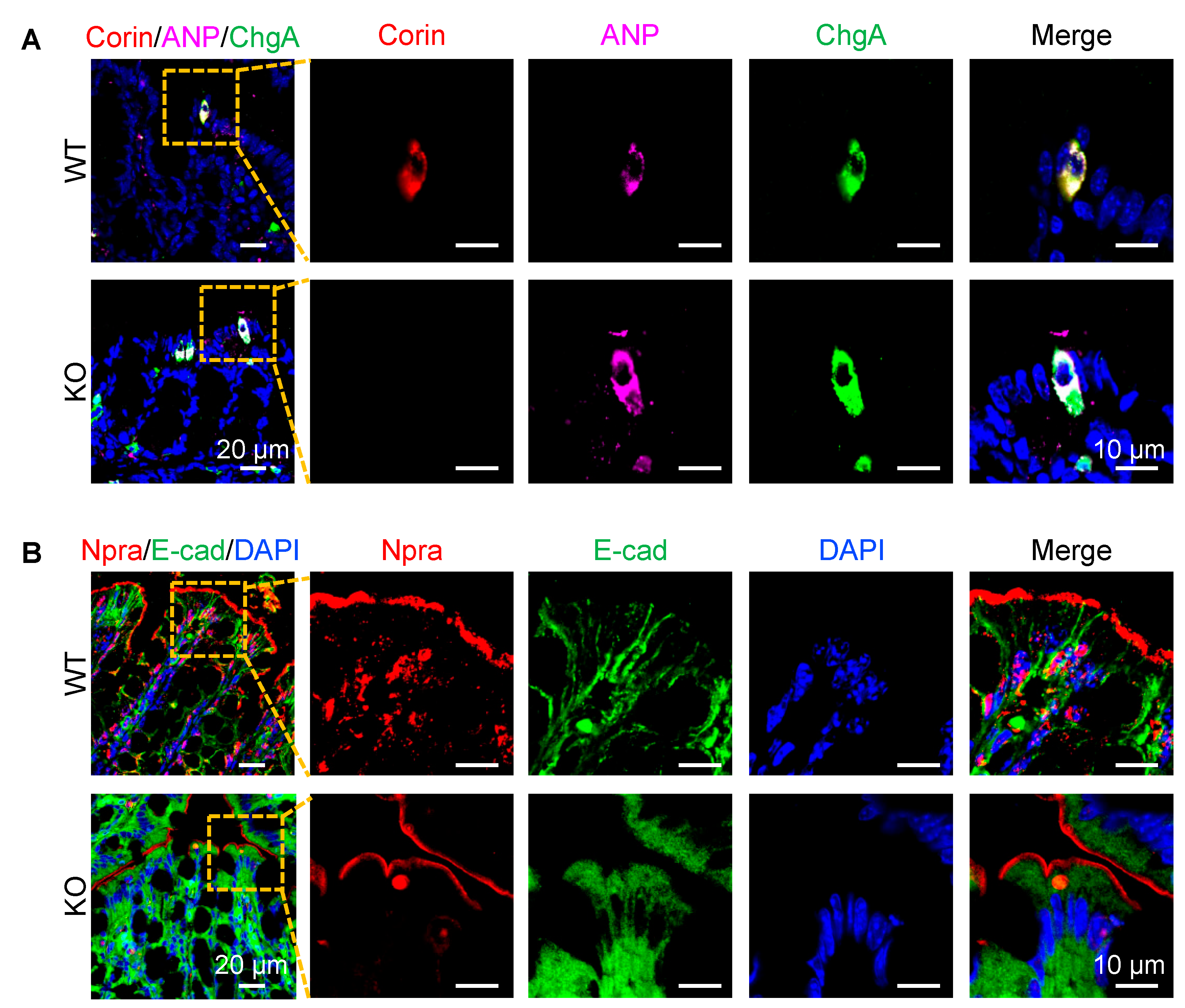
2.3. Immunostaining
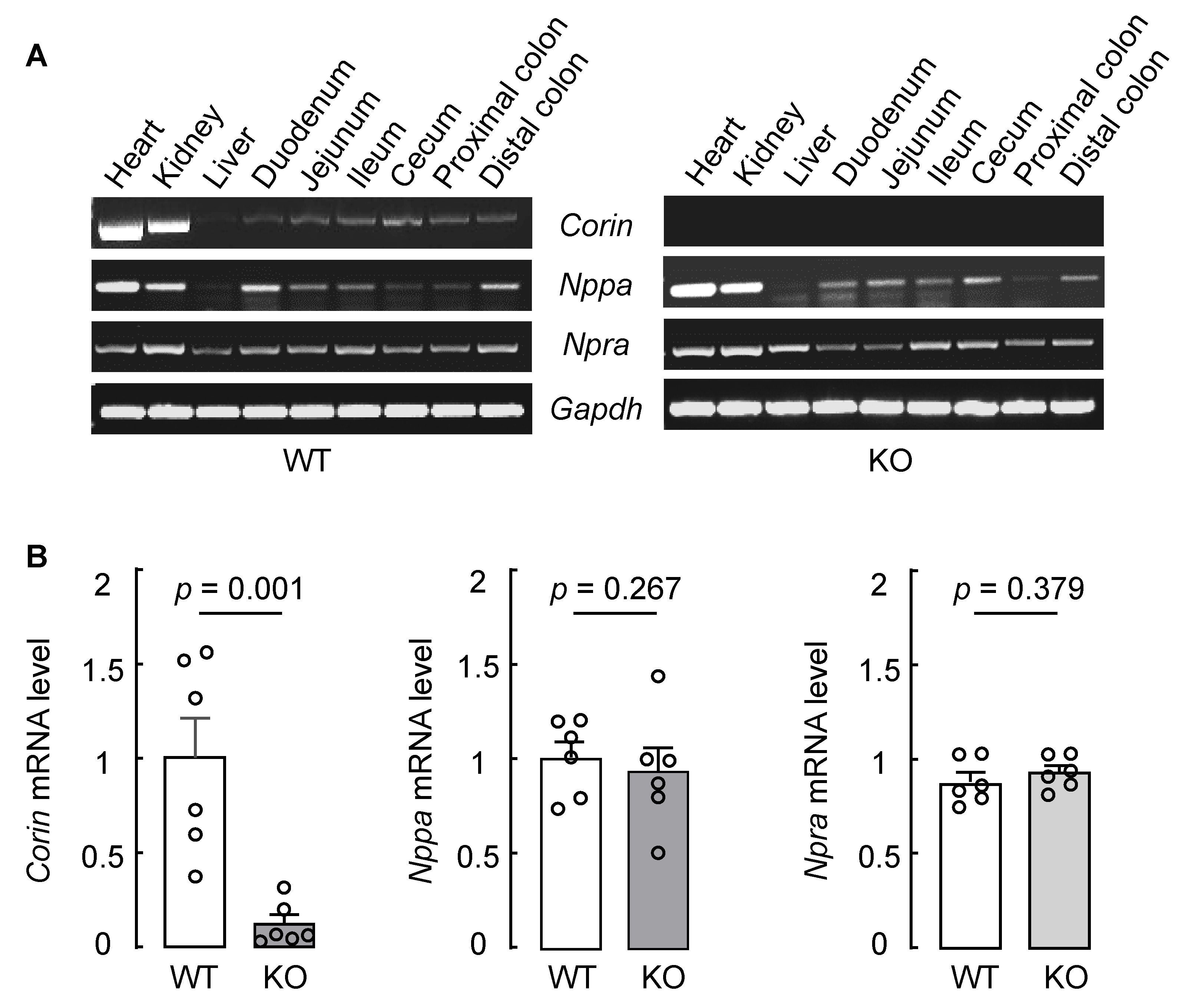
2.4. Gene Expression Analysis by PCR and qRT-PCR
2.5. Aldosterone Treatment
2.6. Fecal Na+ and Cl− Measurements
2.7. Statistics
3. Results
3.1. Corin Protein Expression in Mouse Intestines
3.2. Intestinal Corin, Nppa, and Npra mRNA Expression in WT and Corin KO Mice
3.3. Intestinal Pro-ANP/ANP and Npra Protein Expression in WT and Corin KO Mice
3.4. Co-Localization of Corin and ANP Proteins in Intestinal Enteroendocrine Cells
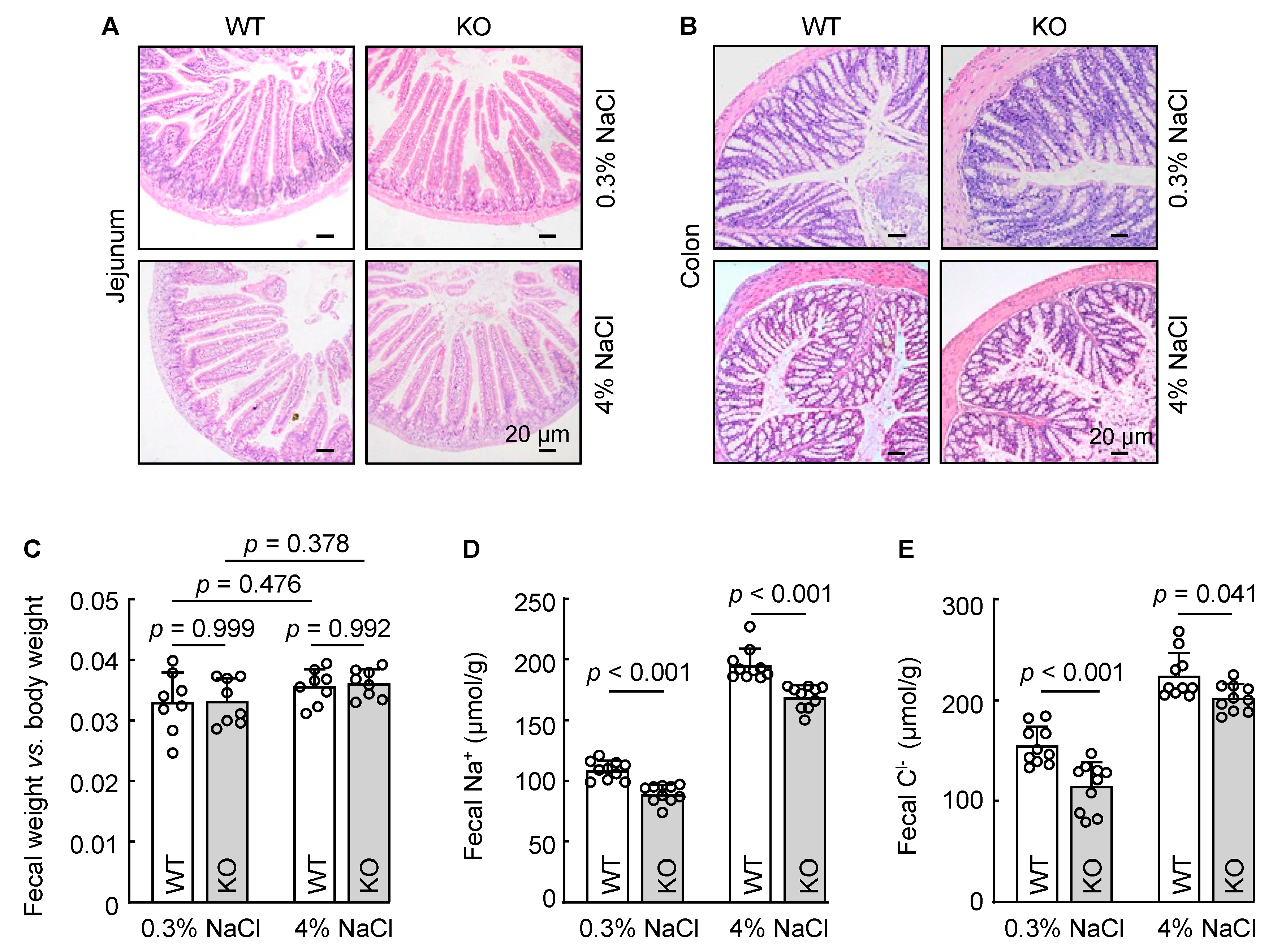
3.5. Reduced Fecal Na+ and Cl− Excretion in Corin KO Mice
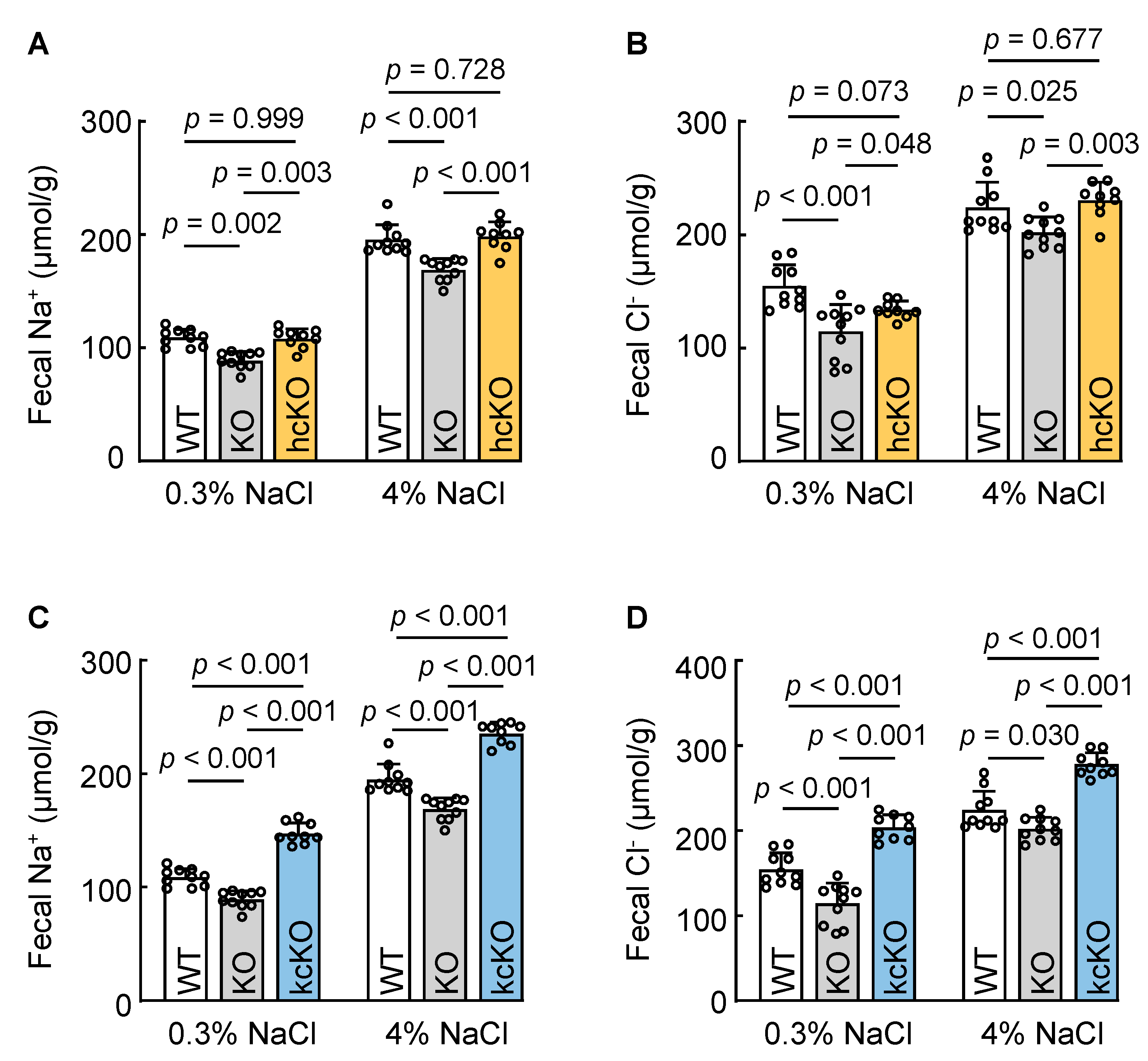
3.6. Cardiac Corin Is Dispensable for Intestinal Na+ and Cl− Excretion in Mice
3.7. Renal-Corin-Deficient Mice Exhibited Increased Fecal Na+ and Cl− Excretion
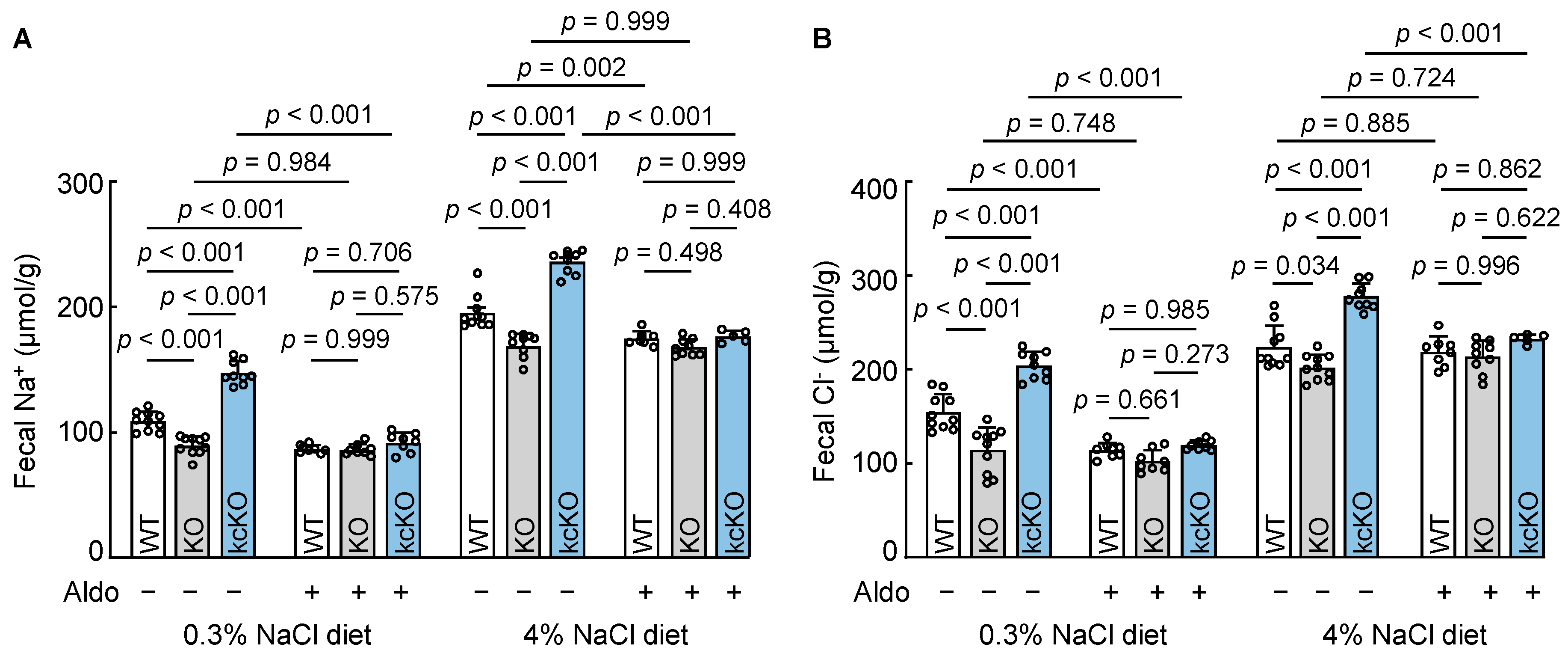
3.8. Effects of Aldosterone on Fecal Na+ and Cl− Excretion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernal, A.; Zafra, M.A.; Simón, M.J.; Mahía, J. Sodium Homeostasis, a Balance Necessary for Life. Nutrients 2023, 15, 395. [Google Scholar] [CrossRef] [PubMed]
- Strazzullo, P.; D’Elia, L.; Kandala, N.B.; Cappuccio, F.P. Salt intake, stroke, and cardiovascular disease: Meta-analysis of prospective studies. BMJ 2009, 339, b4567. [Google Scholar] [CrossRef] [Green Version]
- Osanai, T.; Fujiwara, N.; Saitoh, M.; Sasaki, S.; Tomita, H.; Nakamura, M.; Osawa, H.; Yamabe, H.; Okumura, K. Relationship between salt intake, nitric oxide and asymmetric dimethylarginine and its relevance to patients with end-stage renal disease. Blood Purif. 2002, 20, 466–468. [Google Scholar] [CrossRef] [PubMed]
- Rossier, B.C.; Bochud, M.; Devuyst, O. The Hypertension Pandemic: An Evolutionary Perspective. Physiology 2017, 32, 112–125. [Google Scholar] [CrossRef]
- Ellison, D.H.; Welling, P. Insights into Salt Handling and Blood Pressure. N. Engl. J. Med. 2021, 385, 1981–1993. [Google Scholar] [CrossRef]
- Bie, P. Mechanisms of sodium balance: Total body sodium, surrogate variables, and renal sodium excretion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R945–R962. [Google Scholar] [CrossRef] [Green Version]
