
A Hardware-Based Orientation Detection System Using Dendritic Computation

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
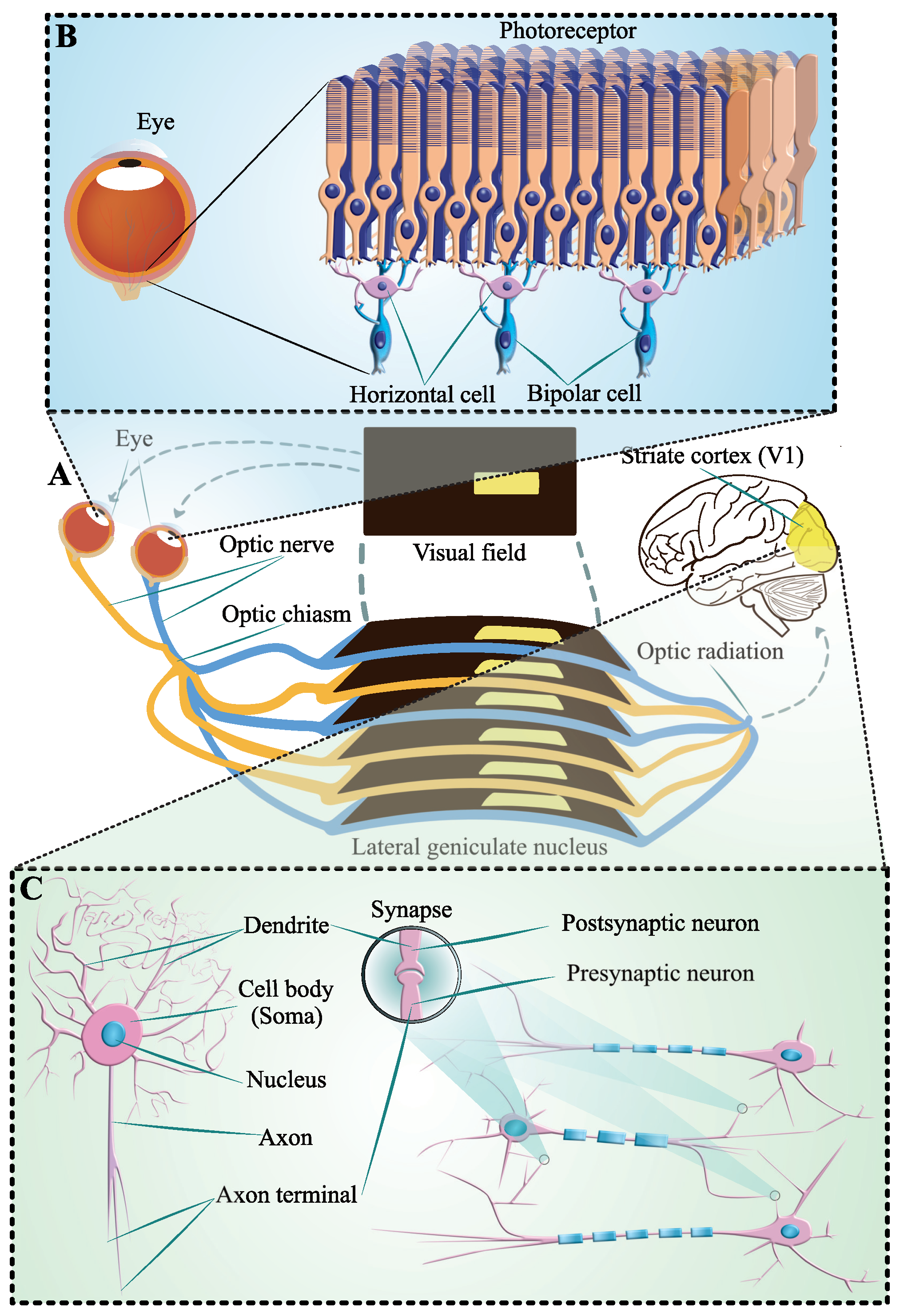
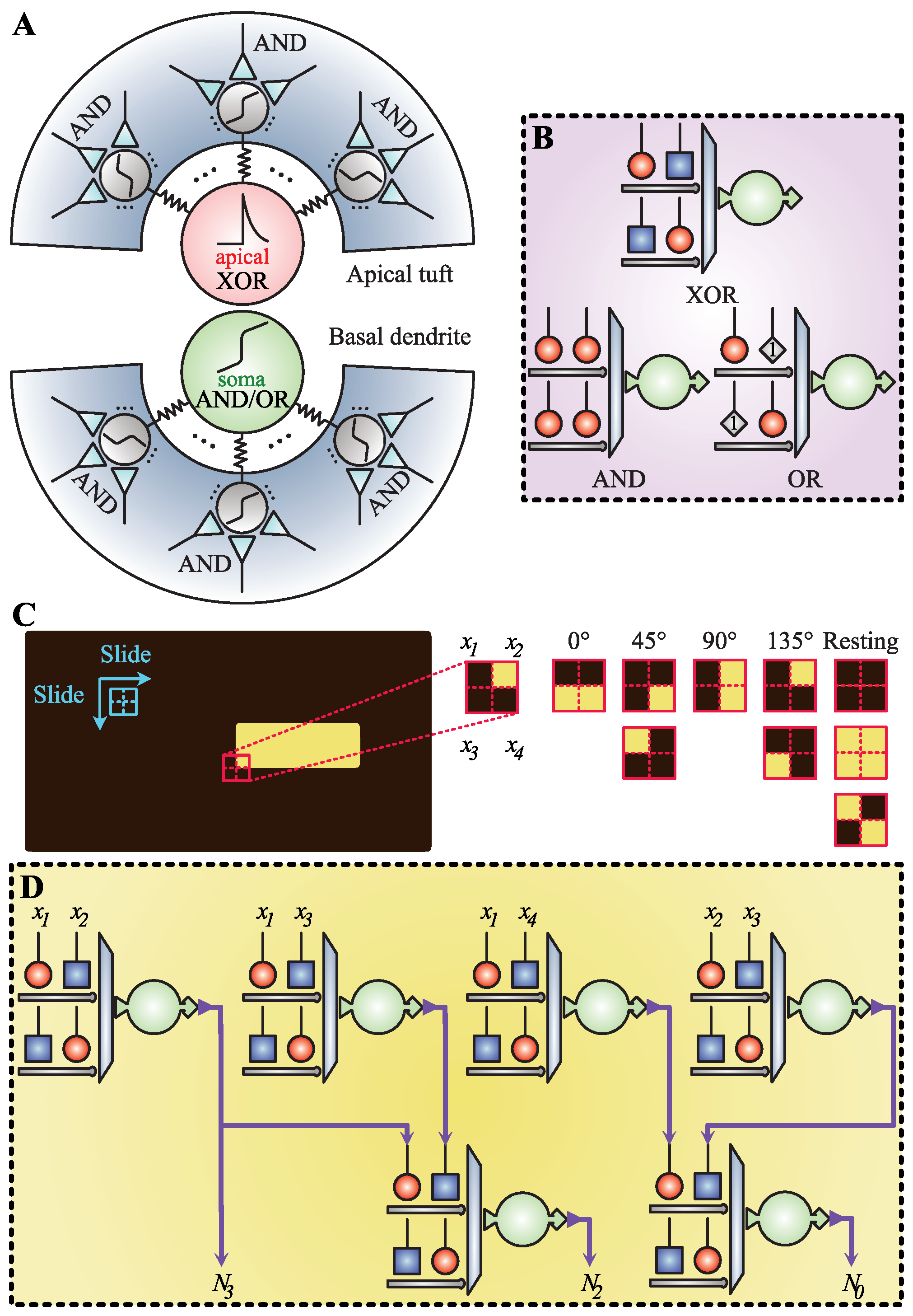
2.1. Dendritic Neuron Model
2.1.1. Model Structure
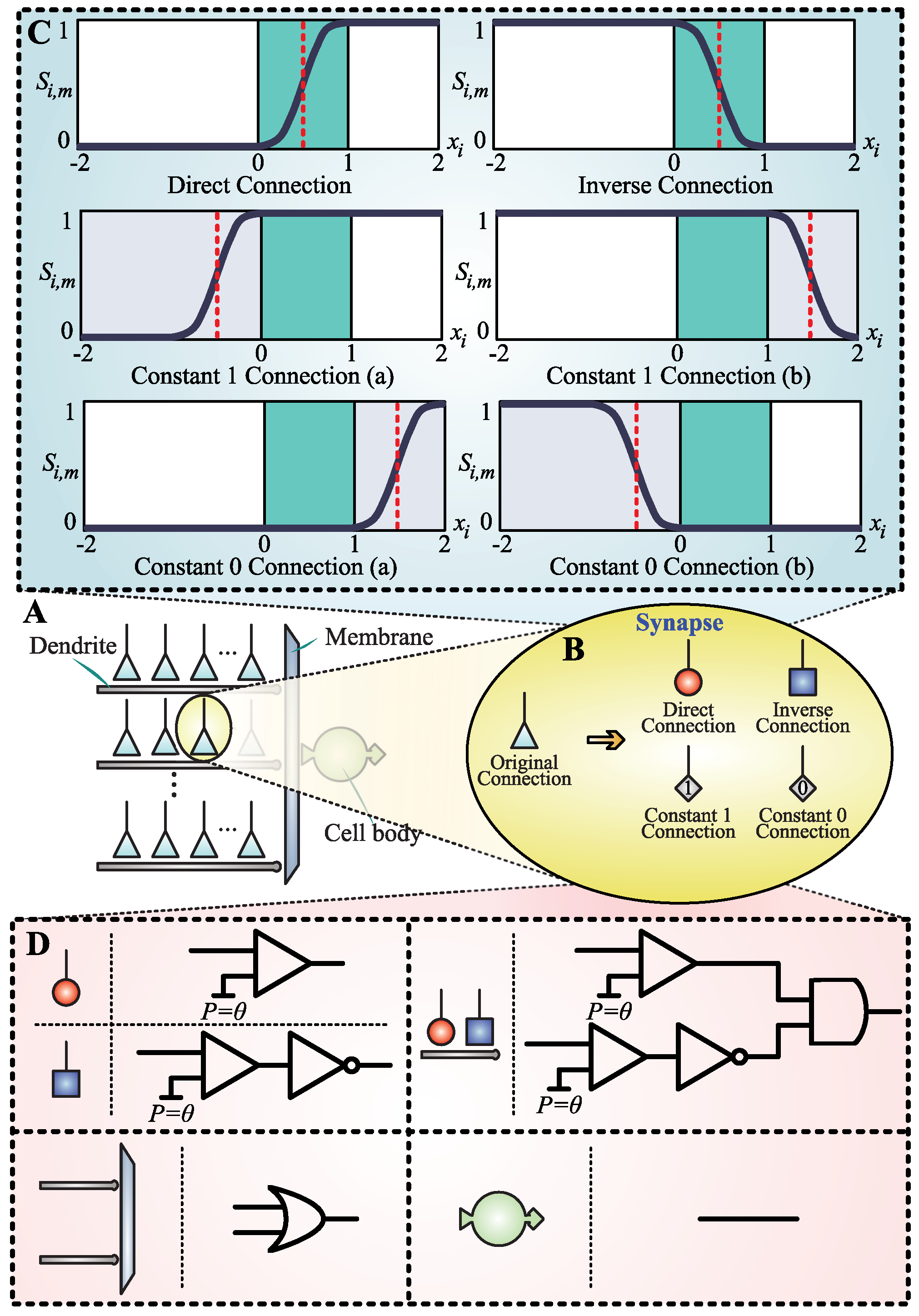
2.1.2. Synapse Evolution Scheme
- Direct connection (, with and .): In the case of a direct connection, the threshold signal and suprathreshold signal can cause an excitatory action potential. An EPSP is generated and delivered to the dendrite of the target neuron, which is approximately 1. Otherwise, the subthreshold signal is represented as an IPSP, and the synapse approximately outputs 0.
- Inverse connection (, with and .): Contrary to the direct connection, an inhibitory action potential is caused by the threshold signal and suprathreshold signal in an inverse connection. It means that the synapse produces an IPSP, and its output is approximately 0. Conversely, the subthreshold causes an excitatory synapse, and an EPSP is triggered. The output of this synapse is close to 1.
