
The Binding of the SARS-CoV-2 Spike Protein to Platelet Factor 4: A Proposed Mechanism for the Generation of Pathogenic Antibodies
 , and
, and
Abstract

1. Introduction
2. Methods
2.1. Ethics
2.2. Reagents
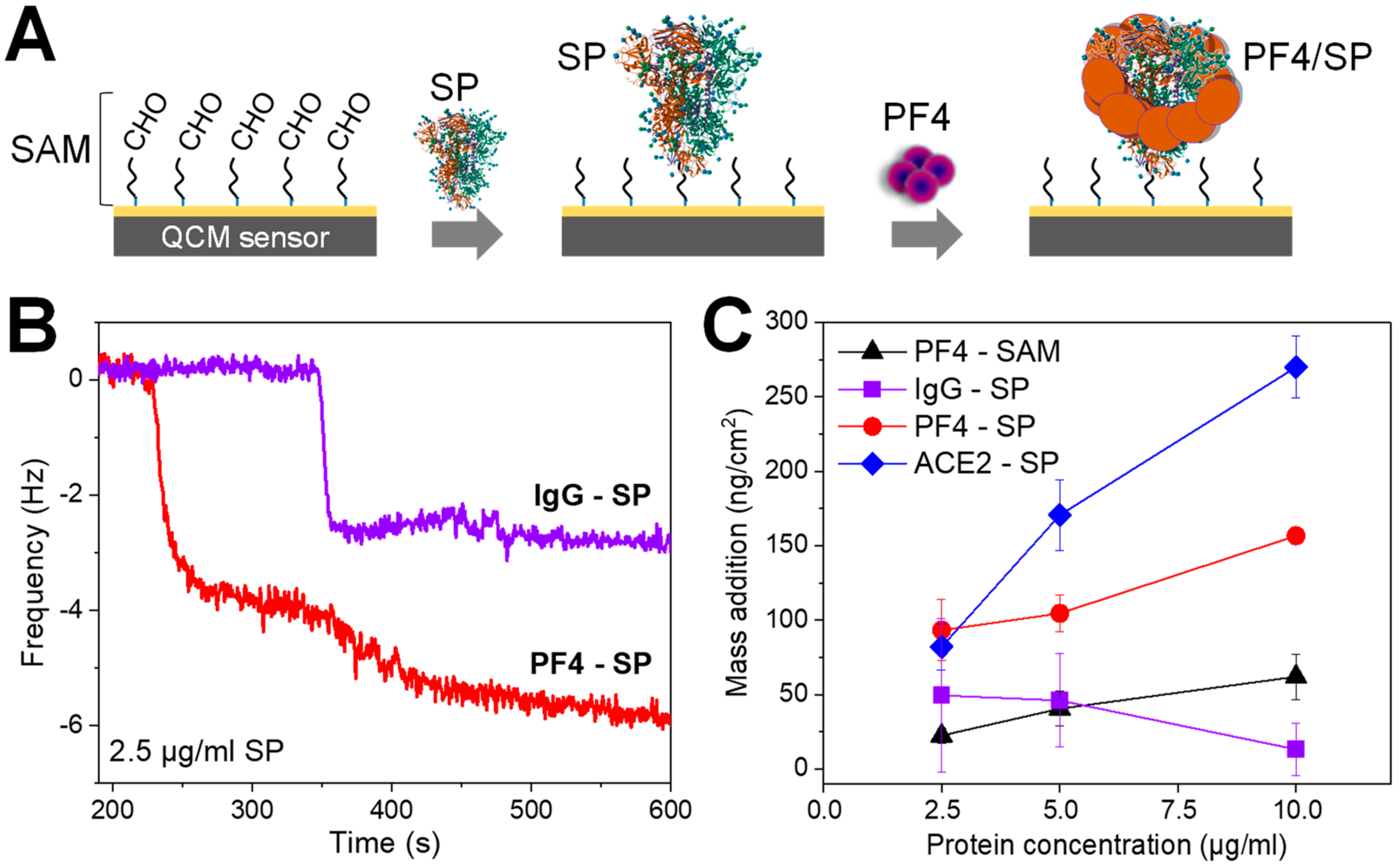
