
Natural Steroidal Lactone Induces G1/S Phase Cell Cycle Arrest and Intrinsic Apoptotic Pathway by Up-Regulating Tumor Suppressive miRNA in Triple-Negative Breast Cancer Cells




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Lines
2.2. Cytotoxicity Assay
2.3. Small RNA Sample Preparation
2.4. Library Construction and Small RNA Next Generation Sequencing
2.5. Differential Expression (DE) Analysis
2.6. Microrna Target Gene Prediction and Functional Enrichment Analysis
2.7. Quantitative RT-PCR Validation of Top 5 up and Top 5 down DEMs
2.8. Transient Transfection of miR-181c-5p Mimic
2.9. Effect of Withaferin A and miR-181c-5p Mimic Co-Treatment on TNBC Cell Viability
2.10. Confocal Microscopy
2.11. Annexin V/FITC and PI Apoptosis Assay
2.12. Expression Analysis of Cell Cycle and Apoptosis Related Genes
2.13. Caspase-Glo® 3/7 Activity
2.14. Western Blotting Analysis of Cell Cycle and Apoptotic Proteins
2.15. Statistical Analysis
3. Results
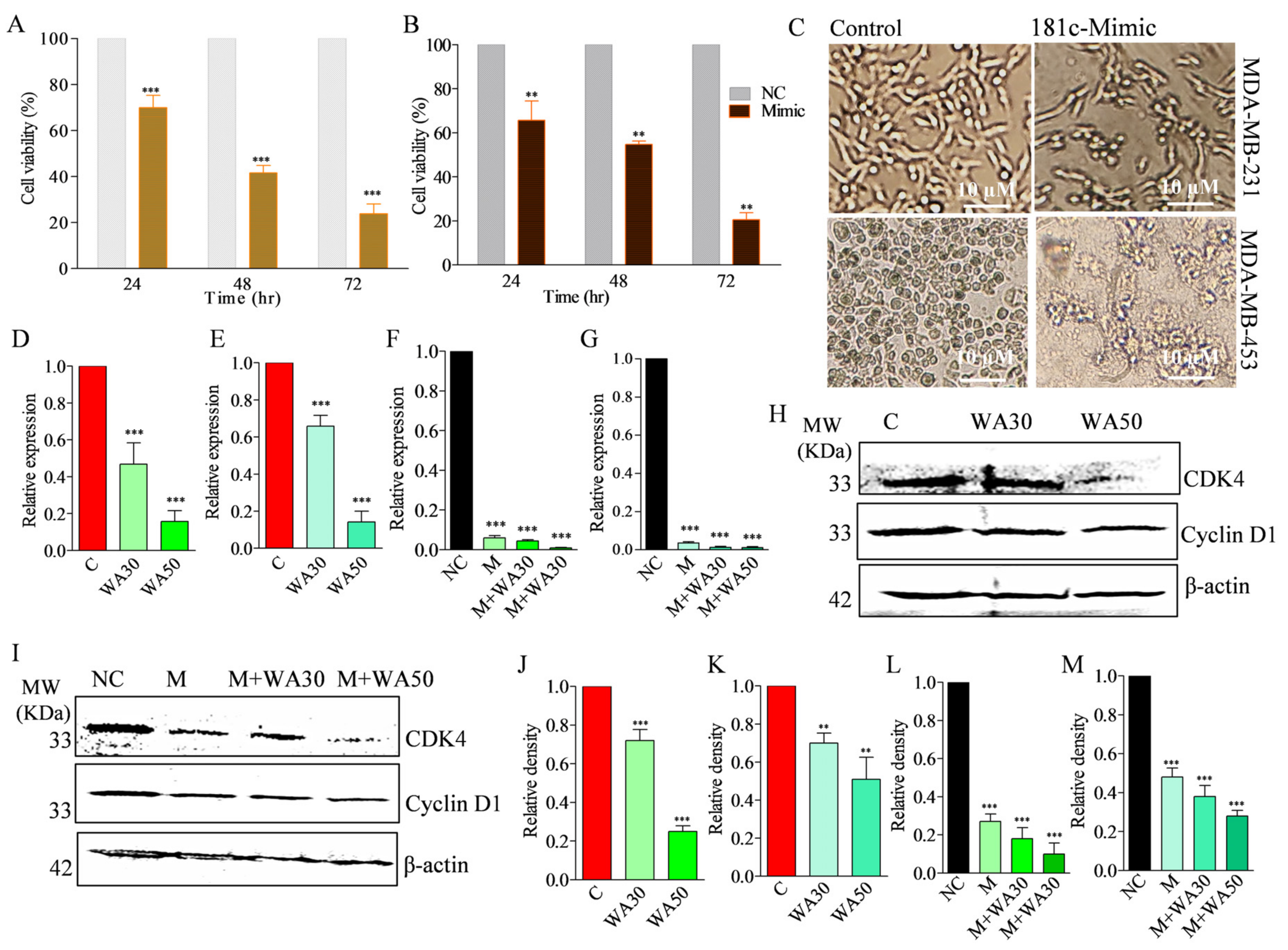
3.1. Withaferin A Induces Cytotoxicity in Triple-Negative Breast Cancer Cells
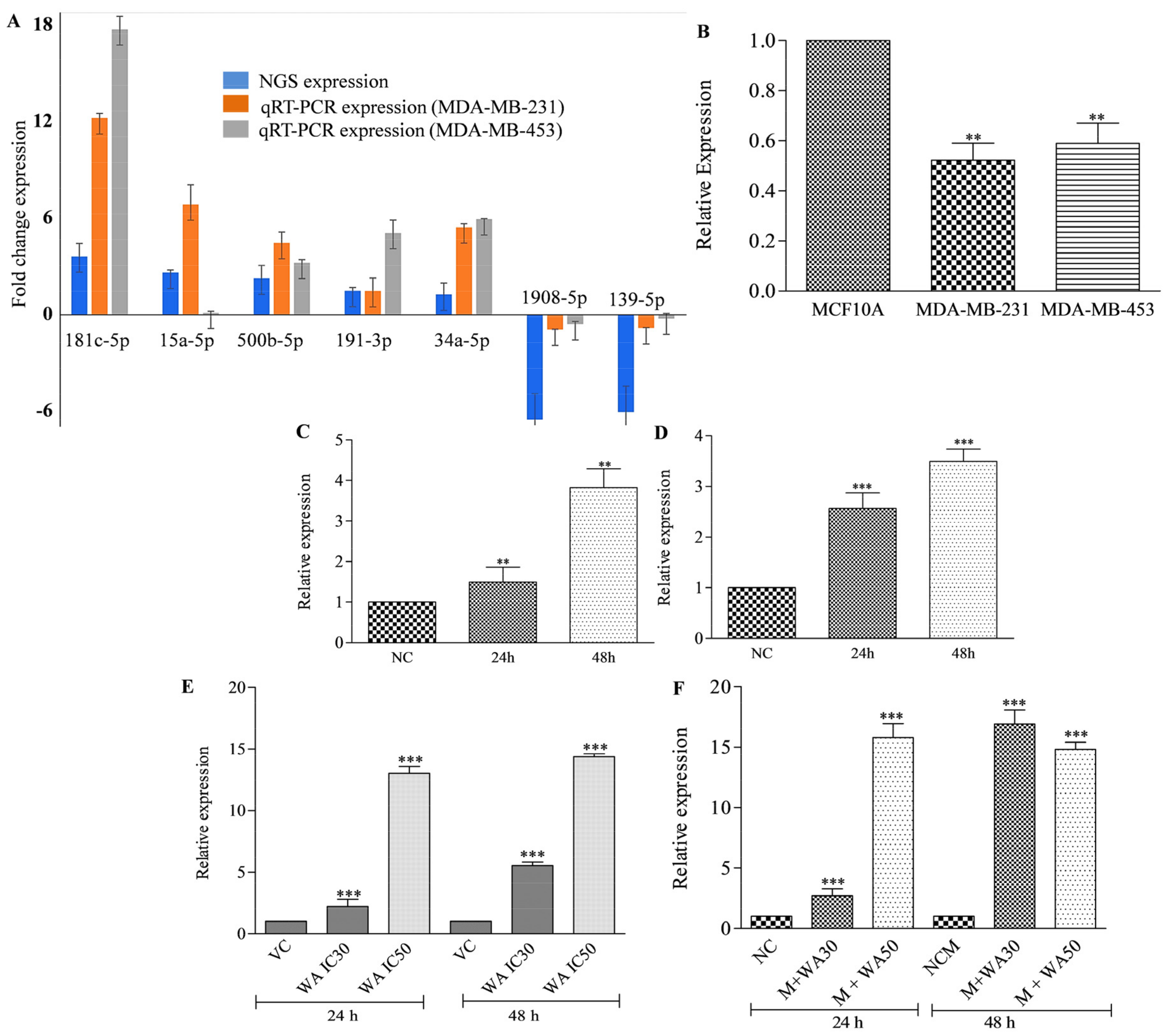
3.2. Withaferin A Treatment Altered MiRNA Expression Profile in MDA-MB-231 Cells
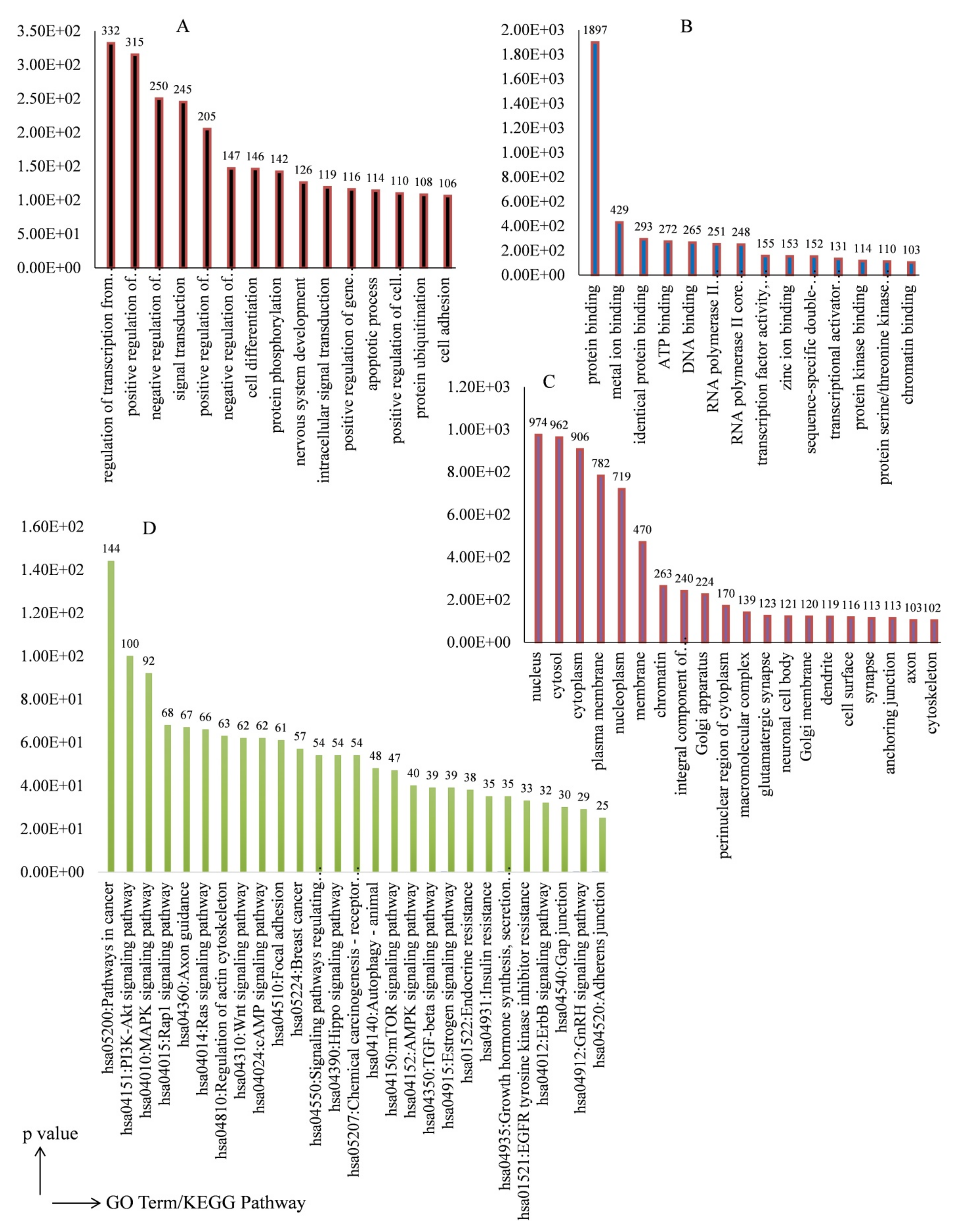
3.3. Target Gene Prediction and GO Enrichment Analysis
3.4. KEGG Pathway Enrichment Analysis
3.5. Withaferin A Potentially Induces Expression of miRNA-181c-5p in TNBC Cells
3.6. Withaferin A and miR-181c-5p Mimic Co-Treatment Decreases Cell Proliferation in TNBC Cells
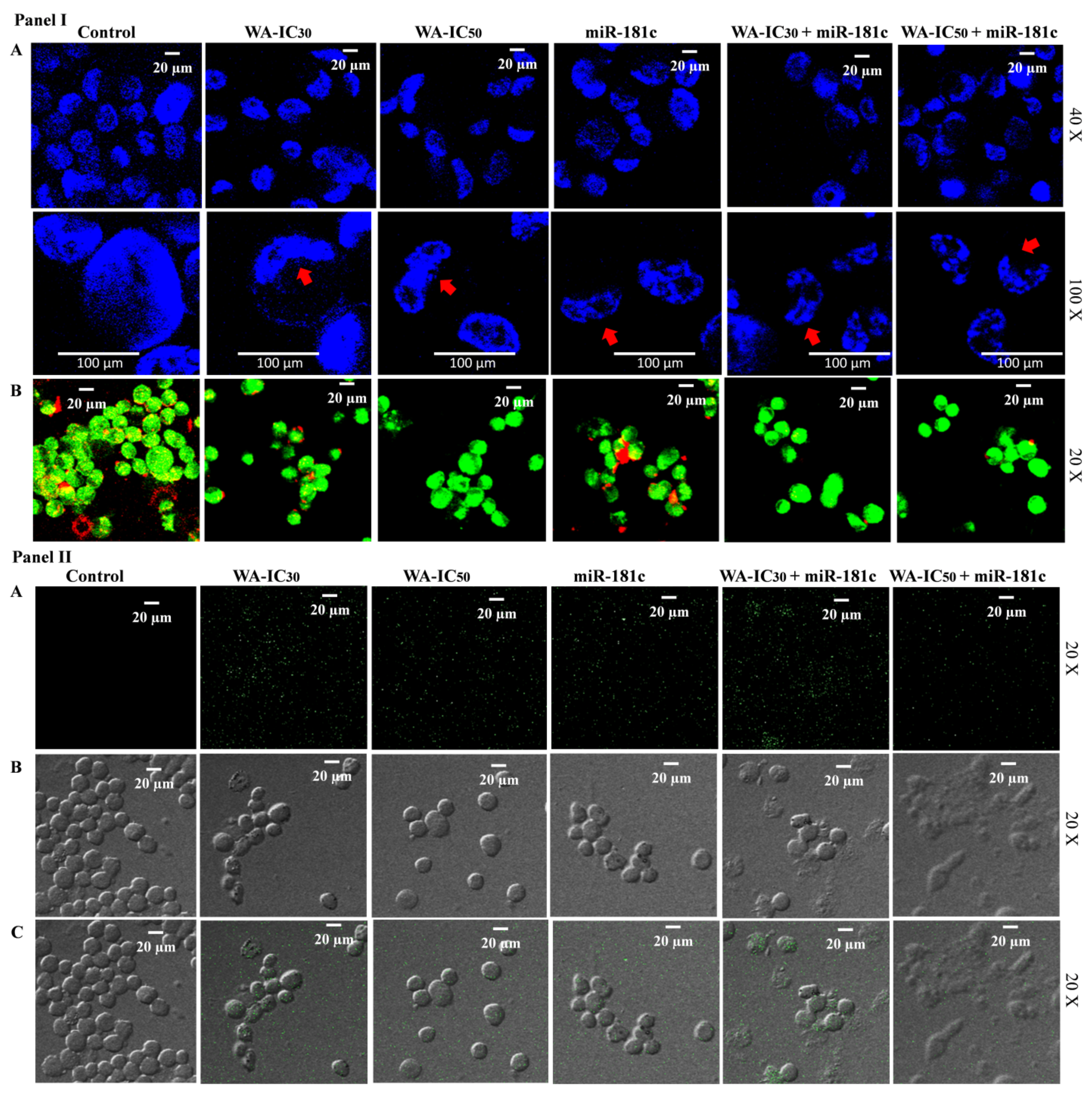
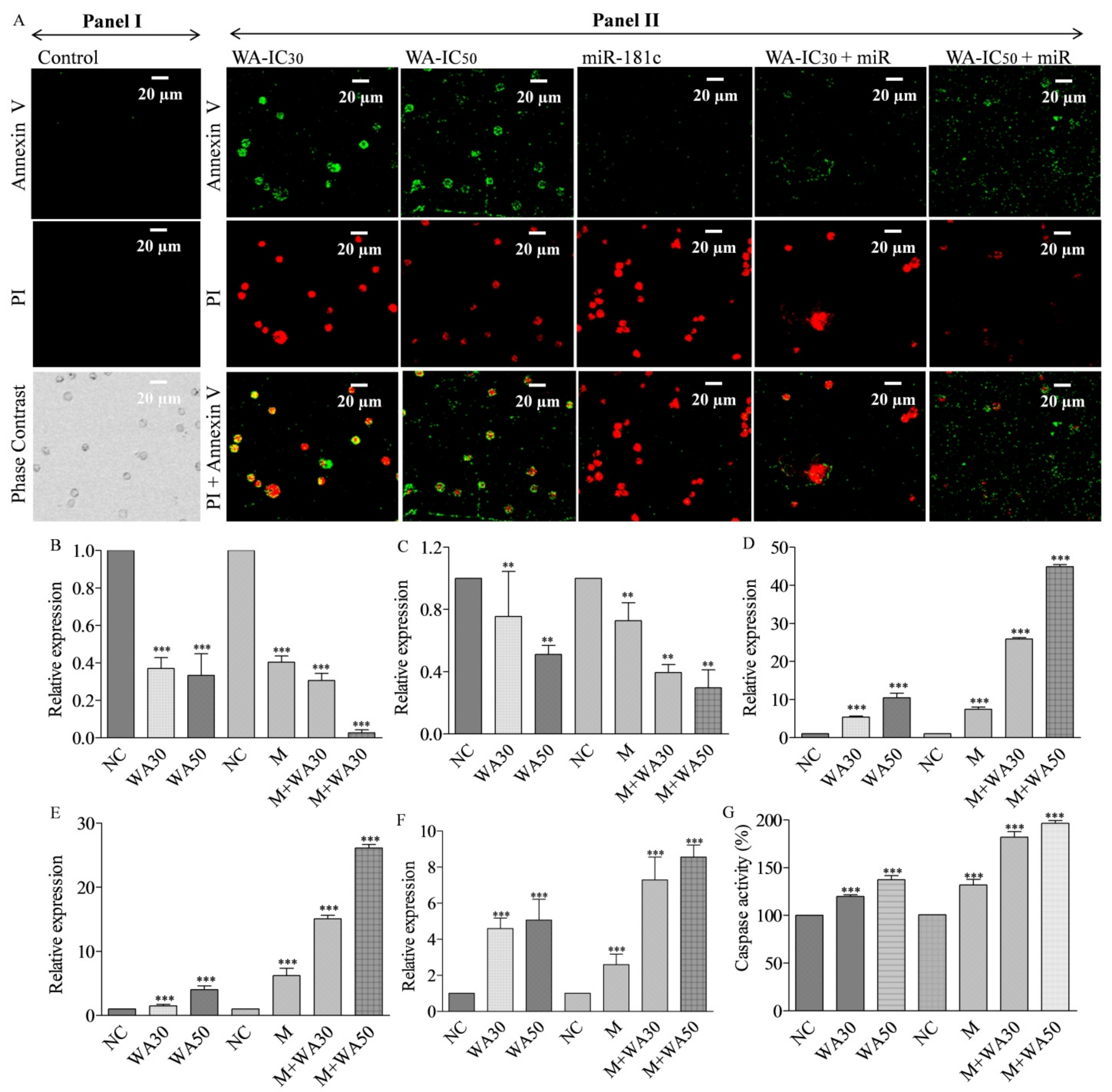
3.7. Withaferin A and miR-181c-5p Mimic Co-Treatment Potentiates Nuclear Morphology Alterations, Mitochondria Membrane Potential Decrease, Reactive Oxygen Species Generation, and Apoptotic Cell Population in MDA-MB-231 Cells
