Trimethylornithine Membrane Lipids: Discovered in Planctomycetes and Identified in Diverse Environments


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction—Amino Acid Containing Membrane Lipids
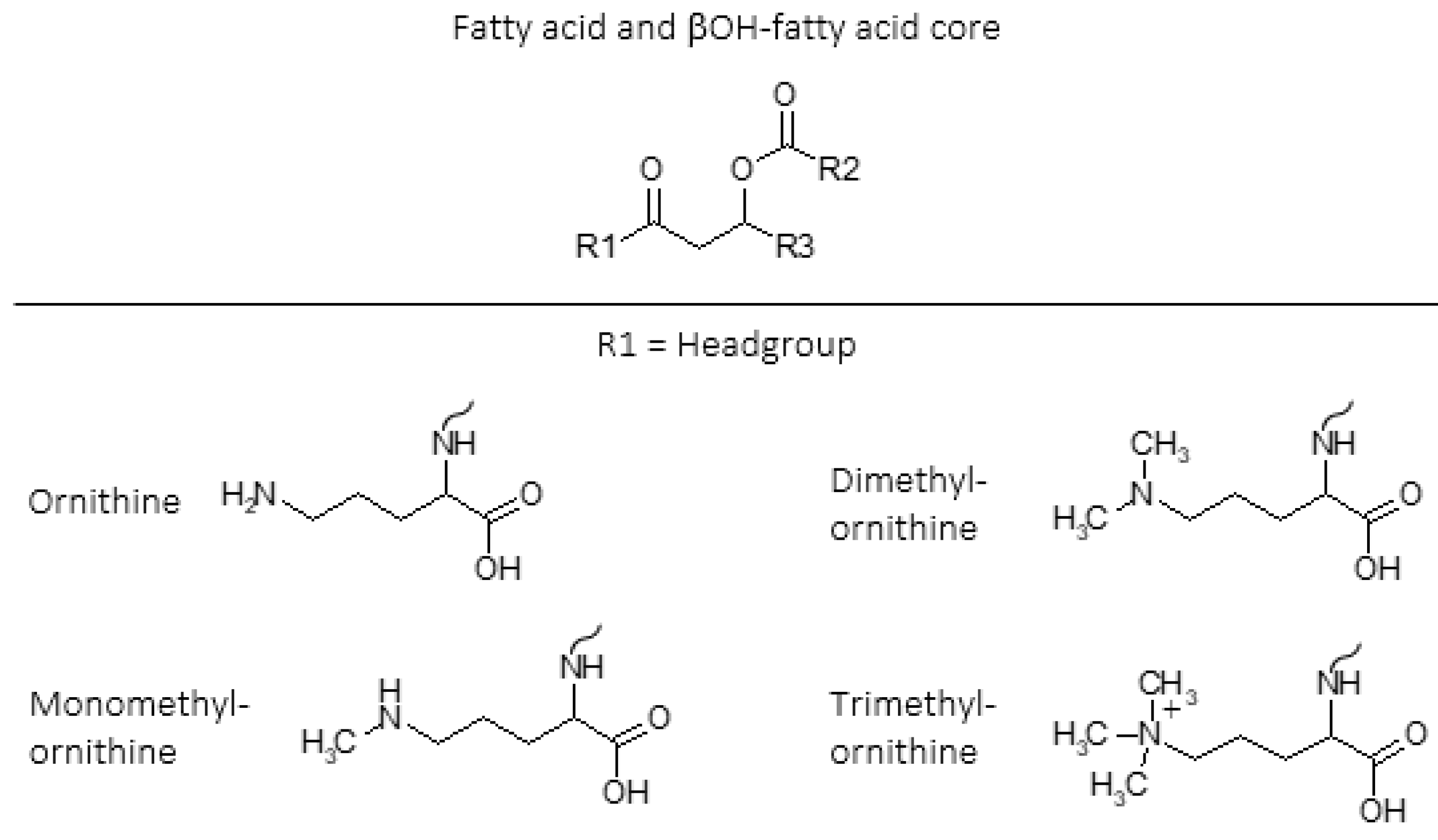
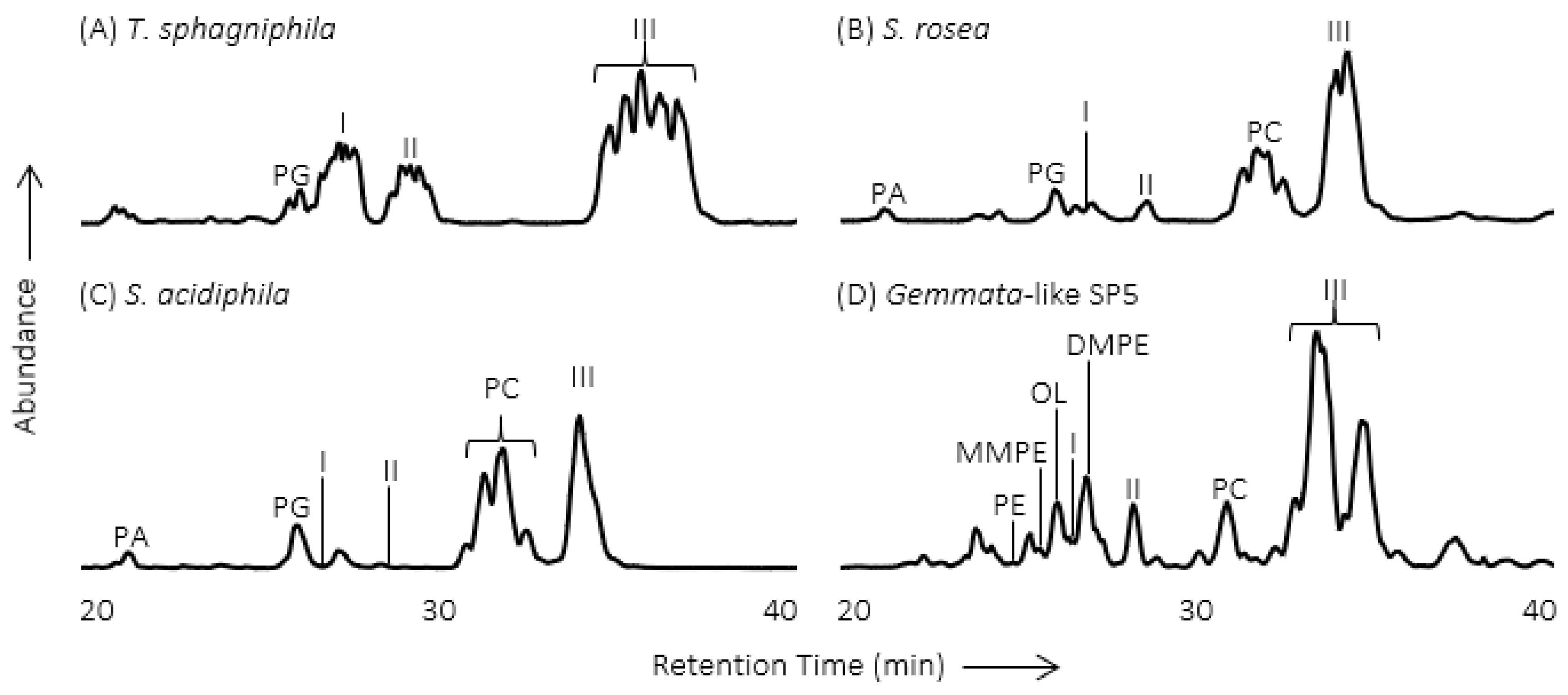
2. Methylated Ornithine Lipids Discovered in Northern Wetland Planctomycetes
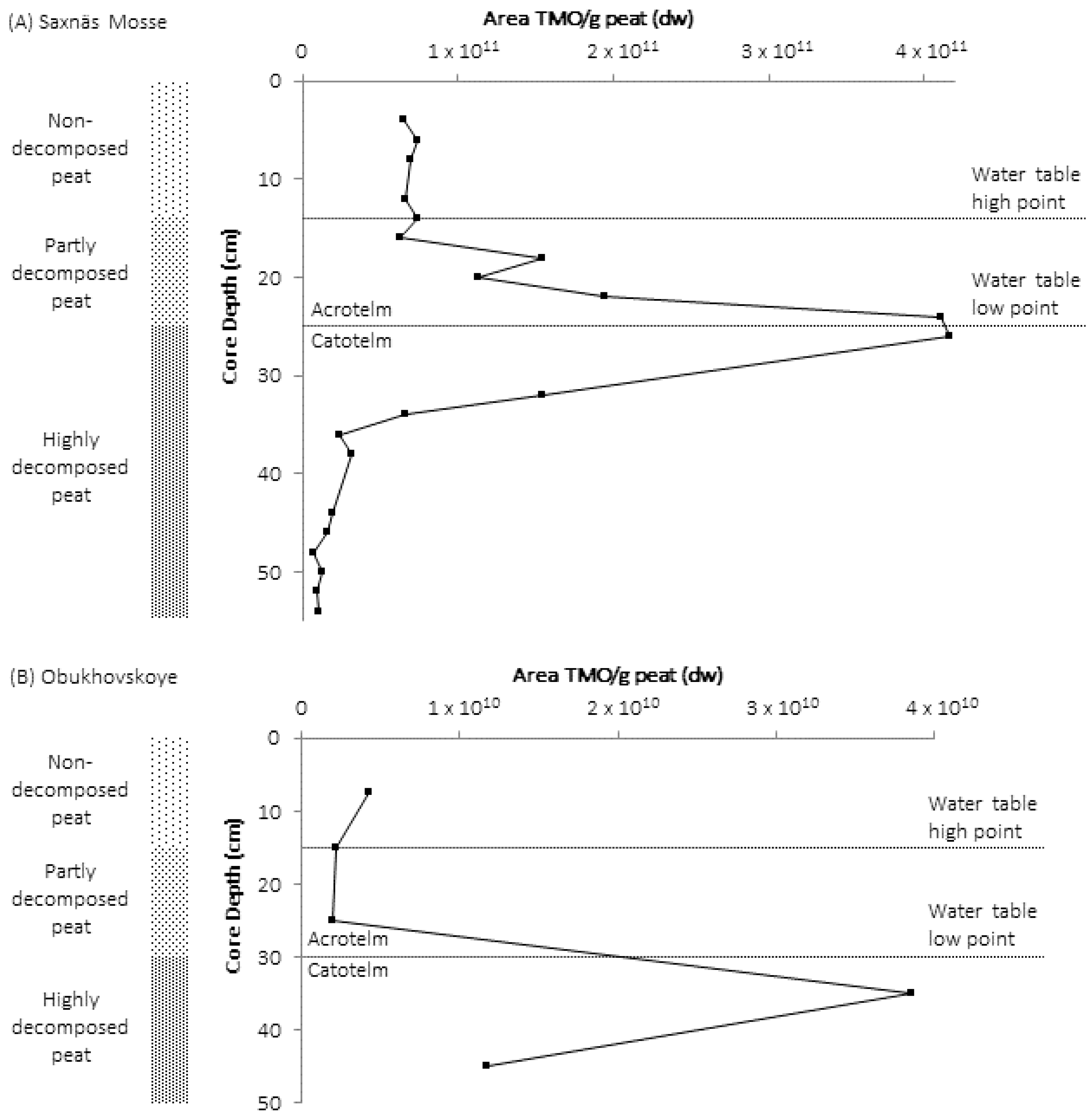
3. High Abundance of TMOs at the Oxic/Anoxic Interface of Northern Wetlands
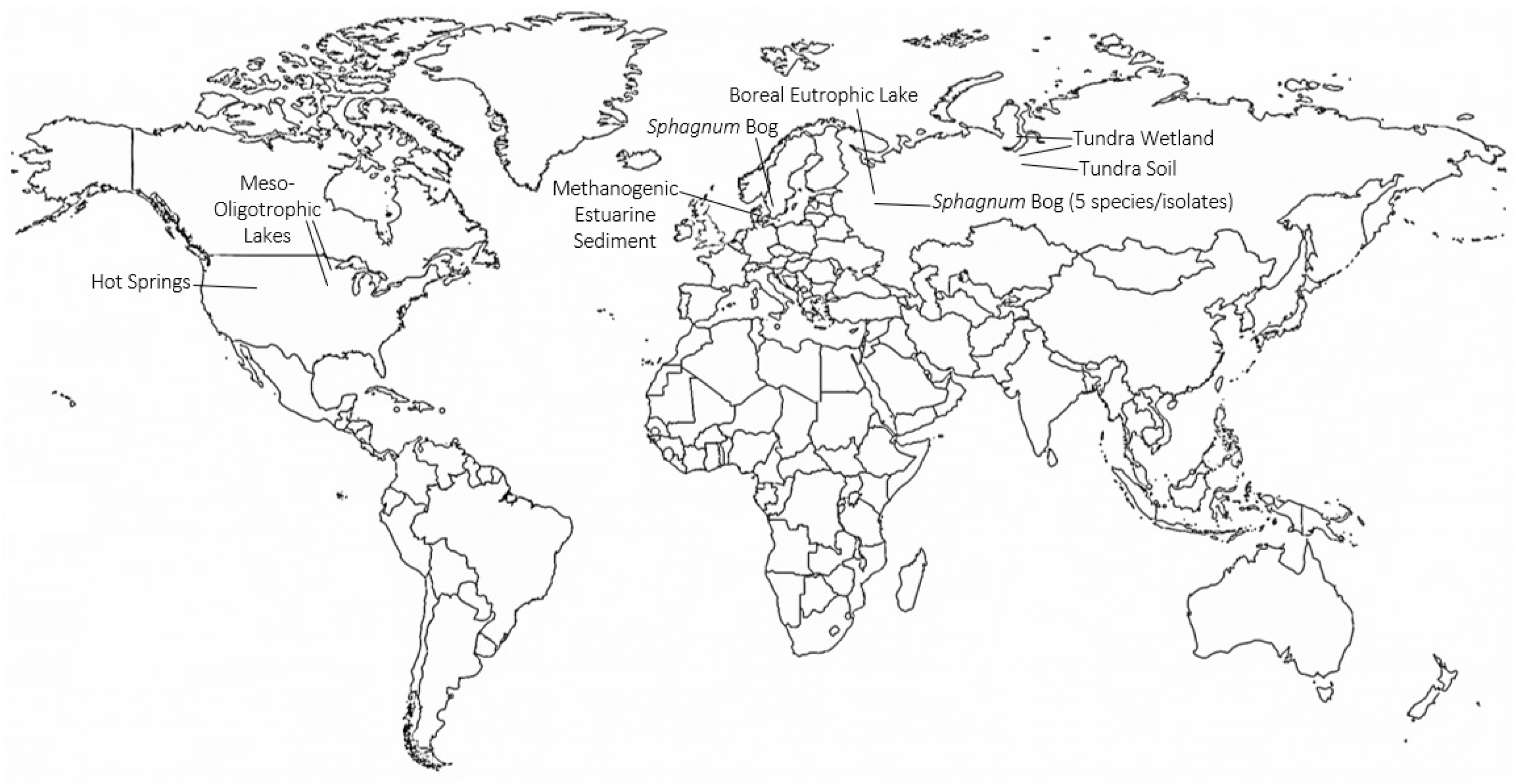
4. Identification of TMOs in Diverse Environments
5. TMOs: Specialized Lipids with Potential Broad Distribution
Funding
Acknowledgments
Conflicts of Interest
References
- Cooper, G.M. The Cell: A Molecular Approach, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2000; ISBN 978-0-87893-106-4. [Google Scholar]
- Sturt, H.F.; Summons, R.E.; Smith, K.; Elvert, M.; Hinrichs, K.-U. Intact polar membrane lipids in prokaryotes and sediments deciphered by high-performance liquid chromatography/electrospray ionization multistage mass spectrometry—New biomarkers for biogeochemistry and microbial ecology. Rapid Commun. Mass Spectrom. 2004, 18, 617–628. [Google Scholar] [CrossRef]
- Schubotz, F.; Wakeham, S.G.; Lipp, J.S.; Fredricks, H.F.; Hinrichs, K.-U. Detection of microbial biomass by intact polar membrane lipid analysis in the water column and surface sediments of the Black Sea. Environ. Microbiol. 2009, 11, 2720–2734. [Google Scholar] [CrossRef] [PubMed]
- White, D.C.; Davis, W.M.; Nickels, J.S.; King, J.D.; Bobbie, R.J. Determination of the sedimentary microbial biomass by extractible lipid phosphate. Oecologia 1979, 40, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Harvey, H.R.; Fallon, R.D.; Patton, J.S. The effect of organic matter and oxygen on the degradation of bacterial membrane lipids in marine sediments. Geochim. Cosmochim. Acta 1986, 50, 795–804. [Google Scholar] [CrossRef]
- Cevc, G. (Ed.) Phospholipids Handbook; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Geiger, O.; González-Silva, N.; López-Lara, I.M.; Sohlenkamp, C. Amino acid-containing membrane lipids in bacteria. Prog. Lipid Res. 2010, 49, 46–60. [Google Scholar] [CrossRef]
- López-Lara, I.M.; Sohlenkamp, C.; Geiger, O. Membrane Lipids in Plant-Associated Bacteria: Their Biosyntheses and Possible Functions. Mol. Plant-Microbe Interact. 2003, 16, 567–579. [Google Scholar] [CrossRef]
