Haplotype- and SNP-Based GWAS for Growth and Wood Quality Traits in Eucalyptus cladocalyx Trees under Arid Conditions


 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
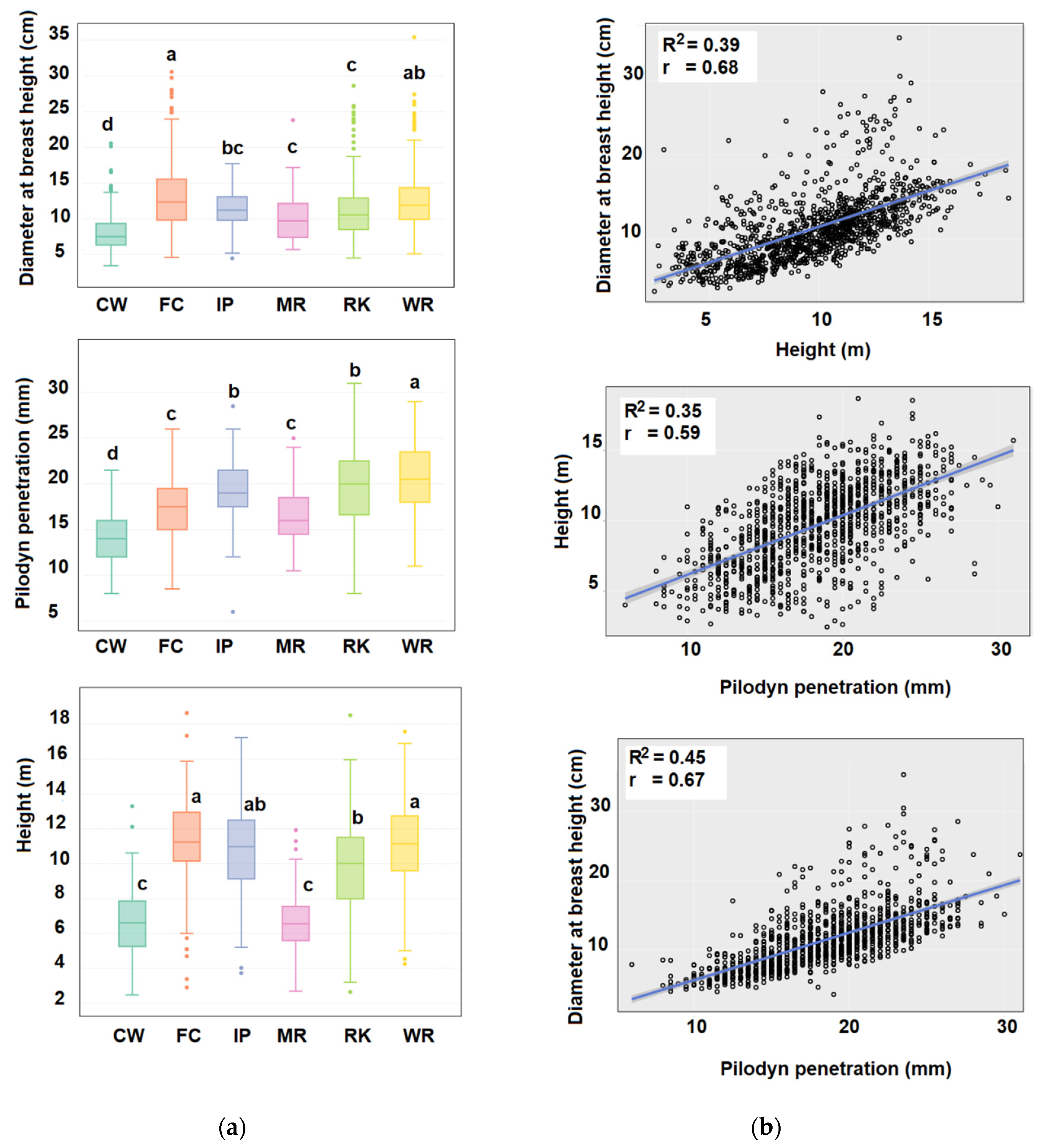
2.1. Phenotypic Diversity and Population Genetic Structure of E. cladocalyx
2.2. Linkage Disequilibrium (LD) Pattern and Haplotype Blocks
2.3. Marker-Trait Associations (MTAs)
2.4. Identification of Candidate Genes Controlling the Variation in Quantitative Traits in E. cladocalyx
2.5. Pleiotropic Loci
3. Discussion
3.1. Phenotypic Variability
3.2. Population Genetic Structure
3.3. Genome-Wide Association Study and Pleiotropic Loci Detection
4. Materials and Methods
4.1. Plant Material and Phenotypic Evaluation
4.2. DNA Extraction and Genotyping
4.3. Estimation of Linkage Disequilibrium (LD) and Identification of Haplotype Blocks
4.4. Genome Wide Association Study (GWAS)
4.5. Detection of Pleiotropic Loci
4.6. Identification of Candidate Genes for Traits Related to Growth and Wood Quality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Hejazi, M.; Liu, L.; Calvin, K.; Clarke, L.; Edmonds, J.; Kyle, P.; Patel, P.; Wise, M.; Davies, E. Balancing global water availability and use at basin scale in an integrated assessment model. Clim. Chang. 2016, 136, 217–231. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The central Chile mega drought (2010–2018): A climate dynamics perspective. Int. J. Climatol. 2019, 40, 421–439. [Google Scholar] [CrossRef]
- Miranda, A.; Lara, A.; Altamirano, A.; Di Bella, C.; González, M.E.; Camarero, J.J. Forest browning trends in response to drought in a highly threatened mediterranean landscape of South America. Ecol. Indic. 2020, 115, 106401. [Google Scholar] [CrossRef]
- Roco, L.; Engler, A.; Bravo-Ureta, B.E.; Jara-Rojas, R. Farmers’ perception of climate change in Mediterranean Chile. Reg. Environ. Chang. 2014, 15, 867–879. [Google Scholar] [CrossRef]
- Ballesta, P.; Mora, F.; Del Pozo, A. Association mapping of drought tolerance indices in wheat: QTL-rich regions on chromosome 4A. Sci. Agric. 2020, 77, e20180153. [Google Scholar] [CrossRef] [Green Version]
- Beech, E.; Rivers, M.; Oldfield, S.; Smith, P.P. GlobalTreeSearch: The first complete global database of tree species and country distributions. J. Sustain. For. 2017, 36, 454–489. [Google Scholar] [CrossRef]
- De Sá Martins, R.; Faria, J.M.R.; Rossini, B.C.; Marino, C.L.; Dos Santos, L.D.; José, A.C. Proteomic analyses unraveling water stress response in two Eucalyptus species originating from contrasting environments for aridity. Mol. Biol. Rep. 2020, 47, 5191–5205. [Google Scholar] [CrossRef]
- Arriagada, O.; do Amaral, A.T., Jr.; Mora, F. Thirteen years under arid conditions: Exploring marker-trait associations in Eucalyptus cladocalyx; for complex traits related to flowering, stem form and growth. Breed. Sci. 2018, 68, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Mora, F.; Ballesta, P.; Serra, N. Bayesian analysis of growth, stem straightness and branching quality in full-sib families of Eucalyptus globulus. Bragantia 2019, 78, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Ballesta, P.; Serra, N.; Guerra, F.P.; Hasbún, R.; Mora, F. Genomic prediction of growth and stem quality traits in Eucalyptus globulus Labill. at its southernmost distribution limit in Chile. Forests 2018, 9, 779. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, C.; Mora-Poblete, F.; Contreras-Soto, R.; Ahmar, S.; Chen, J.T. Genome-wide prediction of complex traits in two outcrossing plant species through Deep Learning and Bayesian Regularized Neural Network. Front. Plant Sci. 2020, 11, 1734. [Google Scholar] [CrossRef] [PubMed]