3.8. Withaferin A and miR-181c-5p Mimic Potentiates Caspase-Mediated Apoptosis Induction in MDA-MB-231 Cells
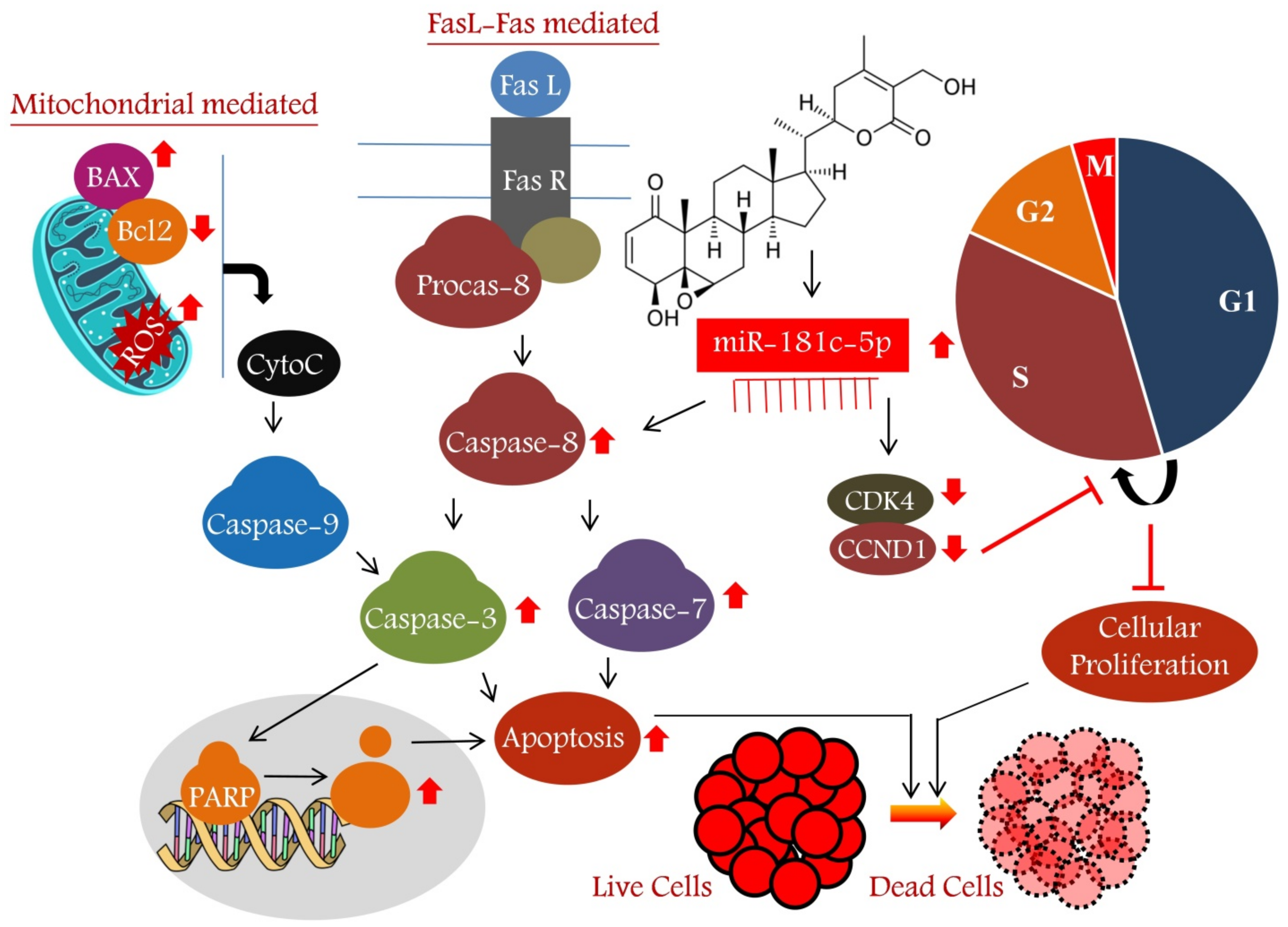
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Simões-Wüst, A.P.; Schürpf, T.; Hall, J.; Stahel, R.A.; Zangemeister-Wittke, U. Bcl-2/bcl-xL bispecific antisense treatment sensitizes breast carcinoma cells to doxorubicin, paclitaxel and cyclophosphamide. Breast Cancer Res. Treat. 2002, 76, 157–166. [Google Scholar] [CrossRef]
- Campbell, K.J.; Dhayade, S.; Ferrari, N.; Sims, A.; Johnson, E.; Mason, S.; Dickson, A.; Ryan, K.M.; Kalna, G.; Edwards, J.; et al. MCL-1 is a prognostic indicator and drug target in breast cancer. Cell Death Dis. 2018, 9, 19. [Google Scholar] [CrossRef]
- Ozretić, P.; Alvir, I.; Sarcevic, B.; Vujaskovic, Z.; Rendic-Miocevic, Z.; Roguljic, A.; Beketic-Oreskovic, L. Apoptosis regulator Bcl-2 is an independent prognostic marker for worse overall survival in triple-negative breast cancer patients. Int. J. Biol. Markers 2018, 33, 109–115. [Google Scholar] [CrossRef]
- Waseem, M.; Ahmad, M.; Srivastava, V.; Rastogi, N.; Serajuddin, M.; Kumar, S.; Mishra, D.; Sankhwar, S.N.; Mahdi, A. Evaluation of miR-711 as Novel Biomarker in Prostate Cancer Progression. Asian Pac. J. Cancer Prev. 2017, 18, 2185–2191. [Google Scholar] [CrossRef]
- Prajapati, K.S.; Shuaib, M.; Kushwaha, P.P.; Singh, A.K.; Kumar, S. Identification of cancer stemness related miRNA(s) using integrated bioinformatics analysis and in vitro validation. 3 Biotech 2021, 11, 446. [Google Scholar] [CrossRef]
- Shuaib, M.; Prajapati, K.S.; Singh, A.K.; Kushwaha, P.P.; Waseem, M.; Kumar, S. Identification of miRNAs and related hub genes associated with the triple negative breast cancer using integrated bioinformatics analysis and in vitro approach. J. Biomol. Struct. Dyn. 2021, 11, 446. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, S.; Pandey, A.K. In Vitro Antibacterial, Antioxidant, and Cytotoxic Activities of Parthenium hysterophorus and Characterization of Extracts by LC-MS Analysis. BioMed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, R.; Dwivedi, A.; Pandey, A.K. In Vitro Antioxidant, Antibacterial, and Cytotoxic Activity and In Vivo Effect of Syngonium podophyllum and Eichhornia crassipes Leaf Extracts on Isoniazid Induced Oxidative Stress and Hepatic Markers. BioMed Res. Int. 2014, 2014, 459452. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, P.P.; Singh, A.K.; Prajapati, K.S.; Shuaib, M.; Fayez, S.; Bringmann, G.; Kumar, S. Induction of apoptosis in breast cancer cells by naphthylisoquinoline alkaloids. Toxicol. Appl. Pharmacol. 2020, 409, 115297. [Google Scholar] [CrossRef]
- Kushwaha, P.P.; Vardhan, P.S.; Kapewangolo, P.; Shuaib, M.; Prajapati, S.K.; Singh, A.K.; Kumar, S. Bulbine frutescens phytochemical inhibits notch signaling pathway and induces apoptosis in triple negative and luminal breast cancer cells. Life Sci. 2019, 234, 116783. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, S. Dietary phytochemicals and their role in cancer chemoprevention. J. Cancer Metastasis Treat. 2021, 7, 51. [Google Scholar] [CrossRef]
- Kumar, S.; Prajapati, K.S.; Shuaib, M.; Kushwaha, P.P.; Tuli, H.S.; Singh, A.K. Five-Decade Update on Chemopreventive and Other Pharmacological Potential of Kurarinone: A Natural Flavanone. Front. Pharmacol. 2021, 12, 737137. [Google Scholar] [CrossRef]
- Mirjalili, M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazón, J. Steroidal Lactones from Withania somnifera, an Ancient Plant for Novel Medicine. Molecules 2009, 14, 2373–2393. [Google Scholar] [CrossRef]
- Hahm, E.R.; Kim, S.H.; Singh, K.B.; Singh, K.; Singh, S.V. A Comprehensive Review and Perspective on Anticancer Mechanisms of Withaferin A in Breast Cancer. Cancer Prevent Res. 2020, 13, 721–734. [Google Scholar] [CrossRef]
- Szic, K.S.V.; Declerck, K.; Crans, R.A.; Diddens, J.; Scherf, D.B.; Gerhäuser, C.; Berghe, W.V. Epigenetic silencing of triple negative breast cancer hallmarks by Withaferin, A. Oncotarget 2017, 8, 40434–40453. [Google Scholar] [CrossRef]
- Vel Szic, K.S.; de Beeck, K.O.; Ratman, D.; Wouters, A.; Beck, I.; Declerck, K.; Heyninck, K.; Fransen, E.; Bracke, M.; De Bosscher, K.; et al. Pharmacological Levels of Withaferin A (Withania somnifera) Trigger Clinically Relevant Anticancer Effects Specific to Triple Negative Breast Cancer Cells. PLoS ONE 2014, 9, e87850. [Google Scholar] [CrossRef]
- Kim, S.-H.; Hahm, E.-R.; Singh, K.B.; Shiva, S.; Stewart-Ornstein, J.; Singh, S.V. RNA-seq reveals novel mechanistic targets of withaferin A in prostate cancer cells. Carcinogenesis 2020, 41, 778–789. [Google Scholar] [CrossRef]
- Singh, A.K.; Shuaib, M.; Prajapati, K.S.; Kumar, S. Rutin Potentially Binds the Gamma Secretase Catalytic Site, Down Regulates the Notch Signaling Pathway and Reduces Sphere Formation in Colonospheres. Metabolites 2022, 12, 926. [Google Scholar] [CrossRef]
- Prajapati, K.S.; Shuaib, M.; Gupta, S.; Kumar, S. Withaferin A mediated changes of miRNA expression in breast cancer-derived mammospheres. Mol. Carcinog. 2022, 61, 876–889. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2013, 42, D68–D73. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Singh, A.K.; Prajapati, K.S.; Kumar, S. Hesperidin potentially interacts with the catalytic site of gamma-secretase and modifies notch sensitive genes and cancer stemness marker expression in colon cancer cells and colonosphere. J. Biomol. Struct. Dyn. 2022, 1–13. [Google Scholar] [CrossRef]
- Yuan, L.; Cai, Y.; Zhang, L.; Liu, S.; Li, P.; Li, X. Promoting Apoptosis, a Promising Way to Treat Breast Cancer with Natural Products: A Comprehensive Review. Front. Pharmacol. 2022, 12, 801662254. [Google Scholar] [CrossRef]
- Lakhani, S.A.; Masud, A.; Kuida, K.; Porter, G.A.; Booth, C.J.; Mehal, W.Z.; Inayat, I.; Flavell, R.A. Caspases 3 and 7: Key Mediators of Mitochondrial Events of Apoptosis. Science 2006, 311, 847–851. [Google Scholar] [CrossRef]
- Mallipeddi, H.; Thyagarajan, A.; Sahu, R.P. Implications of Withaferin-A for triple-negative breast cancer chemoprevention. Biomed. Pharmacother. 2021, 134, 111124. [Google Scholar] [CrossRef]
- Xie, D.; Li, S.; Wu, T.; Wang, X.; Fang, L. MiR-181c suppresses triple-negative breast cancer tumorigenesis by targeting MAP4K4. Pathol.-Res. Pract. 2022, 230, 153763. [Google Scholar] [CrossRef]
- Shinden, Y.; Akiyoshi, S.; Ueo, H.; Nambara, S.; Saito, T.; Komatsu, H.; Ueda, M.; Hirata, H.; Sakimura, S.; Uchi, R.; et al. Diminished expression of MiR-15a is an independent prognostic marker for breast cancer cases. Anticancer Res. 2015, 35, 123–127. [Google Scholar]
- Youness, R.A.; Hafez, H.M.; Khallaf, E.; Assal, R.A.; Motaal, A.A.; Gad, M.Z. The long noncoding RNA sONE represses triple-negative breast cancer aggressiveness through inducing the expression of miR-34a, miR-15a, miR-16, and let-7a. J. Cell. Physiol. 2019, 234, 20286–20297. [Google Scholar] [CrossRef]
- Vishnubalaji, R.; Alajez, N.M. Epigenetic regulation of triple negative breast cancer (TNBC) by TGF-β signaling. Sci. Rep. 2021, 11, 15410. [Google Scholar] [CrossRef]
- Assidicky, R.; Tokat, U.M.; Tarman, I.O.; Saatci, O.; Ersan, P.G.; Raza, U.; Ogul, H.; Riazalhosseini, Y.; Can, T.; Sahin, O. Targeting HIF1-alpha/miR-326/ITGA5 axis potentiates chemotherapy response in triple-negative breast cancer. Breast Cancer Res. Treat. 2022, 193, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Bedja, D.; Campbell, N.; Dunkerly, B.; Chenna, V.; Maitra, A.; Steenbergen, C. miR-181c Regulates the Mitochondrial Genome, Bioenergetics, and Propensity for Heart Failure In Vivo. PLoS ONE 2014, 9, e96820. [Google Scholar] [CrossRef] [PubMed]
- Indrieri, A.; Carrella, S.; Carotenuto, P.; Banfi, S.; Franco, B. The Pervasive Role of the miR-181 Family in Development, Neurodegeneration, and Cancer. Int. J. Mol. Sci. 2020, 21, 2092. [Google Scholar] [CrossRef]
- Mani, S.; Swargiary, G.; Singh, K.K. Natural Agents Targeting Mitochondria in Cancer. Int. J. Mol. Sci. 2020, 21, 6992. [Google Scholar] [CrossRef]
- Hahm, E.-R.; Moura, M.B.; Kelley, E.E.; Van Houten, B.; Shiva, S.; Singh, S.V. Withaferin A-Induced Apoptosis in Human Breast Cancer Cells Is Mediated by Reactive Oxygen Species. PLoS ONE 2011, 6, e23354. [Google Scholar] [CrossRef]
