
Cornus mas and Cornus officinalis—A Comparison of Antioxidant and Immunomodulatory Activities of Standardized Fruit Extracts in Human Neutrophils and Caco-2 Models

 , ,
, ,
Abstract
:1. Introduction
2. Results
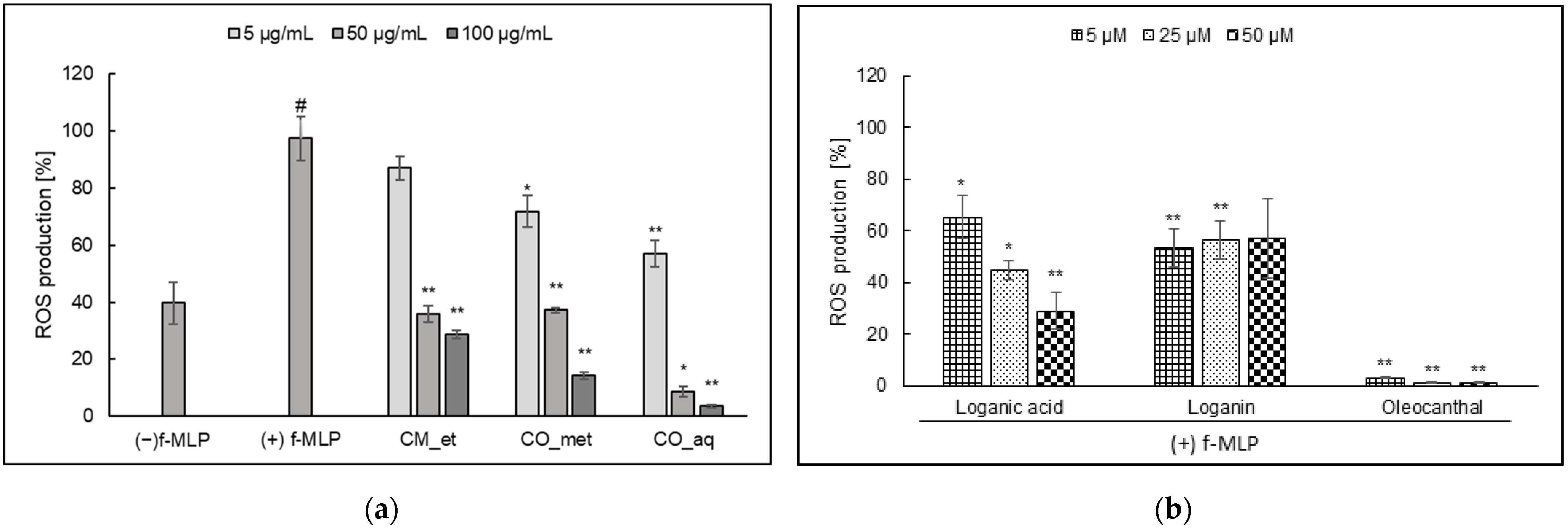
2.1. ROS Generation in PMNs
2.2. Cytokines Secretion by PMNs
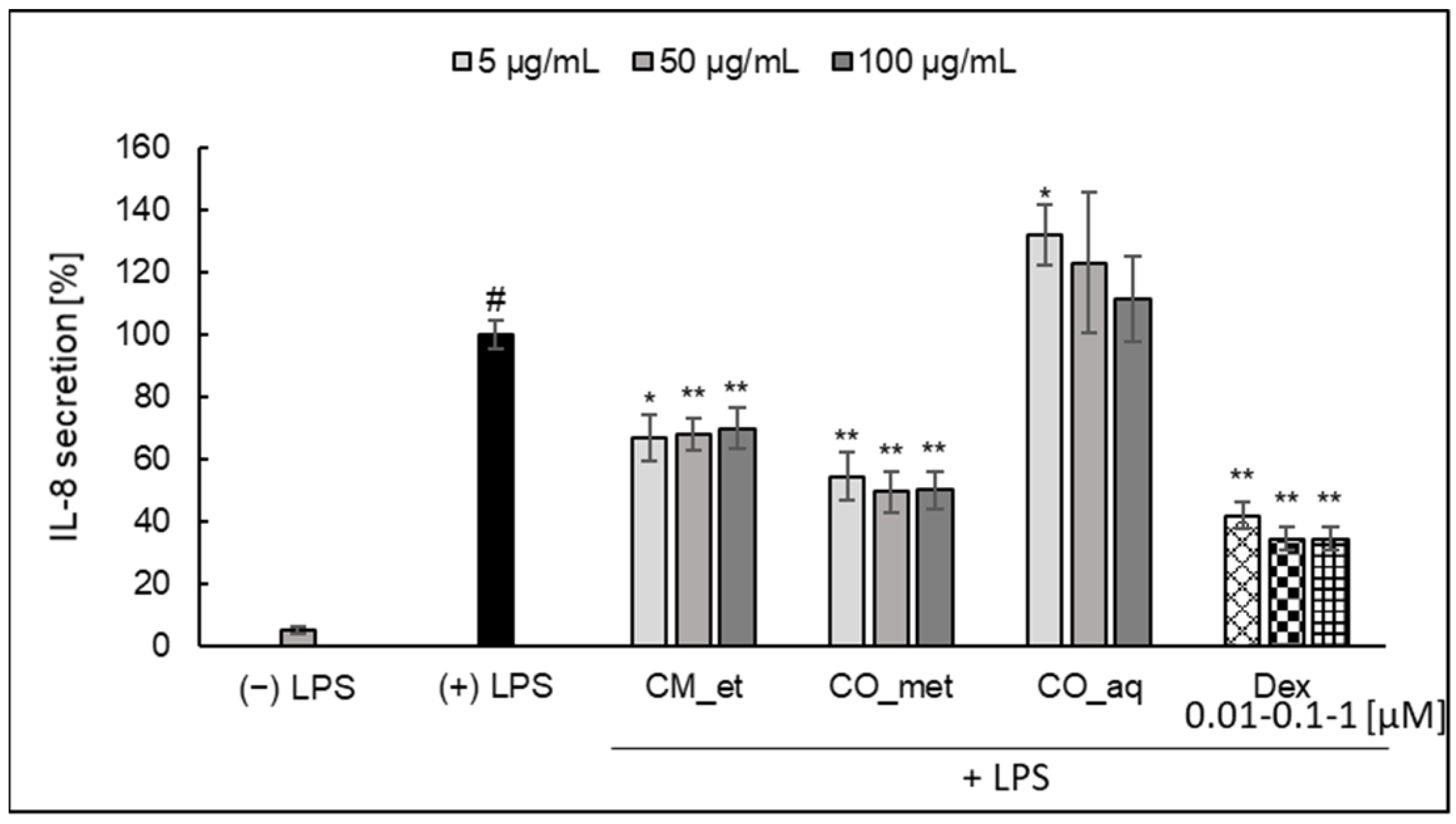
2.2.1. IL-8
2.2.2. TNF-α
2.2.3. IL-1β
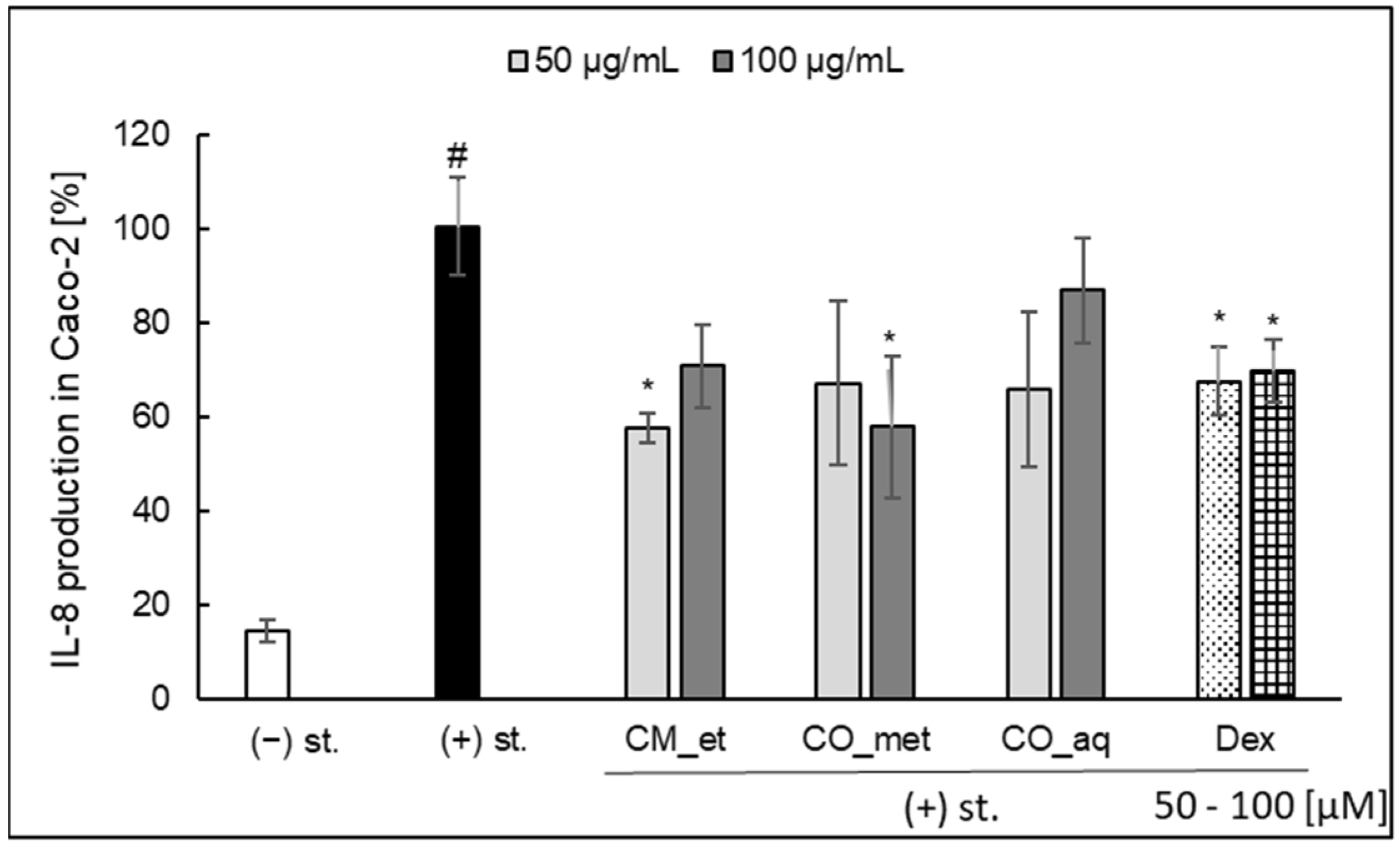
2.3. IL-8 Secretion in Caco-2 Cell Line
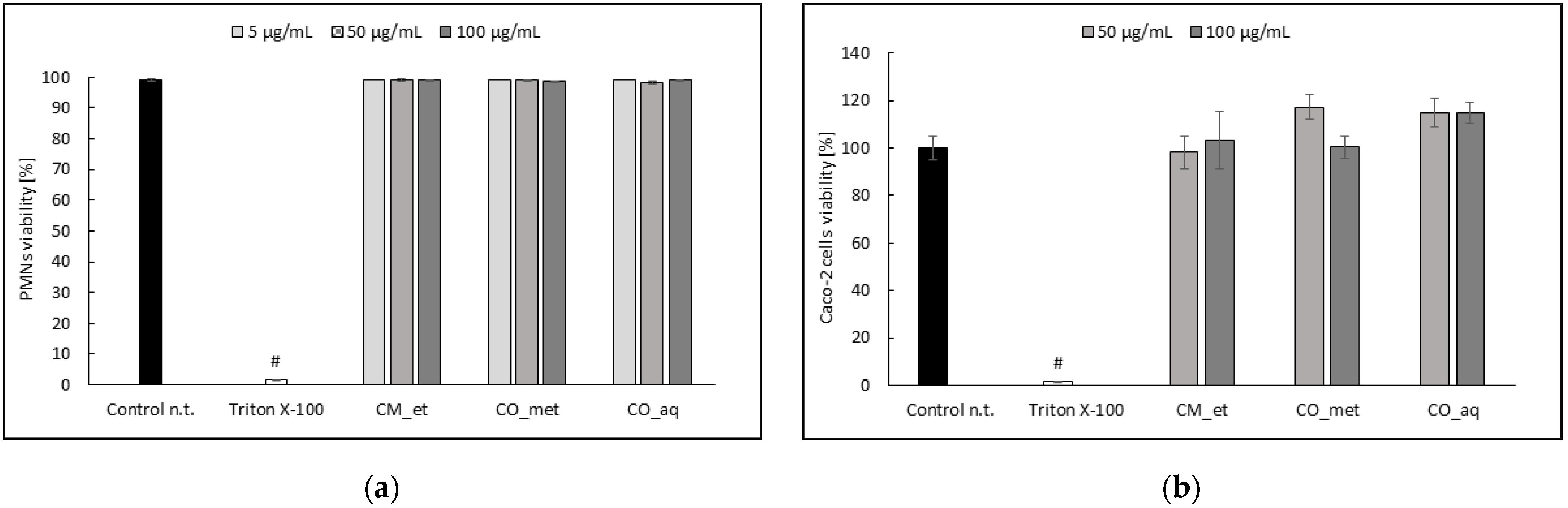