- Bush, D.; Kain, D.; Kanowski, P.; Matheson, C. Genetic parameter estimates informed by a marker-based pedigree: A case study with Eucalyptus cladocalyx in southern Australia. Tree Genet. Genomes 2015, 11, 798. [Google Scholar] [CrossRef]
- Mora, F.; Arriagada, O.; Ballesta, P.; Ruiz, E. Genetic diversity and population structure of a drought-tolerant species of Eucalyptus, using microsatellite markers. J. Plant Biochem. Biotechnol. 2017, 26, 274–281. [Google Scholar] [CrossRef]
- Mora, F.; Gleadow, R.; Perret, S.; Scapim, C.A. Genetic variation for early flowering, survival and growth in sugar gum (Eucalyptus cladocalyx F. Muell) in southern Atacama Desert. Euphytica 2009, 169, 335–344. [Google Scholar] [CrossRef]
- Cané-Retamales, C.; Mora, F.; Vargas-Reeve, F.; Perret, S.; Contreras-Soto, R. Bayesian threshold analysis of breeding values, genetic correlation and heritability of flowering intensity in Eucalyptus cladocalyx under arid conditions. Euphytica 2011, 178, 177–183. [Google Scholar] [CrossRef]
- Valenzuela, C.E.; Ballesta, P.; Maldonado, C.; Baettig, R.; Arriagada, O.; Sousa Mafra, G.; Mora, F. Bayesian mapping reveals large-effect pleiotropic QTLs for wood density and slenderness index in 17-year-old trees of Eucalyptus cladocalyx. Forests 2019, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Woodrow, I.E.; Slocum, D.J.; Gleadow, R.M. Influence of water stress on cyanogenic capacity in Eucalyptus cladocalyx. Funct. Plant Biol. 2002, 29, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Pičmanová, M.; Neilson, E.H.; Motawia, M.S.; Olsen, C.E.; Agerbik, N.; Gray, C.J.; Flitsch, S.; Meier, S.; Silvestro, D.; Jørgensen, K.; et al. A recycling pathway for cyanogenic glycosides evidenced by the comparative metabolic profiling in three cyanogenic plant species. Biochem. J. 2015, 469, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Maseda, P.H.; Fernández, R.J. Stay wet or else: Three ways in which plants can adjust hydraulically to their environment. J. Exp. Bot. 2006, 57, 3963–3977. [Google Scholar] [CrossRef]
- Limousin, J.M.; Longepierre, D.; Huc, R.; Rambal, S. Change in hydraulic traits of Mediterranean Quercus ilex subjected to long-term throughfall exclusion. Tree Physiol. 2010, 30, 1026–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-StPaul, N.K.; Limousin, J.M.; Vogt-Schilb, H.; Rodríguez-Calcerrada, J.; Rambal, S.; Longepierre, D.; Misson, L. The temporal response to drought in a Mediterranean evergreen tree: Comparing a regional precipitation gradient and a throughfall exclusion experiment. Glob. Chang. Biol. 2013, 19, 2413–2426. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Abdullah, M.; Ahmar, S.; Yasir, M.; Iqbal, M.S.; Yasir, M.; Ur Rehman, S.; Ahmed, S.; Rana, R.M.; Ghafoor, A.; et al. Incredible Role of Osmotic Adjustment in Grain Yield Sustainability under Water Scarcity Conditions in Wheat (Triticum aestivum L.). Plants 2020, 9, 1208. [Google Scholar] [CrossRef]
- Dutkowski, G.W.; Potts, B.M. Genetic variation in the susceptibility of Eucalyptus globulus to drought damage. Tree Genet. Genomes 2012, 8, 757–773. [Google Scholar] [CrossRef]