2.3. Quartz Crystal Microbalance (QCM)
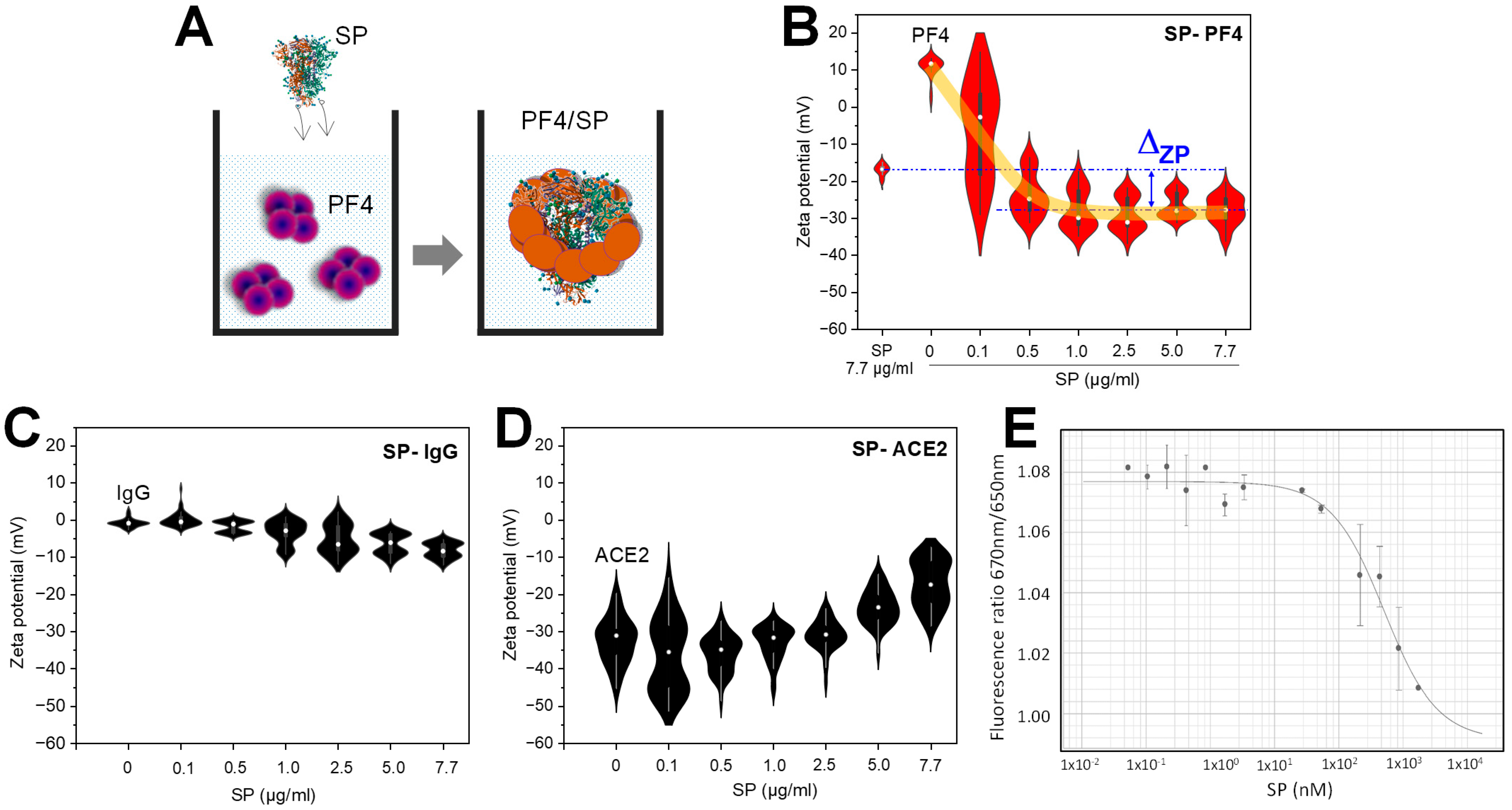
2.4. Dynamic Light Scattering (DLS)
2.5. Isothermal Spectral Shift Analysis (ISSA)
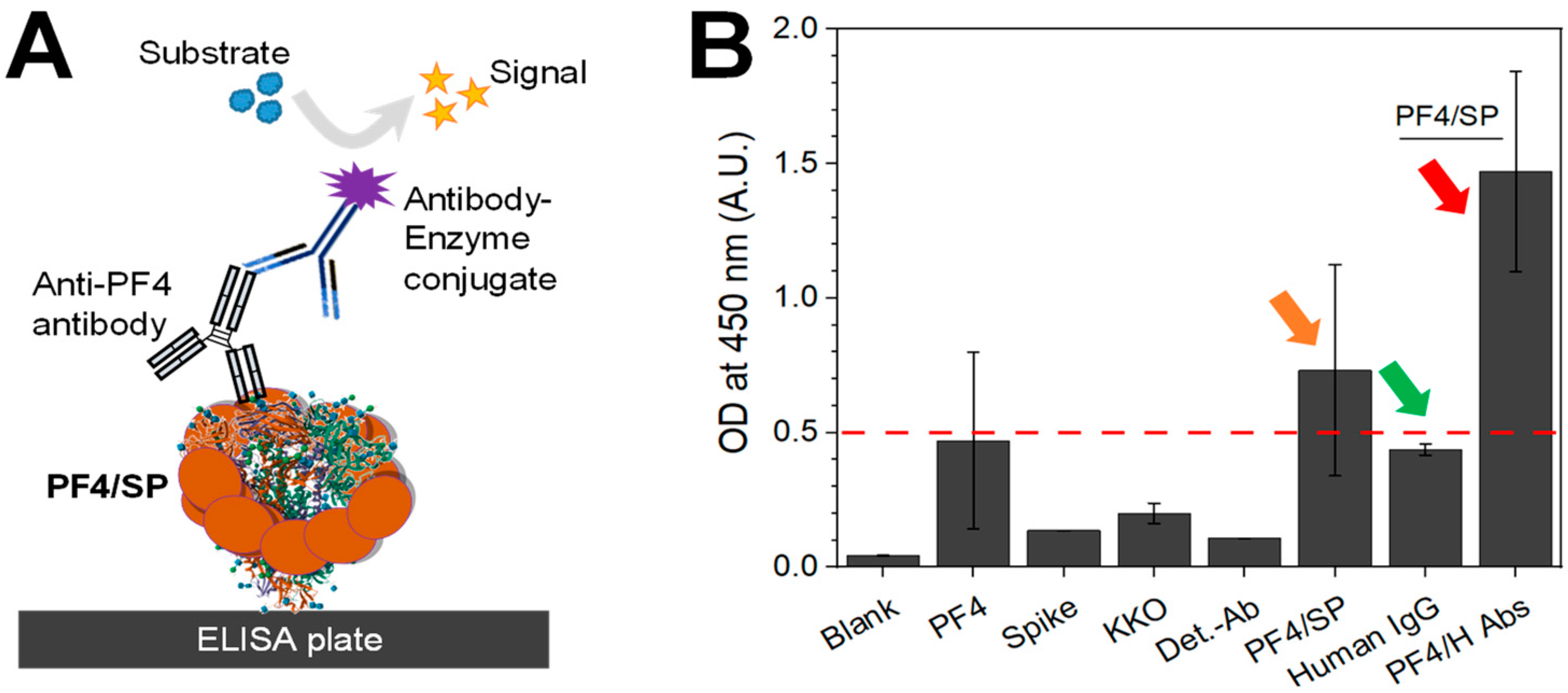