- Constant 1 connection (, with and ; , with and .): There are two cases for constant 1 connection. In both cases, the synapse can only output an EPSP regardless of the input signal. It means that the synapse ignores the input and consistently outputs 1.
- Constant 0 connection (, with and ; , with and .): This is the same as the constant 1 connection in that there are also two cases for the constant 0 connection. However, contrary to the constant 1 connection, the subthreshold, threshold, and suprathreshold signals can all cause an IPSP in a constant 0 connection. In other words, no matter what the input signal is, the synapse remains at approximately 0 output.
2.1.3. Neural Pruning Scheme
- Synaptic pruning stage: In the dendritic function, the multiplication operation is used and any value multiplied by 1 is equal to itself. The output of the synapse in the constant 1 connection state is always 1, which suggests that these synapses do not contribute to dendritic function. Therefore, these synapses can be eliminated without affecting the results.
- Dendritic pruning stage: Similar to the synaptic pruning stage, any value multiplied by 0 is equal to 0, and the output of the synapse in the constant 0 connection always remains 0. Thus, even if there is only one synapse in a constant 0 connection state on a dendrite, the other synapses in the entire dendrite can be ignored. The linear summation operation is adopted in the membranous function, which indicates that the entire dendrites have no effect on the membrane. Those dendrites that contain synapses with a constant 0 connection state can be pruned completely.
2.1.4. Hardware Scheme
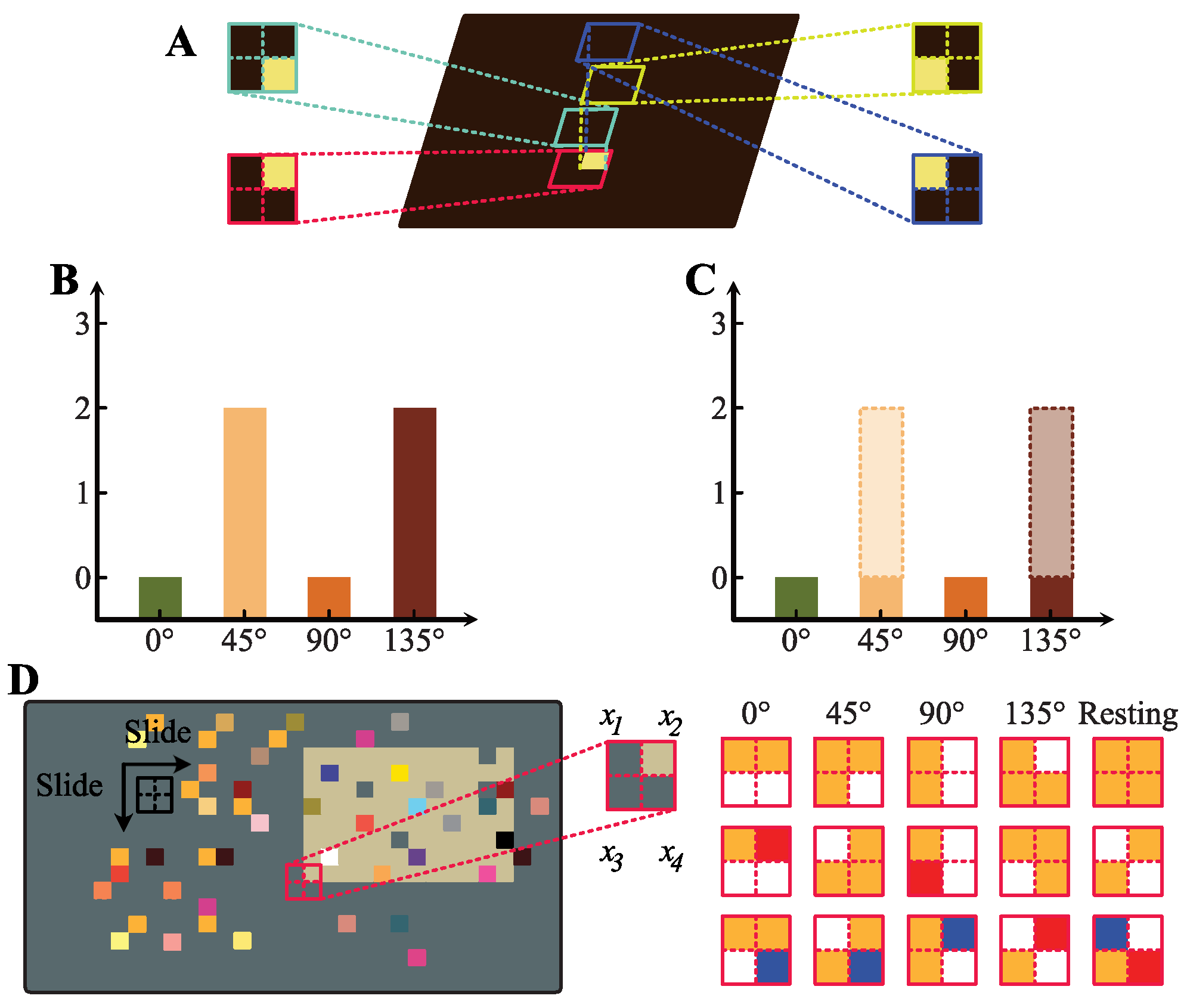
2.2. Orientation Detection Mechanism
3. Experiment and Discussion
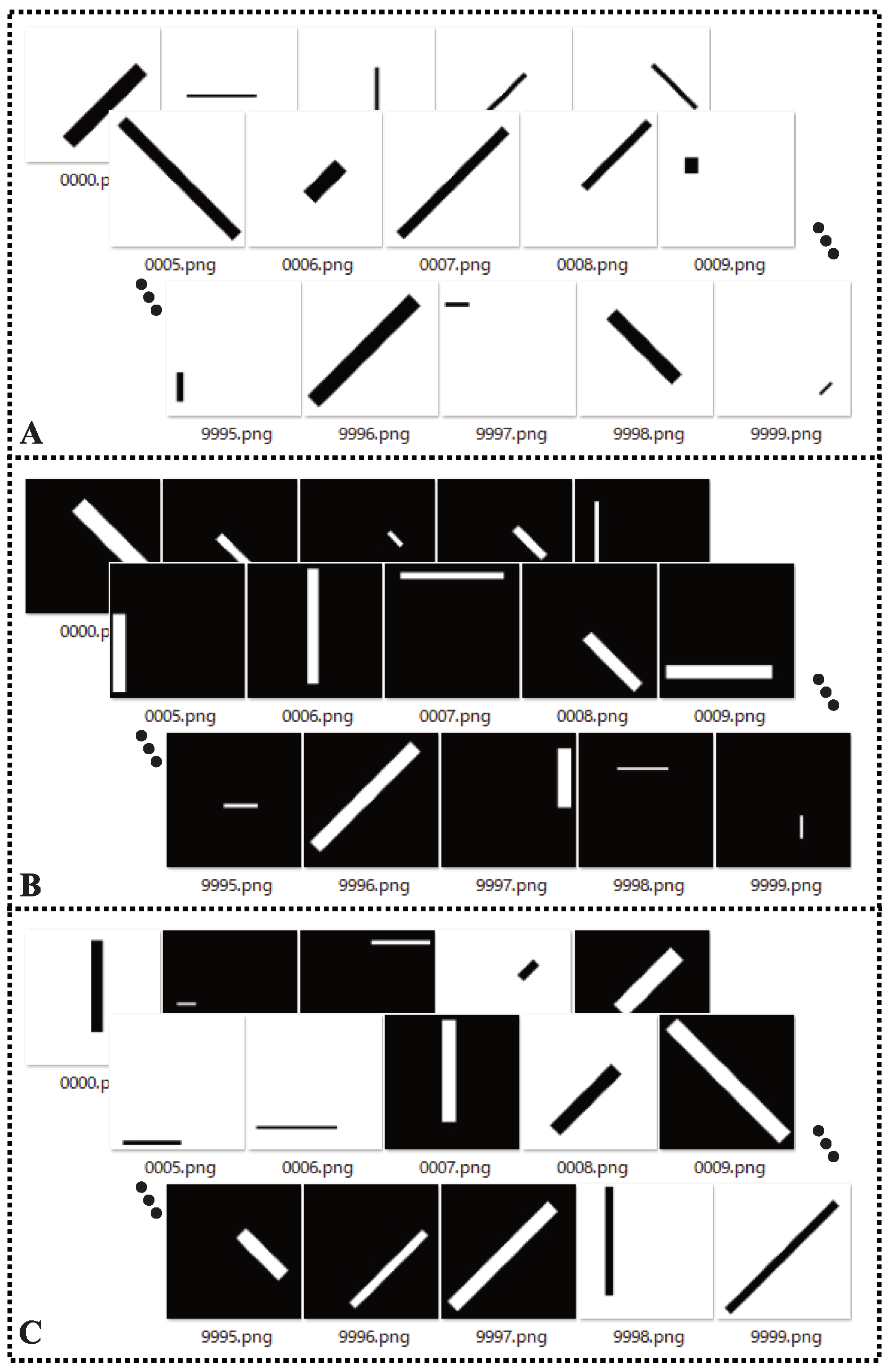
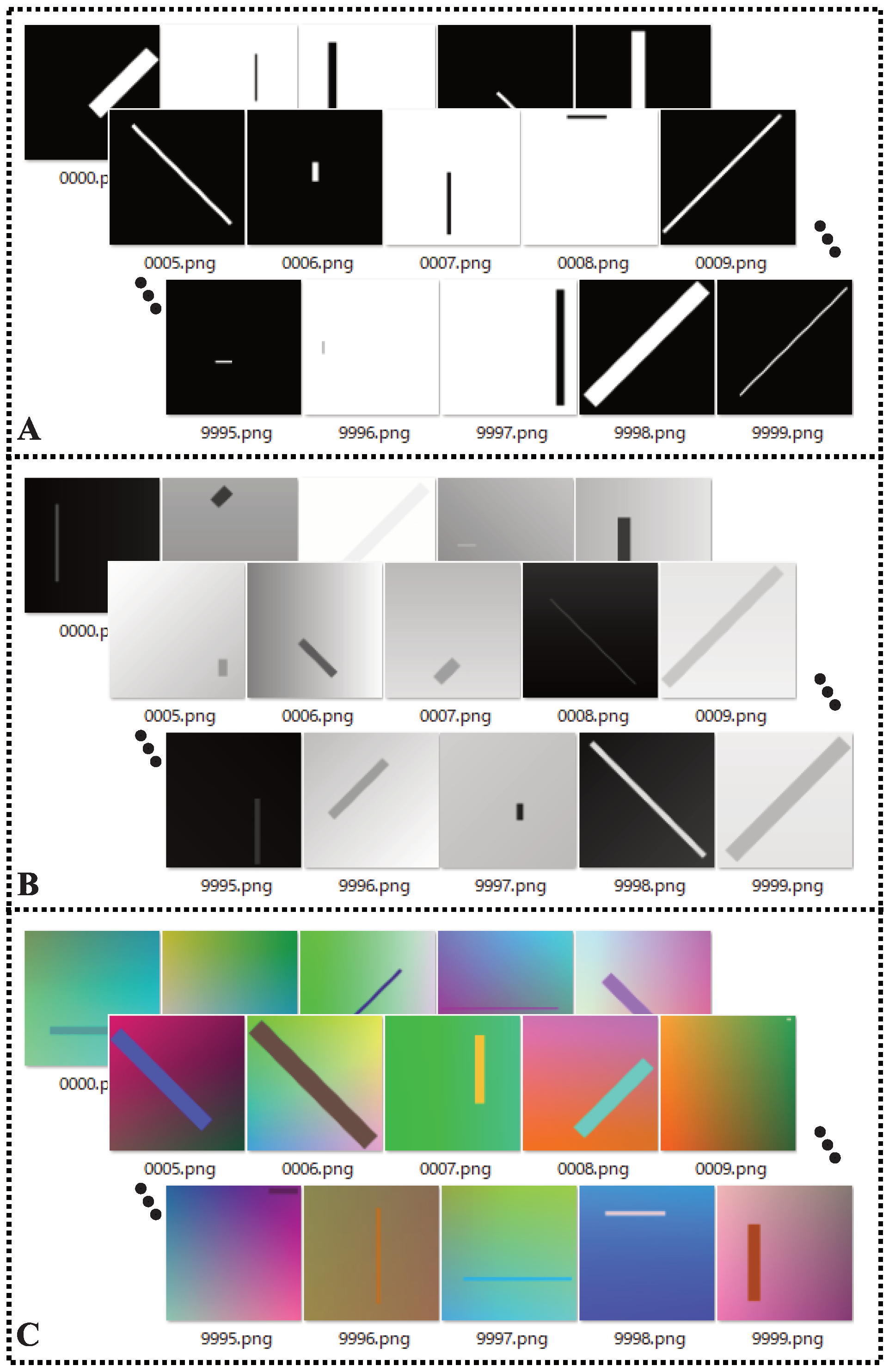
3.1. Experimental Setup
3.2. Performance Evalution Criteria
- Accuracy: This indicates the rate at which the orientations judged by the mechanism match the corresponding target orientations in the datasets. The mean and standard deviation (mean ± std) of the accuracy rates are provided to compare these mechanisms.