- Pfautsch, S.; Harbusch, M.; Wesolowski, A.; Smith, R.; Macfarlane, C.; Tjoelker, M.G.; Reich, P.B.; Adams, M.A. Climate determines vascular traits in the ecologically diverse genus Eucalyptus. Ecol. Lett. 2016, 19, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Hacke, U.G.; Jacobsen, A.L.; Pratt, R.B. Xylem function of aridland shrubs from California, USA: An ecological and evolutionary analysis. Plant Cell Environ. 2009, 32, 1324–1333. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.B.; North, G.B.; Jacobsen, A.L.; Ewers, F.W.; Davis, S.D. Xylem root and shoot hydraulics is linked to life history type in chaparral seedlings. Funct. Ecol. 2010, 24, 70–81. [Google Scholar] [CrossRef]
- Lens, F.; Vos, R.A.; Charrier, G.; Van Der Niet, T.; Merckx, V.; Baas, P.; Aguirre-Gutierrez, J.; Jacobs, B.; Chacón-Dória, L.; Smets, E.; et al. Scalariform-to-simple transition in vessel perforation plates triggered by differences in climate during the evolution of Adoxaceae. Ann. Bot 2016, 118, 1043–1056. [Google Scholar] [CrossRef] [Green Version]
- Petit, G.; Pfautsch, S.; Anfodillo, T.; Adams, M.A. The challenge of tree height in Eucalyptus regnans: When xylem tapering overcomes hydraulic resistance. New Phytol. 2010, 187, 1146–1153. [Google Scholar] [CrossRef]
- Poorter, L.; McDonald, I.; Alarcón, A.; Fichtler, E.; Licona, J.C.; Peña-Claros, M.; Sterck, F.; Villegas, Z.; Sass-Klaassen, U. The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. New Phytol. 2010, 185, 481–492. [Google Scholar] [CrossRef]
- Olson, M.E.; Anfodillo, T.; Rosell, J.A.; Petit, G.; Crivellaro, A.; Isnard, S.; León-Gómez, C.; Alvarado-Cárdenas, L.O.; Castorena, M. Universal hydraulics of the flowering plants: Vessel diameter scales with stem length across angiosperm lineages, habits and climates. Ecol. Lett. 2014, 17, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Givnish, T.J.; Wong, S.C.; Stuart-Williams, H.; Holloway-Phillips, M.; Farquhar, G. Determinants of maximum tree height in Eucalyptus species along a rainfall gradient in Victoria, Australia. Ecology 2014, 95, 2991–3007. [Google Scholar] [CrossRef] [Green Version]
- McLean, E.H.; Prober, S.M.; Stock, W.D.; Steane, D.A.; Potts, B.M.; Vaillancourt, R.E.; Byrne, M. Plasticity of functional traits varies clinally along a rainfall gradient in Eucalyptus tricarpa. Plant Cell Environ. 2014, 37, 1440–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saadaoui, E.; Yahia, K.B.; Dhahri, S.; Jamaa, M.L.B.; Khouja, M.L. An overview of adaptative responses to drought stress in Eucalyptus spp. For. Stud. 2017, 67, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Du, Q.; Yang, X.; Xie, J.; Quan, M.; Xiao, L.; Lu, W.; Tian, J.; Gong, C.; Chen, J.; Li, B. Time-specific and pleiotropic quantitative trait loci coordinately modulate stem growth in Populus. Plant Biotechnol. J. 2019, 17, 608–624. [Google Scholar] [CrossRef] [Green Version]
- Blackman, C.J.; Creek, D.; Maier, C.; Aspinwall, M.J.; Drake, J.E.; Pfautsch, S.; O’Grady, A.; Delzon, S.; Medlyn, B.E.; Tissue, D.T.; et al. Drought response strategies and hydraulic traits contribute to mechanistic understanding of plant dry-down to hydraulic failure. Tree Physiol. 2019, 39, 910–924. [Google Scholar] [CrossRef]
- González-Martínez, S.C.; Huber, D.; Ersoz, E.; Davis, J.M.; Neale, D.B. Association genetics in Pinus taeda L. II. Carbon isotope discrimination. Heredity 2008, 101, 19–26. [Google Scholar] [CrossRef]
- González-Martínez, S.C.; Wheeler, N.C.; Ersoz, E.; Nelson, C.D.; Neale, D.B. Association genetics in Pinus taeda L. I. wood properties traits. Genetics 2007, 17, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Lepoittevin, C.; Harvengt, L.; Plomion, C.; Garnier-Géré, P. Association mapping for growth, straightness and wood chemistry traits in the Pinus pinaster Aquitaine breeding population. Tree Genet. Genomes 2012, 8, 113–126. [Google Scholar] [CrossRef]