- Wagner, C.A. Effect of mineralocorticoids on acid-base balance. Nephron Physiol. 2014, 128, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Theilig, F.; Wu, Q. ANP-induced signaling cascade and its implications in renal pathophysiology. Am. J. Physiol. Renal. Physiol. 2015, 308, F1047–F1055. [Google Scholar] [CrossRef]
- Goetze, J.P.; Bruneau, B.G.; Ramos, H.R.; Ogawa, T.; de Bold, M.K.; de Bold, A.J. Cardiac natriuretic peptides. Nat. Rev. Cardiol. 2020, 17, 698–717. [Google Scholar] [CrossRef]
- Kuhn, M. Molecular Physiology of Membrane Guanylyl Cyclase Receptors. Physiol. Rev. 2016, 96, 751–804. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Wu, F.; Morser, J.; Wu, Q. Corin, a transmembrane cardiac serine protease, acts as a pro-atrial natriuretic peptide-converting enzyme. Proc. Natl. Acad. Sci. USA 2000, 97, 8525–8529. [Google Scholar] [CrossRef]
- Dong, N.; Niu, Y.; Chen, Y.; Sun, S.; Wu, Q. Function and regulation of corin in physiology and disease. Biochem. Soc. Trans. 2020, 48, 1905–1916. [Google Scholar] [CrossRef]
- Yan, W.; Sheng, N.; Seto, M.; Morser, J.; Wu, Q. Corin, a mosaic transmembrane serine protease encoded by a novel cDNA from human heart. J. Biol. Chem. 1999, 274, 14926–14935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, J.D.; Clements, J.A.; Quigley, J.P.; Antalis, T.M. Type II transmembrane serine proteases. Insights into an emerging class of cell surface proteolytic enzymes. J. Biol. Chem. 2001, 276, 857–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, R.; Bugge, T.H. Membrane-anchored serine proteases as regulators of epithelial function. Biochem. Soc. Trans. 2020, 48, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.E.; List, K. Cell surface-anchored serine proteases in cancer progression and metastasis. Cancer Metastasis Rev. 2019, 38, 357–387. [Google Scholar] [CrossRef]
- Hooper, J.D.; Scarman, A.L.; Clarke, B.E.; Normyle, J.F.; Antalis, T.M. Localization of the mosaic transmembrane serine protease corin to heart myocytes. Eur. J. Biochem. 2000, 267, 6931–6937. [Google Scholar] [CrossRef] [PubMed]
- Ichiki, T.; Huntley, B.K.; Heublein, D.M.; Sandberg, S.M.; McKie, P.M.; Martin, F.L.; Jougasaki, M.; Burnett, J.C., Jr. Corin is present in the normal human heart, kidney, and blood, with pro-B-type natriuretic peptide processing in the circulation. Clin. Chem. 2011, 57, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.C.; Knudson, O.; Wu, F.; Morser, J.; Dole, W.P.; Wu, Q. Hypertension