- Nonparametric statistical method: Wilcoxon signed rank test is employed as a nonparametric statistical method to determine whether there are significant differences between the mechanisms [26,98]. The p values computed for all the pairwise comparisons are provided, and the level of significance is set to be 0.05.
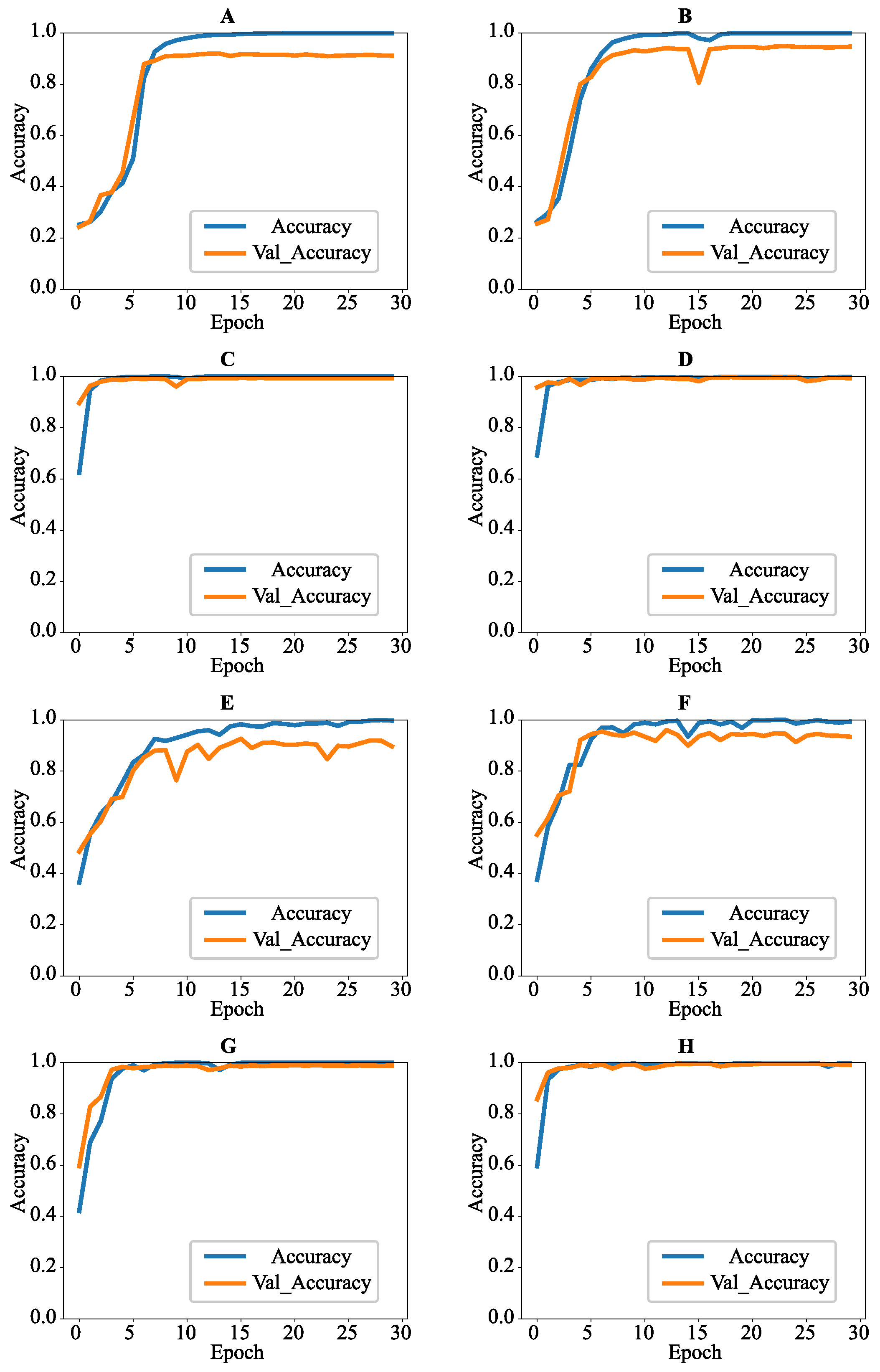
3.3. Comparisons of ODSs
3.4. Comparisons of ODSs and CNNs
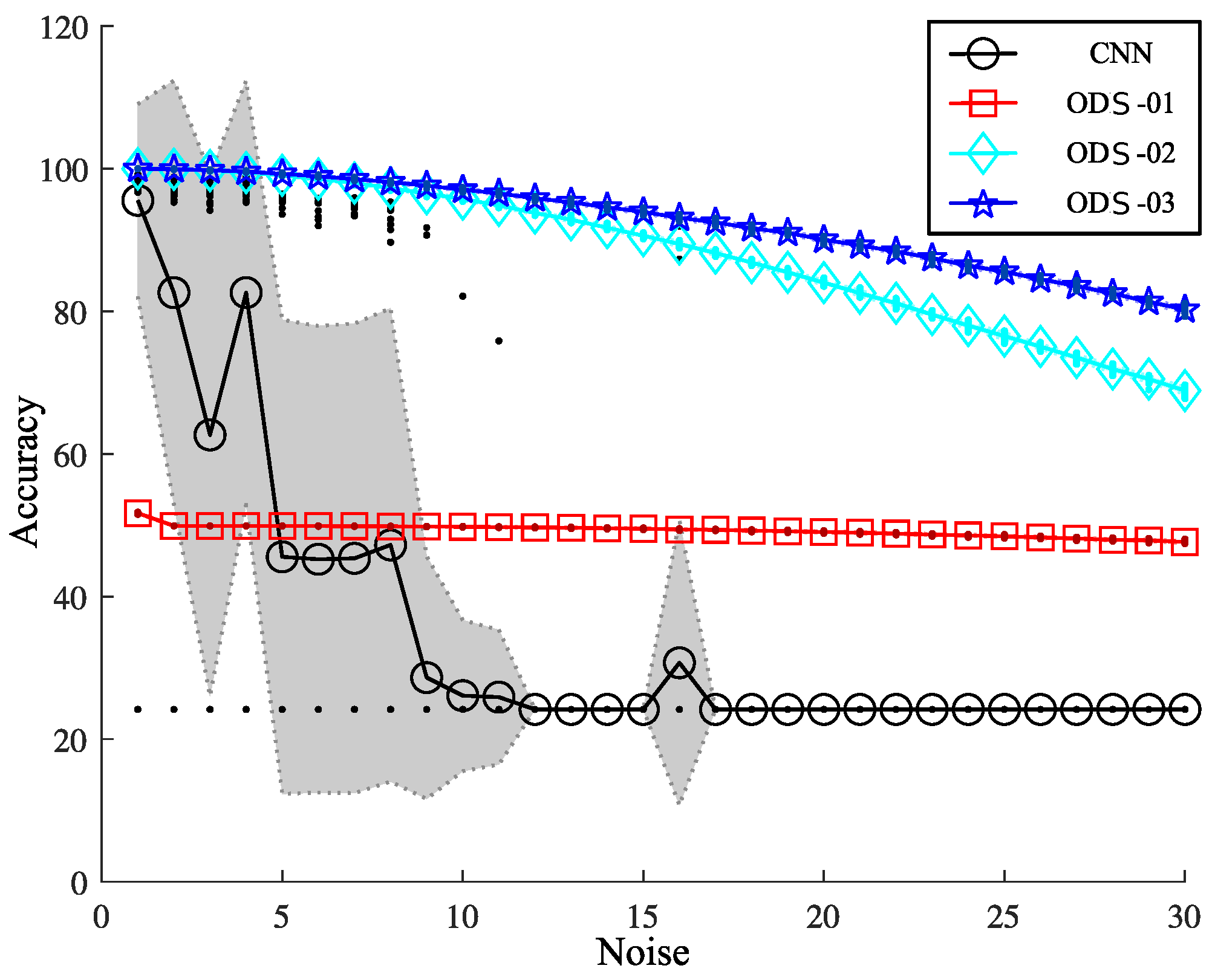
3.5. Performance of ODSs and CNN on Images with Noise
4. Conclusions
- It is precise: The ODSs can make completely correct judgements for all tests without adding noise.