2.4. PMNs and Caco-2 Cells Viability after Cornus Extracts Treatment
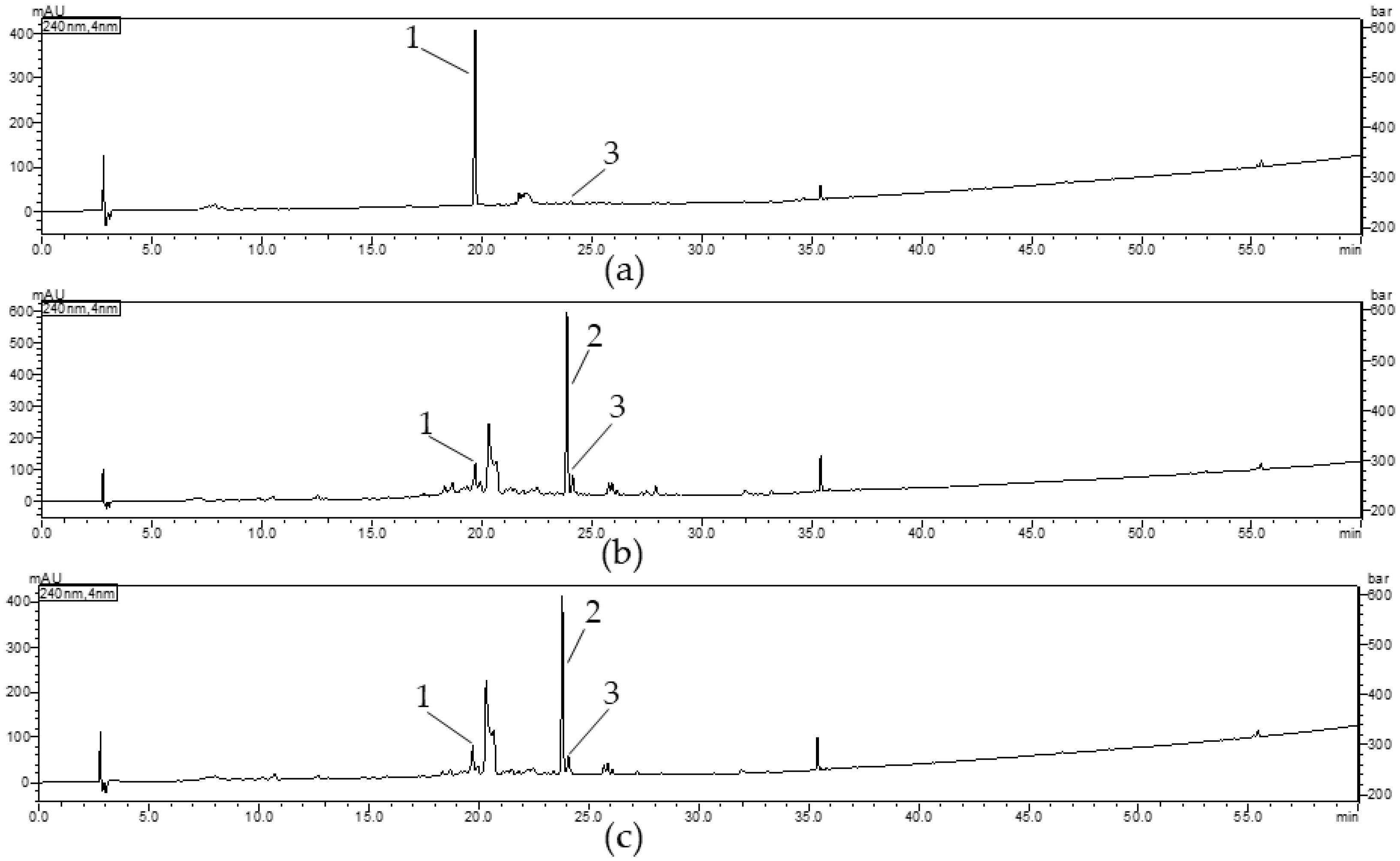
2.5. Qualitative Analysis of Cornus Extracts
2.5.1. Results of Validation Procedures
2.5.2. Quantification of Iridoids in Cornus Extracts
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Extracts Preparations
4.2. General Experimental Conditions in PMNs Culture
4.3. General Experimental Conditions of Caco-2 Cell Culture
4.4. Chromatography Analysis
4.5. Method Validation
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Peschel, W. The Traditional