- Guerra, F.P.; Wegrzyn, J.L.; Sykes, R.; Davis, M.F.; Stanton, B.J.; Neale, D.B. Association genetics of chemical wood properties in black poplar (Populus nigra). New Phytol. 2013, 197, 162–176. [Google Scholar] [CrossRef]
- Porth, I.; Klapšte, J.; Skyba, O.; Hannemann, J.; Mckown, A.D.; Guy, R.D.; Difazio, S.P.; Muchero, W.; Ranjan, P.; Tuskan, G.A.; et al. Genome-wide association mapping for wood characteristics in Populus identifies an array of candidate single nucleotide polymorphisms. New Phytol. 2013, 200, 710–726. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Gong, C.; Wang, Q.; Zhou, D.; Yang, H.; Pan, W.; Li, B.; Zhang, D. Genetic architecture of growth traits in Populus revealed by integrated quantitative trait locus (QTL) analysis and association studies. New Phytol. 2016, 209, 1067–1082. [Google Scholar] [CrossRef] [PubMed]
- Hallingbäck, H.R.; Fogelqvist, J.; Powers, S.J.; Turrion-Gomez, J.; Rossiter, R.; Amey, J.; Martin, T.; Weih, M.; Gyllenstrand, N.; Karp, A.; et al. Association mapping in Salix viminalis L. (Salicaceae)–identification of candidate genes associated with growth and phenology. Gcb Bioenergy 2016, 8, 670–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baison, J.; Vidalis, A.; Zhou, L.; Chen, Z.Q.; Li, Z.; Sillanpää, M.J.; Bernhardsson, C.; Scofield, D.; Forsberg, N.; Grahn, T.; et al. Genome-wide association study identified novel candidate loci affecting wood formation in Norway spruce. Plant J. 2019, 100, 83–100. [Google Scholar] [CrossRef]
- Thumma, B.R.; Baltunis, B.S.; Bell, J.C.; Emebiri, L.C.; Moran, G.F.; Southerton, S.G. Quantitative trait locus (QTL) analysis of growth and vegetative propagation traits in Eucalyptus nitens full-sib families. Tree Genet. Genomes 2010, 6, 877–889. [Google Scholar] [CrossRef]
- Thumma, B.R.; Nolan, M.F.; Evans, R.; Moran, G.F. Polymorphisms in cinnamoyl CoA reductase (CCR) are associated with variation in microfibril angle in Eucalyptus spp. Genetics 2005, 171, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Thavamanikumar, S.; Southerton, S.G.; Bossinger, G.; Thumma, B.R. Dissection of complex traits in forest trees—Opportunities for marker-assisted selection. Tree Genet. Genomes 2013, 9, 627–639. [Google Scholar] [CrossRef]
- Zhang, M.; Zhou, C.; Song, Z.; Weng, Q.; Li, M.; Ji, H.; Mo, X.; Huang, H.; Lu, W.; Luo, J.; et al. The first identification of genomic loci in plants associated with resistance to galling insects: A case study in Eucalyptus L’Hér. (Myrtaceae). Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Müller, B.S.; De Almeida Filho, J.E.; Lima, B.M.; Garcia, C.C.; Missiaggia, A.; Aguiar, A.M.; Takahashi, E.; Kirst, M.; Gezan, S.A.; Silva-Junior, O.B.; et al. Independent and Joint-GWAS for growth traits in Eucalyptus by assembling genome-wide data for 3373 individuals across four breeding populations. New Phytol. 2019, 221, 818–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappa, E.P.; El-Kassaby, Y.A.; Garcia, M.N.; Acuña, C.; Borralho, N.M.; Grattapaglia, D.; Poltri, S.N.M. Impacts of population structure and analytical models in genome-wide association studies of complex traits in forest trees: A case study in Eucalyptus globulus. PLoS ONE 2013, 8, e81267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thavamanikumar, S.; McManus, L.J.; Ades, P.K.; Bossinger, G.; Stackpole, D.J.; Kerr, R.; Hadjigol, S.; Freeman, J.S.; Vaillancourt, R.E.; Zhu, P.; et al. Association mapping for wood quality and growth traits in Eucalyptus globulus ssp. globulus Labill identifies nine stable marker-trait associations for seven traits. Tree Genet. Genomes 2014, 10, 1661–1678. [Google Scholar] [CrossRef]