- It is fast: Very little prior knowledge is used to design the mechanism and no iterative learning is required.
- It is flexible: It has no limitation on the size of the image, and the shape, size, and location of the object in the image has no effect on the accuracy.
- It has a simple structure: The ODSs combine several simple DNMs to complete complex tasks. Only the information of the positional relationship among objects in the image is considered.
- Its structure allows parallel computation: All neurons at the same level can operate in parallel, making detection faster by parallel computation.
- Its anti-noise capability is very powerful: The inhibition scheme bestows the ODSs with the ability to resist noise.
- It can be further enhanced: The slightly more refined structural design in this study significantly improved the accuracy of the mechanism. We can foresee that further refinement will make the mechanism even stronger.
- It can be further extended: In this study, a 2 × 2 receptive field is employed to detect four orientations of the objects. More complex interactions among receptive fields or wider receptive fields are considered to extract orientation information at more angles, which will be investigated in our future works.
- It can be easily implemented by hardware: Benefiting from the easy hardware implementation property of the DNM, the ODSs can also be realized on simple devices such as field programmable gate arrays (FPGA). Being free from floating-point computation can further speed up the processing of data.
- It is suitable for big data: The advantages mentioned above enable the ODS to handle explosive high-dimensional data with ease in the era of big data.
- It is highly interpretable: Different from most black-box models in machine learning, the ODS is designed based on prior knowledge and corroborated with studies in physiology, anatomy, and neuroscience. Hence, we encourage researchers from relevant fields to conduct biological experiments to examine the proposed mechanism. When testing with a tiny light spot, the neurons in the corresponding receptive field show mutual inhibition or no response, as assumed, which can further verify the inhibition scheme in the ODSs. Therefore, we believe that the proposed system may provide a new perspective for understanding the relevant brain functions.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Knutsson, H. Filtering and Reconstruction in Image Processing. Ph.D. Thesis, Linköping University, Linköping, Sweden, 1982. [Google Scholar]
- Bigun, J. Optimal Orientation Detection of Linear Symmetry; Linköping University Electronic Press: Linköping, Sweden, 1987. [Google Scholar]
- Schwartz, J.H.; Jessell, T.M.; Kandel, E.R. Principles of Neural Science; Elsevier: New York, NY, USA, 1991. [Google Scholar]
- Squire, L.; Berg, D.; Bloom, F.E.; Du Lac, S.; Ghosh, A.; Spitzer, N.C. Fundamental Neuroscience; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Fiske, S.T.; Taylor, S.E. Social Cognition; Mcgraw-Hill Book Company: New York, NY, USA, 1991. [Google Scholar]
- Sells, S.B.; Fixott, R.S. Evaluation of research on effects of visual training on visual functions. Am. J. Ophthalmol. 1957, 44, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Medina, J. Brain Rules; Pear Press: Seattle, WA, USA, 2016. [Google Scholar]
- Priebe, N.J. Mechanisms of orientation selectivity in the primary visual cortex. Annu. Rev. Vis. Sci. 2016, 2, 85–107. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.; Oehler, R.; Cowey, A. Retinal ganglion cells that project to the dorsal lateral geniculate nucleus in the macaque monkey. Neuroscience 1984, 12, 1101–1123. [Google Scholar] [CrossRef] [PubMed]
- Hubel, D.H.; Wiesel, T.N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol. 1962, 160, 106–154. [Google Scholar] [CrossRef]
- Kandel, E.R.; Schwartz, J.H.; Jessell, T.M.; Siegelbaum, S.; Hudspeth, A.J.; Mack, S. Principles of Neural Science; McGraw-Hill: New York, NY, USA, 2000; Volume 4. [Google Scholar]
- Solomon, S.G.; Lennie, P. The machinery of colour vision. Nat. Rev. Neurosci. 2007, 8, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Shapley, R.; Hawken, M.J. Color in the cortex: Single-and double-opponent cells. Vis. Res. 2011, 51, 701–717. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.K.; Li, P.; Rashid, M.S.; Callaway, E.M. Color and orientation are jointly coded and spatially organized in primate primary visual cortex. Science 2019, 364, 1275–1279. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Schwartz, G.W. Electrical synapses convey orientation selectivity in the mouse retina. Nat. Commun. 2017, 8, 2025. [Google Scholar] [CrossRef]
- Tanaka, K. Cross-correlation analysis of geniculostriate neuronal relationships in cats. J. Neurophysiol. 1983, 49, 1303–1318. [Google Scholar] [CrossRef]
- Tanaka, K. Organization of geniculate inputs to visual cortical cells in the cat. Vis. Res. 1985, 25, 357–364. [Google Scholar] [CrossRef]
- Reid, R.C.; Alonso, J.M. Specificity of monosynaptic connections from thalamus to visual cortex. Nature 1995, 378, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Bear, M.; Connors, B.; Paradiso, M.A. Neuroscience: Exploring the Brain; Jones & Bartlett Learning LLC: Burlington, MA, USA, 2020. [Google Scholar]
- Nagata, F.; Miki, K.; Imahashi, Y.; Nakashima, K.; Tokuno, K.; Otsuka, A.; Watanabe, K.; Habib, M. Orientation Detection Using a CNN Designed by Transfer Learning of AlexNet. In Proceedings of the 8th IIAE International Conference on Industrial Application Engineering 2020, Matsue, Japan, 26–30 March 2020; Volume 5, pp. 26–30. [Google Scholar]
- Gershgorn, D. The data that transformed AI research—And possibly the world. Quartz 2017, 26, 2013–2017. [Google Scholar]
- Krizhevsky, A.; Sutskever, I.; Hinton, G.E. Imagenet classification with deep convolutional neural networks. Adv. Neural Inf. Process. Syst. 2012, 25, 1097–1105. [Google Scholar] [CrossRef]
- Deshpande, A. The 9 Deep Learning Papers You Need to Know about (Understanding CNNs Part 3); University of California (UCLA): Los Angeles, CA, USA, 2018; p. 12–04. [Google Scholar]
- Yang, L.; Qi, Z.; Liu, Z.; Liu, H.; Ling, M.; Shi, L.; Liu, X. An embedded implementation of CNN-based hand detection and orientation estimation algorithm. Mach. Vis. Appl. 2019, 30, 1071–1082. [Google Scholar] [CrossRef]