Herbal Medicine Directive within the European regulatory framework for herbal products. Bol. Latinoam. Caribe Plantas Med. Aromáticas 2007, 6, 102–111. [Google Scholar]
- Shang, A.; Huwiler, K.; Nartey, L.; Juni, P.; Egger, M. Placebo-controlled trials of Chinese herbal medicine and conventional medicine comparative study. Int. J. Epidemiol. 2007, 36, 1086–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witkamp, R.F.; Norren, K. Let thy food be thy medicine…. when possible. Eur. J. Pharmacol. 2018, 836, 102–114. [Google Scholar] [CrossRef]
- Guideline on Declaration of Herbal Substances and Herbal Preparations in Herbal Medicinal Products/Traditional Herbal Medicinal Products; Committee on Herbal Medicinal Products (HMPC); European Medicines Agency: London, UK, 2009.
- Czerwińska, M.E.; Melzig, M.F. Cornus mas and Cornus officinalis-analogies and differences of two medicinal plants traditionally used. Front. Pharmacol. 2018, 9, 894. [Google Scholar] [CrossRef]
- Dinda, B.; Kyriakopoulos, A.M.; Dinda, S.; Zoumpourlis, V.; Thomaidis, N.S.; Velegraki, A.; Markopoulos, C.; Dinda, M. Cornus mas L. (cornelian cherry), an important European and Asian traditional food and medicine: Ethnomedicine, phytochemistry and pharmacology for its commercial utilization in drug industry. J. Ethnopharmacol. 2016, 193, 670–690. [Google Scholar] [CrossRef] [PubMed]
- Kucharska, A.Z.; Szumny, A.; Sokół-Łętowska, A.; Piórecki, N.; Klymenko, S.V. Iridoids and anthocyanins in cornelian cherry (Cornus mas L.) cultivars. J. Food Comp. Anal. 2015, 40, 95–102. [Google Scholar] [CrossRef]
- Seeram, N.P.; Schutzki, R.; Chandra, A.; