
Metabolic Syndrome and Biotherapeutic Activity of Dairy (Cow and Buffalo) Milk Proteins and Peptides: Fast Food-Induced Obesity Perspective—A Narrative Review
 , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Methodology
3. Obesity
3.1. Diabetes
3.2. Dyslipidemia
4. Management of Obesity
4.1. Nutraceuticals and MS
4.2. Proteins and Peptides from Milk Proteins
4.2.1. Anti-Diabetic Activity
4.2.2. Anti-Dyslipidemic Activity
4.3. Biochemical Mechanisms Considering IPP and VPP as Model Peptides
4.4. Clinical Investigations
4.4.1. Dairy Proteins in Meals and T2DM Subjects
4.4.2. Dairy Proteins in Preload Formulas and T2DM Subjects
4.4.3. Dairy Proteins in Meals and Obese/Overweight Subjects
4.4.4. Dairy Proteins in Preload Formulas and Obese/Overweight Subjects
4.4.5. Dairy Protein Hydrolysates and T2DM Subjects
4.4.6. Co-Consumption of Amino Acids with Dairy Proteins and T2DM Subjects
4.4.7. Contradictory Results
4.4.8. Limitations of Clinical Investigations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [Google Scholar] [CrossRef]
- Wu, M.; Shu, Y.; Wang, L.; Song, L.; Chen, S.; Liu, Y.; Bi, J.; Li, D.; Yang, Y.; Hu, Y.; et al. Visit-to-Visit Variability in the Measurements of Metabolic Syndrome Components and the Risk of All-Cause Mortality, Cardiovascular Disease, and Arterial Stiffness. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2895–2903. [Google Scholar] [CrossRef]
- Williams, V.F.; Oh, G.-T.; Stahlman, S. Incidence and Prevalence of the Metabolic Syndrome Using ICD-9 and ICD-10 Diagnostic Codes, Active Component, U.S. Armed Forces, 2002–2017. MSMR 2018, 25, 20–25. [Google Scholar]
- Alberti, G. Introduction to the Metabolic Syndrome. Eur. Heart J. Suppl. 2005, 7, D3–D5. [Google Scholar] [CrossRef]
- Marangos, P.J.; Okamoto, L.J.; Caro, J.J. Economic Burden of the Components of the Metabolic Syndrome. In Handbook of Disease Burdens and Quality of Life Measures; Preedy, V.R., Watson, R.R., Eds.; Springer: New York, NY, USA, 2010; pp. 1135–1149. ISBN 978-0-387-78665-0. [Google Scholar]
- Jha, B.K.; Sherpa, M.L.; Imran, M.; Mohammed, Y.; Jha, L.A.; Paudel, K.R.; Jha, S.K. Progress in Understanding Metabolic Syndrome and Knowledge of Its Complex Pathophysiology. Diabetology 2023, 4, 134–159. [Google Scholar] [CrossRef]
- Mendrick, D.L.; Diehl, A.M.; Topor, L.S.; Dietert, R.R.; Will, Y.; La Merrill, M.A.; Bouret, S.; Varma, V.; Hastings, K.L.; Schug, T.T.; et al. Metabolic Syndrome and Associated Diseases: From the Bench to the Clinic. Toxicol. Sci. 2018, 162, 36–42. [Google Scholar] [CrossRef]
- VAGUE, J. Sexual differentiation, Factor determining forms of obesity. Presse Med. 1947, 55, 339. [Google Scholar]
- Avogaro, P.; Crepaldi, G.; Enzi, G.; Tiengo, A. Metabolic aspects of essential obesity. Epatologia 1965, 11, 226–238. [Google Scholar]
- O’Neill, S.; O’Driscoll, L. Metabolic Syndrome: A Closer Look at the Growing Epidemic and Its Associated Pathologies. Obes. Rev. 2015, 16, 1–12. [Google Scholar] [CrossRef]
- Reaven, G.M. Role of Insulin Resistance in Human Disease. Diabetes 1988, 37, 1595–1607. [Google Scholar] [CrossRef]
- Kaplan, N.M. The Deadly Quartet. Upper-Body Obesity, Glucose Intolerance, Hypertriglyceridemia, and Hypertension. Arch. Intern. Med. 1989, 149, 1514–1520. [Google Scholar] [CrossRef]
- Haffner, S.M.; Valdez, R.A.; Hazuda, H.P.; Mitchell, B.D.; Morales, P.A.; Stern, M.P. Prospective Analysis of the Insulin-Resistance Syndrome (Syndrome X). Diabetes 1992, 41, 715–722. [Google Scholar] [CrossRef]
- Alberti, K.G.M.M.; Zimmet, P.Z. Definition, Diagnosis and Classification of Diabetes Mellitus and Its Complications. Part 1: Diagnosis and Classification of Diabetes Mellitus. Provisional Report of a WHO Consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Alberti, K.G.M.M.; Zimmet, P.; Shaw, J. Metabolic Syndrome—A New World-Wide Definition. A Consensus Statement from the International Diabetes Federation. Diabet. Med. 2006, 23, 469–480. [Google Scholar] [CrossRef]
- Balkau, B.; Charles, M.A. Comment on the Provisional Report from the WHO Consultation. Diabet. Med. 1999, 16, 442–443. [Google Scholar] [CrossRef]
- Einhorn, D.; Reaven, G.M.; Cobin, R.H.; Ford, E.; Ganda, O.P.; Handelsman, Y.; Hellman, R.; Jellinger, P.S.; Kendall, D.; Krauss, R.M.; et al. American College of Endocrinology Position Statement on the Insulin Resistance Syndrome. Endocr. Pract. 2003, 9, 237–252. [Google Scholar] [CrossRef]
- Wang, W.; Hu, M.; Liu, H.; Zhang, X.; Li, H.; Zhou, F.; Liu, Y.-M.; Lei, F.; Qin, J.-J.; Zhao, Y.-C.; et al. Global Burden of Disease Study 2019 Suggests That Metabolic Risk Factors Are the Leading Drivers of the Burden of Ischemic Heart Disease. Cell Metab. 2021, 33, 1943–1956.e2. [Google Scholar] [CrossRef]
- Ghosh, S.; Dhar, S.; Bhattacharjee, S.; Bhattacharjee, P. Contribution of Environmental, Genetic and Epigenetic Factors to Obesity-Related Metabolic Syndrome. Nucleus 2023, 66, 215–237. [Google Scholar] [CrossRef]
- Noubiap, J.J.; Nansseu, J.R.; Lontchi-Yimagou, E.; Nkeck, J.R.; Nyaga, U.F.; Ngouo, A.T.; Tounouga, D.N.; Tianyi, F.-L.; Foka, A.J.; Ndoadoumgue, A.L.; et al. Geographic Distribution of Metabolic Syndrome and Its Components in the General Adult Population: A Meta-Analysis of Global Data from 28 Million Individuals. Diabetes Res. Clin. Pract. 2022, 188, 109924. [Google Scholar] [CrossRef]
- do Vale Moreira, N.C.; Hussain, A.; Bhowmik, B.; Mdala, I.; Siddiquee, T.; Fernandes, V.O.; Montenegro Júnior, R.M.; Meyer, H.E. Prevalence of Metabolic Syndrome by Different Definitions, and Its Association with Type 2 Diabetes, Pre-Diabetes, and Cardiovascular Disease Risk in Brazil. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 1217–1224. [Google Scholar] [CrossRef]
- Li, W.; Qiu, X.; Ma, H.; Geng, Q. Incidence and Long-Term Specific Mortality Trends of Metabolic Syndrome in the United States. Front. Endocrinol. 2022, 13, 1029736. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Jia, W. Multi-Omics Profiling: The Way towards Precision Medicine in Metabolic Diseases. J. Mol. Cell Biol. 2021, 13, 576–593. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Turner, N. Mitochondrial Dysfunction and Insulin Resistance: An Update. Endocr. Connect. 2015, 4, R1–R15. [Google Scholar] [CrossRef] [PubMed]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-Mediated Dysbiosis Regulates Progression of NAFLD and Obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef]
- Toubal, A.; Treuter, E.; Clément, K.; Venteclef, N. Genomic and Epigenomic Regulation of Adipose Tissue Inflammation in Obesity. Trends Endocrinol. Metab. 2013, 24, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Nieuwdorp, M.; Stroes, E.S.G.; Meijers, J.C.M.; Büller, H. Hypercoagulability in the Metabolic Syndrome. Curr. Opin. Pharmacol. 2005, 5, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Cano-Sancho, G.; Salmon, A.G.; La Merrill, M.A. Association between Exposure to p,p’-DDT and Its Metabolite p,p’-DDE with Obesity: Integrated Systematic Review and Meta-Analysis. Environ. Health Perspect. 2017, 125, 96002. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A Metagenome-Wide Association Study of Gut Microbiota in Type 2 Diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Warrier, M.; Shih, D.M.; Burrows, A.C.; Ferguson, D.; Gromovsky, A.D.; Brown, A.L.; Marshall, S.; McDaniel, A.; Schugar, R.C.; Wang, Z.; et al. The TMAO-Generating Enzyme Flavin Monooxygenase 3 Is a Central Regulator of Cholesterol Balance. Cell Rep. 2015, 10, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Halpern, A.; Mancini, M.C.; Magalhães, M.E.C.; Fisberg, M.; Radominski, R.; Bertolami, M.C.; Bertolami, A.; de Melo, M.E.; Zanella, M.T.; Queiroz, M.S.; et al. Metabolic Syndrome, Dyslipidemia, Hypertension and Type 2 Diabetes in Youth: From Diagnosis to Treatment. Diabetol. Metab. Syndr. 2010, 2, 55. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.C.; Brunzell, J.D. Abdominal Obesity and Dyslipidemia in the Metabolic Syndrome: Importance of Type 2 Diabetes and Familial Combined Hyperlipidemia in Coronary Artery Disease Risk. J. Clin. Endocrinol. Metab. 2004, 89, 2601–2607. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alegría, K.; Arias, C. Functional Consequences of Brain Exposure to Saturated Fatty Acids: From Energy Metabolism and Insulin Resistance to Neuronal Damage. Endocrinol. Diabetes Metab. 2023, 6, e386. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Boppana, A.; Traupman, J.A.; Unson, E.; Maddock, D.A.; Chao, K.; Dobesh, D.P.; Brufsky, A.; Connor, R.I. Impaired Glucose Metabolism in Patients with Diabetes, Prediabetes, and Obesity Is Associated with Severe COVID-19. J. Med. Virol. 2021, 93, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Harlim, A.; Bontong, S.I.; Rahfiludin, M.Z. Relationship between Carbohydrate, Protein, and Fat Consumption with Obesity Incidence in 2014 UKI Faculty of Medicine Students. Amerta Nutr. 2022, 6, 385–391. [Google Scholar] [CrossRef]
- Ussar, S.; Fujisaka, S.; Kahn, C.R. Interactions between Host Genetics and Gut Microbiome in Diabetes and Metabolic Syndrome. Mol. Metab. 2016, 5, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Neumark-Sztainer, D.; Story, M.; Perry, C.; Casey, M.A. Factors Influencing Food Choices of Adolescents: Findings from Focus-Group Discussions with Adolescents. J. Am. Diet. Assoc. 1999, 99, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Kartashov, A.I.; Ebbeling, C.B.; Van Horn, L.; Slattery, M.L.; Jacobs, D.R.; Ludwig, D.S. Fast-Food Habits, Weight Gain, and Insulin Resistance (the CARDIA Study): 15-Year Prospective Analysis. Lancet 2005, 365, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Rouhani, M.H.; Mirseifinezhad, M.; Omrani, N.; Esmaillzadeh, A.; Azadbakht, L. Fast Food Consumption, Quality of Diet, and Obesity among Isfahanian Adolescent Girls. J. Obes. 2012, 2012, 597924. [Google Scholar] [CrossRef] [PubMed]
- Alexander, L.; Christensen, S.M.; Richardson, L.; Ingersoll, A.B.; Burridge, K.; Golden, A.; Karjoo, S.; Cortez, D.; Shelver, M.; Bays, H.E. Nutrition and Physical Activity: An Obesity Medicine Association (OMA) Clinical Practice Statement 2022. Obes. Pillars 2022, 1, 100005. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, Z.; Wang, F.; Mu, H.; Guo, L.; Xiao, J.; Liu, Y.; Li, X. Formation of Starch-Lipid Complexes during the Deep-Frying Process and Its Effects on Lipid Oxidation. Foods 2022, 11, 3083. [Google Scholar] [CrossRef] [PubMed]
- Bordin, K.; Kunitake, M.T.; Aracava, K.K.; Silvia, C.; Trindade, F. Changes in Food Caused by Deep Fat Frying—A Review. Arch. Latinoam. De Nutr. 2013, 63, 5–13. [Google Scholar]
- Ganesan, K.; Xu, B. Deep Frying Cooking Oils Promote the High Risk of Metastases in the Breast-A Critical Review. Food Chem. Toxicol. 2020, 144, 111648. [Google Scholar] [CrossRef]
- Choe, E.; Min, D.B. Chemistry of Deep-Fat Frying Oils. J. Food Sci. 2007, 72, R77–R86. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R. Advanced Lipoxidation End-Products. Chem. Biol. Interact. 2011, 192, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chao, C.; Cai, J.; Niu, B.; Copeland, L.; Wang, S. Starch–Lipid and Starch–Lipid–Protein Complexes: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1056–1079. [Google Scholar] [CrossRef]
- Sharma, C.; Kaur, A.; Thind, S.S.; Singh, B.; Raina, S. Advanced Glycation End-Products (AGEs): An Emerging Concern for Processed Food Industries. J. Food Sci. Technol. 2015, 52, 7561–7576. [Google Scholar] [CrossRef]
- Liauchonak, I.; Qorri, B.; Dawoud, F.; Riat, Y.; Szewczuk, M.R. Non-Nutritive Sweeteners and Their Implications on the Development of Metabolic Syndrome. Nutrients 2019, 11, 644. [Google Scholar] [CrossRef] [PubMed]
- Maharat, M.; Rahimlou, M.; Sioofi, A.; Sajjadi, S.F.; Moosavian, S.P. Association of Major Dietary Patterns with Socioeconomic Status, Obesity, and Contracting COVID-19 among Iranian Adults. Front. Nutr. 2024, 11, 1301634. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [PubMed]
- James, W.P.T. WHO Recognition of the Global Obesity Epidemic. Int. J. Obes. 2008, 32, S120–S126. [Google Scholar] [CrossRef] [PubMed]
- Lathigara, D.; Kaushal, D.; Wilson, R.B. Molecular Mechanisms of Western Diet-Induced Obesity and Obesity-Related Carcinogenesis—A Narrative Review. Metabolites 2023, 13, 675. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Liu, S.; Zhang, C. The Related Metabolic Diseases and Treatments of Obesity. Healthcare 2022, 10, 1616. [Google Scholar] [CrossRef]
- Ward, Z.J.; Bleich, S.N.; Cradock, A.L.; Barrett, J.L.; Giles, C.M.; Flax, C.; Long, M.W.; Gortmaker, S.L. Projected U.S. State-Level Prevalence of Adult Obesity and Severe Obesity. N. Engl. J. Med. 2019, 381, 2440–2450. [Google Scholar] [CrossRef] [PubMed]
- Boutari, C.; Mantzoros, C.S. A 2022 Update on the Epidemiology of Obesity and a Call to Action: As Its Twin COVID-19 Pandemic Appears to Be Receding, the Obesity and Dysmetabolism Pandemic Continues to Rage On. Metabolism 2022, 133, 155217. [Google Scholar] [CrossRef] [PubMed]
- Lobstein, T.; Jackson-Leach, R.; Powis, J.; Brinsden, H.; Gray, M. World Obesity Atlas 2023; World Obesity Federation: London, UK, 2023; pp. 5–25. [Google Scholar]
- Venkatesan, R.; Mohan, V. Obesity—Are We Continuing to Play the Genetic “Blame Game”? Adv. Genom. Genet. 2016, 6, 11–23. [Google Scholar] [CrossRef]
- Scuteri, A.; Sanna, S.; Chen, W.-M.; Uda, M.; Albai, G.; Strait, J.; Najjar, S.; Nagaraja, R.; Orrú, M.; Usala, G.; et al. Genome-Wide Association Scan Shows Genetic Variants in the FTO Gene Are Associated with Obesity-Related Traits. PLoS Genet. 2007, 3, e115. [Google Scholar] [CrossRef] [PubMed]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of Published Genome-Wide Association Studies, Targeted Arrays and Summary Statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [PubMed]
- Pigeyre, M.; Yazdi, F.T.; Kaur, Y.; Meyre, D. Recent Progress in Genetics, Epigenetics and Metagenomics Unveils the Pathophysiology of Human Obesity. Clin. Sci. 2016, 130, 943–986. [Google Scholar] [CrossRef] [PubMed]
- van der Klaauw, A.A.; Farooqi, I.S. The Hunger Genes: Pathways to Obesity. Cell 2015, 161, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.F.; Yeo, G.S.H. The Genetics of Obesity: From Discovery to Biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef]