- Joshi, U.; Guerzhoy, M. Automatic photo orientation detection with convolutional neural networks. In Proceedings of the 2017 14th Conference on Computer and Robot Vision (CRV), Edmonton, AB, Canada, 16–19 May 2017; pp. 103–108. [Google Scholar]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods; John Wiley & Sons: Hoboken, NJ, USA, 2013; Volume 751. [Google Scholar]
- Jiang, Y.; Zhu, X.; Wang, X.; Yang, S.; Li, W.; Wang, H.; Fu, P.; Luo, Z. R2cnn: Rotational region cnn for orientation robust scene text detection. arXiv 2017, arXiv:1706.09579. [Google Scholar]
- Zhao, T. 3D convolutional neural networks for efficient fault detection and orientation estimation. In SEG Technical Program Expanded Abstracts 2019; Society of Exploration Geophysicists: Houston, TX, USA, 2019; pp. 2418–2422. [Google Scholar]
- Guidotti, R.; Monreale, A.; Ruggieri, S.; Turini, F.; Giannotti, F.; Pedreschi, D. A survey of methods for explaining black box models. ACM Comput. Surv. CSUR 2018, 51, 1–42. [Google Scholar] [CrossRef]
- Bilbrey, J.A.; Heindel, J.P.; Schram, M.; Bandyopadhyay, P.; Xantheas, S.S.; Choudhury, S. A look inside the black box: Using graph-theoretical descriptors to interpret a Continuous-Filter Convolutional Neural Network (CF-CNN) trained on the global and local minimum energy structures of neutral water clusters. J. Chem. Phys. 2020, 153, 024302. [Google Scholar] [CrossRef]
- Correia-Silva, J.R.; Berriel, R.F.; Badue, C.; De Souza, A.F.; Oliveira-Santos, T. Copycat CNN: Are random non-Labeled data enough to steal knowledge from black-box models? Pattern Recognit. 2021, 113, 107830. [Google Scholar] [CrossRef]
- Tang, Z.; Tamura, H.; Kuratu, M.; Ishizuka, O.; Tanno, K. A model of the neuron based on dendrite mechanisms. Electron. Commun. Jpn. Part III Fundam. Electron. Sci. 2001, 84, 11–24. [Google Scholar] [CrossRef]
- Tang, Z.; Tamura, H.; Ishizuka, O.; Tanno, K. A neuron model with interaction among synapses. IEEJ Trans. Electron. Inf. Syst. 2000, 120, 1012–1019. [Google Scholar]
- Todo, Y.; Tamura, H.; Yamashita, K.; Tang, Z. Unsupervised learnable neuron model with nonlinear interaction on dendrites. Neural Netw. 2014, 60, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Gao, S.; Cheng, J.; Tang, Z.; Todo, Y. An approximate logic neuron model with a dendritic structure. Neurocomputing 2016, 173, 1775–1783. [Google Scholar] [CrossRef]
- Todo, Y.; Tang, Z.; Todo, H.; Ji, J.; Yamashita, K. Neurons with multiplicative interactions of nonlinear synapses. Int. J. Neural Syst. 2019, 29, 1950012. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Song, S.; Tang, Y.; Gao, S.; Tang, Z.; Todo, Y. Approximate logic neuron model trained by states of matter search algorithm. Knowl.-Based Syst. 2019, 163, 120–130. [Google Scholar] [CrossRef]
- Qian, X.; Tang, C.; Todo, Y.; Lin, Q.; Ji, J. Evolutionary Dendritic Neural Model for Classification Problems. Complexity 2020, 2020, 6296209. [Google Scholar] [CrossRef]
- Song, S.; Chen, X.; Tang, C.; Song, S.; Tang, Z.; Todo, Y. Training an approximate logic dendritic neuron model using social learning particle swarm optimization algorithm. IEEE Access 2019, 7, 141947–141959. [Google Scholar] [CrossRef]
- Ji, J.; Tang, Y.; Ma, L.; Li, J.; Lin, Q.; Tang, Z.; Todo, Y. Accuracy Versus Simplification in an Approximate Logic Neural Model. IEEE Trans. Neural Netw. Learn. Syst. 2020, 32, 5194–5207. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Ji, J.; Tang, Y.; Gao, S.; Tang, Z.; Todo, Y. A novel machine learning technique for computer-aided diagnosis. Eng. Appl. Artif. Intell. 2020, 92, 103627. [Google Scholar] [CrossRef]
- Tang, Y.; Ji, J.; Zhu, Y.; Gao, S.; Tang, Z.; Todo, Y. A differential evolution-oriented pruning neural network model for bankruptcy prediction. Complexity 2019, 2019. [Google Scholar] [CrossRef]
- Song, Z.; Tang, Y.; Ji, J.; Todo, Y. Evaluating a dendritic neuron model for wind speed forecasting. Knowl.-Based Syst. 2020, 201, 106052. [Google Scholar] [CrossRef]
- Song, Z.; Tang, C.; Ji, J.; Todo, Y.; Tang, Z. A Simple Dendritic Neural Network Model-Based Approach for Daily PM2. 5 Concentration Prediction. Electronics 2021, 10, 373. [Google Scholar] [CrossRef]
- Li, B.; Todo, Y.; Tang, Z. Artificial Visual System for Orientation Detection Based on Hubel–Wiesel Model. Brain Sci. 2022, 12, 470. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Todo, Y.; Kobayashi, Y.; Tang, Z.; Li, B. An Artificial Visual System for Motion Direction Detection Based on the Hassenstein–Reichardt Correlator Model. Electronics 2022, 11, 1423. [Google Scholar] [CrossRef]
- Chalupa, L.M.; Williams, R.W. Eye, Retina, and Visual System of the Mouse; Mit Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Brewer, A.A.; Liu, J.; Wade, A.R.; Wandell, B.A. Visual field maps and stimulus selectivity in human ventral occipital cortex. Nat. Neurosci. 2005, 8, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Larsson, J.; Heeger, D.J. Two retinotopic visual areas in human lateral occipital cortex. J. Neurosci. 2006, 26, 13128–13142. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.S. Perceptual adaptation to inverted, reversed, and displaced vision. Psychol. Rev. 1965, 72, 419. [Google Scholar] [CrossRef] [PubMed]
- Tootell, R.B.; Mendola, J.D.; Hadjikhani, N.K.; Ledden, P.J.; Liu, A.K.; Reppas, J.B.; Sereno, M.I.; Dale, A.M. Functional analysis of V3A and related areas in human visual cortex. J. Neurosci. 1997, 17, 7060–7078. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.G. Visual maps in the adult primate cerebral cortex: Some implications for brain development and evolution. Braz. J. Med. Biol. Res. 2002, 35, 1485–1498. [Google Scholar] [CrossRef]
- DeYoe, E.A.; Carman, G.J.; Bandettini, P.; Glickman, S.; Wieser, J.; Cox, R.; Miller, D.; Neitz, J. Mapping striate and extrastriate visual areas in human cerebral cortex. Proc. Natl. Acad. Sci. USA 1996, 93, 2382–2386. [Google Scholar] [CrossRef]
- Engel, S.A.; Glover, G.H.; Wandell, B.A. Retinotopic organization in human visual cortex and the spatial precision of functional MRI. Cereb. Cortex 1997, 7, 181–192. [Google Scholar] [CrossRef]
- Bridge, H. Mapping the visual brain: How and why. Eye 2011, 25, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Wandell, B.A.; Brewer, A.A.; Dougherty, R.F. Visual field map clusters in human cortex. Philos. Trans. R. Soc. Biol. Sci. 2005, 360, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Rajimehr, R.; Tootell, R.B. Does retinotopy influence cortical folding in primate visual cortex? J. Neurosci. 2009, 29, 11149–11152. [Google Scholar] [CrossRef] [PubMed]