in mice lacking the proatrial natriuretic peptide convertase corin. Proc. Natl. Acad. Sci. USA 2005, 102, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cui, Y.; Shen, J.; Jiang, J.; Chen, S.; Peng, J.; Wu, Q. Salt-sensitive hypertension and cardiac hypertrophy in transgenic mice expressing a corin variant identified in blacks. Hypertension 2012, 60, 1352–1358. [Google Scholar] [CrossRef] [Green Version]
- Nigrovic, P.A.; Gray, D.H.; Jones, T.; Hallgren, J.; Kuo, F.C.; Chaletzky, B.; Gurish, M.; Mathis, D.; Benoist, C.; Lee, D.M. Genetic inversion in mast cell-deficient (Wsh) mice interrupts corin and manifests as hematopoietic and cardiac aberrancy. Am. J. Pathol. 2008, 173, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Buckley, C.L.; Stokes, A.J. Corin-deficient W-sh mice poorly tolerate increased cardiac afterload. Regul. Pept. 2011, 172, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Shen, J.; Cui, Y.; Jiang, J.; Chen, S.; Peng, J.; Wu, Q. Impaired sodium excretion and salt-sensitive hypertension in corin-deficient mice. Kidney Int. 2012, 82, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, T.; Yao, S.; Du, M.F.; Mu, J.J.; Chu, C.; Hu, G.L.; Liao, Y.Y.; Chen, C.; Wang, D.; Ma, Q.; et al. Associations of corin genetic polymorphisms with salt sensitivity, blood pressure changes, and hypertension incidence in Chinese adults. J. Clin. Hypertens. 2021, 23, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Dries, D.L.; Victor, R.G.; Rame, J.E.; Cooper, R.S.; Wu, X.; Zhu, X.; Leonard, D.; Ho, S.I.; Wu, Q.; Post, W.; et al. Corin gene minor allele defined by 2 missense mutations is common in blacks and associated with high blood pressure and hypertension. Circulation 2005, 112, 2403–2410. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhou, T.; Niu, Y.; He, M.; Wang, C.; Liu, M.; Yang, J.; Zhang, Y.; Zhou, J.; Fukuda, K.; et al. Identification and functional analysis of CORIN variants in hypertensive patients. Hum. Mutat. 2017, 38, 1700–1710. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yuan, X.; Zhong, Y.; Zhang, Y.; Zhang, S.; Li, S.; Zhao, Y.; Zheng, W.; Liu, J.; Xia, Y.; et al. Single-Nucleotide Polymorphisms in the 3’ Untranslated Region of CORIN Associated with Cardiovascular Diseases in a Chinese Han Population: A Case-Control Study. Front. Cardiovasc. Med. 2021, 8, 625072. [Google Scholar] [CrossRef]
- Dong, N.; Fang, C.; Jiang, Y.; Zhou, T.; Liu, M.; Zhou, J.; Shen, J.; Fukuda, K.; Qin, J.; Wu, Q. Corin mutation R539C from hypertensive patients impairs zymogen activation and generates an inactive alternative ectodomain fragment. J. Biol. Chem. 2013, 288, 7867–7874. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Zhou, T.; Zhang, Y.; Liu, M.; Li, H.; Huang, X.; Liu, Z.; Wu, Y.; Fukuda, K.; Qin, J.; et al. Corin mutations K317E and S472G from preeclamptic patients alter zymogen activation and cell surface targeting. J. Biol. Chem. 2014, 289, 17909–17916. [Google Scholar] [CrossRef] [Green Version]
- Ibebuogu, U.N.; Gladysheva, I.P.; Houng, A.K.; Reed, G.L. Decompensated heart failure is associated with reduced corin levels and decreased cleavage of pro-atrial natriuretic peptide. Circ. Heart Fail. 2011, 4, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Zhu, F.; Shi, J.; Han, X.; Zhou, D.; Liu, Y.; Zhi, Z.; Zhang, F.; Shen, Y.; Ma, J.; et al. Serum Soluble Corin is Decreased in Stroke. Stroke 2015, 46, 1758–1763. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhang, Q.; Zhang, M.; Yu, J.; Ren, L.; Li, J.; Ma, S.; He, Y.; Hu, W.; Peng, H. Soluble Corin Predicts the Risk of Cardiovascular Disease: A 10-Year Follow-Up Study. JACC Asia 2022, 2, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.; Chen, S.; Wang, W.; Zhou, Y.; Wu, Q. Corin in clinical laboratory diagnostics. Clin. Chim. Acta 2012, 413, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zhang, Q.; Peng, H.; Zhong, C.; Guo, D.; Huangfu, X.; Chao, X.; Wang, A.; Jin, J.; Zhang, Y. Predictive value of serum soluble corin in the risk of hyperglycemia: A population-based prospective cohort study in China. Clin. Chim. Acta 2018, 479, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Han, X.; Zhang, X.; Wang, Y.; Wang, T. Circulating soluble corin as a potential biomarker for cardiovascular diseases: A translational review. Clin. Chim. Acta 2018, 485, 106–112. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; He, M.; Zhou, T.; Niu, Y.; Sun, S.; Li, H.; Zhang, C.; Zhang, S.; Liu, M.; et al. Krüppel-like factor 17 upregulates uterine corin expression and promotes spiral artery remodeling in pregnancy. Proc. Natl. Acad. Sci. USA 2020, 117, 19425–19434. [Google Scholar] [CrossRef] [PubMed]
- Kaitu’u-Lino, T.J.; Ye, L.; Tuohey, L.; Dimitriadis, E.; Bulmer, J.; Rogers, P.; Menkhorst, E.; Van Sinderen, M.; Girling, J.E.; Hannan, N.; et al. Corin, an enzyme with a putative role in spiral artery remodeling, is up-regulated in late secretory endometrium and first trimester decidua. Hum. Reprod. 2013, 28, 1172–1180. [Google Scholar] [CrossRef] [Green Version]
- Mika, K.; Marinić, M.; Singh, M.; Muter, J.; Brosens, J.J.; Lynch, V.J. Evolutionary transcriptomics implicates new genes and pathways in human pregnancy and adverse pregnancy outcomes. eLife 2021, 10, e69584. [Google Scholar] [CrossRef]
- Fang, C.; Shen, L.; Dong, L.; Liu, M.; Shi, S.; Dong, N.; Wu, Q. Reduced urinary corin levels in patients with chronic kidney disease. Clin. Sci. 2013, 124, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Yin, Y.; Chen, L.; Chu, C.; Wang, Y.; Lv, Y.; He, M.; Martin, M.; Huang, P.H.; Mu, J.J.; et al. Short-Term High-Salt Diet Increases Corin Level to Regulate the Salt-Water Balance in Humans and Rodents. Am. J. Hypertens. 2018, 31, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Avigad Laron, E.; Aamar, E.; Enshell-Seijffers, D. The Serine Protease Activity of Corin Is Required for Normal Pigment Type Switching. J. Investig. Dermatol. 2019, 139, 257–259. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Zhou, T.; Niu, Y.; Feng, W.; Gu, X.; Xu, W.; Zhang, S.; Wang, Z.; Zhang, Y.; Wang, C.; et al. The protease corin regulates electrolyte homeostasis in eccrine sweat glands. PLoS Biol. 2021, 19, e3001090. [Google Scholar] [CrossRef]
- Nordberg, R.C.; Wang, H.; Wu, Q.; Loboa, E.G. Corin is a key regulator of endochondral ossification and bone development via modulation of vascular endothelial growth factor A expression. J. Tissue Eng. Regen. Med. 2018, 12, 2277–2286. [Google Scholar] [CrossRef]
- Zhou, H.; Zhu, J.; Liu, M.; Wu, Q.; Dong, N. Role of the protease corin in chondrogenic differentiation of human bone marrow-derived mesenchymal stem cells. J. Tissue Eng. Regen. Med. 2018, 12, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, W.; Dong, N.; Lou, J.; Srinivasan, D.K.; Cheng, W.; Huang, X.; Liu, M.; Fang, C.; Peng, J.; et al. Role of corin in trophoblast invasion and uterine spiral artery remodelling in pregnancy. Nature 2012, 484, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Li, S.; Lou, J.; Li, H.; Liu, M.; Dong, N.; Wu, Q. Atrial natriuretic peptide promotes uterine decidualization and a TRAIL-dependent mechanism in spiral artery remodeling. J. Clin. Investig. 2021, 131, e151053. [Google Scholar] [CrossRef]
- Zhou, T.; Zhang, S.; Du, C.; Wang, K.; Gu, X.; Sun, S.; Zhang, X.; Niu, Y.; Wang, C.; Liu, M.; et al. Renal Corin Is Essential for Normal Blood Pressure and Sodium Homeostasis. Int. J. Mol. Sci. 2022, 23, 11251. [Google Scholar] [CrossRef]
- Polzin, D.; Kaminski, H.J.; Kastner, C.; Wang, W.; Krämer, S.; Gambaryan, S.; Russwurm, M.; Peters, H.; Wu, Q.; Vandewalle, A.; et al. Decreased renal corin expression contributes to sodium retention in proteinuric kidney diseases. Kidney Int. 2010, 78, 650–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoury, E.E.; Fokra, A.; Kinaneh, S.; Knaney, Y.; Aronson, D.; Abassi, Z. Distribution of Cardiac and Renal Corin and Proprotein Convertase Subtilisin/Kexin-6 in the Experimental Model of Cardio-Renal Syndrome of Various Severities. Front Physiol. 2021, 12, 673497. [Google Scholar] [CrossRef]
- Hu, X.; Su, M.; Lin, J.; Zhang, L.; Sun, W.; Zhang, J.; Sun, W.; Zhang, J.; Tian, Y.; Qiu, W. Corin Is Downregulated in Renal Ischemia/Reperfusion Injury and Is Associated with Delayed Graft Function after Kidney Transplantation. Dis. Markers 2019, 2019, 9429323. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Romero, M.F. Regulation of electroneutral NaCl absorption by the small intestine. Annu. Rev. Physiol. 2011, 73, 261–281. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.C. Physiology of Electrolyte Transport in the Gut: Implications for Disease. Compr. Physiol. 2019, 9, 947–1023. [Google Scholar] [PubMed]
- Michell, A.R. The gut: The unobtrusive regulator of sodium balance. Perspect. Biol. Med. 1986, 29, 203–213. [Google Scholar] [CrossRef]
- Dong, L.; Wang, H.; Dong, N.; Zhang, C.; Xue, B.; Wu, Q. Localization of corin and atrial natriuretic peptide expression in human renal segments. Clin. Sci. 2016, 130, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Engelstoft, M.S.; Lund, M.L.; Grunddal, K.V.; Egerod, K.L.; Osborne-Lawrence, S.; Poulsen, S.S.; Zigman, J.M.; Schwartz, T.W. Research Resource: A Chromogranin A Reporter for Serotonin and Histamine Secreting Enteroendocrine Cells. Mol. Endocrinol. 2015, 29, 1658–1671. [Google Scholar] [CrossRef]
- Wang, Y.; Song, W.; Wang, J.; Wang, T.; Xiong, X.; Qi, Z.; Fu, W.; Yang, X.; Chen, Y.G. Single-cell transcriptome analysis reveals differential nutrient absorption functions in human intestine. J. Exp. Med. 2020, 217, e20191130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gu, X.; Zhang, Y.; Dong, N.; Wu, Q. Corin: A Key Mediator in Sodium Homeostasis, Vascular Remodeling, and Heart Failure. Biology 2022, 11, 717. [Google Scholar] [CrossRef] [PubMed]
- Armaly, Z.; Assady, S.; Abassi, Z. Corin: A new player in the regulation of salt-water balance and blood pressure. Curr. Opin. Nephrol. Hypertens. 2013, 22, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, I.P.; Robinson, B.R.; Houng, A.K.; Kováts, T.; King, S.M. Corin is co-expressed with pro-ANP and localized on the cardiomyocyte surface in both zymogen and catalytically active forms. J. Mol. Cell. Cardiol. 2008, 44, 131–142. [Google Scholar] [CrossRef]
- Tran, K.L.; Lu, X.; Lei, M.; Feng, Q.; Wu, Q. Upregulation of corin gene expression in hypertrophic cardiomyocytes and failing myocardium. Am. J. Physiol.-Heart Circ. Physiol. 2004, 287, H1625–H1631. [Google Scholar] [CrossRef]
- Wu, F.; Yan, W.; Pan, J.; Morser, J.; Wu, Q. Processing of pro-atrial natriuretic peptide by corin in cardiac myocytes. J. Biol. Chem. 2002, 277, 16900–16905. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Itoh, H. Hypertension as a Metabolic Disorder and the Novel Role of the Gut. Curr. Hypertens. Rep. 2019, 21, 63. [Google Scholar] [CrossRef] [Green Version]
- Giordano, L.; Mihaila, S.M.; Eslami Amirabadi, H.; Masereeuw, R. Microphysiological Systems to Recapitulate the Gut-Kidney Axis. Trends Biotechnol. 2021, 39, 811–823. [Google Scholar] [CrossRef]
- Michell, A.R.; Debnam, E.S.; Unwin, R.J. Regulation of renal function by the gastrointestinal tract: Potential role of gut-derived peptides and hormones. Annu. Rev. Physiol. 2008, 70, 379–403. [Google Scholar] [CrossRef]
- Hatch, M. Intestinal adaptations in chronic kidney disease and the influence of gastric bypass surgery. Exp. Physiol. 2014, 99, 1163–1167. [Google Scholar] [CrossRef] [Green Version]