- Hinney, A.; Körner, A.; Fischer-Posovszky, P. The Promise of New Anti-Obesity Therapies Arising from Knowledge of Genetic Obesity Traits. Nat. Rev. Endocrinol. 2022, 18, 623–637. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Obesity, Metabolic Syndrome, and Cardiovascular Disease. J. Clin. Endocrinol. Metab. 2004, 89, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Choquet, H.; Meyre, D. Genetics of Obesity: What Have We Learned? Curr. Genom. 2011, 12, 169–179. [Google Scholar] [CrossRef]
- Frayling, T.M. Genome-Wide Association Studies Provide New Insights into Type 2 Diabetes Aetiology. Nat. Rev. Genet. 2007, 8, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Duis, J.; Butler, M.G. Syndromic and Nonsyndromic Obesity: Underlying Genetic Causes in Humans. Adv. Biol. 2022, 6, 2101154. [Google Scholar] [CrossRef] [PubMed]
- Şanlı, E.; Kabaran, S. Maternal Obesity, Maternal Overnutrition and Fetal Programming: Effects of Epigenetic Mechanisms on the Development of Metabolic Disorders. Curr. Genom. 2019, 20, 419–427. [Google Scholar] [CrossRef]
- Simmonds, M.; Llewellyn, A.; Owen, C.G.; Woolacott, N. Predicting Adult Obesity from Childhood Obesity: A Systematic Review and Meta-Analysis. Obes. Rev. 2016, 17, 95–107. [Google Scholar] [CrossRef]
- Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2014, 37 (Suppl. S1), S81–S90. [CrossRef] [PubMed]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2018. Diabetes Care 2018, 41, S13–S27. [Google Scholar] [CrossRef] [PubMed]
- Sanyaolu, A.; Okorie, C.; Qi, X.; Locke, J.; Rehman, S. Childhood and Adolescent Obesity in the United States: A Public Health Concern. Glob. Pediatr. Health 2019, 6, 2333794X19891305. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.G.; Williamson, D.F.; Glick, H.A. Direct Medical Cost of Overweight and Obesity in the USA: A Quantitative Systematic Review. Obes. Rev. 2011, 12, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Karamanou, M.; Protogerou, A.; Tsoucalas, G.; Androutsos, G.; Poulakou-Rebelakou, E. Milestones in the History of Diabetes Mellitus: The Main Contributors. World J. Diabetes 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.-F. Global, Regional, and National Burden and Trend of Diabetes in 195 Countries and Territories: An Analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef]
- Kahaly, G.J.; Hansen, M.P. Type 1 Diabetes Associated Autoimmunity. Autoimmun. Rev. 2016, 15, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Leahy, J.L. Pathogenesis of Type 2 Diabetes Mellitus. Arch. Med. Res. 2005, 36, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.M.; Contreras, R.; Chen, W.; Sacks, D.A. Trends in the Prevalence of Preexisting Diabetes and Gestational Diabetes Mellitus among a Racially/Ethnically Diverse Population of Pregnant Women, 1999–2005. Diabetes Care 2008, 31, 899–904. [Google Scholar] [CrossRef]
- Grimaldi, A.; Dudek, A.; Otto-Buczkowska, E. Secondary Diabetes. Med. Des. Mal. Metab. 2008, 2, 571. [Google Scholar] [CrossRef]
- Garvey, W.T.; Olefsky, J.M.; Griffin, J.; Hamman, R.F.; Kolterman, O.G. The Effect of Insulin Treatment on Insulin Secretion and Insulin Action in Type II Diabetes Mellitus. Diabetes 1985, 34, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.F.A.; Thorleifsson, G.; Reynisdottir, I.; Benediktsson, R.; Manolescu, A.; Sainz, J.; Helgason, A.; Stefansson, H.; Emilsson, V.; Helgadottir, A.; et al. Variant of Transcription Factor 7-like 2 (TCF7L2) Gene Confers Risk of Type 2 Diabetes. Nat. Genet. 2006, 38, 320–323. [Google Scholar] [CrossRef]
- Helgason, A.; Pálsson, S.; Thorleifsson, G.; Grant, S.F.A.; Emilsson, V.; Gunnarsdottir, S.; Adeyemo, A.; Chen, Y.; Chen, G.; Reynisdottir, I.; et al. Refining the Impact of TCF7L2 Gene Variants on Type 2 Diabetes and Adaptive Evolution. Nat. Genet. 2007, 39, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of Diabetes: An Overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, P.R.; Kahn, B.B. Glucose Transporters and Insulin Action—Implications for Insulin Resistance and Diabetes Mellitus. N. Engl. J. Med. 1999, 341, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, A.; Mittelman, S.D.; Lamarche, B.; Bergman, R.N.; Giacca, A.; Lewis, G.F. Acute Enhancement of Insulin Secretion by FFA in Humans Is Lost with Prolonged FFA Elevation. Am. J. Physiol. 1999, 276, E1055–E1066. [Google Scholar] [CrossRef] [PubMed]
- Lotta, L.A.; Gulati, P.; Day, F.R.; Payne, F.; Ongen, H.; van de Bunt, M.; Gaulton, K.J.; Eicher, J.D.; Sharp, S.J.; Luan, J.; et al. Integrative Genomic Analysis Implicates Limited Peripheral Adipose Storage Capacity in the Pathogenesis of Human Insulin Resistance. Nat. Genet. 2017, 49, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Biddinger, S.B.; Hernandez-Ono, A.; Rask-Madsen, C.; Haas, J.T.; Alemán, J.O.; Suzuki, R.; Scapa, E.F.; Agarwal, C.; Carey, M.C.; Stephanopoulos, G.; et al. Hepatic Insulin Resistance Is Sufficient to Produce Dyslipidemia and Susceptibility to Atherosclerosis. Cell Metab. 2008, 7, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Pories, W.J.; Dohm, G.L. Diabetes: Have We Got It All Wrong? Hyperinsulinism as the Culprit: Surgery Provides the Evidence. Diabetes Care 2012, 35, 2438–2442. [Google Scholar] [CrossRef] [PubMed]
- da Silva Rosa, S.C.; Nayak, N.; Caymo, A.M.; Gordon, J.W. Mechanisms of Muscle Insulin Resistance and the Cross-Talk with Liver and Adipose Tissue. Physiol. Rep. 2020, 8, e14607. [Google Scholar] [CrossRef] [PubMed]
- Pickering, R.J.; Rosado, C.J.; Sharma, A.; Buksh, S.; Tate, M.; de Haan, J.B. Recent Novel Approaches to Limit Oxidative Stress and Inflammation in Diabetic Complications. Clin. Transl. Immunol. 2018, 7, e1016. [Google Scholar] [CrossRef] [PubMed]
- Ojha, A.; Ojha, U.; Mohammed, R.; Chandrashekar, A.; Ojha, H. Current Perspective on the Role of Insulin and Glucagon in the Pathogenesis and Treatment of Type 2 Diabetes Mellitus. Clin. Pharmacol. 2019, 11, 57–65. [Google Scholar] [CrossRef]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 440649. [Google Scholar] [CrossRef]
- Lo, C.-M.; Obici, S.; Dong, H.H.; Haas, M.; Lou, D.; Kim, D.H.; Liu, M.; D’Alessio, D.; Woods, S.C.; Tso, P. Impaired Insulin Secretion and Enhanced Insulin Sensitivity in Cholecystokinin-Deficient Mice. Diabetes 2011, 60, 2000–2007. [Google Scholar] [CrossRef] [PubMed]
- Hackl, M.T.; Fürnsinn, C.; Schuh, C.M.; Krssak, M.; Carli, F.; Guerra, S.; Freudenthaler, A.; Baumgartner-Parzer, S.; Helbich, T.H.; Luger, A.; et al. Brain Leptin Reduces Liver Lipids by Increasing Hepatic Triglyceride Secretion and Lowering Lipogenesis. Nat. Commun. 2019, 10, 2717. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.; Novick, D.; Rubinstein, M. Modulation of Insulin Activities by Leptin. Science 1996, 274, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, E.E.; Flier, J.S. Adipose Tissue as an Endocrine Organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Katsiki, N.; Mikhailidis, D.P.; Banach, M. Leptin, Cardiovascular Diseases and Type 2 Diabetes Mellitus. Acta Pharmacol. Sin. 2018, 39, 1176–1188. [Google Scholar] [CrossRef] [PubMed]
- Puri, V.; Ranjit, S.; Konda, S.; Nicoloro, S.M.C.; Straubhaar, J.; Chawla, A.; Chouinard, M.; Lin, C.; Burkart, A.; Corvera, S.; et al. Cidea Is Associated with Lipid Droplets and Insulin Sensitivity in Humans. Proc. Natl. Acad. Sci. USA 2008, 105, 7833–7838. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, A.R.; Sztalryd, C. The Perilipins: Major Cytosolic Lipid Droplet-Associated Proteins and Their Roles in Cellular Lipid Storage, Mobilization, and Systemic Homeostasis. Annu. Rev. Nutr. 2016, 36, 471–509. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bai, Y.; Jin, Z.; Svensson, B. Food-Derived Non-Phenolic α-Amylase and α-Glucosidase Inhibitors for Controlling Starch Digestion Rate and Guiding Diabetes-Friendly Recipes. LWT 2022, 153, 112455. [Google Scholar] [CrossRef]
- Del Prete, A.; Iadevaia, M.; Loguercio, C. The Role of Gut Hormones in Controlling the Food Intake. What Is Their Role in Emerging Diseases? Endocrinol. Y Nutr. (Engl. Ed.) 2012, 59, 197–206. [Google Scholar] [CrossRef]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and Insulin Resistance. J. Clin. Invest. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Oxidative Stress and Stress-Activated Signaling Pathways: A Unifying Hypothesis of Type 2 Diabetes. Endocr. Rev. 2002, 23, 599–622. [Google Scholar] [CrossRef] [PubMed]
- Fowler, M.J. Microvascular and Macrovascular Complications of Diabetes. Clin. Diabetes 2008, 26, 77–82. [Google Scholar] [CrossRef]
- Bielka, W.; Przezak, A.; Pawlik, A. The Role of the Gut Microbiota in the Pathogenesis of Diabetes. Int. J. Mol. Sci. 2022, 23, 480. [Google Scholar] [CrossRef] [PubMed]
- Gradisteanu Pircalabioru, G.; Corcionivoschi, N.; Gundogdu, O.; Chifiriuc, M.-C.; Marutescu, L.G.; Ispas, B.; Savu, O. Dysbiosis in the Development of Type I Diabetes and Associated Complications: From Mechanisms to Targeted Gut Microbes Manipulation Therapies. Int. J. Mol. Sci. 2021, 22, 2763. [Google Scholar] [CrossRef] [PubMed]
- Iatcu, C.O.; Steen, A.; Covasa, M. Gut Microbiota and Complications of Type-2 Diabetes. Nutrients 2021, 14, 166. [Google Scholar] [CrossRef] [PubMed]
- Kang, Q.; Yang, C. Oxidative Stress and Diabetic Retinopathy: Molecular Mechanisms, Pathogenetic Role and Therapeutic Implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef] [PubMed]
- Naveen, J.; Baskaran, V. Antidiabetic Plant-Derived Nutraceuticals: A Critical Review. Eur. J. Nutr. 2018, 57, 1275–1299. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.; Jacobs, K.; Haucke, E.; Navarrete Santos, A.; Grune, T.; Simm, A. Role of Advanced Glycation End Products in Cellular Signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Geissler, S.; Hellwig, M.; Zwarg, M.; Markwardt, F.; Henle, T.; Brandsch, M. Transport of the Advanced Glycation End Products Alanylpyrraline and Pyrralylalanine by the Human Proton-Coupled Peptide Transporter HPEPT1. J. Agric. Food Chem. 2010, 58, 2543–2547. [Google Scholar] [CrossRef]
- Tessier, F.J.; Niquet-Léridon, C.; Jacolot, P.; Jouquand, C.; Genin, M.; Schmidt, A.-M.; Grossin, N.; Boulanger, E. Quantitative Assessment of Organ Distribution of Dietary Protein-Bound 13C-Labeled Nɛ-Carboxymethyllysine after a Chronic Oral Exposure in Mice. Mol. Nutr. Food Res. 2016, 60, 2446–2456. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, J.P.; McKinley, B.; Eckel, R.H. The Metabolic Syndrome and Inflammation. Metab. Syndr. Relat. Disord. 2004, 2, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Awazu, M.; Tamiya, M.; Iwasaki, Y.; Harada, A.; Kugisaki, S.; Tanimura, S.; Kohno, M. Targeting the ERK Signaling Pathway as a Potential Treatment for Insulin Resistance and Type 2 Diabetes. Am. J. Physiol. Metab. 2016, 310, E643–E651. [Google Scholar] [CrossRef] [PubMed]
- Sergi, D.; Williams, L.M. Potential Relationship between Dietary Long-Chain Saturated Fatty Acids and Hypothalamic Dysfunction in Obesity. Nutr. Rev. 2020, 78, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, D.; Zhang, X.; Hastoy, B.; Clark, A. The β-Cell Assassin: IAPP Cytotoxicity. J. Mol. Endocrinol. 2017, 59, R121–R140. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, J.; Yang, Y.; Zhang, X. An Update on the Potential Role of Advanced Glycation End Products in Glycolipid Metabolism. Life Sci. 2020, 245, 117344. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Y.; Shen, Y.; Zhang, Y.; Liu, L.; Yang, X. Dietary Polyphenols: Regulate the Advanced Glycation End Products-RAGE Axis and the Microbiota-Gut-Brain Axis to Prevent Neurodegenerative Diseases. Crit. Rev. Food Sci. Nutr. 2023, 63, 9816–9842. [Google Scholar] [CrossRef] [PubMed]
- Hofnagel, O.; Luechtenborg, B.; Weissen-Plenz, G.; Robenek, H. Statins and Foam Cell Formation: Impact on LDL Oxidation and Uptake of Oxidized Lipoproteins via Scavenger Receptors. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2007, 1771, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Kakino, A.; Takeshita, H.; Hayashi, N.; Li, L.; Nakano, A.; Hanasaki-Yamamoto, H.; Fujita, Y.; Imaizumi, Y.; Toyama-Yokoyama, S.; et al. Oxidized LDL (OxLDL) Activates the Angiotensin II Type 1 Receptor by Binding to the Lectin-like OxLDL Receptor. FASEB J. 2015, 29, 3342–3356. [Google Scholar] [CrossRef] [PubMed]
- Karar, J.; Maity, A. PI3K/AKT/MTOR Pathway in Angiogenesis. Front. Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- Arivazhagan, L.; López-Díez, R.; Shekhtman, A.; Ramasamy, R.; Schmidt, A.M. Glycation and a Spark of ALEs (Advanced Lipoxidation End Products)—Igniting RAGE/Diaphanous-1 and Cardiometabolic Disease. Front. Cardiovasc. Med. 2022, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cai, W.; Yu, J.; Liu, H.; He, S.; Zhu, L.; Xu, J. Dietary Advanced Glycation End Products Shift the Gut Microbiota Composition and Induce Insulin Resistance in Mice. Diabetes. Metab. Syndr. Obes. 2022, 15, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Inan-Eroglu, E.; Ayaz, A.; Buyuktuncer, Z. Formation of Advanced Glycation Endproducts in Foods during Cooking Process and Underlying Mechanisms: A Comprehensive Review of Experimental Studies. Nutr. Res. Rev. 2020, 33, 77–89. [Google Scholar] [CrossRef]
- Takeuchi, M.; Sakasai-Sakai, A.; Takata, T.; Takino, J.; Koriyama, Y.; Kikuchi, C.; Furukawa, A.; Nagamine, K.; Hori, T.; Matsunaga, T. Intracellular Toxic AGEs (TAGE) Triggers Numerous Types of Cell Damage. Biomolecules 2021, 11, 387. [Google Scholar] [CrossRef]
- Salazar, J.; Navarro, C.; Ortega, Á.; Nava, M.; Morillo, D.; Torres, W.; Hernández, M.; Cabrera, M.; Angarita, L.; Ortiz, R.; et al. Advanced Glycation End Products: New Clinical and Molecular Perspectives. Int. J. Environ. Res. Public Health 2021, 18, 7236. [Google Scholar] [CrossRef]
- Dong, L.; Li, Y.; Chen, Q.; Liu, Y.; Qiao, Z.; Sang, S.; Zhang, J.; Zhan, S.; Wu, Z.; Liu, L. Research Advances of Advanced Glycation End Products in Milk and Dairy Products: Formation, Determination, Control Strategy and Immunometabolism via Gut Microbiota. Food Chem. 2023, 417, 135861. [Google Scholar] [CrossRef] [PubMed]
- Sacks, F.M.; Lichtenstein, A.H.; Wu, J.H.Y.; Appel, L.J.; Creager, M.A.; Kris-Etherton, P.M.; Miller, M.; Rimm, E.B.; Rudel, L.L.; Robinson, J.G.; et al. Dietary Fats and Cardiovascular Disease: A Presidential Advisory from the American Heart Association. Circulation 2017, 136, e1–e23. [Google Scholar] [CrossRef] [PubMed]