- Hubel, D.H.; Wiesel, T.N. Receptive fields of single neurones in the cat’s striate cortex. In Brain Physiology and Psychology; University of California Press: Berkeley, CA, USA, 2020; pp. 129–150. [Google Scholar]
- Hubel, D.H.; Wiesel, T.N. Receptive fields and functional architecture of monkey striate cortex. J. Physiol. 1968, 195, 215–243. [Google Scholar] [CrossRef] [PubMed]
- Beaudet, A.; Descarries, L. The monoamine innervation of rat cerebral cortex: Synaptic and nonsynaptic axon terminals. Neuroscience 1978, 3, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Palay, S.L.; Chan-Palay, V. Cerebellar Cortex: Cytology and Organization; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Scimemi, A.; Beato, M. Determining the neurotransmitter concentration profile at active synapses. Mol. Neurobiol. 2009, 40, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Azouz, R.; Gray, C.M. Dynamic spike threshold reveals a mechanism for synaptic coincidence detection in cortical neurons in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 8110–8115. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.A.; Stempel, A.V.; Vale, R.; Ruehle, S.; Lefler, Y.; Branco, T. A synaptic threshold mechanism for computing escape decisions. Nature 2018, 558, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.; Narayanan, R. Active dendrites: Colorful wings of the mysterious butterflies. Trends Neurosci. 2008, 31, 309–316. [Google Scholar] [CrossRef]
- Peña, J.L.; Konishi, M. Auditory spatial receptive fields created by multiplication. Science 2001, 292, 249–252. [Google Scholar] [CrossRef]
- Gabbiani, F.; Krapp, H.G.; Koch, C.; Laurent, G. Multiplicative computation in a visual neuron sensitive to looming. Nature 2002, 420, 320–324. [Google Scholar] [CrossRef]
- Kurowski, P.; Gawlak, M.; Szulczyk, P. Muscarinic receptor control of pyramidal neuron membrane potential in the medial prefrontal cortex (mPFC) in rats. Neuroscience 2015, 303, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Bean, B.P. The action potential in mammalian central neurons. Nat. Rev. Neurosci. 2007, 8, 451–465. [Google Scholar] [CrossRef]
- Chakraborty, D.; Truong, D.Q.; Bikson, M.; Kaphzan, H. Neuromodulation of axon terminals. Cereb. Cortex 2018, 28, 2786–2794. [Google Scholar] [CrossRef] [PubMed]
- Choquet, D.; Triller, A. The dynamic synapse. Neuron 2013, 80, 691–703. [Google Scholar] [CrossRef]
- Koch, C. Biophysics of Computation: Information Processing in Single Neurons; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Spruston, N. Pyramidal neurons: Dendritic structure and synaptic integration. Nat. Rev. Neurosci. 2008, 9, 206–221. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; O’Leary, D.D. Axon retraction and degeneration in development and disease. Annu. Rev. Neurosci. 2005, 28, 127–156. [Google Scholar] [CrossRef] [PubMed]
- Zollo, M.; Ahmed, M.; Ferrucci, V.; Salpietro, V.; Asadzadeh, F.; Carotenuto, M.; Maroofian, R.; Al-Amri, A.; Singh, R.; Scognamiglio, I.; et al. PRUNE is crucial for normal brain development and mutated in microcephaly with neurodevelopmental impairment. Brain 2017, 140, 940–952. [Google Scholar] [CrossRef]
- Neniskyte, U.; Gross, C.T. Errant gardeners: Glial-cell-dependent synaptic pruning and neurodevelopmental disorders. Nat. Rev. Neurosci. 2017, 18, 658. [Google Scholar] [CrossRef]
- Ecker, A.S.; Berens, P.; Keliris, G.A.; Bethge, M.; Logothetis, N.K.; Tolias, A.S. Decorrelated neuronal firing in cortical microcircuits. Science 2010, 327, 584–587. [Google Scholar] [CrossRef]
- Vaney, D.I.; Sivyer, B.; Taylor, W.R. Direction selectivity in the retina: Symmetry and asymmetry in structure and function. Nat. Rev. Neurosci. 2012, 13, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, G.M.; Brayton, R.K. Logic operations are properties of computer-simulated interactions between excitable dendritic spines. Neuroscience 1987, 21, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Shadlen, M.N.; Newsome, W.T. The variable discharge of cortical neurons: Implications for connectivity, computation, and information coding. J. Neurosci. 1998, 18, 3870–3896. [Google Scholar] [CrossRef] [PubMed]
- Chklovskii, D.B.; Mel, B.; Svoboda, K. Cortical rewiring and information storage. Nature 2004, 431, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Polsky, A.; Mel, B.W.; Schiller, J. Computational subunits in thin dendrites of pyramidal cells. Nat. Neurosci. 2004, 7, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Eyal, G.; Verhoog, M.B.; Testa-Silva, G.; Deitcher, Y.; Benavides-Piccione, R.; DeFelipe, J.; De Kock, C.P.; Mansvelder, H.D.; Segev, I. Human cortical pyramidal neurons: From spines to spikes via models. Front. Cell. Neurosci. 2018, 12, 181. [Google Scholar] [CrossRef] [PubMed]
- Gidon, A.; Zolnik, T.A.; Fidzinski, P.; Bolduan, F.; Papoutsi, A.; Poirazi, P.; Holtkamp, M.; Vida, I.; Larkum, M.E. Dendritic action potentials and computation in human layer 2/3 cortical neurons. Science 2020, 367, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, P.D. Neural Computing: Theory and Practice; Van Nostrand Reinhold Co.: New York, NY, USA, 1989. [Google Scholar]
- Beale, R.; Jackson, T. Neural Computing—An Introduction; CRC Press: Boca Raton, FL, USA, 1990. [Google Scholar]
- Wasserman, P.D. Advanced Methods in Neural Computing; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1993. [Google Scholar]
- Deco, G.; Obradovic, D. An Information-Theoretic Approach to Neural Computing; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Maren, A.J.; Harston, C.T.; Pap, R.M. Handbook of Neural Computing Applications; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Alonso, J.M.; Chen, Y. Receptive field. Scholarpedia 2009, 4, 5393. [Google Scholar] [CrossRef]
- Finn, I.M.; Priebe, N.J.; Ferster, D. The emergence of contrast-invariant orientation tuning in simple cells of cat visual cortex. Neuron 2007, 54, 137–152. [Google Scholar] [CrossRef]
- Hansel, D.; van Vreeswijk, C. The mechanism of orientation selectivity in primary visual cortex without a functional map. J. Neurosci. 2012, 32, 4049–4064. [Google Scholar] [CrossRef]
- Koch, E.; Jin, J.; Wang, Y.; Kremkow, J.; Alonso, J.M.; Zaidi, Q. Cross-orientation suppression and the topography of orientation preferences. J. Vis. 2015, 15, 1000. [Google Scholar] [CrossRef]
- Martinez, L.M.; Wang, Q.; Reid, R.C.; Pillai, C.; Alonso, J.M.; Sommer, F.T.; Hirsch, J.A. Receptive field structure varies with layer in the primary visual cortex. Nat. Neurosci. 2005, 8, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Frégnac, Y.; Bathellier, B. Cortical correlates of low-level perception: From neural circuits to percepts. Neuron 2015, 88, 110–126. [Google Scholar] [CrossRef] [PubMed]