- Zhang, L.-Y.; Yu, J.-Y.; Leng, Y.-L.; Zhu, R.-R.; Liu, H.-X.; Wang, X.-Y.; Yang, T.-T.; Guo, Y.-N.; Tang, J.-L.; Zhang, X.-C. MiR-181c sensitizes ovarian cancer cells to paclitaxel by targeting GRP78 through the PI3K/Akt pathway. Cancer Gene Ther. 2021, 29, 770–783. [Google Scholar] [CrossRef]
- Sun, J.; Ma, Q.; Li, B.; Wang, C.; Mo, L.; Zhang, X.; Tang, F.; Wang, Q.; Yan, X.; Yao, X.; et al. RPN2 is targeted by miR-181c and mediates glioma progression and temozolomide sensitivity via the wnt/β-catenin signaling pathway. Cell Death Dis. 2020, 11, 890. [Google Scholar] [CrossRef]
- Zhuang, L.; Qu, H.; Cong, J.; Dai, H.; Liu, X. MiR-181c affects estrogen-dependent endometrial carcinoma cell growth by targeting PTEN. Endocr. J. 2019, 66, 523–533. [Google Scholar] [CrossRef]
- Ai, J.; Gong, C.; Wu, J.; Gao, J.; Liu, W.; Liao, W.; Wu, L. MicroRNA-181c suppresses growth and metastasis of hepatocellular carcinoma by modulating NCAPG. Cancer Manag. Res. 2019, 11, 3455–3467. [Google Scholar] [CrossRef]
- Li, N.; Cheng, C.; Wang, T. MiR-181c-5p Mitigates Tumorigenesis in Cervical Squamous Cell Carcinoma via Targeting Glycogen Synthase Kinase 3β Interaction Protein (GSKIP). OncoTargets Ther. 2020, 13, 4495–4505. [Google Scholar] [CrossRef]
- Wang, H.-C.; Hu, H.-H.; Chang, F.-R.; Tsai, J.-Y.; Kuo, C.-Y.; Wu, Y.-C.; Wu, C.-C. Different effects of 4β-hydroxywithanolide E and withaferin A, two withanolides from Solanaceae plants, on the Akt signaling pathway in human breast cancer cells. Phytomedicine 2018, 53, 213–222. [Google Scholar] [CrossRef]
- Zhou, J.; Luo, Y.-H.; Wang, J.-R.; Lu, B.-B.; Wang, K.-M.; Tian, Y. Gambogenic Acid Induction of Apoptosis in a Breast Cancer Cell Line. Asian Pac. J. Cancer Prev. 2013, 14, 7601–7605. [Google Scholar] [CrossRef]