- Dybiec, J.; Baran, W.; Dąbek, B.; Fularski, P.; Młynarska, E.; Radzioch, E.; Rysz, J.; Franczyk, B. Advances in Treatment of Dyslipidemia. Int. J. Mol. Sci. 2023, 24, 13288. [Google Scholar] [CrossRef] [PubMed]
- Arvanitis, M.; Lowenstein, C.J. Dyslipidemia. Ann. Intern. Med. 2023, 176, ITC81–ITC96. [Google Scholar] [CrossRef] [PubMed]
- Rosenson, R.S.; Najera, S.D.; Hegele, R.A. Heterozygous Familial Hypercholesterolemia Presenting as Chylomicronemia Syndrome. J. Clin. Lipidol. 2017, 11, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Pirillo, A.; Casula, M.; Olmastroni, E.; Norata, G.D.; Catapano, A.L. Global Epidemiology of Dyslipidaemias. Nat. Rev. Cardiol. 2021, 18, 689–700. [Google Scholar] [CrossRef]
- Schwarz, J.-M.; Linfoot, P.; Dare, D.; Aghajanian, K. Hepatic de Novo Lipogenesis in Normoinsulinemic and Hyperinsulinemic Subjects Consuming High-Fat, Low-Carbohydrate and Low-Fat, High-Carbohydrate Isoenergetic Diets123. Am. J. Clin. Nutr. 2003, 77, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L.; Schwarz, J.M.; Keim, N.L.; Griffen, S.C.; Bremer, A.A.; Graham, J.L.; Hatcher, B.; Cox, C.L.; Dyachenko, A.; Zhang, W.; et al. Consuming Fructose-Sweetened, Not Glucose-Sweetened, Beverages Increases Visceral Adiposity and Lipids and Decreases Insulin Sensitivity in Overweight/Obese Humans. J. Clin. Invest. 2009, 119, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Strable, M.S.; Ntambi, J.M. Genetic Control of de Novo Lipogenesis: Role in Diet-Induced Obesity. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Sears, B.; Perry, M. The Role of Fatty Acids in Insulin Resistance. Lipids Health Dis. 2015, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.R.; Pissios, P.; Otu, H.; Roberson, R.; Xue, B.; Asakura, K.; Furukawa, N.; Marino, F.E.; Liu, F.-F.; Kahn, B.B.; et al. A High-Fat, Ketogenic Diet Induces a Unique Metabolic State in Mice. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1724–E1739. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.M.; Botezelli, J.D.; da Cruz Rodrigues, K.C.; Mekary, R.A.; Cintra, D.E.; Pauli, J.R.; da Silva, A.S.R.; Ropelle, E.R.; de Moura, L.P. Fructose Consumption in the Development of Obesity and the Effects of Different Protocols of Physical Exercise on the Hepatic Metabolism. Nutrients 2017, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Ameer, F.; Scandiuzzi, L.; Hasnain, S.; Kalbacher, H.; Zaidi, N. De Novo Lipogenesis in Health and Disease. Metabolism 2014, 63, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Herman, M.A.; Birnbaum, M.J. Molecular Aspects of Fructose Metabolism and Metabolic Disease. Cell Metab. 2021, 33, 2329–2354. [Google Scholar] [CrossRef] [PubMed]
- Fraser, F.; Corstorphine, C.G.; Zammit, V.A. Topology of Carnitine Palmitoyltransferase I in the Mitochondrial Outer Membrane. Biochem. J. 1997, 323 Pt. 3, 711–718. [Google Scholar] [CrossRef]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced Glycation End Products and Oxidative Stress in Type 2 Diabetes Mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef]
- Ganda, O.P. Triglyceride-Rich Lipoproteins, Remnant-Cholesterol, and Atherosclerotic Cardiovascular Disease. Curr. Opin. Lipidol. 2023, 34, 105–113. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Chen, Y.; Wang, Z.; He, H.; Yu, P. Cellular Uptake, Metabolism and Sensing of Long-Chain Fatty Acids. Front. Biosci. Landmark 2023, 28, 10. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, A.; Aryal, P.; Wen, J.; Syed, I.; Vazirani, R.P.; Moraes-Vieira, P.M.; Camporez, J.P.; Gallop, M.R.; Perry, R.J.; Peroni, O.D.; et al. Absence of Carbohydrate Response Element Binding Protein in Adipocytes Causes Systemic Insulin Resistance and Impairs Glucose Transport. Cell Rep. 2017, 21, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Aarsland, A.; Chinkes, D.; Wolfe, R.R. Hepatic and Whole-Body Fat Synthesis in Humans during Carbohydrate Overfeeding. Am. J. Clin. Nutr. 1997, 65, 1774–1782. [Google Scholar] [CrossRef] [PubMed]
- Bolsoni-Lopes, A.; Alonso-Vale, M.I.C. Lipolysis and Lipases in White Adipose Tissue—An Update. Arch. Endocrinol. Metab. 2015, 59, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Barrows, B.R.; Parks, E.J. Contributions of Different Fatty Acid Sources to Very Low-Density Lipoprotein-Triacylglycerol in the Fasted and Fed States. J. Clin. Endocrinol. Metab. 2006, 91, 1446–1452. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, K.L.; Smith, C.I.; Schwarzenberg, S.J.; Jessurun, J.; Boldt, M.D.; Parks, E.J. Sources of Fatty Acids Stored in Liver and Secreted via Lipoproteins in Patients with Nonalcoholic Fatty Liver Disease. J. Clin. Invest. 2005, 115, 1343–1351. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.E.; Ramos-Roman, M.A.; Browning, J.D.; Parks, E.J. Increased de Novo Lipogenesis Is a Distinct Characteristic of Individuals with Nonalcoholic Fatty Liver Disease. Gastroenterology 2014, 146, 726–735. [Google Scholar] [CrossRef]
- Kim, J.Y.; Nasr, A.; Tfayli, H.; Bacha, F.; Michaliszyn, S.F.; Arslanian, S. Increased Lipolysis, Diminished Adipose Tissue Insulin Sensitivity, and Impaired β-Cell Function Relative to Adipose Tissue Insulin Sensitivity in Obese Youth With Impaired Glucose Tolerance. Diabetes 2017, 66, 3085–3090. [Google Scholar] [CrossRef] [PubMed]
- Morigny, P.; Houssier, M.; Mouisel, E.; Langin, D. Adipocyte Lipolysis and Insulin Resistance. Biochimie 2016, 125, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Roden, M. Mechanisms of Disease: Hepatic Steatosis in Type 2 Diabetes—Pathogenesis and Clinical Relevance. Nat. Clin. Pract. Endocrinol. Metab. 2006, 2, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Milner, K.; Gupta, A.; Lopaschuk, D.G. Myocardial Energy Substrate Metabolism in Heart Failure: From Pathways to Therapeutic Targets. Curr. Pharm. Des. 2015, 21, 3654–3664. [Google Scholar] [CrossRef] [PubMed]
- Mato, J.M.; Alonso, C.; Noureddin, M.; Lu, S.C. Biomarkers and Subtypes of Deranged Lipid Metabolism in Non-Alcoholic Fatty Liver Disease. World J. Gastroenterol. 2019, 25, 3009–3020. [Google Scholar] [CrossRef] [PubMed]
- Mach, F.; Baigent, C.; Catapano, A.L.; Koskinas, K.C.; Casula, M.; Badimon, L.; Chapman, M.J.; De Backer, G.G.; Delgado, V.; Ference, B.A.; et al. 2019 ESC/EAS Guidelines for the Management of Dyslipidaemias: Lipid Modification to Reduce Cardiovascular Risk. Eur. Heart J. 2020, 41, 111–188. [Google Scholar] [CrossRef]
- Nair, P.A.; Patel, C.R.; Ganjiwale, J.D.; Diwan, N.G.; Jivani, N.B. Xanthelasma Palpebrarum with Arcus Cornea: A Clinical and Biochemical Study. Indian J. Dermatol. 2016, 61, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic Syndrome: Pathophysiology, Management, and Modulation by Natural Compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Dobrowolski, P.; Prejbisz, A.; Kuryłowicz, A.; Baska, A.; Burchardt, P.; Chlebus, K.; Dzida, G.; Jankowski, P.; Jaroszewicz, J.; Jaworski, P.; et al. Metabolic Syndrome—A New Definition and Management Guidelines. A joint position paper by the Polish Society of Hypertension, Polish Society for the Treatment of Obesity, Polish Lipid Association, Polish Association for Study of Liver, Polish Society of Family Medicine, Polish Society of Lifestyle Medicine, Division of Prevention and Epidemiology Polish Cardiac Society, “Club 30” Polish Cardiac Society, and Division of Metabolic and Bariatric Surgery Society of Polish Surgeons. Arch. Med. Sci. 2022, 18, 1133–1156. [Google Scholar] [CrossRef] [PubMed]
- Padwal, R.S.; Majumdar, S.R. Drug Treatments for Obesity: Orlistat, Sibutramine, and Rimonabant. Lancet 2007, 369, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Lillich, F.F.; Imig, J.D.; Proschak, E. Multi-Target Approaches in Metabolic Syndrome. Front. Pharmacol. 2021, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jimenez, F. Metabolic Syndrome—A Common and Dangerous Health Problem. US Cardiol. 2007, 4, 37–40. [Google Scholar] [CrossRef]
- Lim, R.; Beekley, A.; Johnson, D.C.; Davis, K.A. Early and Late Complications of Bariatric Operation. Trauma Surg. Acute Care Open 2018, 3, e000219. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.H.; Singh, R.B.; Mehta, V.; Sakshi; Asif, M.; Goyal, K.; Balodhi, A.; Manglik, P.; Sharma, A.; Chahal, A. Impact of Exercise Training Duration on Obesity and Cardiometabolic Biomarkers: A Systematic Review. J. Diabetes Metab. Disord. 2023, 22, 155–174. [Google Scholar] [CrossRef] [PubMed]
- Moschonis, G.; Trakman, G.L. Overweight and Obesity: The Interplay of Eating Habits and Physical Activity. Nutrients 2023, 15, 2896. [Google Scholar] [CrossRef] [PubMed]
- Parmar, R.M.; Can, A.S. Dietary Approaches to Obesity Treatment; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Lee, V. Introduction to the Dietary Management of Obesity in Adults. Clin. Med. J. R. Coll. Physicians Lond. 2023, 23, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Shaik Mohamed Sayed, U.F.; Moshawih, S.; Goh, H.P.; Kifli, N.; Gupta, G.; Singh, S.K.; Chellappan, D.K.; Dua, K.; Hermansyah, A.; Ser, H.L.; et al. Natural Products as Novel Anti-Obesity Agents: Insights into Mechanisms of Action and Potential for Therapeutic Management. Front. Pharmacol. 2023, 14, 1182937. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.; Ng, S.W.; Tan, J.Z.X.; Gupta, G.; Negi, P.; Thangavelu, L.; Balusamy, S.R.; Perumalsamy, H.; Yap, W.H.; Singh, S.K.; et al. Natural Products in the Management of Obesity: Fundamental Mechanisms and Pharmacotherapy. S. Afr. J. Bot. 2021, 143, 176–197. [Google Scholar] [CrossRef]
- Miller, E.G.; Nowson, C.A.; Dunstan, D.W.; Kerr, D.A.; Menzies, D.; Daly, R.M. Effects of Whey Protein plus Vitamin D Supplementation Combined with Progressive Resistance Training on Glycaemic Control, Body Composition, Muscle Function and Cardiometabolic Risk Factors in Middle-Aged and Older Overweight/Obese Adults with Type 2 Dia. Diabetes. Obes. Metab. 2021, 23, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, L.J.; Veronese, N.; Di Bella, G.; Cusumano, C.; Parisi, A.; Tagliaferri, F.; Ciriminna, S.; Barbagallo, M. Mediterranean Diet in the Management and Prevention of Obesity. Exp. Gerontol. 2023, 174, 112121. [Google Scholar] [CrossRef] [PubMed]
- Del Bo’, C.; Perna, S.; Allehdan, S.; Rafique, A.; Saad, S.; AlGhareeb, F.; Rondanelli, M.; Tayyem, R.F.; Marino, M.; Martini, D.; et al. Does the Mediterranean Diet Have Any Effect on Lipid Profile, Central Obesity and Liver Enzymes in Non-Alcoholic Fatty Liver Disease (NAFLD) Subjects? A Systematic Review and Meta-Analysis of Randomized Control Trials. Nutrients 2023, 15, 2250. [Google Scholar] [CrossRef] [PubMed]
- Hoyas, I.; Leon-Sanz, M. Nutritional Challenges in Metabolic Syndrome. J. Clin. Med. 2019, 8, 1301. [Google Scholar] [CrossRef] [PubMed]
- Castro-Barquero, S.; Ruiz-León, A.M.; Sierra-Pérez, M.; Estruch, R.; Casas, R. Dietary Strategies for Metabolic Syndrome: A Comprehensive Review. Nutrients 2020, 12, 2983. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.M.; Shalaby, M.A.; El-Shiekh, R.A.; El-Banna, H.A.; Emam, S.R.; Bakr, A.F. Metabolic Syndrome: Risk Factors, Diagnosis, Pathogenesis, and Management with Natural Approaches. Food Chem. Adv. 2023, 3, 100335. [Google Scholar] [CrossRef]
- Hsieh, C.C.; Hernández-Ledesma, B.; Fernández-Tomé, S.; Weinborn, V.; Barile, D.; De Moura Bell, J.M.L.N. Milk Proteins, Peptides, and Oligosaccharides: Effects against the 21st Century Disorders. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef]
- McGregor, R.A.; Poppitt, S.D. Milk Protein for Improved Metabolic Health: A Review of the Evidence. Nutr. Metab. 2013, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- Ricci-Cabello, I.; Olalla Herrera, M.; Artacho, R. Possible Role of Milk-Derived Bioactive Peptides in the Treatment and Prevention of Metabolic Syndrome. Nutr. Rev. 2012, 70, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Paddon-Jones, D.; Westman, E.; Mattes, R.D.; Wolfe, R.R.; Astrup, A.; Westerterp-Plantenga, M. Protein, Weight Management, and Satiety. Am. J. Clin. Nutr. 2008, 87, 1558S–1561S. [Google Scholar] [CrossRef] [PubMed]
- Achaglinkame, M.A.; Dari, L.; Mörlein, D. A Review of Dairy Production and Utilization in Ghana and Benin. Discov. Food 2023, 3, 13. [Google Scholar] [CrossRef]
- Garau, V.; Manis, C.; Scano, P.; Caboni, P. Compositional Characteristics of Mediterranean Buffalo Milk and Whey. Dairy 2021, 2, 469–488. [Google Scholar] [CrossRef]
- Mathai, J.K.; Liu, Y.; Stein, H.H. Values for Digestible Indispensable Amino Acid Scores (DIAAS) for Some Dairy and Plant Proteins May Better Describe Protein Quality than Values Calculated Using the Concept for Protein Digestibility-Corrected Amino Acid Scores (PDCAAS). Br. J. Nutr. 2017, 117, 490–499. [Google Scholar] [CrossRef]
- Roy, D.; Ye, A.; Moughan, P.J.; Singh, H. Composition, Structure, and Digestive Dynamics of Milk From Different Species—A Review. Front. Nutr. 2020, 7. [Google Scholar] [CrossRef]
- Jenness, R. Composition of Milk BT—Fundamentals of Dairy Chemistry; Wong, N.P., Jenness, R., Keeney, M., Marth, E.H., Eds.; Springer US: Boston, MA, USA, 1988; pp. 1–38. ISBN 978-1-4615-7050-9. [Google Scholar]
- Hossein, S.; Shahbazi, R.; Esmaeili, S.; Sohrabvandi, S. Health-Related Aspects of Milk Proteins. Iran. J. Pharm. Res. 2016, 15, 573–591. [Google Scholar]
- Sindayikengera, S.; Xia, W. Nutritional Evaluation of Caseins and Whey Proteins and Their Hydrolysates from Protamex. J. Zhejiang Univ. Sci. B 2006, 7, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.-P.; Maubois, J.-L.; Beaufrère, B. Slow and Fast Dietary Proteins Differently Modulate Postprandial Protein Accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.L. The Rationale for Consuming Protein Blends in Sports Nutrition. J. Am. Coll. Nutr. 2009, 28 (Suppl. S4), 464S–472S. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.R.; Falvo, M.J. Protein—Which Is Best? J. Sports Sci. Med. 2004, 3, 118–130. [Google Scholar] [PubMed]
- Bendtsen, L.Q.; Lorenzen, J.K.; Bendsen, N.T.; Rasmussen, C.; Astrup, A. Effect of Dairy Proteins on Appetite, Energy Expenditure, Body Weight, and Composition: A Review of the Evidence from Controlled Clinical Trials. Adv. Nutr. 2013, 4, 418–438. [Google Scholar] [CrossRef]