- Sattler, T.; Leibe, B.; Kobbelt, L. Efficient & effective prioritized matching for large-scale image-based localization. IEEE Trans. Pattern Anal. Mach. Intell. 2016, 39, 1744–1756. [Google Scholar] [PubMed]
- Russakovsky, O.; Deng, J.; Su, H.; Krause, J.; Satheesh, S.; Ma, S.; Huang, Z.; Karpathy, A.; Khosla, A.; Bernstein, M.; et al. Imagenet large scale visual recognition challenge. Int. J. Comput. Vis. 2015, 115, 211–252. [Google Scholar] [CrossRef]
- Derrac, J.; García, S.; Molina, D.; Herrera, F. A practical tutorial on the use of nonparametric statistical tests as a methodology for comparing evolutionary and swarm intelligence algorithms. Swarm Evol. Comput. 2011, 1, 3–18. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orientation | |||
|---|---|---|---|
| 0 | 1 | 0 | |
| 1 | 1 | 0 | |
| 0/1 | 0 | 1 | |
| 0/1 | 1 | 1 | |
| Resting | 0/1 | 0 | 0 |
| 100 × 100 pixels | |||
| Datasets | ODS-01 | ODS-02 | ODS-03 |
| Binary-WB | 100.00 | 100.00 | 100.00 |
| Binary-BW | 100.00 | 100.00 | 100.00 |
| Binary-Mix | 100.00 | 100.00 | 100.00 |
| Grey | 100.00 | 100.00 | 100.00 |
| Color | 100.00 | 100.00 | 100.00 |
| 1000 × 1000 pixels | |||
| Datasets | ODS-01 | ODS-02 | ODS-03 |
| Binary-Mix | 100.00 | 100.00 | 100.00 |
| Grey | 100.00 | 100.00 | 100.00 |
| Color | 100.00 | 100.00 | 100.00 |
| 3024 × 3024 pixels | |||
| Datasets | ODS-01 | ODS-02 | ODS-03 |
| Real | 73.21 | 75.00 | 100.00 |
| Datasets | CNN-04 | CNN-30 | ODS-01 | ODS-02 | ODS-03 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean ± Std | p | Mean ± Std | p | Mean ± Std | p | Mean ± Std | p | Mean ± Std | |
| Binary-WB | 93.46 ± 0.69 | 9.07 | 95.23 ± 0.45 | 9.07 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 |
| Binary-BW | 91.89 ± 12.87 | 9.11 | 96.33 ± 0.40 | 9.02 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 |
| Binary-Mix | 94.21 ± 2.83 | 9.09 | 94.08 ± 9.17 | 9.08 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 |
| Grey | 90.87 ± 1.66 | 9.09 | 90.53 ± 2.25 | 9.12 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 |
| Color | 89.17 ± 12.25 | 9.10 | 89.17 ± 12.25 | 9.10 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 | 1.00 | 100.00 ± 0.00 |
| CNNs with different convolutional layers on color image dataset | |||||||||
| Layers-02 | 94.67 ± 13.11 | 9.08 | 95.05 ± 13.16 | 9.04 | - | - | - | - | - |
| Layers-03 | 98.55 ± 1.36 | 9.05 | 98.08 ± 0.93 | 9.06 | - | - | - | - | - |
| Layers-04 | 93.44 ± 15.47 | 9.09 | 98.47 ± 1.19 | 9.04 | - | - | - | - | - |
| Orientation Detection System | Device | Type | Duration |
|---|---|---|---|
| ODS | GPU | NVIDIA Tesla P100 | 3 min 2 s |
| 1-Channel CNN | GPU | NVIDIA Tesla P100 | 4 min 47 s |
| 4-Layer CNN | GPU | NVIDIA Tesla P100 | 5 min 58 s |
| EfN | GPU | NVIDIA Tesla P100 | 29 min 14 s |
| Noise | CNN | ODS-01 | ODS-02 | ODS-03 | |||
|---|---|---|---|---|---|---|---|
| Mean ± Std | p | Mean ± Std | p | Mean ± Std | p | Mean ± Std | |
| 01 | 95.55 ± 13.50 | 9.11 | 51.69 ± 0.08 | 9.04 | 99.99 ± 0.01 | 3.00 | 99.99 ± 0.01 |
| 02 | 82.61 ± 29.73 | 9.12 | 49.90 ± 0.00 | 8.52 | 99.92 ± 0.02 | 2.18 | 99.92 ± 0.03 |
| 03 | 62.65 ± 36.63 | 9.01 | 49.90 ± 0.00 | 8.70 | 99.73 ± 0.04 | 6.84 | 99.78 ± 0.04 |
| 04 | 82.61 ± 29.73 | 9.12 | 49.89 ± 0.01 | 8.93 | 99.44 ± 0.05 | 1.10 | 99.59 ± 0.05 |
| 05 | 45.56 ± 33.25 | 9.08 | 49.89 ± 0.01 | 9.05 | 99.06 ± 0.08 | 1.58 | 99.28 ± 0.08 |
| 06 | 45.22 ± 32.72 | 9.04 | 49.87 ± 0.01 | 9.01 | 98.54 ± 0.11 | 9.09 | 98.92 ± 0.08 |
| 07 | 45.35 ± 32.92 | 9.12 | 49.86 ± 0.02 | 9.06 | 97.99 ± 0.11 | 9.06 | 98.54 ± 0.13 |
| 08 | 47.23 ± 33.19 | 9.09 | 49.83 ± 0.03 | 9.05 | 97.28 ± 0.13 | 9.09 | 98.11 ± 0.10 |
| 09 | 28.63 ± 17.01 | 9.08 | 49.81 ± 0.02 | 9.06 | 96.49 ± 0.15 | 9.09 | 97.61 ± 0.13 |
| 10 | 26.09 ± 10.58 | 9.09 | 49.77 ± 0.03 | 9.10 | 95.73 ± 0.15 | 9.09 | 97.06 ± 0.15 |
| 11 | 25.88 ± 9.44 | 9.11 | 49.73 ± 0.04 | 9.09 | 94.85 ± 0.17 | 9.08 | 96.53 ± 0.16 |
| 12 | 24.16 ± 0.00 | 9.05 | 49.68 ± 0.04 | 9.05 | 93.78 ± 0.18 | 9.09 | 95.85 ± 0.15 |
| 13 | 24.16 ± 0.00 | 9.09 | 49.64 ± 0.05 | 9.08 | 92.84 ± 0.23 | 9.12 | 95.30 ± 0.20 |
| 14 | 24.16 ± 0.00 | 9.10 | 49.56 ± 0.05 | 9.01 | 91.72 ± 0.22 | 9.10 | 94.58 ± 0.21 |
| 15 | 24.16 ± 0.00 | 9.10 | 49.51 ± 0.05 | 9.00 | 90.59 ± 0.20 | 9.10 | 93.94 ± 0.21 |
| 16 | 30.70 ± 19.98 | 9.12 | 49.40 ± 0.07 | 9.12 | 89.43 ± 0.31 | 9.09 | 93.11 ± 0.23 |
| 17 | 24.16 ± 0.00 | 9.11 | 49.35 ± 0.06 | 9.10 | 88.17 ± 0.26 | 9.12 | 92.47 ± 0.25 |
| 18 | 24.16 ± 0.00 | 9.08 | 49.25 ± 0.08 | 9.11 | 86.83 ± 0.25 | 9.09 | 91.65 ± 0.27 |
| 19 | 24.16 ± 0.00 | 9.02 | 49.15 ± 0.09 | 9.04 | 85.46 ± 0.30 | 9.10 | 91.00 ± 0.20 |
| 20 | 24.16 ± 0.00 | 9.08 | 49.04 ± 0.07 | 9.10 | 84.01 ± 0.28 | 9.10 | 90.04 ± 0.28 |
| 21 | 24.16 ± 0.00 | 9.09 | 48.94 ± 0.10 | 9.09 | 82.54 ± 0.34 | 9.10 | 89.23 ± 0.23 |
| 22 | 24.16 ± 0.00 | 9.10 | 48.82 ± 0.10 | 9.08 | 81.13 ± 0.35 | 9.12 | 88.39 ± 0.24 |
| 23 | 24.16 ± 0.00 | 9.12 | 48.68 ± 0.09 | 9.11 | 79.53 ± 0.28 | 9.11 | 87.45 ± 0.29 |
| 24 | 24.16 ± 0.00 | 9.10 | 48.56 ± 0.10 | 9.10 | 77.99 ± 0.39 | 9.11 | 86.39 ± 0.29 |
| 25 | 24.16 ± 0.00 | 9.11 | 48.45 ± 0.13 | 9.09 | 76.52 ± 0.44 | 9.10 | 85.55 ± 0.30 |
| 26 | 24.16 ± 0.00 | 9.12 | 48.28 ± 0.11 | 9.09 | 75.05 ± 0.30 | 9.08 | 84.49 ± 0.25 |
| 27 | 24.16 ± 0.00 | 9.12 | 48.13 ± 0.12 | 9.12 | 73.51 ± 0.38 | 9.12 | 83.54 ± 0.37 |
| 28 | 24.16 ± 0.00 | 9.12 | 47.95 ± 0.11 | 9.11 | 71.90 ± 0.45 | 9.12 | 82.49 ± 0.33 |
| 29 | 24.16 ± 0.00 | 9.10 | 47.87 ± 0.12 | 9.08 | 70.50 ± 0.39 | 9.06 | 81.32 ± 0.27 |
| 30 | 24.16 ± 0.00 | 9.12 | 47.65 ± 0.14 | 9.12 | 68.87 ± 0.47 | 9.11 | 80.25 ± 0.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nomura, M.; Chen, T.; Tang, C.; Todo, Y.; Sun, R.; Li, B.; Tang, Z. A Hardware-Based Orientation Detection System Using Dendritic Computation. Electronics 2024, 13, 1367. https://doi.org/10.3390/electronics13071367
Nomura M, Chen T, Tang C, Todo Y, Sun R, Li B, Tang Z. A Hardware-Based Orientation Detection System Using Dendritic Computation. Electronics. 2024; 13(7):1367. https://doi.org/10.3390/electronics13071367
Chicago/Turabian StyleNomura, Masahiro, Tianqi Chen, Cheng Tang, Yuki Todo, Rong Sun, Bin Li, and Zheng Tang. 2024. "A Hardware-Based Orientation Detection System Using Dendritic Computation" Electronics 13, no. 7: 1367. https://doi.org/10.3390/electronics13071367