- Sul, J.H.; Martin, L.S.; Eskin, E. Population structure in genetic studies: Confounding factors and mixed models. PLoS Genet. 2018, 14, e1007309. [Google Scholar] [CrossRef]
- Vargas-Reeve, F.; Mora, F.; Perret, S.; Scapim, C.A. Heritability of stem straightness and genetic correlations in Eucalyptus cladocalyx in the semi-arid region of Chile. Crop Breed. Appl. Biotechnol. 2013, 13, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Bush, D.; McCarthy, K.; Meder, R. Genetic variation of natural durability traits in Eucalyptus cladocalyx (sugar gum). Ann. For. Sci. 2011, 68, 1057–1066. [Google Scholar] [CrossRef]
- Pittermann, J.; Sperry, J.S.; Wheeler, J.K.; Hacke, U.G.; Sikkema, E.H. Mechanical reinforcement of tracheids compromises the hydraulic efficiency of conifer xylem. Plant Cell Environ. 2006, 29, 1618–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocozza, C.; Giovannelli, A.; Traversi, M.L.; Castro, G.; Cherubini, P.; Tognetti, R. Do tree-ring traits reflect different water deficit responses in young poplar clones (Populus × canadensis Mönch I-214 and P. deltoides Dvina)? Trees 2011, 25, 975–985. [Google Scholar] [CrossRef]
- Hietz, P.; Rosner, S.; Hietz-Seifert, U.; Wright, S.J. Wood traits related to size and life history of trees in a Panamanian rainforest. New Phytol. 2017, 213, 170–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanne, A.E.; Westoby, M.; Falster, D.S.; Ackerly, D.D.; Loarie, S.R.; Arnold, S.E.J.; Coomes, D.A. Angiosperm wood structure: Global patterns in vessel anatomy and their relation to wood density and potential conductivity. Am. J. Bot. 2010, 97, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stackpole, D.J.; Vaillancourt, R.E.; De Aguigar, M.; Potts, B.M. Age trends in genetic parameters for growth and wood density in Eucalyptus globulus. Tree Genet. Genomes 2010, 6, 179–193. [Google Scholar] [CrossRef]
- McDonald, M.W.; Rawlings, M.; Butcher, P.A.; Bell, J.C. Regional divergence and inbreeding in Eucalyptus cladocalyx (Myrtaceae). Aust. J. Bot. 2003, 51, 393–403. [Google Scholar] [CrossRef]
- Sallam, A.H.; Conley, E.; Prakapenka, D.; Da, Y.; Anderson, J.A. Improving Prediction Accuracy Using Multi-allelic Haplotype Prediction and Training Population Optimization in Wheat. G3 Genes Genomes Genet. 2020, 10, 2265–2273. [Google Scholar] [CrossRef] [PubMed]
- Goudet, J.; Kay, T.; Weir, B.S. How to estimate kinship. Mol. Ecol. 2018, 27, 4121–4135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, N. Structural biology of the major facilitator superfamily transporters. Ann. Rev. Biophys. 2015, 44, 257–283. [Google Scholar] [CrossRef] [PubMed]
- Remy, E.; Cabrito, T.R.; Baster, P.; Batista, R.A.; Teixeira, M.C.; Friml, J.; Sa-Correia, I.; Duque, P. A major facilitator superfamily transporter plays a dual role in polar auxin transport and drought stress tolerance in Arabidopsis. Plant Cell 2013, 25, 901–926. [Google Scholar] [CrossRef] [Green Version]
- Spokevicius, A.V.; Tibbits, J.; Rigault, P.; Nolin, M.A.; Müller, C.; Merchant, A. Medium term water deficit elicits distinct transcriptome responses in Eucalyptus species of contrasting environmental origin. BMC Genom. 2017, 18, 284. [Google Scholar] [CrossRef] [Green Version]
- Toujani, W.; Muñoz-Bertomeu, J.; Flores-Tornero, M.; Rosa-Téllez, S.; Anoman, A.D.; Alseekh, S.; Fernie, A.R.; Ros, R. Functional characterization of the plastidial 3-phosphoglycerate dehydrogenase family in Arabidopsis. Plant Physiol. 2013, 163, 1164–1178. [Google Scholar] [CrossRef] [Green Version]