- Hidayat, K.; Du, X.; Shi, B.-M. Milk in the Prevention and Management of Type 2 Diabetes: The Potential Role of Milk Proteins. Diabetes. Metab. Res. Rev. 2019, 35, e3187. [Google Scholar] [CrossRef] [PubMed]
- Sackesen, C.; Altintas, D.U.; Bingol, A.; Bingol, G.; Buyuktiryaki, B.; Demir, E.; Kansu, A.; Kuloglu, Z.; Tamay, Z.; Sekerel, B.E. Current Trends in Tolerance Induction in Cow’s Milk Allergy: From Passive to Proactive Strategies. Front. Pediatr. 2019, 7, 372. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, B.; Indirli, G.C.; Bianchi, A.; Arasi, S.; Caimmi, D.; Dondi, A.; La Grutta, S.; Panetta, V.; Verga, M.C.; Calvani, M. Specific IgE and Skin Prick Tests to Diagnose Allergy to Fresh and Baked Cow’s Milk According to Age: A Systematic Review. Ital. J. Pediatr. 2017, 43, 93. [Google Scholar] [CrossRef] [PubMed]
- Wal, J.-M. Cow’s Milk Proteins/Allergens. Ann. Allergy Asthma Immunol. 2002, 89, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Shandilya, U.K.; Panwar, V.; Haq, M.R.; Kapila, R.; Kansal, V.K. IgE and IgG Cross Reactivity between Cow and Buffalo Milk Proteins in Swiss Albino Mice Model. Milchwissenschaft 2012, 67, 235–238. [Google Scholar]
- Villa, C.; Costa, J.; Oliveira, M.B.P.P.; Mafra, I. Bovine Milk Allergens: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 137–164. [Google Scholar] [CrossRef] [PubMed]
- Restani, P.; Ballabio, C.; Di Lorenzo, C.; Tripodi, S.; Fiocchi, A. Molecular Aspects of Milk Allergens and Their Role in Clinical Events. Anal. Bioanal. Chem. 2009, 395, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Klaassen Kamdar, A.; Labuza, T.P. Storage Stability of Food Protein Hydrolysates—A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 1169–1192. [Google Scholar] [CrossRef] [PubMed]
- Ramos, O.L.; Pereira, R.N.; Martins, A.; Rodrigues, R.; Fuciños, C.; Teixeira, J.A.; Pastrana, L.; Malcata, F.X.; Vicente, A.A. Design of Whey Protein Nanostructures for Incorporation and Release of Nutraceutical Compounds in Food. Crit. Rev. Food Sci. Nutr. 2017, 57, 1377–1393. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, C.; Sforza, S.; Palazzo, P.; Lambertini, F.; Paolella, S.; Zennaro, D.; Rafaiani, C.; Ferrara, R.; Bernardi, M.L.; Santoro, M.; et al. Tolerability of a Fully Maturated Cheese in Cow’s Milk Allergic Children: Biochemical, Immunochemical, and Clinical Aspects. PLoS ONE 2012, 7, e40945. [Google Scholar] [CrossRef] [PubMed]
- Manson, W. Nutritional Aspects of Milk Protein. Int. J. Dairy Technol. 1975, 28, 203–209. [Google Scholar] [CrossRef]
- Park, Y.W.; Nam, M.S. Bioactive Peptides in Milk and Dairy Products: A Review. Korean J. Food Sci. Anim. Resour. 2015, 35, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Aluko, R.E.; Hati, S.; Solanki, D. Bioactive Peptides in the Management of Lifestyle-Related Diseases: Current Trends and Future Perspectives. Crit. Rev. Food Sci. Nutr. 2022, 62, 4593–4606. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. The Scientific Evidence for the Role of Milk Protein-Derived Bioactive Peptides in Humans: A Review. J. Funct. Foods 2015, 17, 640–656. [Google Scholar] [CrossRef]
- Koirala, P.; Dahal, M.; Rai, S.; Dhakal, M.; Nirmal, N.P.; Maqsood, S.; Al-Asmari, F.; Buranasompob, A. Dairy Milk Protein–Derived Bioactive Peptides: Avengers Against Metabolic Syndrome. Curr. Nutr. Rep. 2023, 12, 299–317. [Google Scholar] [CrossRef] [PubMed]
- Meisel, H.; Bockelmann, W. Bioactive Peptides Encrypted in Milk Proteins: Proteolytic Activation and Thropho-Functional Properties. Antonie Van Leeuwenhoek 1999, 76, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Zaky, A.A.; Simal-Gandara, J.; Eun, J.B.; Shim, J.H.; Abd El-Aty, A.M. Bioactivities, Applications, Safety, and Health Benefits of Bioactive Peptides From Food and By-Products: A Review. Front. Nutr. 2022, 8, 1–18. [Google Scholar] [CrossRef]
- Abd El-Salam, M.H.; El-Shibiny, S. Preparation, Properties, and Uses of Enzymatic Milk Protein Hydrolysates. Crit. Rev. Food Sci. Nutr. 2017, 57, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Salam, M.H.; El-Shibiny, S. Reduction of Milk Protein Antigenicity by Enzymatic Hydrolysis and Fermentation. A Review. Food Rev. Int. 2021, 37, 276–295. [Google Scholar] [CrossRef]
- Meisel, H. Biochemical Properties of Regulatory Peptides Derived from Milk Proteins. Biopolym. Pept. Sci. Sect. 1997, 43, 119–128. [Google Scholar] [CrossRef]
- Mellander, O. The Physiological Importance of the Casein Phosphopeptide Calcium Salts. II. Peroral Calcium Dosage of Infants. Acta Soc. Med. Ups. 1950, 55, 247–255. [Google Scholar] [PubMed]
- Xu, Q.; Yan, X.; Zhang, Y.; Wu, J. Current Understanding of Transport and Bioavailability of Bioactive Peptides Derived from Dairy Proteins: A Review. Int. J. Food Sci. Technol. 2019, 54, 1930–1941. [Google Scholar] [CrossRef]
- Nath, A.; Eren, B.A.; Zinia Zaukuu, J.-L.; Koris, A.; Pásztorné-Huszár, K.; Szerdahelyi, E.; Kovacs, Z. Detecting the Bitterness of Milk-Protein-Derived Peptides Using an Electronic Tongue. Chemosens. 2022, 10, 215. [Google Scholar] [CrossRef]
- Maruyama, S.; Nakagomi, K.; Tomizuka, N.; Suzuki, H. Angiotensin I-Converting Enzyme Inhibitor Derived from an Enzymatic Hydrolysate of Casein. Ii. Isolation and Bradykinin-Potentiating Activity on the Uterus and the Ileum of Rats. Agric. Biol. Chem. 1985, 49, 1405–1409. [Google Scholar] [CrossRef]
- Maeno, M.; Yamamoto, N.; Takano, T. Identification of an Antihypertensive Peptide from Casein Hydrolysate Produced by a Proteinase from Lactobacillus Helveticus CP790. J. Dairy Sci. 1996, 79, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Mullally, M.M.; Meisel, H.; Fitzgerald, R.J. Angiotensin-I-Converting Enzyme Inhibitory Activities of Gastric and Pancreatic Proteinase Digests of Whey Proteins. Int. Dairy J. 1997, 7, 299–303. [Google Scholar] [CrossRef]
- Yan, D.; Chen, D.; Shen, J.; Xiao, G.; van Wijnen, A.J.; Im, H.J. Bovine Lactoferricin Is Anti-Inflammatory and Anti-Catabolic in Human Articular Cartilage and Synovium. J. Cell. Physiol. 2013, 228, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, R.K.; Kanwar, J.R. Immunomodulatory Lactoferrin in the Regulation of Apoptosis Modulatory Proteins in Cancer. Protein Pept. Lett. 2013, 20, 450–458. [Google Scholar] [PubMed]
- Furlong, S.J.; Mader, J.S.; Hoskin, D.W. Bovine Lactoferricin Induces Caspase-Independent Apoptosis in Human B-Lymphoma Cells and Extends the Survival of Immune-de Fi Cient Mice Bearing B-Lymphoma Xenografts. Exp. Mol. Pathol. 2010, 88, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Meisel, H. Biochemical Properties of Peptides Encrypted in Bovine Milk Proteins. Curr. Med. Chem. 2005, 12, 1905–1919. [Google Scholar] [CrossRef] [PubMed]
- Yvon, M.; Beucher, S.; Guilloteau, P.; Le Huerou-Luron, I.; Corring, T. Effects of Caseinomacropeptide (CMP) on Digestion Regulation. Reprod. Nutr. Dev. 1994, 34, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Córdova-Dávalos, L.E.; Jiménez, M.; Salinas, E. Glycomacropeptide Bioactivity and Health: A Review Highlighting Action Mechanisms and Signaling Pathways. Nutrients 2019, 11, 598. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, R.J.; Meisel, H. Lactokinins: Whey Protein-Derived ACE Inhibitory Peptides. Nahr. Food 1999, 43, 165–167. [Google Scholar] [CrossRef]
- Pepe, G.; Tenore, G.C.; Mastrocinque, R.; Stusio, P.; Campiglia, P. Potential Anticarcinogenic Peptides from Bovine Milk. J. Amino Acids 2013, 2013, 939804. [Google Scholar] [CrossRef] [PubMed]
- Shazly, A.B.; He, Z.; El-Aziz, M.A.; Zeng, M.; Zhang, S.; Qin, F.; Chen, J. Fractionation and Identification of Novel Antioxidant Peptides from Buffalo and Bovine Casein Hydrolysates. Food Chem. 2017, 232, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Wang, X.; Hu, Z.; Wu, Z.; Wang, L.; Wang, H.; Yang, M. Identification and Characterization of an Angiotensin-Converting Enzyme Inhibitory Peptide Derived from Bovine Casein. Peptides 2018, 99, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Pan, D.; Zhang, T.; Liu, C.; Zhang, J.; Su, M.; Wu, Z.; Zeng, X.; Sun, Y.; Guo, Y. Novel Milk Casein-Derived Peptides Decrease Cholesterol Micellar Solubility and Cholesterol Intestinal Absorption in Caco-2 Cells. J. Dairy Sci. 2020, 103, 3924–3936. [Google Scholar] [CrossRef] [PubMed]
- Mudgil, P.; Kamal, H.; Priya Kilari, B.; Mohd Salim, M.A.S.; Gan, C.-Y.; Maqsood, S. Simulated Gastrointestinal Digestion of Camel and Bovine Casein Hydrolysates: Identification and Characterization of Novel Anti-Diabetic Bioactive Peptides. Food Chem. 2021, 353, 129374. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M. Biofunctional Peptides from Milk Proteins: Mineral Binding and Cytomodulatory Effects. Curr. Pharm. Des. 2003, 9, 1289–1295. [Google Scholar] [CrossRef]
- Minervini, F.; Algaron, F.; Rizzello, C.G.; Fox, P.F.; Monnet, V.; Gobbetti, M. Angiotensin I-Converting-Enzyme-Inhibitory and Antibacterial Peptides from Lactobacillus Helveticus PR4 Proteinase-Hydrolyzed Caseins of Milk from Six Species. Appl. Environ. Microbiol. 2003, 69, 5297–5305. [Google Scholar] [CrossRef] [PubMed]
- Hern, B.; Miralles, B.; Amigo, L.; Ramos, M. Identification of Antioxidant and ACE-Inhibitory Peptides in Fermented Milk. J. Sci. Food Agric. 2005, 1048, 1041–1048. [Google Scholar] [CrossRef]
- Meisel, H. Overview on Milk Protein-Derived Peptides. Int. Dairy J. 1998, 8, 363–373. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.; Otte, J.; De Gobba, C.; Osman, A.; Hamad, E. Angiotensin I-Converting Enzyme Inhibitory Activity and Antioxidant Capacity of Bioactive Peptides Derived from Enzymatic Hydrolysis of Buffalo Milk Proteins. Int. Dairy J. 2017, 66, 91–98. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Jahandideh, F.; Davidge, S.T.; Wu, J. Milk-Derived Tripeptides IPP (Ile-Pro-Pro) and VPP (Val-Pro-Pro) Enhance Insulin Sensitivity and Prevent Insulin Resistance in 3T3-F442A Preadipocytes. J. Agric. Food Chem. 2018, 66, 10179–10187. [Google Scholar] [CrossRef]
- Zhao, Q.; Wei, G.; Li, K.; Duan, S.; Ye, R.; Huang, A. Identification and Molecular Docking of Novel α-Glucosidase Inhibitory Peptides from Hydrolysates of Binglangjiang Buffalo Casein. LWT 2022, 156, 113062. [Google Scholar] [CrossRef]
- Basilicata, M.G.; Pepe, G.; Adesso, S.; Ostacolo, C.; Sala, M.; Sommella, E.; Scala, M.C.; Messore, A.; Autore, G.; Marzocco, S.; et al. Antioxidant Properties of Buffalo-Milk Dairy Products: A β-Lg Peptide Released after Gastrointestinal Digestion of Buffalo Ricotta Cheese Reduces Oxidative Stress in Intestinal Epithelial Cells. Int. J. Mol. Sci. 2018, 19, 1955. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, V.P.; Kapila, S.; Kemgang, T.S.; Reddi, S.; Kapila, R.; Muthukumar, S.; Rajesh, D. Isolation and Characterization of Angiotensin Converting Enzyme Inhibitory Peptide from Buffalo Casein. Int. J. Pept. Res. Ther. 2021, 27, 1481–1491. [Google Scholar] [CrossRef]
- Shanmugam, V.P.; Kapila, S.; Sonfack, T.K.; Kapila, R. Antioxidative Peptide Derived from Enzymatic Digestion of Buffalo Casein. Int. Dairy J. 2015, 42, 1–5. [Google Scholar] [CrossRef]
- Kimmerlin, T.; Seebach, D.; Hilvert, D. Synthesis of Β3-Peptides and Mixed α/Β3-Peptides by Thioligation. Helv. Chim. Acta 2002, 85, 1812–1826. [Google Scholar] [CrossRef]
- Guzmán, F.; Barberis, S.; Illanes, A. Peptide Synthesis: Chemical or Enzymatic. Electron. J. Biotechnol. 2007, 10, 279–314. [Google Scholar] [CrossRef]
- Alpine Roads. Nobell Foods Food Compositions Comprising Milk Proteins Produced in Transgenic Plants. U.S. Patent Application No. 16/862,011, 2021. [Google Scholar]
- Dallas, D.; Nielsen, S.D. Milk Peptidomics to Identify Functional Peptides and for Quality Control of Dairy Products. Methods Mol. Biol. 2018, 1719, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.D.; Beverly, R.L.; Qu, Y.; Dallas, D.C. Milk Bioactive Peptide Database: A Comprehensive Database of Milk Protein-Derived Bioactive Peptides and Novel Visualization. Food Chem. 2017, 232, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Kussmann, M.; Affolter, M. Proteomics at the Center of Nutrigenomics: Comprehensive Molecular Understanding of Dietary Health Effects. Nutrition 2009, 25, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Comerford, K.B.; Pasin, G. Gene-Dairy Food Interactions and Health Outcomes: A Review of Nutrigenetic Studies. Nutrients 2017, 9, 710. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; Cermeño, M.; Khalesi, M.; Kleekayai, T.; Amigo-Benavent, M. Application of in Silico Approaches for the Generation of Milk Protein-Derived Bioactive Peptides. J. Funct. Foods 2020, 64, 103636. [Google Scholar] [CrossRef]
- Iwaniak, A.; Mogut, D. Metabolic Syndrome-Preventive Peptides Derived from Milk Proteins and Their Presence in Cheeses: A Review. Appl. Sci. 2020, 10, 2772. [Google Scholar] [CrossRef]
- Chen, G.-C.; Szeto, I.M.Y.; Chen, L.-H.; Han, S.-F.; Li, Y.-J.; van Hekezen, R.; Qin, L.-Q. Dairy Products Consumption and Metabolic Syndrome in Adults: Systematic Review and Meta-Analysis of Observational Studies. Sci. Rep. 2015, 5, 14606. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Gutiérrez-López, G.F.; Dávila-Ortiz, G. Use of Proteomics and Peptidomics Methods in Food Bioactive Peptide Science and Engineering. Food Eng. Rev. 2012, 4, 224–243. [Google Scholar] [CrossRef]
- Nilsson, M.; Holst, J.J.; Björck, I.M.E. Metabolic Effects of Amino Acid Mixtures and Whey Protein in Healthy Subjects: Studies Using Glucose-Equivalent Drinks. Am. J. Clin. Nutr. 2007, 85, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Dangin, M.; Boirie, Y.; Garcia-Rodenas, C.; Gachon, P.; Fauquant, J.; Callier, P.; Ballèvre, O.; Beaufrère, B. The Digestion Rate of Protein Is an Independent Regulating Factor of Postprandial Protein Retention. Am. J. Physiol. Metab. 2001, 280, E340–E348. [Google Scholar] [CrossRef] [PubMed]
- Calbet, J.A.L.; Holst, J.J. Gastric Emptying, Gastric Secretion and Enterogastrone Response after Administration of Milk Proteins or Their Peptide Hydrolysates in Humans. Eur. J. Nutr. 2004, 43, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R.; Crombach, N.; Gijsen, A.P.; Walrand, S.; Fauquant, J.; Kies, A.K.; Lemosquet, S.; Saris, W.H.M.; Boirie, Y.; van Loon, L.J.C. Ingestion of a Protein Hydrolysate Is Accompanied by an Accelerated in Vivo Digestion and Absorption Rate When Compared with Its Intact Protein. Am. J. Clin. Nutr. 2009, 90, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Morifuji, M.; Ishizaka, M.; Baba, S.; Fukuda, K.; Matsumoto, H.; Koga, J.; Kanegae, M.; Higuchi, M. Comparison of Different Sources and Degrees of Hydrolysis of Dietary Protein: Effect on Plasma Amino Acids, Dipeptides, and Insulin Responses in Human Subjects. J. Agric. Food Chem. 2010, 58, 8788–8797. [Google Scholar] [CrossRef] [PubMed]
- Veldhorst, M.A.B.; Nieuwenhuizen, A.G.; Hochstenbach-Waelen, A.; van Vught, A.J.A.H.; Westerterp, K.R.; Engelen, M.P.K.J.; Brummer, R.-J.M.; Deutz, N.E.P.; Westerterp-Plantenga, M.S. Dose-Dependent Satiating Effect of Whey Relative to Casein or Soy. Physiol. Behav. 2009, 96, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Pistrosch, F.; Natali, A.; Hanefeld, M. Is Hyperglycemia a Cardiovascular Risk Factor? Diabetes Care 2011, 34, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Claessens, M.; Saris, W.H.M.; van Baak, M.A. Glucagon and Insulin Responses after Ingestion of Different Amounts of Intact and Hydrolysed Proteins. Br. J. Nutr. 2008, 100, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Gerich, J.E. Clinical Significance, Pathogenesis, and Management of Postprandial Hyperglycemia. Arch. Intern. Med. 2003, 163, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Ellis, V.; Dhaliwal, S. Effects of Whey Protein Isolate on Body Composition, Lipids, Insulin and Glucose in Overweight and Obese Individuals. Br. J. Nutr. 2010, 104, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food Protein-Derived Bioactive Peptides in Management of Type 2 Diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Features of Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides from Dietary Proteins. J. Food Biochem. 2019, 43, e12451. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Hoffmann, T.; Wagner, L.; Wermann, M.; Heiser, U.; Kiefersauer, R.; Huber, R.; Bode, W.; Demuth, H.-U.; Brandstetter, H. The Crystal Structure of Dipeptidyl Peptidase IV (CD26) Reveals Its Functional Regulation and Enzymatic Mechanism. Proc. Natl. Acad. Sci. USA 2003, 100, 5063–5068. [Google Scholar] [CrossRef]
- Guasch, L.; Ojeda, M.J.; González-Abuín, N.; Sala, E.; Cereto-Massagué, A.; Mulero, M.; Valls, C.; Pinent, M.; Ardévol, A.; Garcia-Vallvé, S.; et al. Identification of Novel Human Dipeptidyl Peptidase-IV Inhibitors of Natural Origin (Part I): Virtual Screening and Activity Assays. PLoS ONE 2012, 7, e44971. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Inhibition of Dipeptidyl Peptidase IV (DPP-IV) by Tryptophan Containing Dipeptides. Food Funct. 2013, 4, 1843–1849. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Inhibition of Dipeptidyl Peptidase IV (DPP-IV) by Proline Containing Casein-Derived Peptides. J. Funct. Foods 2013, 5, 1909–1917. [Google Scholar] [CrossRef]
- Sowmya, K.; Mala, D.; Bhat, M.I.; Kumar, N.; Bajaj, R.K.; Kapila, S.; Kapila, R. Bio-Accessible Milk Casein Derived Tripeptide (LLY) Mediates Overlapping Anti- Inflammatory and Anti-Oxidative Effects under Cellular (Caco-2) and in Vivo Milieu. J. Nutr. Biochem. 2018, 62, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.; Kumar, N.; Kapila, S.; Mada, S.B.; Reddi, S.; Vij, R.; Kapila, R. Buffalo Casein Derived Peptide Can Alleviates H2O2 Induced Cellular Damage and Necrosis in Fibroblast Cells. Exp. Toxicol. Pathol. 2017, 69, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Mada, S.B.; Reddi, S.; Kumar, N.; Kumar, R.; Kapila, S.; Kapila, R.; Trivedi, R.; Karvande, A.; Ahmad, N. Antioxidative Peptide from Milk Exhibits Antiosteopenic Effects through Inhibition of Oxidative Damage and Bone-Resorbing Cytokines in Ovariectomized Rats. Nutrition 2017, 43–44, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Plant Protein-Derived Antioxidant Peptides: Isolation, Identification, Mechanism of Action and Application in Food Systems: A Review. Trends Food Sci. Technol. 2020, 105, 308–322. [Google Scholar] [CrossRef]
- Kumar, N.; Devi, S.; Mada, S.B.; Reddi, S.; Kapila, R.; Kapila, S. Anti-Apoptotic Effect of Buffalo Milk Casein Derived Bioactive Peptide by Directing Nrf2 Regulation in Starving Fibroblasts. Food Biosci. 2020, 35, 100566. [Google Scholar] [CrossRef]
- Iwasa, M.; Takezoe, S.; Kitaura, N.; Sutani, T.; Miyazaki, H.; Aoi, W. A Milk Casein Hydrolysate-Derived Peptide Enhances Glucose Uptake through the AMP-Activated Protein Kinase Signalling Pathway in Skeletal Muscle Cells. Exp. Physiol. 2021, 106, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Kondrashina, A.; Brodkorb, A.; Giblin, L. Dairy-Derived Peptides for Satiety. J. Funct. Foods 2020, 66, 103801. [Google Scholar] [CrossRef]
- Kaelberer, M.M.; Buchanan, K.L.; Klein, M.E.; Barth, B.B.; Montoya, M.M.; Shen, X.; Bohórquez, D. V A Gut-Brain Neural Circuit for Nutrient Sensory Transduction. Science 2018, 361, eaat5236. [Google Scholar] [CrossRef] [PubMed]
- Hall, W.L.; Millward, D.J.; Long, S.J.; Morgan, L.M. Casein and Whey Exert Different Effects on Plasma Amino Acid Profiles, Gastrointestinal Hormone Secretion and Appetite. Br. J. Nutr. 2003, 89, 239–248. [Google Scholar] [CrossRef]
- Demling, R.H.; DeSanti, L. Effect of a Hypocaloric Diet, Increased Protein Intake and Resistance Training on Lean Mass Gains and Fat Mass Loss in Overweight Police Officers. Ann. Nutr. Metab. 2000, 44, 21–29. [Google Scholar] [CrossRef]
- Acheson, K.J.; Blondel-Lubrano, A.; Oguey-Araymon, S.; Beaumont, M.; Emady-Azar, S.; Ammon-Zufferey, C.; Monnard, I.; Pinaud, S.; Nielsen-Moennoz, C.; Bovetto, L. Protein Choices Targeting Thermogenesis and Metabolism. Am. J. Clin. Nutr. 2011, 93, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Froetschel, M.A.; Azain, M.J.; Edwards, G.L.; Barb, C.R.; Amos, H.E. Opioid and Cholecystokinin Antagonists Alleviate Gastric Inhibition of Food Intake by Premeal Loads of Casein in Meal-Fed Rats. J. Nutr. 2001, 131, 3270–3276. [Google Scholar] [CrossRef] [PubMed]
- Pupovac, J.; Anderson, G.H. Dietary Peptides Induce Satiety via Cholecystokinin-A and Peripheral Opioid Receptors in Rats. J. Nutr. 2002, 132, 2775–2780. [Google Scholar] [CrossRef] [PubMed]
- Toedebusch, R.G.; Childs, T.E.; Hamilton, S.R.; Crowley, J.R.; Booth, F.W.; Roberts, M.D. Postprandial Leucine and Insulin Responses and Toxicological Effects of a Novel Whey Protein Hydrolysate-Based Supplement in Rats. J. Int. Soc. Sports Nutr. 2012, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, G.; Reimann, F.; Gribble, F.M. Intestinal Sensing of Nutrients. In Appetite Control. Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 309–335. [Google Scholar] [CrossRef]
- Santos-Hernández, M.; Miralles, B.; Amigo, L.; Recio, I. Intestinal Signaling of Proteins and Digestion-Derived Products Relevant to Satiety. J. Agric. Food Chem. 2018, 66, 10123–10131. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Rouvinen-Watt, K. The Role of Food Peptides in Lipid Metabolism during Dyslipidemia and Associated Health Conditions. Int. J. Mol. Sci. 2015, 16, 9303–9313. [Google Scholar] [CrossRef]
- Karpe, F. Postprandial Lipoprotein Metabolism and Atherosclerosis. J. Intern. Med. 1999, 246, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S. Physiological Regulation of Lipoprotein Lipase. Biochim. Biophys. Acta 2014, 1841, 919–933. [Google Scholar] [CrossRef]
- Mudgil, P.; Baba, W.N.; Kamal, H.; FitzGerald, R.J.; Hassan, H.M.; Ayoub, M.A.; Gan, C.-Y.; Maqsood, S. A Comparative Investigation into Novel Cholesterol Esterase and Pancreatic Lipase Inhibitory Peptides from Cow and Camel Casein Hydrolysates Generated upon Enzymatic Hydrolysis and In-Vitro Digestion. Food Chem. 2022, 367, 130661. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qi, X.; Guan, K.; Gu, Y.; Wang, R.; Li, Q.; Ma, Y. Peptides Released from Bovine α-Lactalbumin by Simulated Digestion Alleviated Free Fatty Acids-Induced Lipid Accumulation in HepG2 Cells. J. Funct. Foods 2021, 85, 104618. [Google Scholar] [CrossRef]
- Hlais, S.; Reslan, D.R.A.; Sarieddine, H.K.; Nasreddine, L.; Taan, G.; Azar, S.; Obeid, O.A. Effect of Lysine, Vitamin B(6), and Carnitine Supplementation on the Lipid Profile of Male Patients with Hypertriglyceridemia: A 12-Week, Open-Label, Randomized, Placebo-Controlled Trial. Clin. Ther. 2012, 34, 1674–1682. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, A.; Petzke, K.J.; Klaus, S. Dietary L-Leucine and L-Alanine Supplementation Have Similar Acute Effects in the Prevention of High-Fat Diet-Induced Obesity. Amino Acids 2013, 44, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Youn, B. Hypolipidemic Roles of Casein-Derived Peptides by Regulation of Trans-Intestinal Cholesterol Excretion and Bile Acid Synthesis. Nutrients 2020, 12, 3058. [Google Scholar] [CrossRef]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 8267234. [Google Scholar] [CrossRef] [PubMed]
- Tonolo, F.; Folda, A.; Cesaro, L.; Scalcon, V.; Marin, O.; Ferro, S.; Bindoli, A.; Rigobello, M.P. Milk-Derived Bioactive Peptides Exhibit Antioxidant Activity through the Keap1-Nrf2 Signaling Pathway. J. Funct. Foods 2020, 64, 103696. [Google Scholar] [CrossRef]
- Qi, X.; Chen, H.; Guan, K.; Sun, Y.; Wang, R.; Li, Q.; Ma, Y. Novel Xanthine Oxidase Inhibitory Peptides Derived from Whey Protein: Identification, in Vitro Inhibition Mechanism and in Vivo Activity Validation. Bioorg. Chem. 2022, 128, 106097. [Google Scholar] [CrossRef] [PubMed]
- Putnam, K.; Shoemaker, R.; Yiannikouris, F.; Cassis, L.A. The Renin-Angiotensin System: A Target of and Contributor to Dyslipidemias, Altered Glucose Homeostasis, and Hypertension of the Metabolic Syndrome. Am. J. Physiol. Circ. Physiol. 2012, 302, H1219–H1230. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative Stress, Insulin Resistance, Dyslipidemia and Type 2 Diabetes Mellitus. World J. Diabetes 2015, 6, 456. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, Oxidative Stress, and Obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.J.; Olza, J.; Gil, Á.; Aguilera, C.M. Chapter 1—Oxidative Stress and Inflammation in Obesity and Metabolic Syndrome. In Obesity; del Moral, A.M., Aguilera García, C.M.B.T.-O., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 1–15. ISBN 978-0-12-812504-5. [Google Scholar]
- Meyer, J.; Bütikofer, U.; Walther, B.; Wechsler, D.; Sieber, R. Hot Topic: Changes in Angiotensin-Converting Enzyme Inhibition and Concentrations of the Tripeptides Val-Pro-Pro and Ile-Pro-Pro during Ripening of Different Swiss Cheese Varieties. J. Dairy Sci. 2009, 92, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, C.; Xue, J.; Kwok, L.Y.; Yang, J.; Zhang, H.; Menghe, B. Characterization of Angiotensin-Converting Enzyme Inhibitory Activity of Fermented Milk Produced by Lactobacillus Helveticus. J. Dairy Sci. 2015, 98, 5113–5124. [Google Scholar] [CrossRef] [PubMed]
- Sipola, M.; Finckenberg, P.; Korpela, R.; Vapaatalo, H.; Nurminen, M.-L. Effect of Long-Term Intake of Milk Products on Blood Pressure in Hypertensive Rats. J. Dairy Res. 2002, 69, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Kawaguchi, K.; Yamamoto, N. Study of the Mechanism of Antihypertensive Peptides VPP and IPP in Spontaneously Hypertensive Rats by DNA Microarray Analysis. Eur. J. Pharmacol. 2009, 620, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kurihara, J.; Suzuki, S.; Nagashima, K.; Hosono, H.; Itagaki, F. The Influence of Dietary Peptide Inhibitors of Angiotensin-Converting Enzyme on the Hypotensive Effects of Enalapril. J. Pharm. Health Care Sci. 2015, 1, 17. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, S.M.; Kim, J.Y.; Choue, R.W. Effects of IPP and VPP Supplementation on Blood Pressure, ACE Activity and NO Production in Spontaneously Hypertensive Rats. FASEB J. 2008, 22, 1112.3. [Google Scholar] [CrossRef]
- Hirota, T.; Ohki, K.; Kawagishi, R.; Kajimoto, Y.; Mizuno, S.; Nakamura, Y.; Kitakaze, M. Casein Hydrolysate Containing the Antihypertensive Tripeptides Val-Pro-Pro and Ile-Pro-Pro Improves Vascular Endothelial Function Independent of Blood Pressure-Lowering Effects: Contribution of the Inhibitory Action of Angiotensin-Converting Enzyme. Hypertens. Res. 2007, 30, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Fekete, Á.A.; Givens, D.I.; Lovegrove, J.A. Casein-Derived Lactotripeptides Reduce Systolic and Diastolic Blood Pressure in a Meta-Analysis of Randomised Clinical Trials. Nutrients 2015, 7, 659–681. [Google Scholar] [CrossRef] [PubMed]
- de Leeuw, P.W.; van der Zander, K.; Kroon, A.A.; Rennenberg, R.M.W.; Koning, M.M.G. Dose-dependent Lowering of Blood Pressure by Dairy Peptides in Mildly Hypertensive Subjects. Blood Press. 2009, 18, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Aubin, F.; Azais-Braesco, V.; Borghi, C. Do the Lactotripeptides Isoleucine–Proline–Proline and Valine–Proline–Proline Reduce Systolic Blood Pressure in European Subjects? A Meta-Analysis of Randomized Controlled Trials. Am. J. Hypertens. 2013, 26, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Engberink, M.F.; Schouten, E.G.; Kok, F.J.; van Mierlo, L.A.J.; Brouwer, I.A.; Geleijnse, J.M. Lactotripeptides Show No Effect on Human Blood Pressure. Hypertension 2008, 51, 399–405. [Google Scholar] [CrossRef]
- van Mierlo, L.A.J.; Koning, M.M.G.; van der Zander, K.; Draijer, R. Lactotripeptides Do Not Lower Ambulatory Blood Pressure in Untreated Whites: Results from 2 Controlled Multicenter Crossover Studies2. Am. J. Clin. Nutr. 2009, 89, 617–623. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, R.J.; Murray, B.A.; Walsh, D.J. Hypotensive Peptides from Milk Proteins. J. Nutr. 2004, 134, 980S–988S. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Liao, W.; Davidge, S.T.; Wu, J. Milk-Derived Tripeptides IPP (Ile-Pro-Pro) and VPP (Val-Pro-Pro) Differentially Modulate Angiotensin II Effects on Vascular Smooth Muscle Cells. J. Funct. Foods 2017, 30, 151–158. [Google Scholar] [CrossRef]
- Nakamura, T.; Hirota, T.; Mizushima, K.; Ohki, K.; Naito, Y.; Yamamoto, N.; Yoshikawa, T. Milk-Derived Peptides, Val-Pro-Pro and Ile-Pro-Pro, Attenuate Atherosclerosis Development in Apolipoprotein e-Deficient Mice: A Preliminary Study. J. Med. Food 2013, 16, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Turpeinen, A.M.; Kumpu, M.; Rönnback, M.; Seppo, L.; Kautiainen, H.; Jauhiainen, T.; Vapaatalo, H.; Korpela, R. Antihypertensive and Cholesterol-Lowering Effects of a Spread Containing Bioactive Peptides IPP and VPP and Plant Sterols. J. Funct. Foods 2009, 1, 260–265. [Google Scholar] [CrossRef]