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
3. Results
3.1. The Binding of SARS-CoV-2 SP to PF4 by the Mean of the Quartz Crystal Microbalance (QCM)
3.2. The Binding of SARS-CoV-2 SP to PF4 Alters Protein Structures
3.3. The Confirmation of PF4–SP Binding in the Enzyme-Linked Immunosorbent Assay (ELISA)
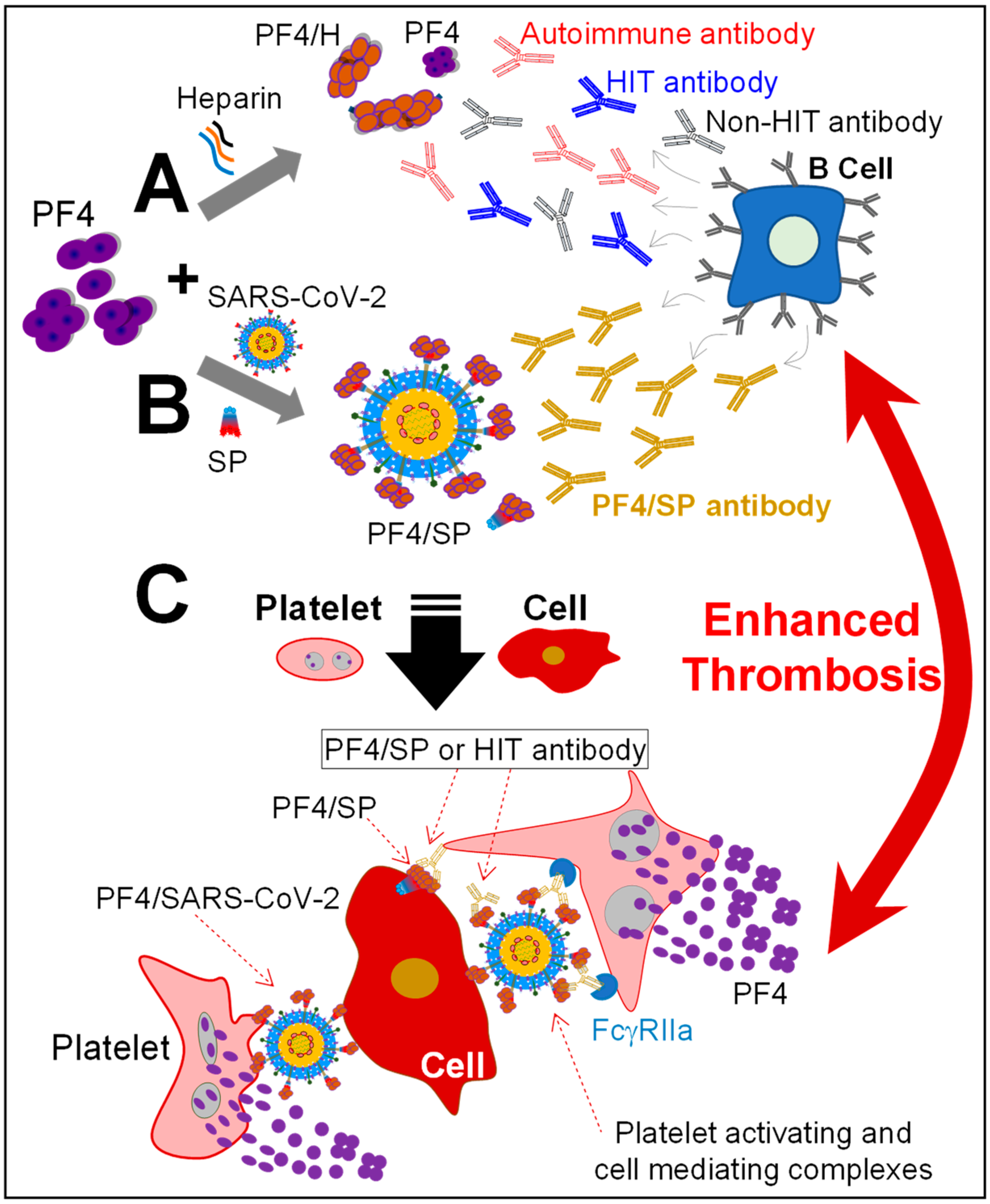
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. WHO Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 4 October 2023).