- Vences-Guzmán, M.Á.; Geiger, O.; Sohlenkamp, C. Ornithine lipids and their structural modifications: From A to E and beyond. FEMS Microbiol. Lett. 2012, 335, 1–10. [Google Scholar] [CrossRef]
- Vences-Guzmán, M.Á.; Guan, Z.; Escobedo-Hinojosa, W.I.; Bermúdez-Barrientos, J.R.; Geiger, O.; Sohlenkamp, C. Discovery of a bifunctional acyltransferase responsible for ornithine lipid synthesis in Serratia proteamaculans. Environ. Microbiol. 2015, 17, 1487–1496. [Google Scholar] [CrossRef]
- Tahara, Y.; Yamada, Y.; Kondo, K. A New Lysine-containing Lipid Isolated from Agrobacterium tumefaciens. Agric. Biol. Chem. 1976, 40, 1449–1450. [Google Scholar] [CrossRef]
- Moore, E.K.; Hopmans, E.C.; Rijpstra, W.I.C.; Sanchez Andrea, I.; Villanueva, L.; Wienk, H.; Schoutsen, F.; Stams, A.; Sinninghe Damste, J. Lysine and novel hydroxylysine lipids in soil bacteria: Amino acid membrane lipid response to temperature and pH in Pseudopedobacter saltans. Front. Microbiol. 2015, 6, 637. [Google Scholar] [CrossRef]
- Hilker, D.R.; Gross, M.L.; Knocke, H.W.; Shively, J.M. The interpretation of the mass spectrum of an ornithine-containing lipid from Thiobacillus thiooxidans. Biomed. Mass Spectrom. 1978, 5, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Cerny, R.L.; Tomer, K.B.; Gross, M.L. Desorption ionization combined with tandem mass spectrometry: Advantages for investigating complex lipids, disaccharides and organometallic complexes. Org. Mass Spectrom. 1986, 21, 655–660. [Google Scholar] [CrossRef]
- Zhang, X.; Ferguson-Miller, S.M.; Reid, G.E. Characterization of ornithine and glutamine lipids extracted from cell membranes of Rhodobacter sphaeroides. J. Am. Soc. Mass Spectrom. 2009, 20, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.K.; Hopmans, E.C.; Rijpstra, W.I.C.; Villanueva, L.; Dedysh, S.N.; Kulichevskaya, I.S.; Wienk, H.; Schoutsen, F.; Damsté, J.S.S. Novel Mono-, Di-, and Trimethylornithine Membrane Lipids in Northern Wetland Planctomycetes. Appl. Environ. Microbiol. 2013, 79, 6874–6884. [Google Scholar] [CrossRef]
- Moore, E.K.; Hopmans, E.C.; Rijpstra, W.I.C.; Villanueva, L.; Damsté, J.S.S. Elucidation and identification of amino acid containing membrane lipids using liquid chromatography/high-resolution mass spectrometry. Rapid Commun. Mass Spectrom. 2016, 30, 739–750. [Google Scholar] [CrossRef]
- Weissenmayer, B.; Gao, J.-L.; López-Lara, I.M.; Geiger, O. Identification of a gene required for the biosynthesis of ornithine-derived lipids. Mol. Microbiol. 2002, 45, 721–733. [Google Scholar] [CrossRef]
- Gao, J.-L.; Weissenmayer, B.; Taylor, A.M.; Thomas-Oates, J.; López-Lara, I.M.; Geiger, O. Identification of a gene required for the formation of lyso-ornithine lipid, an intermediate in the biosynthesis of ornithine-containing lipids. Mol. Microbiol. 2004, 53, 1757–1770. [Google Scholar] [CrossRef]
- Vences-Guzmán, M.Á.; Guan, Z.; Ormeño-Orrillo, E.; González-Silva, N.; López-Lara, I.M.; Martínez-Romero, E.; Geiger, O.; Sohlenkamp, C. Hydroxylated ornithine lipids increase stress tolerance in Rhizobium tropici CIAT899. Mol. Microbiol. 2011, 79, 1496–1514. [Google Scholar] [CrossRef]
- González-Silva, N.; López-Lara, I.M.; Reyes-Lamothe, R.; Taylor, A.M.; Sumpton, D.; Thomas-Oates, J.; Geiger, O. The Dioxygenase-Encoding olsD Gene from Burkholderia cenocepacia Causes the Hydroxylation of the Amide-Linked Fatty Acyl Moiety of Ornithine-Containing Membrane Lipids. Biochemistry 2011, 50, 6396–6408. [Google Scholar] [CrossRef]
- Gibbons, H.S.; Lin, S.; Cotter, R.J.; Raetz, C.R.H. Oxygen Requirement for the Biosynthesis of theS-2-Hydroxymyristate Moiety in Salmonella typhimurium Lipid a function of LpxO, a new Fe2+/α-Ketoglutarate-Dependent Dioxygenase Homologue. J. Biol. Chem. 2000, 275, 32940–32949. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage. Soil Biol. Biochem. 1995, 27, 753–760. [Google Scholar] [CrossRef]
- Biasi, C.; Rusalimova, O.; Meyer, H.; Kaiser, C.; Wanek, W.; Barsukov, P.; Junger, H.; Richter, A. Temperature-dependent shift from labile to recalcitrant carbon sources of arctic heterotrophs. Rapid Commun. Mass Spectrom. 2005, 19, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Dorrepaal, E.; Toet, S.; van Logtestijn, R.S.P.; Swart, E.; van de Weg, M.J.; Callaghan, T.V.; Aerts, R. Carbon respiration from subsurface peat accelerated by climate warming in the subarctic. Nature 2009, 460, 616–619. [Google Scholar] [CrossRef]
- Lai, D.Y.F. Methane Dynamics in Northern Peatlands: A Review. Pedosphere 2009, 19, 409–421. [Google Scholar] [CrossRef]
- Gorham, E. Northern Peatlands: Role in the Carbon Cycle and Probable Responses to Climatic Warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef]
- Bain, C.G.; Bonn, A.; Stoneman, R.; Chapman, S.; Coupar, A.; Evans, M.; Gearey, B.; Howat, M.; Joosten, H.; Keenleyside, C.; et al. IUCN UK Commission of Inquiry on Peatlands; IUCN UK Peatland Programme: Edinburgh, UK, 2011. [Google Scholar]
- Sundh, I.; Nilsson, M.; Granberg, G.; Svensson, B.H. Depth distribution of microbial production and oxidation of methane in northern boreal peatlands. Microb. Ecol. 1994, 27, 253–265. [Google Scholar] [CrossRef]
- Basiliko, N.; Henry, K.; Gupta, V.; Moore, T.; Driscoll, B.; Dunfield, P. Controls on bacterial and archaeal community structure and greenhouse gas production in natural, mined, and restored Canadian peatlands. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef]
- McCalley, C.K.; Woodcroft, B.J.; Hodgkins, S.B.; Wehr, R.A.; Kim, E.-H.; Mondav, R.; Crill, P.M.; Chanton, J.P.; Rich, V.I.; Tyson, G.W.; et al. Methane dynamics regulated by microbial community response to permafrost thaw. Nature 2014, 514, 478–481. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Pankratov, T.A.; Dedysh, S.N. Detection of representatives of the Planctomycetes in Sphagnum peat bogs by molecular and cultivation approaches. Microbiology 2006, 75, 329–335. [Google Scholar] [CrossRef]
- Dedysh, S.N. Cultivating Uncultured Bacteria from Northern Wetlands: Knowledge Gained and Remaining Gaps. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef]
- Serkebaeva, Y.M.; Kim, Y.; Liesack, W.; Dedysh, S.N. Pyrosequencing-Based Assessment of the Bacteria Diversity in Surface and Subsurface Peat Layers of a Northern Wetland, with Focus on Poorly Studied Phyla and Candidate Divisions. PLoS ONE 2013, 8, e63994. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Damsté, J.S.S. Acidobacteria. In eLS; American Cancer Society: Atlanta, GA, USA, 2018; pp. 1–10. ISBN 978-0-470-01590-2. [Google Scholar]
- Dedysh, S.N.; Ivanova, A.A. Planctomycetes in boreal and subarctic wetlands: Diversity patterns and potential ecological functions. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef] [PubMed]
- Kulichevskaia, I.S.; Belova, S.E.; Kevbrin, V.V.; Dedysh, S.N.; Zavarzin, G.A. Analysis of the bacterial community developing in the course of Sphagnum moss decomposition. Mikrobiologiia 2007, 76, 702–710. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Ivanova, A.O. Abundance, Diversity, and Depth Distribution of Planctomycetes in Acidic Northern Wetlands. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef]
- Bremer, J.; Greenberg, D.M. Methyl transfering enzyme system of microsomes in the biosynthesis of lecithin (phosphatidylcholine). Biochim. Biophys. Acta 1961, 46, 205–216. [Google Scholar] [CrossRef]
- Yamashita, S.; Oshima, A.; Nikawa, J.; Hosaka, K. Regulation of the phosphatidylethanolamine methylation pathway in Saccharomyces cerevisiae. Eur. J. Biochem. 1982, 128, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Gaynor, P.M.; Gill, T.; Toutenhoofd, S.; Summers, E.F.; McGraw, P.; Homann, M.J.; Henry, S.A.; Carman, G.M. Regulation of phosphatidylethanolamine methyltransferase and phospholipid methyltransferase by phospholipid precursors in Saccharomyces cerevisiae. Biochim. Biophys. Acta Gene Struct. Expr. 1991, 1090, 326–332. [Google Scholar] [CrossRef]
- Escobedo-Hinojosa, W.I.; Vences-Guzmán, M.Á.; Schubotz, F.; Sandoval-Calderón, M.; Summons, R.E.; López-Lara, I.M.; Geiger, O.; Sohlenkamp, C. OlsG (Sinac_1600) Is an Ornithine LipidN-Methyltransferase from the PlanctomyceteSingulisphaera acidiphila. J. Biol. Chem. 2015, 290, 15102–15111. [Google Scholar] [CrossRef]
- Freer, E.; Moreno, E.; Moriyón, I.; Pizarro-Cerdá, J.; Weintraub, A.; Gorvel, J.P. Brucella-Salmonella lipopolysaccharide chimeras are less permeable to hydrophobic probes and more sensitive to cationic peptides and EDTA than are their native Brucella sp. counterparts. J. Bacteriol. 1996, 178, 5867–5876. [Google Scholar] [CrossRef]
- Pankratov, T.A.; Ivanova, A.O.; Dedysh, S.N.; Liesack, W. Bacterial populations and environmental factors controlling cellulose degradation in an acidic Sphagnum peat. Environ. Microbiol. 2011, 13, 1800–1814. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Rütters, H.; Sass, H.; Cypionka, H.; Rullkötter, J. Phospholipid analysis as a tool to study complex microbial communities in marine sediments. J. Microbiol. Methods 2002, 48, 149–160. [Google Scholar] [CrossRef]
- Weijers, J.W.H.; Schouten, S.; van der Linden, M.; van Geel, B.; Sinninghe Damsté, J.S. Water table related variations in the abundance of intact archaeal membrane lipids in a Swedish peat bog. FEMS Microbiol. Lett. 2004, 239, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Peterse, F.; Hopmans, E.C.; Schouten, S.; Mets, A.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S. Identification and distribution of intact polar branched tetraether lipids in peat and soil. Org. Geochem. 2011, 42, 1007–1015. [Google Scholar] [CrossRef]
- Moore, E.K.; Villanueva, L.; Hopmans, E.C.; Rijpstra, W.I.C.; Mets, A.; Dedysh, S.N.; Damsté, J.S.S. Abundant Trimethylornithine Lipids and Specific Gene Sequences Are Indicative of Planctomycete Importance at the Oxic/Anoxic Interface in Sphagnum-Dominated Northern Wetlands. Appl. Environ. Microbiol. 2015, 81, 6333–6344. [Google Scholar] [CrossRef]
- Roslev, P.; King, G.M. Regulation of methane oxidation in a freshwater wetland by water table changes and anoxia. FEMS Microbiol. Ecol. 1996, 19, 105–115. [Google Scholar] [CrossRef]
- Angel, R.; Claus, P.; Conrad, R. Methanogenic archaea are globally ubiquitous in aerated soils and become active under wet anoxic conditions. ISME J. 2012, 6, 847–862. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Ivanova, A.A.; Suzina, N.E.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S.; Dedysh, S.N. Paludisphaera borealis gen. nov., sp. nov., a hydrolytic planctomycete from northern wetlands, and proposal of Isosphaeraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 837–844. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Ivanova, A.A.; Detkova, E.N.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S.; Dedysh, S.N. Tundrisphaera lichenicola gen. nov., sp. nov., a psychrotolerant representative of the family Isosphaeraceae from lichen-dominated tundra soils. Int. J. Syst. Evol. Microbiol. 2017, 67, 3583–3589. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Ivanova, A.A.; Baulina, O.I.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S.; Dedysh, S.N. Fimbriiglobus ruber gen. nov., sp. nov., a Gemmata-like planctomycete from Sphagnum peat bog and the proposal of Gemmataceae fam. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 218–224. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Naumoff, D.G.; Miroshnikov, K.K.; Ivanova, A.A.; Philippov, D.A.; Hakobyan, A.; Rijpstra, W.I.C.; Damsté, J.S.S.; Liesack, W.; Dedysh, S.N. Limnoglobus roseus gen. nov., sp. nov., a novel freshwater planctomycete with a giant genome from the family Gemmataceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Kulichevskaya, I.S.; Ivanova, A.A.; Naumoff, D.G.; Beletsky, A.V.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S.; Mardanov, A.V.; Ravin, N.V.; Dedysh, S.N. Frigoriglobus tundricola gen. nov., sp. nov., a psychrotolerant cellulolytic planctomycete of the family Gemmataceae from a littoral tundra wetland. Syst. Appl. Microbiol. 2020, 43, 126129. [Google Scholar] [CrossRef] [PubMed]