- Elvira-Matelot, E.; Hachet, M.; Shamandi, N.; Comella, P.; Sáez-Vásquez, J.; Zytnicki, M.; Vaucheret, H. Arabidopsis RNASE THREE LIKE2 modulates the expression of protein-coding genes via 24-nucleotide small interfering RNA-directed DNA methylation. Plant Cell 2016, 28, 406–425. [Google Scholar] [CrossRef]
- Ranf, S.; Gisch, N.; Schäffer, M.; Illig, T.; Westphal, L.; Knirel, Y.A.; Sánchez-Carballo, P.M.; Zähringer, U.; Hückelhoven, R.; Lee, J.; et al. A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat. Immunol. 2015, 16, 426–433. [Google Scholar] [CrossRef]
- Smakowska, E.; Kong, J.; Busch, W.; Belkhadir, Y. Organ-specific regulation of growth-defense tradeoffs by plants. Curr. Opin. Plant Biol. 2016, 29, 129–137. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.-M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, R.T.; Fortes, A.M.; Pinheiro, C.; Pereira, H. Comparison of good-and bad-quality cork: Application of high-throughput sequencing of phellogenic tissue. J. Exp. Bot. 2014, 65, 4887–4905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Cui, D.; Ye, Z.H. Xyloglucan O-acetyltransferases from Arabidopsis thaliana and Populus trichocarpa catalyze acetylation of fucosylated galactose residues on xyloglucan side chains. Planta 2018, 248, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Roje, S. S-Adenosyl-L-methionine: Beyond the universal methyl group donor. Phytochemistry 2006, 67, 1686–1698. [Google Scholar] [CrossRef] [PubMed]
- Ooijen, V.; Lukasik, G.; Van Den Burg, E.; Vossen, H.A.; Cornelissen, J.H.; Takken, B.J. The small heat shock protein 20 RSI2 interacts with and is required for stability and function of tomato resistance protein I-2. Plant J. 2010, 63, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Raes, J.; Rohde, A.; Christensen, J.H.; Van de Peer, Y.; Boerjan, W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.B.; McNear, D.H. Induced transcriptional profiling of phenylpropanoid pathway genes increased flavonoid and lignin content in Arabidopsis leaves in response to microbial products. BMC Plant Biol. 2014, 14, 84. [Google Scholar] [CrossRef] [Green Version]
- Baron, K.N.; Schroeder, D.F.; Stasolla, C. GEm-Related 5 (GER5), an ABA and stress-responsive GRAM domain protein regulating seed development and inflorescence architecture. Plant Sci. 2014, 223, 153–166. [Google Scholar] [CrossRef]
- Hossain, Z.; Pillai, B.V.S.; Gruber, M.Y.; Yu, M.; Amyot, L.; Hannoufa, A. Transcriptome profiling of Brassica napus stem sections in relation to differences in lignin content. BMC Genom. 2018, 19, 255. [Google Scholar] [CrossRef] [Green Version]
- Bonaventure, G.; Salas, J.J.; Pollard, M.R.; Ohlrogge, J.B. Disruption of the FATB gene in Arabidopsis demonstrates an essential role of saturated fatty acids in plant growth. Plant Cell 2003, 15, 1020–1033. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Pérez, A.J.; Venegas-Calerón, M.; Vaistij, F.E.; Salas, J.J.; Larson, T.R.; Garcés, R.; Graham, I.A.; Martínez-Force, E. Reduced expression of FatA thioesterases in Arabidopsis affects the oil content and fatty acid composition of the seeds. Planta 2012, 235, 629–639. [Google Scholar] [CrossRef]