- Hanif, A.; Khan, S.; Mantri, N.; Hanif, S.; Saleh, M.; Alla, Y.; Chinta, S.; Shrestha, N.; Ji, W.; Attwood, K.; et al. Thrombotic complications and anticoagulation in COVID-19 pneumonia: A New York City hospital experience. Ann. Hematol. 2020, 99, 2323–2328. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Q.; Wang, Y.; Wu, Y.; Xu, J.; Yu, Y.; Shang, Y. Thrombocytopenia and Its Association with Mortality in Patients with COVID-19. J. Thromb. Haemost. 2020, 18, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Heamost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Chen, S.; Li, X.; Liu, S.; Wang, F. Prevalence of venous thromboembolism in patients with severe novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 1421–1424. [Google Scholar] [CrossRef] [PubMed]
- Middeldorp, S.; Coppens, M.; van Haaps, T.F.; Foppen, M.; Vlaar, A.P.; Muller, M.C.A.; Bouman, C.C.S.; Beenen, L.F.M.; Kootte, R.S.; Heijmans, J.; et al. Incidence of venous thromboembolism in hospitalized patients with COVID-19. J. Thromb. Haemost. 2020, 18, 1995–2002. [Google Scholar] [CrossRef] [PubMed]
- Klok, F.A.; Kruip, M.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Confirmation of the high cumulative incidence of thrombotic complications in critically ill ICU patients with COVID-19: An updated analysis. Thromb. Res. 2020, 191, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Warkentin, T.E.; Kaatz, S. COVID-19 versus HIT hypercoagulability. Thromb. Res. 2020, 196, 38–51. [Google Scholar] [CrossRef]
- Warkentin, T.E.; Levine, M.N.; Hirsh, J.; Horsewood, P.; Roberts, R.S.; Gent, M.; Kelton, J.G. Heparin-Induced Thrombocytopenia in Patients Treated with Low-Molecular-Weight Heparin or Unfractionated Heparin. N. Engl. J. Med. 1995, 332, 1330–1335. [Google Scholar] [CrossRef]
- Chen, L.Y.; Khan, N.; Lindenbauer, A.; Nguyen, T.H. When Will Fondaparinux Induce Thrombocytopenia? Bioconjug. Chem. 2022, 33, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Warkentin, T.E.; Kelton, J.G. A 14-year study of heparin-induced thrombocytopenia. Am. J. Med. 1996, 101, 502–507. [Google Scholar] [CrossRef]
- Dragonetti, D.; Guarini, G.; Pizzuti, M. Detection of anti-heparin-PF4 complex antibodies in COVID-19 patients on heparin therapy. Blood Transfus. 2020, 18, 328. [Google Scholar] [CrossRef]
- Althaus, K.; Marini, I.; Zlamal, J.; Pelzl, L.; Singh, A.; Haberle, H.; Mehrlander, M.; Hammer, S.; Schulze, H.; Bitzer, M.; et al. Antibody-induced procoagulant platelets in severe COVID-19 infection. Blood 2021, 137, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Miao, H.; Li, S.; Zhang, P.; Gerber, G.F.; Follmann, D.; Ji, H.; Zeger, S.L.; Chertow, D.S.; Quinn, T.C.; et al. Anti-PF4 antibodies associated with disease severity in COVID-19. Proc. Natl. Acad. Sci. USA 2022, 119, e2213361119. [Google Scholar] [CrossRef] [PubMed]
- Ueland, T.; Hausberg, I.; Mortberg, T.V.; Dahl, T.B.; Lerum, T.V.; Michelsen, A.; Ranheim, T.; Nezvalova Henriksen, K.; Dyrhol-Riise, A.M.; Holme, P.A.; et al. Anti-PF4/polyanion antibodies in COVID-19 patients are associated with disease severity and pulmonary pathology. Platelets 2022, 33, 640–644. [Google Scholar] [CrossRef]
- Thachil, J.; Tang, N.; Gando, S.; Falanga, A.; Cattaneo, M.; Levi, M.; Clark, C.; Iba, T. ISTH interim guidance on recognition and management of coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, M.; Bertinato, E.M.; Birocchi, S.; Brizio, C.; Malavolta, D.; Manzoni, M.; Muscarella, G.; Orlandi, M. Pulmonary Embolism or Pulmonary Thrombosis in COVID-19? Is the Recommendation to Use High-Dose Heparin for Thromboprophylaxis Justified? Thromb. Haemost. 2020, 120, 1230–1232. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, J.; Klamroth, R.; Langer, F.; Albisetti, M.; von Auer, C.; Ay, C.; Korte, W.; Scharf, R.E.; Potzsch, B.; Greinacher, A. Diagnosis and Management of Vaccine-Related Thrombosis following AstraZeneca COVID-19 Vaccination: Guidance Statement from the GTH. Hamostaseologie 2021, 41, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Greinacher, A.; Thiele, T.; Warkentin, T.E.; Weisser, K.; Kyrle, P.A.; Eichinger, S. Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. N. Engl. J. Med. 2021, 384, 2092–2101. [Google Scholar] [CrossRef] [PubMed]
- Scully, M.; Singh, D.; Lown, R.; Poles, A.; Solomon, T.; Levi, M.; Goldblatt, D.; Kotoucek, P.; Thomas, W.; Lester, W. Pathologic Antibodies to Platelet Factor 4 after ChAdOx1 nCoV-19 Vaccination. N. Engl. J. Med. 2021, 384, 2202–2211. [Google Scholar] [CrossRef]
- Althaus, K.; Moller, P.; Uzun, G.; Singh, A.; Beck, A.; Bettag, M.; Bosmuller, H.; Guthoff, M.; Dorn, F.; Petzold, G.C.; et al. Antibody-mediated procoagulant platelets in SARS-CoV-2- vaccination associated immune thrombotic thrombocytopenia. Haematologica 2021, 106, 2170. [Google Scholar] [CrossRef]
- Zuo, Y.; Estes, S.K.; Ali, R.A.; Gandhi, A.A.; Yalavarthi, S.; Shi, H.; Sule, G.; Gockman, K.; Madison, J.A.; Zuo, M.; et al. Prothrombotic autoantibodies in serum from patients hospitalized with COVID-19. Sci. Transl. Med. 2020, 12, eabd3876. [Google Scholar] [CrossRef]
- Nguyen, T.H. Single-molecule force spectroscopy applied to heparin-induced thrombocytopenia. J. Mol. Recognit. 2017, 30, e2585. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Greinacher, A. Effect of pH and ionic strength on the binding strength of anti-PF4/polyanion antibodies. Eur. Biophys. J. 2017, 46, 795–801. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Greinacher, A. Platelet factor 4/heparin complexes present epitopes differently on solid-phase vs platelet surfaces. Blood 2017, 129, 3498–3501. [Google Scholar] [CrossRef] [PubMed]
- Bui, V.C.; Nguyen, T.H. The Role of Single-Molecule Force Spectroscopy in Unraveling Typical and Autoimmune Heparin-induced Thrombocytopenia. Int. J. Mol. Sci. 2018, 19, 1054. [Google Scholar] [CrossRef] [PubMed]
- Rauova, L.; Poncz, M.; McKenzie, S.E.; Reilly, M.P.; Arepally, G.; Weisel, J.W.; Nagaswami, C.; Cines, D.B.; Sachais, B.S. Ultralarge complexes of PF4 and heparin are central to the pathogenesis of heparin-induced thrombocytopenia. Blood 2005, 105, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Greinacher, A. Heparin-Induced Thrombocytopenia. N. Engl. J. Med. 2015, 373, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Greinacher, A.; Delcea, M. Quantitative description of thermodynamic and kinetic properties of the platelet factor 4/heparin bonds. Nanoscale 2015, 7, 10130–10139. [Google Scholar] [CrossRef]
- Hui, M.; Sheppard, J.I.; Li, N.; Warkentin, T.E. Neutrophil and Monocyte Counts in Heparin-Induced Thrombocytopenia. Thromb. Haemost. 2019, 119, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Chong, B.H.; Pitney, W.R.; Castaldi, P.A.; Warkentin, T.E.; Kelton, J.G.; Warkentin, T.E. Heparin-induced Thrombocytopenia: To w a rds Consensus. Thromb. Haemost. 1998, 79, 1–7. [Google Scholar]
- Cines, D.B. Disorders Associated with Antibodies to Endothelial-Cells. Rev. Infect. Dis. 1989, 11, S705–S711. [Google Scholar] [CrossRef]
- Visentin, G.P.; Ford, S.E.; Scott, J.P.; Aster, R.H. Antibodies from Patients with Heparin-Induced Thrombocytopenia/Thrombosis Are Specific for Platelet Factor-4 Complexed with Heparin or Bound to Endothelial-Cells. J. Clin. Investig. 1994, 93, 81–88. [Google Scholar] [CrossRef]