- Bale, N.J.; Hopmans, E.C.; Schoon, P.L.; Kluijver, A.d.; Downing, J.A.; Middelburg, J.J.; Damsté, J.S.S.; Schouten, S. Impact of trophic state on the distribution of intact polar lipids in surface waters of lakes. Limnol. Oceanogr. 2016, 61, 1065–1077. [Google Scholar] [CrossRef]
- Ozuolmez, D.; Moore, E.K.; Hopmans, E.C.; Sinninghe Damsté, J.S.; Stams, A.J.M.; Plugge, C.M. Butyrate Conversion by Sulfate-Reducing and Methanogenic Communities from Anoxic Sediments of Aarhus Bay, Denmark. Microorganisms 2020, 8, 606. [Google Scholar] [CrossRef]
- Boyer, G.M.; Schubotz, F.; Summons, R.E.; Woods, J.; Shock, E.L. Carbon Oxidation State in Microbial Polar Lipids Suggests Adaptation to Hot Spring Temperature and Redox Gradients. Front. Microbiol. 2020, 11, 229. [Google Scholar] [CrossRef]
- Wörmer, L.; Gajendra, N.; Schubotz, F.; Matys, E.D.; Evans, T.W.; Summons, R.E.; Hinrichs, K.-U. A micrometer-scale snapshot on phototroph spatial distributions: mass spectrometry imaging of microbial mats in Octopus Spring, Yellowstone National Park. Geobiology 2020, 18, 742–759. [Google Scholar] [CrossRef]
- Jauhiainen, J.; Takahashi, H.; Heikkinen, J.E.P.; Martikainen, P.J.; Vasander, H. Carbon fluxes from a tropical peat swamp forest floor. Glob. Chang. Biol. 2005, 11, 1788–1797. [Google Scholar] [CrossRef]
- Hirano, T.; Segah, H.; Kusin, K.; Limin, S.; Takahashi, H.; Osaki, M. Effects of disturbances on the carbon balance of tropical peat swamp forests. Glob. Chang. Biol. 2012, 18, 3410–3422. [Google Scholar] [CrossRef]
- Kanokratana, P.; Uengwetwanit, T.; Rattanachomsri, U.; Bunterngsook, B.; Nimchua, T.; Tangphatsornruang, S.; Plengvidhya, V.; Champreda, V.; Eurwilaichitr, L. Insights into the phylogeny and metabolic potential of a primary tropical peat swamp forest microbial community by metagenomic analysis. Microb. Ecol. 2011, 61, 518–528. [Google Scholar] [CrossRef]
- Bowen, J.L.; Morrison, H.G.; Hobbie, J.E.; Sogin, M.L. Salt marsh sediment diversity: A test of the variability of the rare biosphere among environmental replicates. ISME J. 2012, 6, 2014–2023. [Google Scholar] [CrossRef]
- Osburn, C.L.; Mikan, M.P.; Etheridge, J.R.; Burchell, M.R.; Birgand, F. Seasonal variation in the quality of dissolved and particulate organic matter exchanged between a salt marsh and its adjacent estuary. J. Geophys. Res. Biogeosciences 2015, 120, 1430–1449. [Google Scholar] [CrossRef]