- Lee, D.; Yu, J.S.; Ryoo, R.; Kim, J.-C.; Jang, T.S.; Kang, K.S.; Kim, K.H. Pulveraven A from the fruiting bodies of Pulveroboletus ravenelii induces apoptosis in breast cancer cell via extrinsic apoptotic signaling pathway. J. Antibiot. 2021, 74, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Machin, R.P.; Veleiro, A.S.; Nicotra, V.E.; Oberti, J.C.; Padrón, J.M. Antiproliferative Activity of Withanolides against Human Breast Cancer Cell Lines. J. Nat. Prod. 2010, 73, 966–968. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shuaib, M.; Prajapati, K.S.; Gupta, S.; Kumar, S. Natural Steroidal Lactone Induces G1/S Phase Cell Cycle Arrest and Intrinsic Apoptotic Pathway by Up-Regulating Tumor Suppressive miRNA in Triple-Negative Breast Cancer Cells. Metabolites 2023, 13, 29. https://doi.org/10.3390/metabo13010029
Shuaib M, Prajapati KS, Gupta S, Kumar S. Natural Steroidal Lactone Induces G1/S Phase Cell Cycle Arrest and Intrinsic Apoptotic Pathway by Up-Regulating Tumor Suppressive miRNA in Triple-Negative Breast Cancer Cells. Metabolites. 2023; 13(1):29. https://doi.org/10.3390/metabo13010029
Chicago/Turabian StyleShuaib, Mohd, Kumari Sunita Prajapati, Sanjay Gupta, and Shashank Kumar. 2023. "Natural Steroidal Lactone Induces G1/S Phase Cell Cycle Arrest and Intrinsic Apoptotic Pathway by Up-Regulating Tumor Suppressive miRNA in Triple-Negative Breast Cancer Cells" Metabolites 13, no. 1: 29. https://doi.org/10.3390/metabo13010029
APA StyleShuaib, M., Prajapati, K. S., Gupta, S., & Kumar, S. (2023). Natural Steroidal Lactone Induces G1/S Phase Cell Cycle Arrest and Intrinsic Apoptotic Pathway by Up-Regulating Tumor Suppressive miRNA in Triple-Negative Breast Cancer Cells. Metabolites, 13(1), 29. https://doi.org/10.3390/metabo13010029