- Warkentin, T.E.; Basciano, P.A.; Knopman, J.; Bernstein, R.A. Spontaneous heparin-induced thrombocytopenia syndrome: 2 new cases and a proposal for defining this disorder. Blood 2014, 123, 3651–3654. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Medvedev, N.; Delcea, M.; Greinacher, A. Anti-platelet factor 4/polyanion antibodies mediate a new mechanism of autoimmunity. Nat. Commun. 2017, 8, 14945. [Google Scholar] [CrossRef]
- Langer, A.; Bartoschik, T.; Cehlar, O.; Duhr, S.; Baaske, P.; Streicher, W. A New Spectral Shift-Based Method to Characterize Molecular Interactions. Assay Drug Dev. Technol. 2022, 20, 83–94. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Wesche, J.; Raschke, R.; Strobel, U.; Bui, V.C.; Delcea, M.; Greinacher, A. Reactivity of platelet-activating and nonplatelet-activating anti-PF4/heparin antibodies in enzyme immunosorbent assays under different conditions. J. Thromb. Haemost. 2019, 17, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Xu, Y.; Brandt, S.; Mandelkow, M.; Raschke, R.; Strobel, U.; Delcea, M.; Zhou, W.; Liu, J.; Greinacher, A. Characterization of the interaction between platelet factor 4 and homogeneous synthetic low molecular weight heparins. J. Thromb. Haemost. 2020, 18, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Vayne, C.; Nguyen, T.H.; Rollin, J.; Charuel, N.; Poupon, A.; Pouplard, C.; Normann, N.; Gruel, Y.; Greinacher, A. Characterization of New Monoclonal PF4-Specific Antibodies as Useful Tools for Studies on Typical and Autoimmune Heparin-Induced Thrombocytopenia. Thromb. Haemost. 2020, 121, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Greinacher, A. Distinct Binding Characteristics of Pathogenic Anti-Platelet Factor-4/Polyanion Antibodies to Antigens Coated on Different Substrates: A Perspective on Clinical Application. ACS Nano 2018, 12, 12030–12041. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.Z.; Chen, L.Y.; Lindenbauer, A.; Pliquett, U.; Rothe, H.; Nguyen, T.H. Label-Free Detection and Characterization of Heparin-Induced Thrombocytopenia (HIT)-like Antibodies. ACS Omega 2021, 6, 25926–25939. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.Z.; Martin, D.; Pliquett, U.; Zaikou, Y.; Thomas, N.; Heinrich, D.; Kohler, J.M.; Nguyen, T.H. High-frequency Contactless Sensor for the Detection of Heparin-Induced Thrombocytopenia Antibodies via Platelet Aggregation. Int. J. Mol. Sci. 2022, 23, 14395. [Google Scholar] [CrossRef]
- Passariello, M.; Vetrei, C.; Amato, F.; De Lorenzo, C. Interactions of Spike-RBD of SARS-CoV-2 and Platelet Factor 4: New Insights in the Etiopathogenesis of Thrombosis. Int. J. Mol. Sci. 2021, 22, 8562. [Google Scholar] [CrossRef]
- Cox, M.; Peacock, T.P.; Harvey, W.T.; Hughes, J.; Wright, D.W.; Consortium, C.-G.U.; Willett, B.J.; Thomson, E.; Gupta, R.K.; Peacock, S.J.; et al. SARS-CoV-2 variant evasion of monoclonal antibodies based on in vitro studies. Nat. Rev. Microbiol. 2023, 21, 112–124. [Google Scholar] [CrossRef]
- Wang, Q.; Iketani, S.; Li, Z.; Liu, L.; Guo, Y.; Huang, Y.; Bowen, A.D.; Liu, M.; Wang, M.; Yu, J.; et al. Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell 2023, 186, 279–286.e278. [Google Scholar] [CrossRef]
- Arora, P.; Kempf, A.; Nehlmeier, I.; Schulz, S.R.; Jack, H.M.; Pohlmann, S.; Hoffmann, M. Omicron sublineage BQ.1.1 resistance to monoclonal antibodies. Lancet Infect. Dis. 2023, 23, 22–23. [Google Scholar] [CrossRef]