- Sun, H.; Jiang, J.; Cui, L.; Feng, W.; Wang, Y.; Zhang, J. Soil organic carbon stabilization mechanisms in a subtropical mangrove and salt marsh ecosystems. Sci. Total Environ. 2019, 673, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Oremland, R.S.; Marsh, L.M.; Polcin, S. Methane production and simultaneous sulphate reduction in anoxic, salt marsh sediments. Nature 1982, 296, 143–145. [Google Scholar] [CrossRef]
- Metcalf, J.L.; Xu, Z.Z.; Bouslimani, A.; Dorrestein, P.; Carter, D.O.; Knight, R. Microbiome Tools for Forensic Science. Trends Biotechnol. 2017, 35, 814–823. [Google Scholar] [CrossRef]
- Langley, N.R.; Wood, P.; Herling, P.; Steadman, D.W. Forensic Postmortem Interval Estimation from Skeletal Muscle Tissue: A Lipidomics Approach. Forensic Anthropol. 2019, 2, 152–157. [Google Scholar] [CrossRef]
- Pechal, J.L.; Crippen, T.L.; Benbow, M.E.; Tarone, A.M.; Dowd, S.; Tomberlin, J.K. The potential use of bacterial community succession in forensics as described by high throughput metagenomic sequencing. Int. J. Legal Med. 2014, 128, 193–205. [Google Scholar] [CrossRef]
- Howard, G.T.; Duos, B.; Watson-Horzelski, E.J. Characterization of the soil microbial community associated with the decomposition of a swine carcass. Int. Biodeterior. Biodegrad. 2010, 64, 300–304. [Google Scholar] [CrossRef]
- Cobaugh, K.L.; Schaeffer, S.M.; DeBruyn, J.M. Functional and Structural Succession of Soil Microbial Communities below Decomposing Human Cadavers. PLoS ONE 2015, 10, e0130201. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moore, E.K. Trimethylornithine Membrane Lipids: Discovered in Planctomycetes and Identified in Diverse Environments. Metabolites 2021, 11, 49. https://doi.org/10.3390/metabo11010049
Moore EK. Trimethylornithine Membrane Lipids: Discovered in Planctomycetes and Identified in Diverse Environments. Metabolites. 2021; 11(1):49. https://doi.org/10.3390/metabo11010049
Chicago/Turabian StyleMoore, Eli K. 2021. "Trimethylornithine Membrane Lipids: Discovered in Planctomycetes and Identified in Diverse Environments" Metabolites 11, no. 1: 49. https://doi.org/10.3390/metabo11010049
APA StyleMoore, E. K. (2021). Trimethylornithine Membrane Lipids: Discovered in Planctomycetes and Identified in Diverse Environments. Metabolites, 11(1), 49. https://doi.org/10.3390/metabo11010049