- Marmol, F.; Badaruddin, M.; Baig, A.; Ye, M.; Wysocki, J.; Tahaei, E.; Welling, P.; Bamberg, K.; Batlle, D. Fecal ammonium in mice with CKD: Gastrointestinal sequestration by sodium zirconium cyclosilicate. Am. J. Physiol.-Ren. Physiol. 2023, 324, F464–F471. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M.; Freel, R.W.; Vaziri, N.D. Local upregulation of colonic angiotensin II receptors enhances potassium excretion in chronic renal failure. Am. J. Physiol. 1998, 274, F275–F282. [Google Scholar] [CrossRef] [PubMed]
- Malsure, S.; Wang, Q.; Charles, R.P.; Sergi, C.; Perrier, R.; Christensen, B.M.; Maillard, M.; Rossier, B.C.; Hummler, E. Colon-specific deletion of epithelial sodium channel causes sodium loss and aldosterone resistance. J. Am. Soc. Nephrol. 2014, 25, 1453–1464. [Google Scholar] [CrossRef] [Green Version]
- Booth, R.E.; Johnson, J.P.; Stockand, J.D. Aldosterone. Adv. Physiol. Educ. 2002, 26, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Negussie, A.B.; Dell, A.C.; Davis, B.A.; Geibel, J.P. Colonic Fluid and Electrolyte Transport 2022: An Update. Cells 2022, 11, 1712. [Google Scholar] [CrossRef] [PubMed]
- Masilamani, S.; Kim, G.H.; Mitchell, C.; Wade, J.B.; Knepper, M.A. Aldosterone-mediated regulation of ENaC alpha, beta, and gamma subunit proteins in rat kidney. J. Clin. Investig. 1999, 104, R19–R23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotin, D.; Staub, O. Function and Regulation of the Epithelial Na(+) Channel ENaC. Compr. Physiol. 2021, 11, 2017–2045. [Google Scholar] [PubMed]
- Rossier, B.C.; Staub, O.; Hummler, E. Genetic dissection of sodium and potassium transport along the aldosterone-sensitive distal nephron: Importance in the control of blood pressure and hypertension. FEBS Lett. 2013, 587, 1929–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, X.; Wang, K.; Li, W.; He, M.; Zhou, T.; Liu, M.; Wu, Q.; Dong, N. Corin Deficiency Diminishes Intestinal Sodium Excretion in Mice. Biology 2023, 12, 945. https://doi.org/10.3390/biology12070945
Gu X, Wang K, Li W, He M, Zhou T, Liu M, Wu Q, Dong N. Corin Deficiency Diminishes Intestinal Sodium Excretion in Mice. Biology. 2023; 12(7):945. https://doi.org/10.3390/biology12070945
Chicago/Turabian StyleGu, Xiabing, Kun Wang, Wenguo Li, Meiling He, Tiantian Zhou, Meng Liu, Qingyu Wu, and Ningzheng Dong. 2023. "Corin Deficiency Diminishes Intestinal Sodium Excretion in Mice" Biology 12, no. 7: 945. https://doi.org/10.3390/biology12070945