- Bui, V.C.; Gebicka, P.; Hippe, H.; Raschke, R.; Nguyen, T.L.; Greinacher, A.; Nguyen, T.H. Physicochemical Characteristics of Platelet Factor 4 under Various Conditions are Relevant for Heparin-Induced Thrombocytopenia Testing. J. Phys. Chem. B 2020, 124, 1438–1443. [Google Scholar] [CrossRef]
- Suvarna, S.; Espinasse, B.; Qi, R.; Lubica, R.; Poncz, M.; Cines, D.B.; Wiesner, M.R.; Arepally, G.M. Determinants of PF4/heparin immunogenicity. Blood 2007, 110, 4253–4260. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Flamholz, A.; Phillips, R.; Milo, R. SARS-CoV-2 (COVID-19) by the numbers. eLife 2020, 9, e57309. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Schirmer, U.; Widder, M.; Gruel, Y.; Rollin, J.; Zipfel, P.F.; Nguyen, T.H. Breast cancer cell-based ELISA: A potential material for better detection of heparin-induced thrombocytopenia antibodies. J. Mater. Chem. B 2022, 10, 7708–7716. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Apte, G.; Lindenbauer, A.; Frant, M.; Nguyen, T.H. Effect of HIT Components on the Development of Breast Cancer Cells. Life 2021, 11, 832. [Google Scholar] [CrossRef] [PubMed]
- Aleem, A.; Nadeem, A.J. Coronavirus (COVID-19) Vaccine-Induced Immune Thrombotic Thrombocytopenia (VITT); StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Swank, Z.; Senussi, Y.; Manickas-Hill, Z.; Yu, X.G.; Li, J.Z.; Alter, G.; Walt, D.R. Persistent Circulating Severe Acute Respiratory Syndrome Coronavirus 2 Spike Is Associated With Post-acute Coronavirus Disease 2019 Sequelae. Clin. Infect. Dis. 2023, 76, e487–e490. [Google Scholar] [CrossRef] [PubMed]
- Zaid, Y.; Puhm, F.; Allaeys, I.; Naya, A.; Oudghiri, M.; Khalki, L.; Limami, Y.; Zaid, N.; Sadki, K.; Ben El Haj, R.; et al. Platelets Can Associate with SARS-CoV-2 RNA and Are Hyperactivated in COVID-19. Circ. Res. 2020, 127, 1404–1418. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Wang, H.; Chen, L.Y.; Echtermeyer, D.; Pliquett, U. Modulating SARS-CoV-2 Spike Protein Reactivity through Moderate Electric Fields: A Pathway to Innovative Therapies. ACS Omega 2023, 8, 45952–45960. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adding Material | Blank | Spike Control (SpikeCon) | KKO Control (KKOCon) | Detection Ab Control (Det-AbCon) | PF4/SP |
|---|---|---|---|---|---|
| SP | - | + | + | - | + |
| PF4 | - | - | - | - | + |
| KKO | - | - | + | - | + |
| Detection Ab | - | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.-H.; Chen, L.-Y.; Khan, N.Z.; Lindenbauer, A.; Bui, V.-C.; Zipfel, P.F.; Heinrich, D. The Binding of the SARS-CoV-2 Spike Protein to Platelet Factor 4: A Proposed Mechanism for the Generation of Pathogenic Antibodies. Biomolecules 2024, 14, 245. https://doi.org/10.3390/biom14030245
Nguyen T-H, Chen L-Y, Khan NZ, Lindenbauer A, Bui V-C, Zipfel PF, Heinrich D. The Binding of the SARS-CoV-2 Spike Protein to Platelet Factor 4: A Proposed Mechanism for the Generation of Pathogenic Antibodies. Biomolecules. 2024; 14(3):245. https://doi.org/10.3390/biom14030245
Chicago/Turabian StyleNguyen, Thi-Huong, Li-Yu Chen, Nida Zaman Khan, Annerose Lindenbauer, Van-Chien Bui, Peter F. Zipfel, and Doris Heinrich. 2024. "The Binding of the SARS-CoV-2 Spike Protein to Platelet Factor 4: A Proposed Mechanism for the Generation of Pathogenic Antibodies" Biomolecules 14, no. 3: 245. https://doi.org/10.3390/biom14030245
APA StyleNguyen, T.-H., Chen, L.-Y., Khan, N. Z., Lindenbauer, A., Bui, V.-C., Zipfel, P. F., & Heinrich, D. (2024). The Binding of the SARS-CoV-2 Spike Protein to Platelet Factor 4: A Proposed Mechanism for the Generation of Pathogenic Antibodies. Biomolecules, 14(3), 245. https://doi.org/10.3390/biom14030245