- Brady, S.M.; Zhang, L.; Megraw, M.; Martinez, N.J.; Jiang, E.; Yi, C.S.; Liu, W.; Zeng, A.; Taylor-Teeples, M.; Kim, D.; et al. A stele-enriched gene regulatory network in the Arabidopsis root. Mol. Syst. Biol. 2011, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor-Teeples, M.; Lin, L.; De Lucas, M.; Turco, G.; Toal, T.W.; Gaudinier, A.; Young, N.F.; Trabucco, G.M.; Veling, M.T.; Lamothe, R.; et al. An Arabidopsis gene regulatory network for secondary cell wall synthesis. Nature 2015, 517, 571–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Ye, Z. Secondary Cell Walls: Biosynthesis, Patterned Deposition and Transcriptional Regulation. Plant Cell Physiol. 2015, 56, 195–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Ji, M.; Wen, B.; Liu, L.; Li, S.; Chen, X.; Gao, D.; Li, L. GOLDEN 2-LIKE Transcription Factors of Plants. Front. Plant Sci. 2016, 7, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Sun, Y.; Ahmed, R.I.; Ren, A.; Xie, M. Research Progress on Plant RING-Finger Proteins. Genes 2019, 10, 973. [Google Scholar] [CrossRef] [Green Version]
- Seguel, A.; Jelenska, J.; Herrera-Vásquez, A.; Marr, S.K.; Joyce, M.B.; Gagesch, K.R.; Shakoor, N.; Jiang, S.-C.; Fonseca, A.; Wildermuth, M.C.; et al. PROHIBITIN3 forms complexes with ISOCHORISMATE SYNTHASE1 to regulate stress-induced salicylic acid biosynthesis in Arabidopsis. Plant Physiol. 2018, 176, 2515–2531. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Yang, C.; Zhang, S. The Arabidopsis PHB3 is a pleiotropic regulator for plant development. Plant Signal. Behav. 2019, 14, 1656036. [Google Scholar] [CrossRef]
- Werner, F.A.; Homeier, J. Is tropical montane forest heterogeneity promoted by a resource-driven feedback cycle? Evidence from nutrient relations, herbivory and litter decomposition along a topographical gradient. Funct. Ecol. 2015, 29, 430–440. [Google Scholar] [CrossRef]
- Jullien, D.; Widmann, R.; Loup, C.; Thibaut, B. Relationship between tree morphology and growth stress in mature European beech stands. Ann. For. Sci. 2013, 70, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Ballesta, P.; Bush, D.; Silva, F.F.; Mora, F. Genomic Predictions Using Low-Density SNP Markers, Pedigree and GWAS Information: A Case Study with the Non-Model Species Eucalyptus cladocalyx. Plants 2020, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Desrousseaux, D.; Sandron, F.; Siberchicot, A.; Cierco-Ayrolles, C.; Mangin, B.; Siberchicot, M.A. Package ‘LDcorSV’; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Hill, W.G.; Weir, B.S. Variances and covariances of squared linkage disequilibria in finite populations. Theor. Popul. Biol. 1988, 33, 54–78. [Google Scholar] [CrossRef]
- Breseghello, F.; Sorrells, M.E. Association mapping of kernel size and milling quality in wheat (Triticum aestivum L.) cultivars. Genetics 2006, 172, 1165–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, J.; Schaffner, R.; Nguyen, A.; Moore, H.; Roy, T.; Blumenstiel, S.; Higgins, T.; DeFelice, D.; Lochner, J.; Faggart, L.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Ballesta, P.; Maldonado, C.; Pérez-Rodríguez, P.; Mora, F. SNP and haplotype-based genomic selection of quantitative traits in Eucalyptus globulus. Plants 2019, 8, 331. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Bi, I.V.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Endelman, J.B.; Jannink, J.L. Shrinkage estimation of the realized relationship matrix. G3 Genes Genomes Genet. 2012, 2, 1405–1413. [Google Scholar] [CrossRef]
- Pritchard, M.; Stephens, R.; Donnelly, V. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, C.; Regnaut, R.; Goudet, T. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SNPTEST. Available online: https://mathgen.stats.ox.ac.uk/genetics_software/snptest/snptest.html (accessed on 17 November 2020).
- Maldonado, C.; Mora, F.; Scapim, C.A.; Coan, M. Genome-wide haplotype-based association analysis of key traits of plant lodging and architecture of maize identifies major determinants for leaf angle: Hap LA4. PLoS ONE 2019, 14, e0212925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myburg, A.A.; Grattapaglia, D.; Tuskan, G.A.; Hellsten, U.; Hayes, R.D.; Grimwood, J.; Jenkins, J.; Lindquist, E.; Tice, H.; Bauer, D.; et al. The genome of Eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait a | MTA b | ID c | BF d | PPA e | Candidate Gene | Mutation |
|---|---|---|---|---|---|---|
| HT-DBH | QTL6:3208 | Eucgr.F02605 | 103.44 | 0.35 | E3 ubiquitin-protein ligase RNF13 Protease-associated (PA) RING/U-box zinc finger family protein f | Missense g |
| QTL2:901 | Eucgr.B02601 | 71.65 | 0.27 | Lipoyltransferase 2 (LPT2) | Silent | |
| HT-STR | QTL5:2482 | - | 66.77 | 0.26 | No annotation | - |
| HT-PIL | QTL6:2976 | Eucgr.F01050 | 48.05 | 0.17 | S-locus lectin protein kinase | - |
| DBH-PIL | QTL6:3208 | Eucgr.F02605 | 37.85 | 0.22 | E3 ubiquitin-protein ligase RNF13 Protease-associated (PA) RING/U-box zinc finger family protein f | Missense g |
| DBH-SLD | QTL1:32 | - | 51.89 | 0.19 | No annotation | - |
| QTL10:5177 | Eucgr.J02293 | 44.17 | 0.17 | Prohibitin-3 (PHB3) | - | |
| QTL8:4173 | - | 39.40 | 0.15 | No annotation | - | |
| DBH-BHT | QTL8:4670 | - | 33.40 | 0.22 | No annotation | - |
| DBH-PIL | QTL1:32 | - | 53.37 | 0.42 | No annotation | - |
| QTL6:3208 | Eucgr.F02605 | 140.29 | 0.23 | E3 ubiquitin-protein ligase RNF13 Protease-associated (PA) RING/U-box zinc finger family protein f | Missense g | |
| QTL3:1461 | - | 57.40 | 0.20 | No annotation | - | |
| QTL2:546 | Eucgr.B01596 | 48.73 | 0.20 | Fosfatase Domain | - | |
| QTL6:3236 | Eucgr.F02951 | 46.91 | 0.15 | Ribosome L39 (RPL39) | - |
| Region of Origin/Provenance | N° Families | Latitude | Longitude | Annual Precipitation (mm) |
|---|---|---|---|---|
| Kangaroo Island | ||||
| Flinders Chase | 8 | 35°57′ S | 136°42′ E | 637.9 |
| Eyre Peninsula | ||||
| Cowell | 10 | 33°38′ S | 136°40′ E | 405.0 |
| Marble Range | 4 | 34°30′ S | 135°30′ E | 485.1 |
| Flinders Ranges | ||||
| Mount Remarkable | 16 | 32°43′ S | 138°06′ E | 242.8 |
| Wirrabara | 9 | 33°06′ S | 138°14′ E | 256.6 |
| Chile-Illapel | 2 | 31°40′ S | 71°14′ W | 240.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela, C.E.; Ballesta, P.; Ahmar, S.; Fiaz, S.; Heidari, P.; Maldonado, C.; Mora-Poblete, F. Haplotype- and SNP-Based GWAS for Growth and Wood Quality Traits in Eucalyptus cladocalyx Trees under Arid Conditions. Plants 2021, 10, 148. https://doi.org/10.3390/plants10010148
Valenzuela CE, Ballesta P, Ahmar S, Fiaz S, Heidari P, Maldonado C, Mora-Poblete F. Haplotype- and SNP-Based GWAS for Growth and Wood Quality Traits in Eucalyptus cladocalyx Trees under Arid Conditions. Plants. 2021; 10(1):148. https://doi.org/10.3390/plants10010148
Chicago/Turabian StyleValenzuela, Camilo E., Paulina Ballesta, Sunny Ahmar, Sajid Fiaz, Parviz Heidari, Carlos Maldonado, and Freddy Mora-Poblete. 2021. "Haplotype- and SNP-Based GWAS for Growth and Wood Quality Traits in Eucalyptus cladocalyx Trees under Arid Conditions" Plants 10, no. 1: 148. https://doi.org/10.3390/plants10010148
APA StyleValenzuela, C. E., Ballesta, P., Ahmar, S., Fiaz, S., Heidari, P., Maldonado, C., & Mora-Poblete, F. (2021). Haplotype- and SNP-Based GWAS for Growth and Wood Quality Traits in Eucalyptus cladocalyx Trees under Arid Conditions. Plants, 10(1), 148. https://doi.org/10.3390/plants10010148