- Liu, L.; Yu, S.; Bu, T.; He, G.; Li, S.; Wu, J. Casein Hydrolysate Alleviates Adipose Chronic Inflammation in High Fat-Diet Induced Obese C57BL/6J Mice through MAPK Pathway. Nutrients 2023, 15, 1813. [Google Scholar] [CrossRef] [PubMed]
- Aihara, K.; Osaka, M.; Yoshida, M. Oral Administration of the Milk Casein-Derived Tripeptide Val-Pro-Pro Attenuates High-Fat Diet-Induced Adipose Tissue Inflammation in Mice. Br. J. Nutr. 2014, 112, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Wu, J. Milk-Derived Tripeptides IPP (Ile-Pro-Pro) and VPP (Val-Pro-Pro) Promote Adipocyte Differentiation and Inhibit Inflammation in 3T3-F442A Cells. PLoS ONE 2015, 10, e0117492. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zheng, L.; Zhao, M. Molecular Targets and Mechanisms of Casein-Derived Tripeptides Ile-Pro-Pro and Val-Pro-Pro on Hepatic Glucose Metabolism. J. Agric. Food Chem. 2023, 71, 18802–18814. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zheng, J.; Zhou, M.; Li, S.; He, G.; Wu, J. Peptide Analogues of VPP and IPP with Improved Glucose Uptake Activity in L6 Myotubes Can Be Released from Cereal Proteins. J. Agric. Food Chem. 2021, 69, 2875–2883. [Google Scholar] [CrossRef] [PubMed]
- van Meijl, L.E.C.; Vrolix, R.; Mensink, R.P. Dairy Product Consumption and the Metabolic Syndrome. Nutr. Res. Rev. 2008, 21, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.L.; Zhou, J.; Kim, J.E.; Campbell, W.W. Incorporating Milk Protein Isolate into an Energy-Restricted Western-Style Eating Pattern Augments Improvements in Blood Pressure and Triglycerides, but Not Body Composition Changes in Adults Classified as Overweight or Obese: A Randomized Controlled Tria. Nutrients 2020, 12, 851. [Google Scholar] [CrossRef] [PubMed]
- Frid, A.H.; Nilsson, M.; Holst, J.J.; Björck, I.M.E. Effect of Whey on Blood Glucose and Insulin Responses to Composite Breakfast and Lunch Meals in Type 2 Diabetic Subjects. Am. J. Clin. Nutr. 2005, 82, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Manders, R.J.F.; Wagenmakers, A.J.M.; Koopman, R.; Zorenc, A.H.G.; Menheere, P.P.C.A.; Schaper, N.C.; Saris, W.H.M.; van Loon, L.J.C. Co-Ingestion of a Protein Hydrolysate and Amino Acid Mixture with Carbohydrate Improves Plasma Glucose Disposal in Patients with Type 2 Diabetes. Am. J. Clin. Nutr. 2005, 82, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.; Noakes, M.; Trenerry, C.; Clifton, P.M. Energy Intake, Ghrelin, and Cholecystokinin after Different Carbohydrate and Protein Preloads in Overweight Men. J. Clin. Endocrinol. Metab. 2006, 91, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Manders, R.J.F.; Praet, S.F.E.; Meex, R.C.R.; Koopman, R.; de Roos, A.L.; Wagenmakers, A.J.M.; Saris, W.H.M.; van Loon, L.J.C. Protein Hydrolysate/Leucine Co-Ingestion Reduces the Prevalence of Hyperglycemia in Type 2 Diabetic Patients. Diabetes Care 2006, 29, 2721–2722. [Google Scholar] [CrossRef] [PubMed]
- Manders, R.J.; Koopman, R.; Sluijsmans, W.E.; van den Berg, R.; Verbeek, K.; Saris, W.H.; Wagenmakers, A.J.; van Loon, L.J. Co-Ingestion of a Protein Hydrolysate with or without Additional Leucine Effectively Reduces Postprandial Blood Glucose Excursions in Type 2 Diabetic Men. J. Nutr. 2006, 136, 1294–1299. [Google Scholar] [CrossRef] [PubMed]
- Tessari, P.; Kiwanuka, E.; Cristini, M.; Zaramella, M.; Enslen, M.; Zurlo, C.; Garcia-Rodenas, C. Slow versus Fast Proteins in the Stimulation of Beta-Cell Response and the Activation of the Entero-Insular Axis in Type 2 Diabetes. Diabetes. Metab. Res. Rev. 2007, 23, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Stevens, J.E.; Cukier, K.; Maddox, A.F.; Wishart, J.M.; Jones, K.L.; Clifton, P.M.; Horowitz, M.; Rayner, C.K. Effects of a Protein Preload on Gastric Emptying, Glycemia, and Gut Hormones After a Carbohydrate Meal in Diet-Controlled Type 2 Diabetes. Diabetes Care 2009, 32, 1600–1602. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, L.S.; Hartvigsen, M.L.; Brader, L.J.; Astrup, A.; Schrezenmeir, J.; Holst, J.J.; Thomsen, C.; Hermansen, K. Differential Effects of Protein Quality on Postprandial Lipemia in Response to a Fat-Rich Meal in Type 2 Diabetes: Comparison of Whey, Casein, Gluten, and Cod Protein. Am. J. Clin. Nutr. 2009, 90, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Claessens, M.; Van Baak, M.A.; Monsheimer, S.; Saris, W.H.M. The Effect of a Low-Fat, High-Protein or High-Carbohydrate Ad Libitum Diet on Weight Loss Maintenance and Metabolic Risk Factors. Int. J. Obes. 2009, 33, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Ellis, V.; Ho, S. Acute Effects of Whey Protein Isolate on Cardiovascular Risk Factors in Overweight, Post-Menopausal Women. Atherosclerosis 2010, 212, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Brader, L.; Holm, L.; Mortensen, L.; Thomsen, C.; Astrup, A.; Holst, J.J.; de Vrese, M.; Schrezenmeir, J.; Hermansen, K. Acute Effects of Casein on Postprandial Lipemia and Incretin Responses in Type 2 Diabetic Subjects. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Geerts, B.F.; Van Dongen, M.G.J.; Flameling, B.; Moerland, M.M.; Kam, M.L.D.; Cohen, A.F.; Romijn, J.A.; Gerhardt, C.C.; Kloek, J.; Burggraaf, J. Hydrolyzed Casein Decreases Postprandial Glucose Concentrations in T2DM Patients Irrespective of Leucine Content. J. Diet. Suppl. 2011, 8, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Jonker, J.T.; Wijngaarden, M.A.; Kloek, J.; Groeneveld, Y.; Gerhardt, C.; Brand, R.; Kies, A.K.; Romijn, J.A.; Smit, J.W.A. Effects of Low Doses of Casein Hydrolysate on Post-Challenge Glucose and Insulin Levels. Eur. J. Intern. Med. 2011, 22, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, L.S.; Holmer-Jensen, J.; Hartvigsen, M.L.; Jensen, V.K.; Astrup, A.; De Vrese, M.; Holst, J.J.; Thomsen, C.; Hermansen, K. Effects of Different Fractions of Whey Protein on Postprandial Lipid and Hormone Responses in Type 2 Diabetes. Eur. J. Clin. Nutr. 2012, 66, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Holmer-Jensen, J.; Mortensen, L.S.; Astrup, A.; de Vrese, M.; Holst, J.J.; Thomsen, C.; Hermansen, K. Acute Differential Effects of Dietary Protein Quality on Postprandial Lipemia in Obese Non-Diabetic Subjects. Nutr. Res. 2013, 33, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, M.; Madadlou, A. Influence of Whey Protein and Its Hydrolysate on Prehypertension and Postprandial Hyperglycaemia in Adult Men. Int. Dairy J. 2013, 33, 62–66. [Google Scholar] [CrossRef]
- Van Meijl, L.E.C.; Mensink, R.P. Effects of Milk and Milk Constituents on Postprandial Lipid and Glucose Metabolism in Overweight and Obese Men. Br. J. Nutr. 2013, 110, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Manders, R.J.F.; Hansen, D.; Zorenc, A.H.G.; Dendale, P.; Kloek, J.; Saris, W.H.M.; van Loon, L.J.C. Protein Co-Ingestion Strongly Increases Postprandial Insulin Secretion in Type 2 Diabetes Patients. J. Med. Food 2014, 17, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Jakubowicz, D.; Froy, O.; Ahrén, B.; Boaz, M.; Landau, Z.; Bar-Dayan, Y.; Ganz, T.; Barnea, M.; Wainstein, J. Incretin, Insulinotropic and Glucose-Lowering Effects of Whey Protein Pre-Load in Type 2 Diabetes: A Randomised Clinical Trial. Diabetologia 2014, 57, 1807–1811. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Jesudason, D.R.; Stevens, J.E.; Keogh, J.B.; Jones, K.L.; Clifton, P.M.; Horowitz, M.; Rayner, C.K. Sustained Effects of a Protein ‘Preload’ on Glycaemia and Gastric Emptying over 4 Weeks in Patients with Type 2 Diabetes: A Randomized Clinical Trial. Diabetes Res. Clin. Pract. 2015, 108, e31–e34. [Google Scholar] [CrossRef] [PubMed]
- Hoefle, A.S.; Bangert, A.M.; Stamfort, A.; Gedrich, K.; Rist, M.J.; Lee, Y.-M.; Skurk, T.; Daniel, H. Metabolic Responses of Healthy or Prediabetic Adults to Bovine Whey Protein and Sodium Caseinate Do Not Differ. J. Nutr. 2015, 145, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Bohl, M.; Bjørnshave, A.; Rasmussen, K.V.; Schioldan, A.G.; Amer, B.; Larsen, M.K.; Dalsgaard, T.K.; Holst, J.J.; Herrmann, A.; O’Neill, S.; et al. Dairy Proteins, Dairy Lipids, and Postprandial Lipemia in Persons with Abdominal Obesity (DairyHealth): A 12-Wk, Randomized, Parallel-Controlled, Double-Blinded, Diet Intervention Study. Am. J. Clin. Nutr. 2015, 101, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Jakubowicz, D.; Wainstein, J.; Landau, Z.; Ahren, B.; Barnea, M.; Bar-Dayan, Y.; Froy, O. High-Energy Breakfast Based on Whey Protein Reduces Body Weight, Postprandial Glycemia and HbA1C in Type 2 Diabetes. J. Nutr. Biochem. 2017, 49, 1–7. [Google Scholar] [CrossRef]
- Almario, R.U.; Buchaxn, W.M.; Rocke, D.M.; Karakas, S.E. Glucose-Lowering Effect of Whey Protein Depends upon Clinical Characteristics of Patients with Type 2 Diabetes. BMJ Open Diabetes Res. Care 2017, 5, e000420. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, K.A.; Lucero, A.; Stoner, L.; Faulkner, J.; Whitfield, P.; Krebs, J.; Rowlands, D.S. Nil Whey Protein Effect on Glycemic Control after Intense Mixed-Mode Training in Type 2 Diabetes. Med. Sci. Sports Exerc. 2018, 50, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Horner, K.; Drummond, E.; O’Sullivan, V.; S C Sri Harsha, P.; Brennan, L. Effects of a Casein Hydrolysate versus Intact Casein on Gastric Emptying and Amino Acid Responses. Eur. J. Nutr. 2019, 58, 955–964. [Google Scholar] [CrossRef] [PubMed]
- King, D.G.; Walker, M.; Campbell, M.D.; Breen, L.; Stevenson, E.J.; West, D.J. A Small Dose of Whey Protein Co-Ingested with Mixed-Macronutrient Breakfast and Lunch Meals Improves Postprandial Glycemia and Suppresses Appetite in Men with Type 2 Diabetes: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2018, 107, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Bjørnshave, A.; Holst, J.J.; Hermansen, K. Pre-Meal Effect of Whey Proteins on Metabolic Parameters in Subjects with and without Type 2 Diabetes: A Randomized, Crossover Trial. Nutrients 2018, 10, 122. [Google Scholar] [CrossRef]
- Bjørnshave, A.; Holst, J.J.; Hermansen, K. A Pre-Meal of Whey Proteins Induces Differential Effects on Glucose and Lipid Metabolism in Subjects with the Metabolic Syndrome: A Randomised Cross-over Trial. Eur. J. Nutr. 2019, 58, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Sartorius, T.; Weidner, A.; Dharsono, T.; Boulier, A.; Wilhelm, M.; Schön, C. Postprandial Effects of a Proprietary Milk Protein Hydrolysate Containing Bioactive Peptides in Prediabetic Subjects. Nutrients 2019, 11, 1700. [Google Scholar] [CrossRef]
- Basciani, S.; Camajani, E.; Contini, S.; Persichetti, A.; Risi, R.; Bertoldi, L.; Strigari, L.; Prossomariti, G.; Watanabe, M.; Mariani, S.; et al. Very-Low-Calorie Ketogenic Diets With Whey, Vegetable, or Animal Protein in Patients With Obesity: A Randomized Pilot Study. J. Clin. Endocrinol. Metab. 2020, 105, 2939–2949. [Google Scholar] [CrossRef] [PubMed]
- Memelink, R.G.; Pasman, W.J.; Bongers, A.; Tump, A.; van Ginkel, A.; Tromp, W.; Wopereis, S.; Verlaan, S.; de Vogel-van den Bosch, J.; Weijs, P.J.M. Effect of an Enriched Protein Drink on Muscle Mass and Glycemic Control during Combined Lifestyle Intervention in Older Adults with Obesity and Type 2 Diabetes: A Double-Blind RCT. Nutrients 2020, 13, 64. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; D’Angelo, A.; Maffioli, P. Change of Some Oxidative Stress Parameters after Supplementation with Whey Protein Isolate in Patients with Type 2 Diabetes. Nutrition 2020, 73, 110700. [Google Scholar] [CrossRef] [PubMed]
- Manders, R.J.F.; Praet, S.F.E.; Vikström, M.H.; Saris, W.H.M.; van Loon, L.J.C. Protein Hydrolysate Co-Ingestion Does Not Modulate 24 h Glycemic Control in Long-Standing Type 2 Diabetes Patients. Eur. J. Clin. Nutr. 2009, 63, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Holmer-Jensen, J.; Hartvigsen, M.L.; Mortensen, L.S.; Astrup, A.; de Vrese, M.; Holst, J.J.; Thomsen, C.; Hermansen, K. Acute Differential Effects of Milk-Derived Dietary Proteins on Postprandial Lipaemia in Obese Non-Diabetic Subjects. Eur. J. Clin. Nutr. 2012, 66, 32–38. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Weisse, K.; Brandsch, C.; Zernsdorf, B.; Nkengfack Nembongwe, G.S.; Hofmann, K.; Eder, K.; Stangl, G.I. Lupin Protein Compared to Casein Lowers the LDL Cholesterol: HDL Cholesterol-Ratio of Hypercholesterolemic Adults. Eur. J. Nutr. 2010, 49, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, L.Q.; Lorenzen, J.K.; Gomes, S.; Liaset, B.; Holst, J.J.; Ritz, C.; Reitelseder, S.; Sjödin, A.; Astrup, A. Effects of Hydrolysed Casein, Intact Casein and Intact Whey Protein on Energy Expenditure and Appetite Regulation: A Randomised, Controlled, Cross-over Study. Br. J. Nutr. 2014, 112, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Nouri, M.; Tarighat-Esfanjani, A.; Sadra, V.; Ghasempour, Z.; Jafarabadi, M.A.; Tajfar, P.; Gargari, B.P. Effects of Whey Protein Concentrate on Glycemic Status, Lipid Profile, and Blood Pressure in Overweight/Obese Women with Type 2 Diabetes Mellitus: A Randomized Placebo-Controlled Clinical Trial. Turk. J. Endocrinol. Metab. 2022, 26, 148–155. [Google Scholar] [CrossRef]








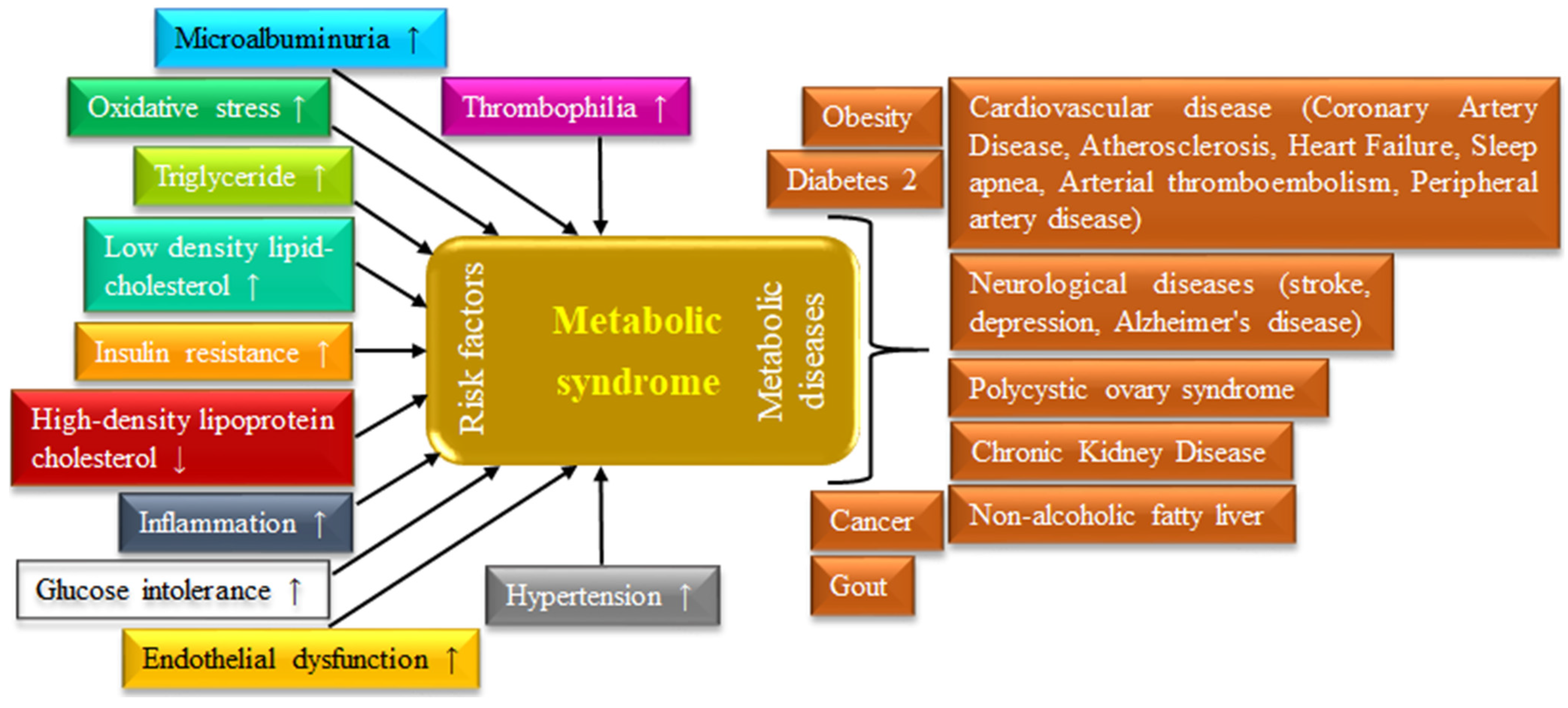{kind=link}
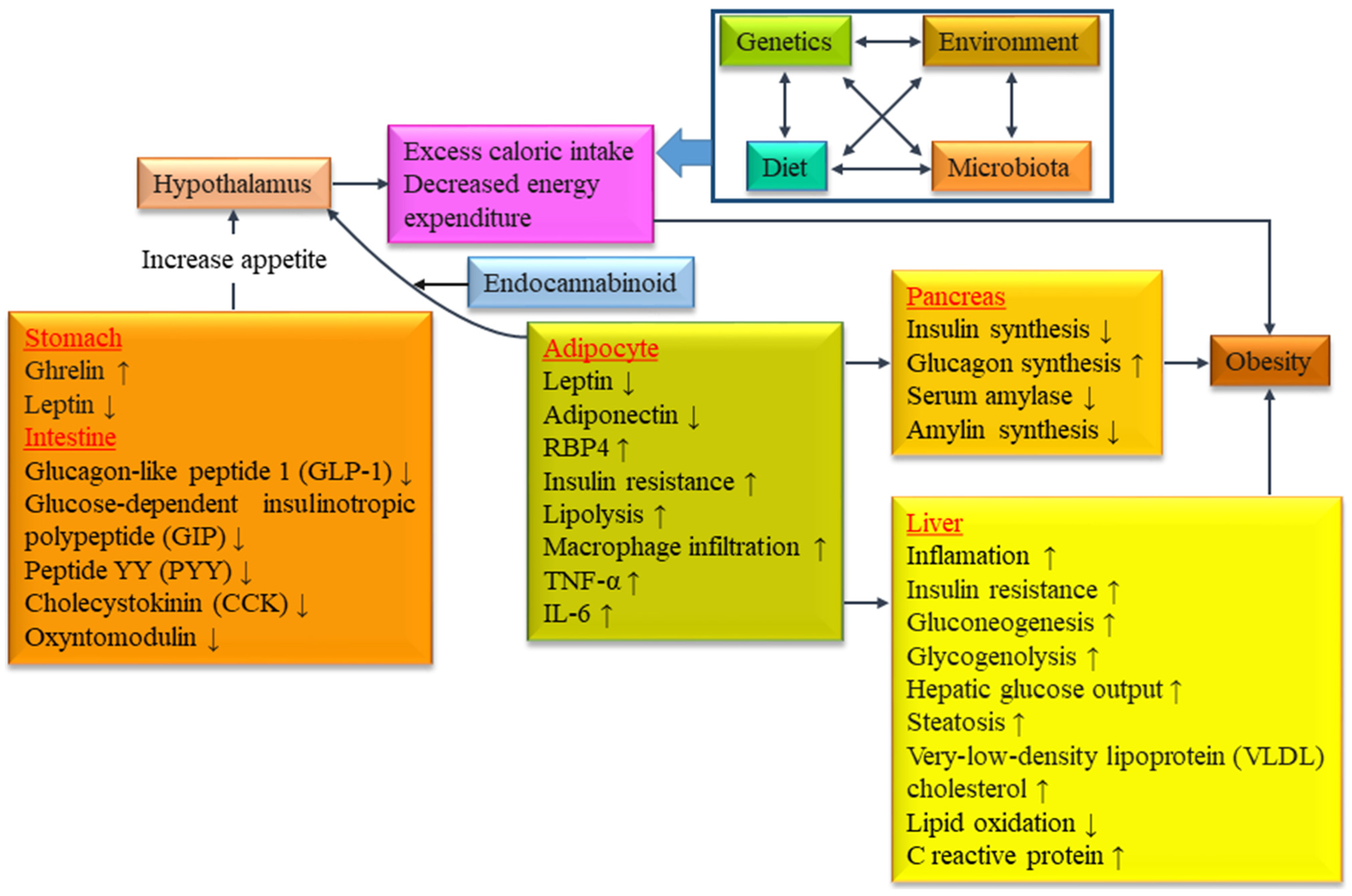{kind=link}
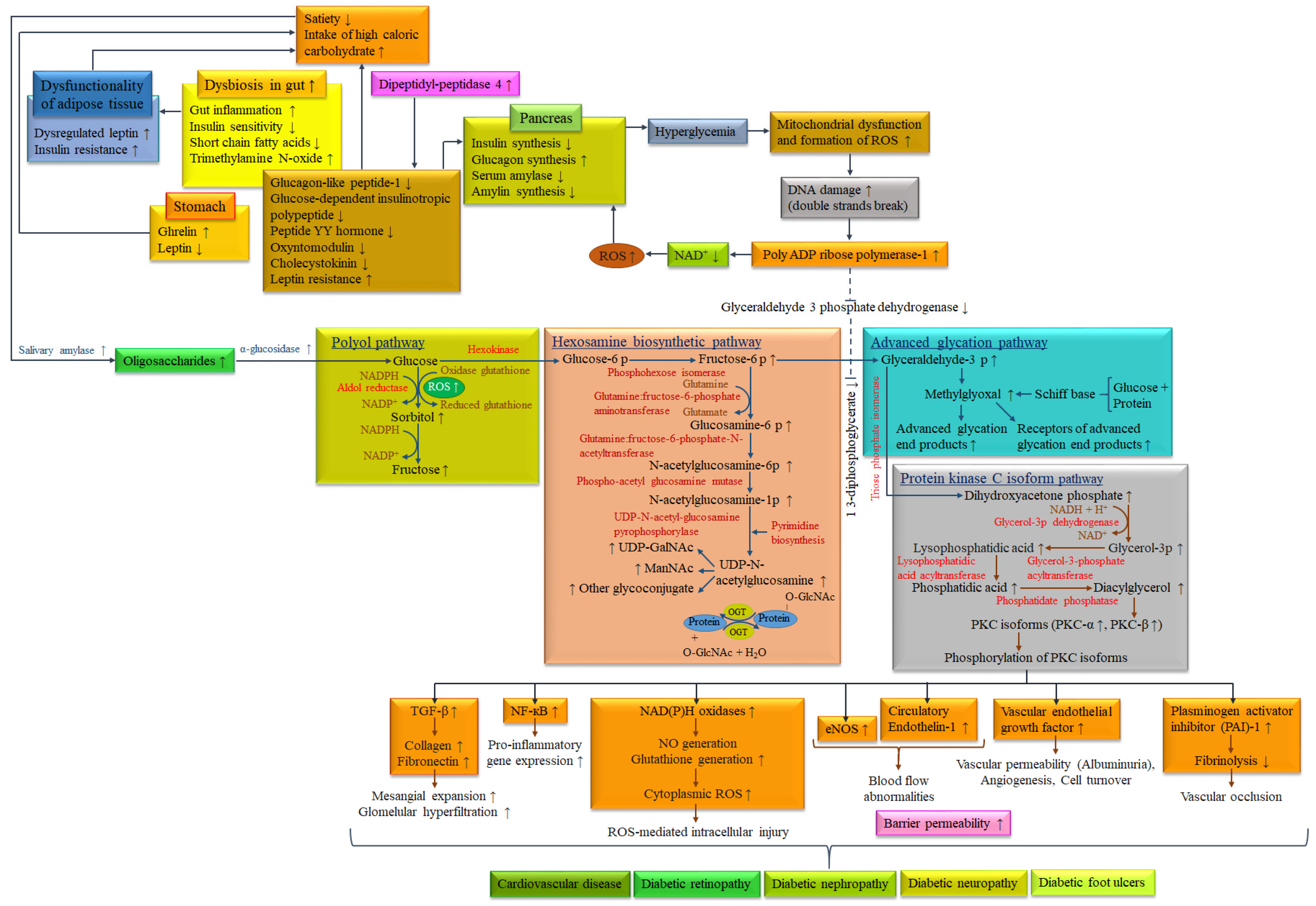{kind=link}
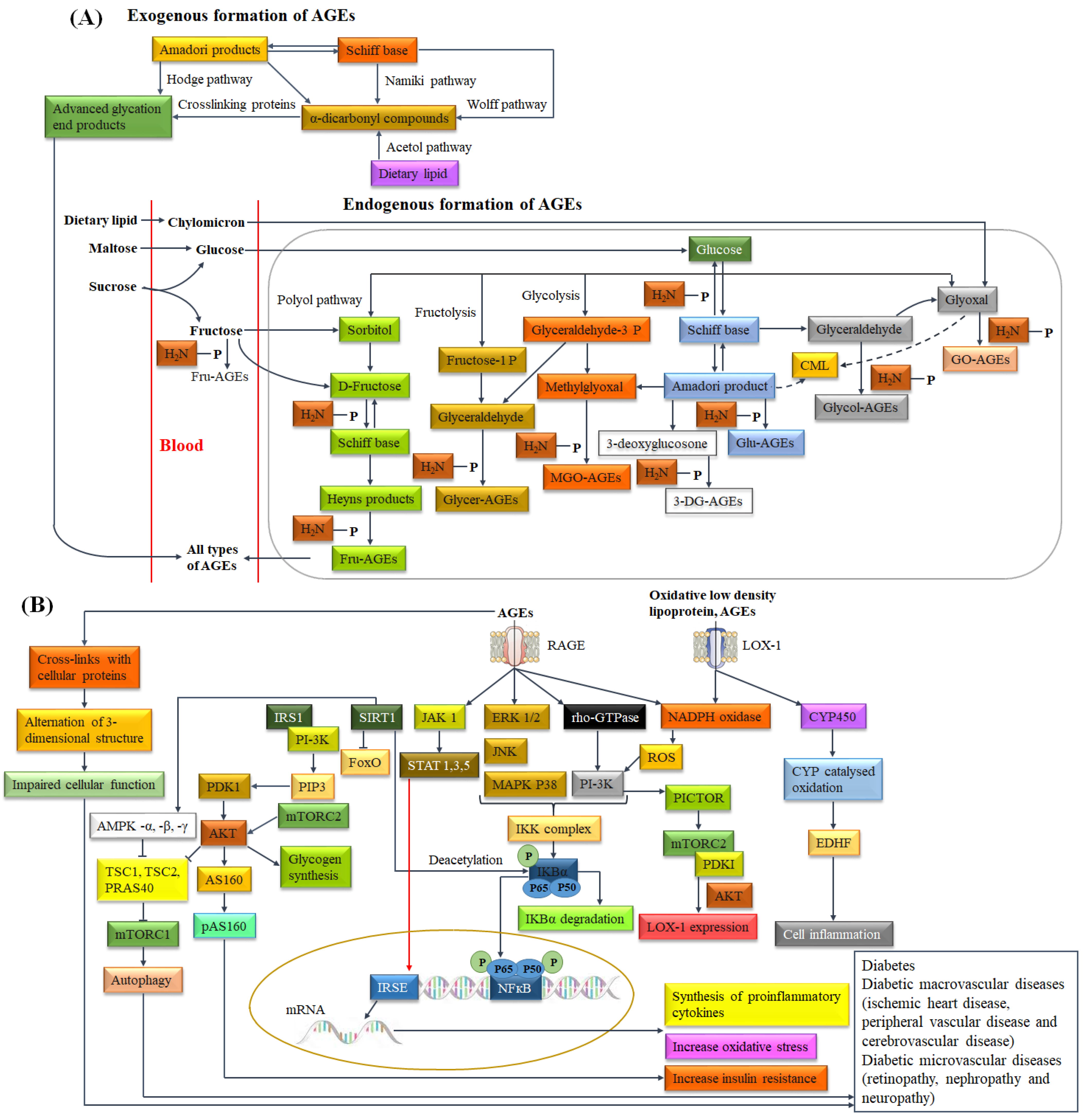{kind=link}
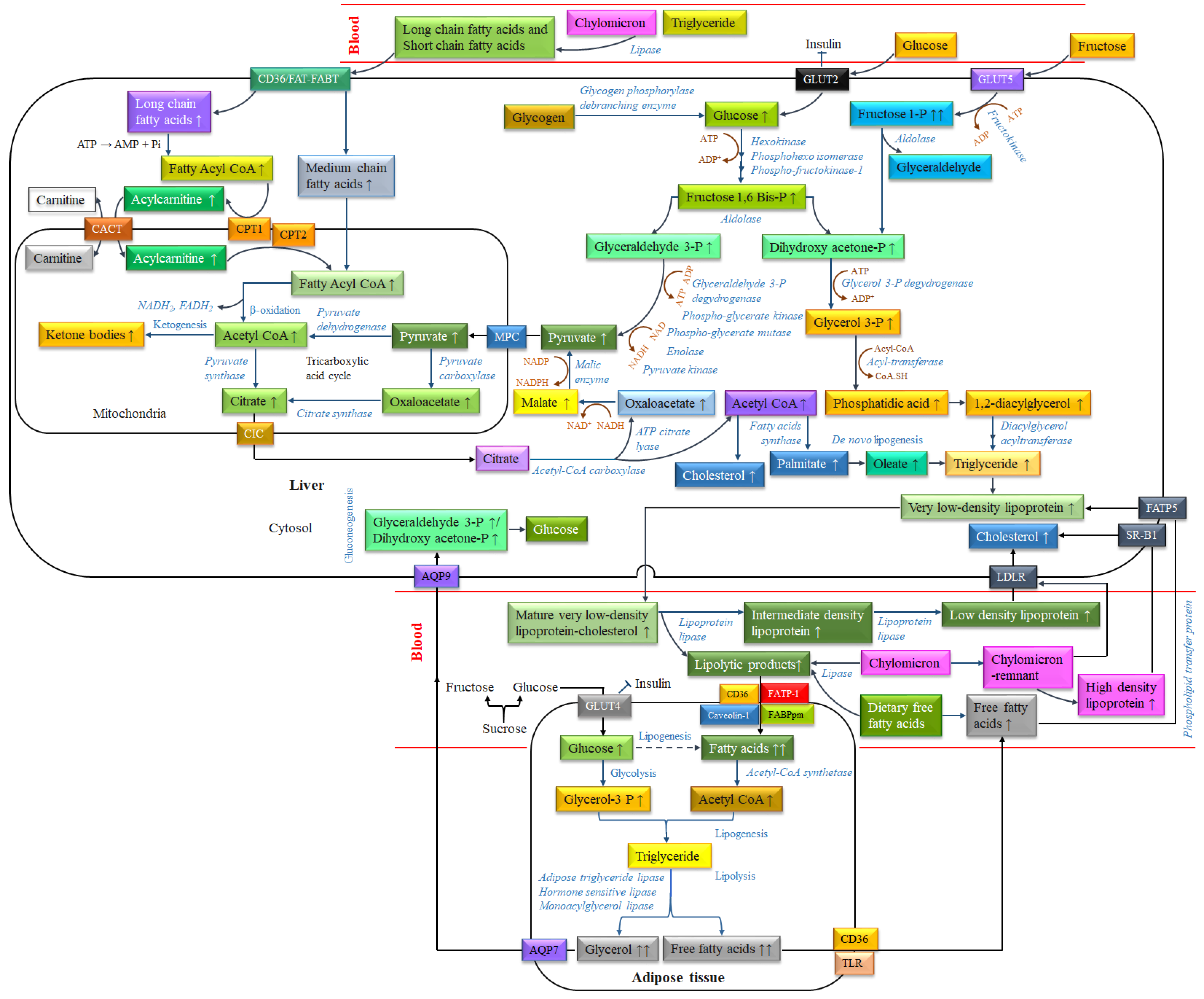{kind=link}
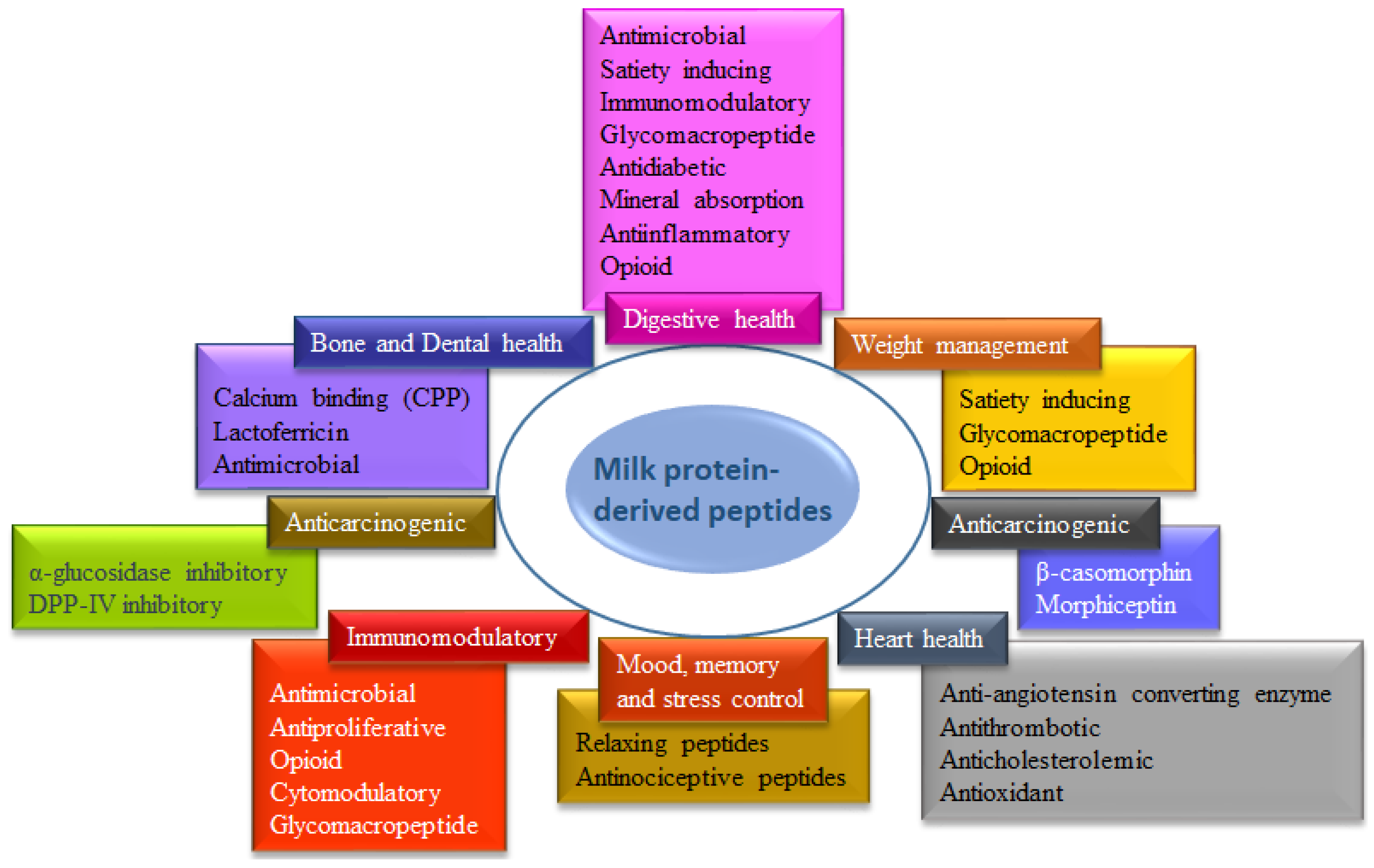{kind=link}
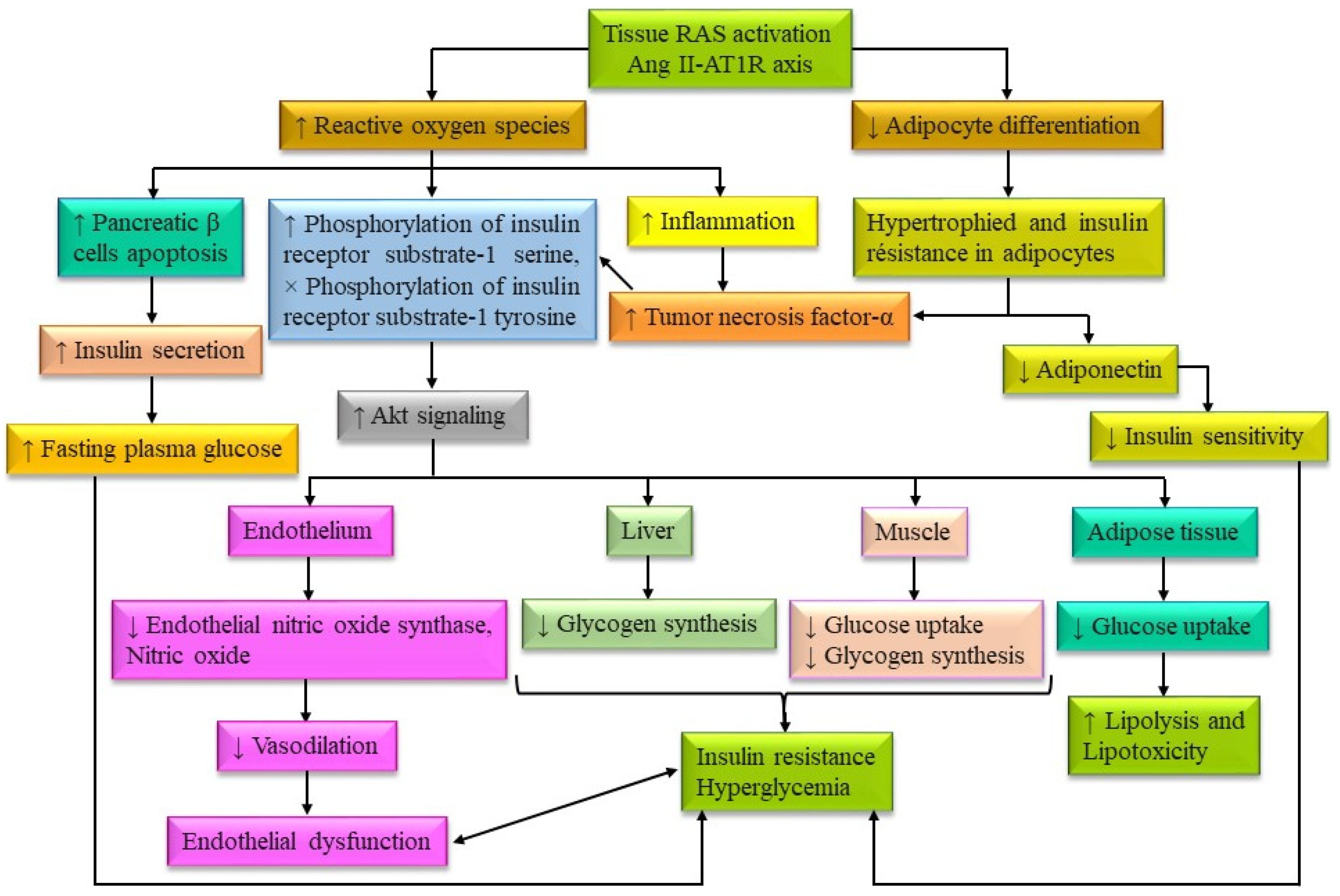{kind=link}
| WHO | IDF | EGIR | AACE | |
|---|---|---|---|---|
| Insulin resistance | Impaired glucose tolerance, Impaired fasting glucose, T2DM, or Lowered insulin sensitivity * plus any 2 of the following. | - | Plasma insulin >75th percentile plus any 2 of the following | Impaired fasting glucose or Impaired fasting glucose plus any of the following based on the clinical judgment |
| Blood glucose | Impaired glucose tolerance, Impaired fasting glucose, or T2DM | ≥100 mg·dL−1 (includes diabetes) ** | Impaired glucose tolerance or Impaired fasting glucose (but not diabetes) | Impaired fasting glucose or Impaired fasting glucose (but not diabetes) |
| Dyslipidemia | TG ≥ 150 mg·dL−1 and/or High density lipoprotein cholesterol < 35 mg·dL−1 in men or <39 mg·dL−1 in women. | TG ≥ 150 mg·dL−1 or on receiving treatment for TG. High density lipoprotein cholesterol < 40 mg·dL−1 in men or <50 mg·dL−1 in women or on receiving treatment for High density lipoprotein cholesterol. | TG ≥ 150 mg·dL−1 and/or High density lipoprotein cholesterol < 39 mg·dL−1 in men or women. | TG ≥ 150 mg·dL−1 and High density lipoprotein cholesterol < 40 mg·dL−1 in men or <50 mg·dL−1 in women |
| Blood pressure | ≥140/90 mm Hg | ≥130 mm Hg systolic or ≥85 mm Hg diastolic or on receiving treatment for hypertension. | ≥140/90 mm Hg or on receiving treatment for hypertension. | ≥130/85 mm Hg |
| Body weight | Men: waist-to-hip ratio >0.90; Women: waist-to-hip ratio >0.85 and/or BMI > 30 kg·m−2. | Increased waist circumference (population specific) plus any 2 of the following. | Waist circumference ≥94 cm in men or ≥80 cm in women. | BMI ≥ 25 kg·m−2 |
| Others | Microalbuminuria: Urinary excretion rate of >20 mg·min−1 or albumin: creatinine ratio of >30 mg·g−1. | - | - | Other features of insulin resistance *** |
| Casein Protein (g·L−1) | Whey Protein (g·L−1) | |||||
|---|---|---|---|---|---|---|
| Total Protein for Cow (24.6–28) and Buffalo (32–40) Milk | Total Protein for Cow (5.5–7) and Buffalo (6) Milk | |||||
| αs1-Casein | αs2-Casein | β-Casein | κ-Casein | β-Lactoglobulin | α-Lactalbumin | |
| Cow | 8–10.7 | 2.8–3.4 | 8.6–9.3 | 2.3–3.3 | 3.2–3.3 | 1.2–1.3 |
| Buffalo | 8.9 | 5.1 | 12.6–20.9 | 4.1–5.4 | 3.9 | 1.4 |
| Fraction of Protein | Approximate Molecular Weight (kDa) [196,197] | Amino Acids (-) [196,197] | Isoelectric Point (-) [197,198] | Denaturation Temperature (°C) [198,199] | Allergome Name [200] | |
|---|---|---|---|---|---|---|
| Casein | αs1-casein | 23.6 | 199 | 4.9–5.0 | - | Bos d 9 |
| αs2-casein | 25.2 | 207 | 5.2–5.4 | - | Bos d 10 | |
| β-casein | 24 | 209 | 5.1–5.4 | - | Bos d 11 | |
| κ-casein | 19 | 169 | 5.4–5.6 | - | Bos d 12 | |
| Whey | β-lactoglobulin | 18.3 | 162 | 5.3 | 71.9 | Bos d 5 |
| α-lactalbumin | 14.2 | 123 | 4.8 | 64.3 | Bos d 4 | |
| Immunoglobulins (IgG, IgA, IgM) | IgG 150–161, IgA 385–417, IgM 1000 | - | IgG 5.5–8.3 | 72 | - | |
| Bovine serum albumin | 66.4 | 583 | 4.9–5.1 | 72–74 | Bos d 6 | |
| Lactoferrin | 80 | 703 | 8.7 | 72–85 | - | |
| Lactoperoxidase | 78 | 612 | 9.8 | 70 | - | |
| Peptide Sequence | Precursor Protein | Source | Hydrolysis Enzymes | Bioactivities | Ref. |
|---|---|---|---|---|---|
| FFVAP | α-casein | Cow milk | Trypsin | ACE inhibitory | [214] |
| KVLPVPQ | β-casein | Proteinase | Anti-hypertensive | [215] | |
| YKVPQL | β-casein | Proteinase | ACE inhibitory | [215] | |
| KVLPVP | β-casein | Carboxypeptidase | ACE inhibitory | [215] | |
| KVLPVP | β-casein | Carboxypeptidase | Antihypertensive | [215] | |
| YGLF | α-lactalbumin | Trypsin, Pepsin | ACE inhibitory | [216] | |
| YLLF | β-lactoglobulin | Trypsin, Pepsin | ACE inhibitory | [216] | |
| KKLGAPSITCVRRAF | Lactoferrin | Pepsin | Anti-inflammatory | [217] | |
| KKLGAPSITCVRRAF | Lactoferrin | Pepsin | Antioxidant | [218] | |
| KKLGAPSITCVRRAF | Lactoferrin | Pepsin | Anticancer | [219] | |
| MAIPPKKNQDK | κ-casein | Chymosin, Trypsin | Antithrombotic | [220] | |
| AVESTVATLEDΣPEVIESPPE, where Σ is Ser(P) | κ-casein | Chymosin | Modulatory of satiety | [221] | |
| TVQVTSTAV, MAIPKKNQDKTEIP | κ-casein | Chymosin, Papain | Anti-obesity | [222] | |
| PGPIPN | β-casein | Trypsin, Chymotrypsin | Immunomodulatory | [220] | |
| YPSYGLNY | κ-casein | Trypsin, Pepsin, Chymotrypsin | Anti-opioid | [220] | |
| YGLF | α-lactalbumin | Trypsin/Pepsin | ACE inhibitory, Anti-opioid | [223] | |
| YLLF | β-lactoglobulin | Trypsin/Pepsin | ACE inhibitory, Anti-opioid | [223] | |
| HIRL | β-lactoglobulin | Trypsin/Pepsin | ACE inhibitory | [223] | |
| RYLGYL, RYLGYLE, YVPFP | α-casein | - | Anticancer | [224] | |
| YPFPGPI, YPFPG | β-casein | - | Anticancer | [224] | |
| FKCRRWQWRMKK, LGAPSITCVRRAF | Lactoferrin | - | Anticancer | [224] | |
| PYPQ, YFYPE, EMPFK, PQSV | Casein | Trypsin, Alcalase | Antioxidant | [225] | |
| YQKFPQYLQY | Casein | Pepsin, Trypsin | Antihypertensive | [226] | |
| LQPE, VAPFPE, TDVEN, VLPVPQ | Milk casein | Neutrase | Cholesterol lowering | [227] | |
| HLPGRG, QNVLPLH, PLMLP, MFE, GPAHCLL, ACGP | Milk casein | Alcalase, Pronase E | Antidiabetic activity (inhibit three diabetic-related enzymes; such as DPP-IV, α-glucosidase and α-amylase) | [228] | |
| RELEELNVPGEIVEΣLΣΣΣEESITRINK | β-casein | Chymotrypsin | Immuno-stimulatory | [229] | |
| LVYPFPGPI | β-casein | Buffalo milk | Proteinase | ACE inhibitory | [230] |
| FVAPFPE | αs1-casein | Trypsin | ACE inhibitory | [231] | |
| YQQPVL | β-casein | Fermentation + Pepsin + Trypsin | ACE inhibitory | [232] | |
| FPGPIPK, IPPK, IVPN, QPPQ | β-casein | Papain, Pepsin, or Trypsin | ACE inhibitory | [233] | |
| IPP/VPP | β-casein | - | Anti-diabetic, Antihypertensive | [205] | |
| IPP/VPP | β-casein | Proteases | Antihypertensive | [234] | |
| RNAVPITPTLNR | αs2-casein | Protease | Antidiabetic/α-glucosidase inhibitory | [235] | |
| TKVIPYVRYL | αs2-casein | Protease | Antidiabetic/α-glucosidase inhibitory | [235] | |
| YLGYLEQLLR | αs2-casein | Protease | Antidiabetic/α-glucosidase inhibitory | [235] | |
| FALPQYLK | αs2-casein | Protease | Antidiabetic/α-glucosidase inhibitory | [235] | |
| YVEELKPTPEGDL | β-lactoglobulin | Pepsin | Antioxidant | [236] | |
| VLPVPQK | β-casein | Pepsin + Trypsin | ACE inhibitory | [237] | |
| VLPVPQK | β-casein | Pepsin + Trypsin | Antioxidant | [238] | |
| YPSG, HPFA, KFQ | β-casein | Papain, Pepsin, Trypsin | Antioxidant | [233] | |
| RELEE, TVA, MEDNKQ | Casein | Trypsin, Alcalase | Antioxidant | [225] |
| Commercial Brand Name | Food Type | Peptide Sequence | Health/Functional Claim | Manufacturers |
|---|---|---|---|---|
| Ameal bp®, Ameal peptide® | Tablets | VPP, IPP derived from β-casein and κ-casein | Reduction in blood pressure | Calpis Co., Tokyo, Japan |
| BioZate 1 | Hydrolyzed whey protein isolate | Fragments from β-lactoglobulin | Reduction in blood pressure | Davisco Foods International Inc., Savage, MN, USA |
| BiPRO WPI | WPI | GMP (106–109) | Antithrombic and Anticariogenic | Davisco Foods International Inc., Savage, MN, USA |
| Calpis | Sour milk | IPP and VPP | Hypotensive | Calpis Co., Tokyo, Japan |
| Calpico®/Calpis® AMEALs | Fermented milk | IPP and VPP | Hypotensive | Calpis Co., Tokyo, Japan |
| C12 peptide | Peptide ingredient | FFVAPFPEVFGK | Hypotensive | DMV International, Veghel, The Netherland |
| Evolus® | Fermented milk | IPP and VPP | Reduction in blood pressure | Valio Ltd., Helsinki, Finland |
| GC tooth mousse | Water-based creme | Caseinomacropeptide | Anticariogenic | GC Europe N.V., Leuven, Belgium |
| Glycomacropeptide (GMP) | Fresh cheese, WPI | κ-casein f(106–169) | Anticariogenic and Antithrombotic | Davisco Foods International Inc., Savage, MN, USA |
| insuVidaTM | Tablets | Casein hydrolysate | Antidiabetic | DSM Food Specialties, Delft, The Netherland |
| Lactium® | Milk protein hydrolysate | αs1-casein f(91–100) YLGYLEQLLR | Reduction in stress effects | Ingredia Nutritional, Arras, France |
| LACPRODAN-DI-3065 | Healthy food formula | Whey protein hydrolysate (WPH) | Benefits to sufferers from impaired digestion as a result of surgery, illness or health conditions, such as short bowel syndrome, pancreatic insufficiency and inflammatory bowel disease. | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-DI-3091 | Healthy food formula | WPH | Supplements for patients with maldigestion or malabsorption. | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-DI-3092 | Healthy food formula | WPH | Supplements for patients with maldigestion or malabsorption. | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-IF-3070 | Healthy food formula | WPH | Sustainable gastrointestinal system for infant | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-IF-3071 | Healthy food formula | WPH | Sustainable gastrointestinal system for infant | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-IF-3080 | Healthy infant food formula | WPH | Lower allergenic | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-IF-3090 | Healthy infant food formula | WPH | Lower allergenic | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-DI-2021 | Healthy food formula | Casein phosphopeptides | Increase calcium absorption and bone health | Arla Foods Ingredients Group P/S, Viby, Denmark |
| LACPRODAN-CGMP-10 and CGMP-20 | Healthy food formula | Fragment from α-lactalbumin | Improves sleep and memory | Arla Foods Ingredients Group P/S, Viby, Denmark |
| MI pasteTM/MI paste plusTM | Toothpaste | Caseinomacropeptide | Anticariogenic | GC America, Alsip, IL, USA |
| Pep2Dia® | Capsule (vegetable fiber with 350 mg of milk protein hydrolysate/capsule) | RP | Antidiabetic | Ingredia S.A., Arras Cedex, France |
| PROTARMOR™ 80 | Ingredient | Casein hydrolysates | Weight loss | Armor Proteines, Maen Roch, France |
| RecaldentTM | Ingredient | Caseinomacropeptide | Anticariogenic | Cadbury Enterprises Pte. Ltd., Jurong, Singapore |
| Trident xtra careTM | Chewing gum | Caseinomacropeptide | Anticariogenic | Cadbury Adams, East Hanover, NJ, USA |
| Publication Year/Reference | Study Design, Duration | Characteristics and Number of Subjects (n) | Dairy Milk Proteins Formula | Comparison Formula | Effects and Remarks | |
|---|---|---|---|---|---|---|
| Hyperglycemia | Dyslipidemia | |||||
| 2005/[320] | Randomized, Acute (2 separate occasions with ≥1 week gap between each) | T2DM (14) | Whey protein (27.6 g) | Ham (96 g) and Lactose (5.3 g) | ↓ Postprandial plasma glucose by whey protein. ↑ Plasma insulin response and postprandial GIP by whey protein. → GLP-1 by whey protein and ham. | - |
| 2005/[321] | Randomized order and a double-blind (2 trials, separated by a 2 weeks gap between each) | T2DM (10) | No diabetes (n = 10), 0.7 g carbohydrate/kg body weight/h (50% glucose and 50% maltodextrin) with or without 0.35 g/kg body weight/h of a protein hydrolysate and AA mixture (50% casein hydrolysate, 25% free Leu and 25% free phenylalanine) every 15 min until t = 165 min | No diabetes (n = 9), 0.7 g carbohydrate/kg body weight/h (50% glucose and 50% maltodextrin) with or without 0.35 g/kg body weight/h of a protein hydrolysate and AA mixture (50% casein hydrolysate, 25% free Leu and 25% free phenylalanine) every 15 min until t = 165 min | ↑ Plasma insulin and ↓ plasma glucose responses by carbohydrate + protein trial than carbohydrate trial in diabetic subjects and matched to control subjects. | - |
| 2006/[322] | Randomized cross-over (4 occasions with 7 days gap between each) | Overweight (19) | Preloads made by water solutions of (a) WPI (55 g), (b) Calcium caseinate (55 g), (c) Glucose (60 g), (d) Lactose (56 g) | - | ↓ Acute appetite and energy intake by casein- or whey-, lactose- than glucose-preload. ↓ Postprandial plasma glucose by protein (mean value of casein and WPI)-preload than lactose- and glucose-preload. → Plasma insulin by different preloads. ↑ CCK response and ↓ ghrelin by protein preload. | - |
| 2006/[323] | Randomized, double-blind, placebo-controlled, cross-over, acute challenge (Participants consumed 3 meals/day in a single 24 h period) | T2DM (11), Healthy (11) | T2DM (11), Casein hydrolysate (0.3 g/kg body weight) + Leu (0.1 g/kg body weight) + meal (64% carbohydrate, 25% fat and 11% protein) | Healthy (11), Flavored water + meal (64% carbohydrate, 25% fat and 11% protein) | ↓ Plasma glucose by casein hydrolysate in average 24 h by casein and Leu supplemented meal. | - |
| 2006/[324] | Acute, randomized, double-blind (3 trials, separated by at least 1 week gap between each) | T2DM (10), Healthy (10) | T2DM (10), (a) Carbohydrate (50% glucose and 50% maltodextrin) (0.7 g/kg body weight), (b) Carbohydrate + Casein hydrolysate (0.7 g/kg body weight + 0.3 g/kg body weight), (c) Carbohydrate + Casein hydrolysate + Leu (0.7 g/kg body weight + 0.3 g/kg body weight + 0.1 g/kg body weight) | Healthy (10), Similar diet like diabetic group | ↑ Insulin response and ↓ plasma glucose response in subjects with T2DM and control subjects by Carbohydrate + Casein hydrolysate and Carbohydrate + Casein hydrolysate + Leu for both T2DM and healthy subjects (Carbohydrate diet basis). | - |
| 2007/[325] | Double-blind, randomized, controlled cross-over clinical trial (5 days study with 2 weeks gap between each) | Non-obese, T2DM (12) | Mixed meal (≈31% carbohydrates, ≈17% lipids and ≈52% proteins as total calories) with (a) WPI (fast protein), (b) micellar casein (slow protein), (c) a mixture of FAAs resembling the AA composition of micellar casein. | - | ↓ Postprandial plasma glucose by FAA meal than WPI meal and casein meal. ↑ Plasma BCAAs, EAAs, C-peptide, insulin and pro-insulin concentrations by WPI meal than casein meal and similar with FAA meal. ↓ Plasma GLP-1 response by casein meal than WPI meal. ↑ Plasma GIP response by WPI meal and casein meal than FAA meal. | - |
| 2009/[326] | Acute (3 occasions) | T2DM (8) | (a) Pre-meal: 55 g whey in 350 mL beef soup, (b) Main-meal: 55 g whey in potato | No whey in pre-meal and main meal | ↑ Plasma insulin, GIP and CCK by whey diet. ↓ Gastric emptying and postprandial glycemia by whey diet. | - |
| 2009/[327] | Randomized crossover (4 separate occasions with 2–5 weeks gap between each) | T2DM (12) | Whey protein (45 g) in meal (80 g fat and 45 g carbohydrate) | (a) Casein (45 g), (b) Cod (45 g), (c) Gluten (45 g) in meal (80 g fat and 45 g carbohydrate) | ↑ Postprandial plasma insulin and incretins (GLP-1, GIP) by whey protein. → Glucagon by all types of proteins. | ↑ Postprandial lipemia by whey protein than other proteins. |
| 2009/[328] | Randomized (Energy restriction period of 5–6 weeks followed by a weight maintenance period of 12 weeks with 1 week in-between to change back from liquid to normal food) | Overweight and obese (48) | (a) Casein, (b) Whey protein supplements (2 × 25 g/day) | Maltodextrin | ↑ Plasma glucagon. Fasting glucose in normal range by protein diet. → Plasma insulin resistance by casein and whey proteins. | ↑ Serum TG by maltodextrin. → Serum total cholesterol, leptin, adiponectin, LDL-C and HDL-C by protein diets. |
| 2010/[329] | Randomized, single blind, three-way crossover design (3 separate intervention days, each preceded by a 1 week washout period) | Overweight or obese (20) | (a) WPI (45 g), (b) Sodium caseinate (45 g) | Glucose (45 g) | ↓ Plasma glucose by WPI than sodium caseinate and glucose. | ↓ Serum TG-enriched lipoprotein by WPI than casein and glucose. → Serum total cholesterol, LDL-C, HDL-C, NEFA, apolipoprotein B-48, insulin and leptin. |
| 2010/[330] | Randomized, controlled, cross-over, acute challenge trial (4 test meals with intervals of >2 weeks) | T2DM (11) | (a) Casein (45 g) + Carbohydrate (45 g) + Fat (80 g), (b) Casein (45 g) + Fat (80 g), (c) Carbohydrate (45 g) + Fat (80 g) | Fat (80 g) | ↑ Plasma insulin and glucagon by Casein + Fat meal and Casein + Carbohydrate + Fat meal than Fat-meal. ↑ GIP response by Casein + Carbohydrate + Fat meal. → GLP-1 response. | → Serum TG and retinyl palmitate in the chylomicron-rich fraction for all meals. ↑ Retinyl palmitate in the chylomicron-poor fraction by 45 g of protein as casein. |
| 2011/[259] | Randomized parallel (12 weeks) | Overweight and obese (70) | (a) WPI (27 g) in 250 mL of water, (b) Sodium caseinate (27 g) in 250 mL of water. Twice in a day | Glucose (27 g) in 250 mL of water | → Plasma glucose by WPI than control or casein meal. ↑ Fasting plasma insulin by whey protein. | ↓ Fasting serum TG, plasma total cholesterol and LDL-C by WPI. |
| 2011/[331] | Randomized double-blind partial cross-over (3 out of 4 treatments separated by a week gap between each) | T2DM (36) | (a) Intact casein, (b) Casein hydrolysate, (c) Casein hydrolysate with Leu | Carbohydrate (maltodextrin and glucose monohydrate) without (a) intact casein, (b) unhydrolyzed casein, (c) Casein hydrolysate and Leu | ↓ Plasma glucose by casein hydrolysate with or without Leu. | - |
| 2011/[332] | Randomized placebo controlled double blind (Separate 3 days with 1 week gap between each) | T2DM (13) | In a single oral bolus (300 mL) containing 50 g of carbohydrates (50% glucose and 50% maltodextrin and casein hydrolysate (6 g or 12 g) | No casein hydrolysate | → Plasma insulin and glucose by 6 g of casein hydrolysate. ↑ Post-challenge plasma insulin and ↓ glucose levels by 12 g of casein hydrolysate. | - |
| 2012/[333] | Single blind crossover (4 separate occasions with a washout period ⩾2 weeks) | T2DM (12) | Protein supplement ((a) LACPRODAN-ALPHA-10 (45 g), (b) LACPRODAN-DI-9224 (45 g), (c) LACPRODAN CGMP-10 (45 g), (d) LACPRODAN-DI-3065 (45 g)) + Fat (80 g) + Carbohydrate meal (45 g) | - | ↑ Plasma glucose, insulin, glucagon and GLP-1 by LACPRODAN-DI-3065 than other proteins in fat-carbohydrate meals. | → Postprandial serum TG by all dietary proteins. ↑ Retinyl palmitate in the chylomicron-rich fraction by LACPRODAN-DI-3065 than dietary proteins. → FFA and Retinyl palmitate in the chylomicron-poor fraction by all dietary proteins. |
| 2013/[334] | Randomized crossover (4 different meals in different days with a 2 weeks washout period between each meal) | Obese non-diabetic (11) | WPI + Fat (80 g) + Carbohydrate (45 g) | Carbohydrate (45 g) + Fat (80 g) + (a) Cod or casein (45 g), (b) gluten protein (45 g) | ↑ Biomarkers (GLP-1, insulin, glucagon) of postprandial glycemia, ↓ Postprandial GIP by WPI. The larger initial plasma insulin and glucagon response after whey meal did not correlate with the initial GLP-1 or GIP responses. | ↓ Postprandial serum TG by WPI than cod and gluten proteins. ↓ NEFA by whey and casein than cod and gluten proteins. → Retinyl palmitate in chylomicron by all types of dietary proteins. |
| 2013/[335] | Double-blind, randomized (Each trial was performed on a distinct day with 3 days intervals between each trial) | T2DM (10) | (a) WPH beverage (0.1 g/kg body weight, 0.2 g/kg body weight and 0.4 g/kg body weight), (b) WPI beverage (0.1 g/kg body weight, 0.2 g/kg body weight and 0.4 g/kg body weight) | Distilled water | ↑ Postprandial plasma insulin and ↓ post-challenge plasma glucose by 0.2 g/kg body weight WPH or 0.4 g/kg body weight WPI. ↑ Postprandial plasma insulin and ↓ concomitant glucose level to normal range at 2 h after 0.2 g/kg body weight WPH. | - |
| 2013/[336] | Randomized crossover (4 test days, which were separated by a washout period of at least 3 days) | Obese men (16) | Butter cake (high fat) + (a) 500 mL milk or (b) 500 mL water + milk protein 23.4 g or (c) 500 mL water + calcium 2.3 g | Butter cake + 500 mL water | ↑ Plasma insulin and ↓ glucose by milk protein. ↑ Plasma concentrations of total AAs, EAAs and non-EAAs by milk and protein milk. | ↑ Serum TG by milk protein. ↓ Apolipoprotein B-48 by calcium meal compared with milk. → NEFA by all drinks. |
| 2014/[337] | Double-blind randomized and cross-over (3 trials was separated by at least 6 days gap between each) | T2DM (60) | (a) Carbohydrate + Intact casein (0.7 g/kg body weight + 0.3 g/kg body weight), (b) Carbohydrate + Casein hydrolysate (0.7 g/kg body weight + 0.3 g/kg body weight) | Carbohydrate (50% glucose + 50% maltodextrin) (0.7 g/kg body weight) | ↑ Plasma insulin and ↓ plasma glucose responses by Carbohydrate + Protein diet than Carbohydrate diet. ↑ Plasma insulin response by Carbohydrate + Casein hydrolysate diet than Carbohydrate + Intact protein diet. ↓ Plasma glucose response by Carbohydrate + Casein hydrolysate diet than Carbohydrate + Intact protein diet. | - |
| 2014/[338] | Randomized, open-label crossover (2 meals in 2 separate days and a at least 2 weeks gap between each) | T2DM (15) | Preload: 50 g whey in 250 mL water | Preload: 250 mL water | ↓ Plasma glucose, ↑ plasma insulin, ↑ C-peptide, ↑ GLP-1, and → DPP4 by whey pre-load. | - |
| 2015/[339] | Randomized, cross-over study (4 weeks) | T2DM (7) | Preload: WPI (25 g) + 25 g chocolate-flavor in 100 mL water | Preload: 25 g chocolate-flavor without WPI in 100 mL water | ↓ Postprandial plasma blood glucose and peak blood glucose by WPI formula. | - |
| 2015/[340] | Single-center, randomized, single blind (3 blind challenges on 3 different occasions with a minimum of 2 days gap between them) | Healthy (15), Prediabetic (15) | 50 g casein or WPI + 50 g of maltodextrin with dextrose equivalent 19 + 2 g of hydroxyproline + 10 g of lactulose in 300 mL of water. | 50 g of maltodextrin with dextrose equivalent 19 + 2 g of hydroxyproline + 10 g of lactulose in 300 mL of water. | ↑ Plasma insulin, glucagon, C-peptide, GIP, GLP-1 and satiety by casein and whey proteins than glucose. Plasma glucose by casein and whey proteins than glucose. → All effects two proteins. | - |
| 2015/[341] | Randomized, parallel-controlled, double-blinded (Each test meal with 12 weeks intervention period) | Obese (52) | (a) Whey + Low medium-chain saturated fatty acids, (b) Whey + High medium-chain saturated fatty acids, (c) Casein + Low medium-chain saturated fatty acids, (d) Casein + High medium-chain saturated fatty acids | - | ↓ Postprandial apolipoprotein B-48 response after whey compared with casein independently of fatty acid composition. ↑ Postprandial plasma GLP-1 by casein compared with whey. → Postprandial plasma insulin, glucose, glucagon, or GIP among groups. | → Postprandial serum triacylglycerol and FFA among groups. |
| 2017/[342] | Randomized open-label parallel-arm (12 weeks, dietary intake on 3 days in each week) | T2DM (56) | (a) Whey protein breakfast: 25% fat + 50% carbohydrates + 25% protein (28 g whey from 42 g total protein). (b) Protein breakfast: 25% fat + 50% carbohydrates + 25% (42 g) protein mainly from eggs (7 g), tuna (20 g), soya (7 g). | Carbohydrate breakfast: 25% fat + 64% carbohydrates + 11% (17 g) soya protein | ↓ Overall plasma postprandial incremental area under curve (iAUC) for glucose, ghrelin and hunger scores by whey protein breakfast and protein breakfast than carbohydrate breakfast. ↑ Postprandial plasma overall iAUC for insulin, C-peptide, GLP-1 and satiety scores by whey protein breakfast than protein breakfast and carbohydrate breakfast. | - |
| 2017/[343] | (a) Parallel-armed acute challenge (One serving (21 g) of whey protein) and (b) Crossover design (continuous glucose monitoring (CGM) twice, over 2 consecutive weeks, 3.5 days each week) | n = 18 underwent a challenge test (not crossover design). n = 22 underwent CGM and controlled feeding twice (crossover design), Two consecutive weeks: one week WPI and other week placebo diet. | WPI (21 g protein + 3 g carbohydrate + 0.5 g fat) | Indigestible potato starch (1 g protein + 25 g carbohydrate 20 g fiber + 0.5 g fat) | Acute challenge studies: ↑ Plasma insulin, ↑ GLP-1, ↓ Plasma glucose and ↓ Ghrelin by WPI diet. Placebo diet had no effect. | ↓ Hypertriglyceridemia by WPI diet. |
| 2018/[344] | Randomized, double-blind, placebo-controlled (10 weeks) | T2DM (24) | WPI beverage (20 g protein + 10 g carbohydrate + 3 g milk fat) | Without WPI (30 g carbohydrate + 3 g milk fat) | ↓ Fasting plasma blood glucose and insulin resistance by whey protein diet. | - |
| 2018/[345] | Randomized crossover (2 separate test days with 1 week gap between each) | Overweight and obese (9) | Breakfast (74 g carbohydrate, 2.1 g fat and 8.5 g protein in 100 mL orange juice) with 10% (w/v) solution of a novel casein hydrolysate | Breakfast (74 g carbohydrate, 2.1 g fat and 8.5 g protein in 100 mL orange juice) with 10% (w/v) solution of sodium caseinate (intact protein). | → Gastric emptying outcome by intact casein and hydrolysate. ↑ Insulin response and ↓ glucose response by casein hydrolysate compared to intact casein. | - |
| 2018/[346] | Randomized, single-blind crossover (3 separated occasions separated by 7 days gap between each) | T2DM (11) | (a) Intact whey protein (15 g), (b) WPH (15 g) before mixed-macronutrient breakfast and lunch meals, separated by 3 h. | Flavored water | ↓ Plasma glucose by WPH (early) and intact whey protein. ↑ Satiety and Plasma insulin by both WPH and intact whey protein. | - |
| 2018/[347] | Acute, randomized, cross-over (Test days separated by a washout period of approximately 1 week gap between each) | Non-diabetic (12), T2DM (12) | Preload: 20 g whey protein in 200 mL water as pre-meal or part of the fat-rich meal. | Preload: 200 mL water | Plasma ↑ insulin, ↑ glucagon, ↑ GIP and ↓ gastric emptying in subjects with and without T2DM by whey protein pre-meal. Plasma ↑ insulin, ↑ glucagon, ↑ GIP and ↓ gastric emptying in subjects with and without T2DM by pre-meal than main meal. | → Postprandial TG, apolipoprotein B-48 and NEFA in subjects with and without T2DM. |
| 2018/[348] | Acute, randomized, cross-over (The test days were separated by a wash-out period of approximately 1 week gap between each) | MS (20) | Preload: (a) 10 g whey protein, (b) 20 g whey protein | Preload: No whey protein | ↑ Insulin response, ↑ postprandial glucagon, ↓ glucose and ↓ gastric emptying after a pre-meal with 20 g whey protein than 10 g whey protein and placebo. → GIP by whey protein pre-meal. | → TG, apolipoprotein B-48 and FFA by whey protein pre-meal. |
| 2019/[349] | Randomized, double-blind, placebo-controlled, monocentric, 3-way-cross-over (6 weeks, a wash-out period of 7 days between each study day) | Prediabetic (21) | Milk protein hydrolysate (1.4 g and 2.8 g), 2 dosages/day | Maltodextrin with dextrose equivalent of 9 | ↓ iAUC of plasma glucose by milk protein hydrolysate in dose-dependent manner. → Insulinotropic properties were insignificant. | - |
| 2020/[350] | Prospective randomized pilot study (45 days) | Obese and insulin resistance (48) | Protein diet (90 g/meal), 5 meals/day for (a) whey protein, (b) vegetable protein (soya, green peas, or cereals), (c) animal protein (meat, fish, egg) | - | ↓ Insulin resistance and fasting glycemia by whey protein. | Serum ↓ total cholesterol, LDL-C and TG by whey protein. |
| 2020/[351] | Double-blind randomized clinical trial (13 weeks) | Obese and type 2 (pre-)diabetes (123) | 21 g of Leu-enriched whey protein ((3 g total Leu), 9 g carbohydrates, 3 g fat, 800 IU cholecalciferol (Vitamin D3)) | Carbohydrate (25 g) and fat (6 g) mix | ↓ Insulin resistance and postprandial plasma glucose by Leu-enriched whey protein. | - |
| 2020/[352] | Single-center randomized, placebo-controlled (3 months) | T2DM (120) | Cys-rich (2.7%) WPI with a standardized lactoferrin in 100 mL of water | 5 g casein in 100 mL of water | ↓ Fasting plasma glucose by Cys-rich WPI diet. | Serum ↓ total cholesterol, triacylglyceride and LDL-C levels by Cys-rich (2.7%) WPI than placebo. |
| 2020/[319] | Randomized, parallel, placebo-controlled, double-blind study (20 weeks: 1 week pre-study measurement period, a 3 weeks baseline period and a 16 weeks energy restriction intervention period | Overweight or obese | n = 21, Unhealthy western-style eating patterns + 1.5 g total protein (MPI)/kg body weight/day | n = 23, Unhealthy western-style eating patterns + 0.8 g total protein (MPI)/kg body weight/day | → Fasting plasma insulin and glucose | Serum ↓ fasting TG, → total cholesterol and LDL-C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdisa, K.B.; Szerdahelyi, E.; Molnár, M.A.; Friedrich, L.; Lakner, Z.; Koris, A.; Toth, A.; Nath, A. Metabolic Syndrome and Biotherapeutic Activity of Dairy (Cow and Buffalo) Milk Proteins and Peptides: Fast Food-Induced Obesity Perspective—A Narrative Review. Biomolecules 2024, 14, 478. https://doi.org/10.3390/biom14040478
Abdisa KB, Szerdahelyi E, Molnár MA, Friedrich L, Lakner Z, Koris A, Toth A, Nath A. Metabolic Syndrome and Biotherapeutic Activity of Dairy (Cow and Buffalo) Milk Proteins and Peptides: Fast Food-Induced Obesity Perspective—A Narrative Review. Biomolecules. 2024; 14(4):478. https://doi.org/10.3390/biom14040478
Chicago/Turabian StyleAbdisa, Kenbon Beyene, Emőke Szerdahelyi, Máté András Molnár, László Friedrich, Zoltán Lakner, András Koris, Attila Toth, and Arijit Nath. 2024. "Metabolic Syndrome and Biotherapeutic Activity of Dairy (Cow and Buffalo) Milk Proteins and Peptides: Fast Food-Induced Obesity Perspective—A Narrative Review" Biomolecules 14, no. 4: 478. https://doi.org/10.3390/biom14040478
APA StyleAbdisa, K. B., Szerdahelyi, E., Molnár, M. A., Friedrich, L., Lakner, Z., Koris, A., Toth, A., & Nath, A. (2024). Metabolic Syndrome and Biotherapeutic Activity of Dairy (Cow and Buffalo) Milk Proteins and Peptides: Fast Food-Induced Obesity Perspective—A Narrative Review. Biomolecules, 14(4), 478. https://doi.org/10.3390/biom14040478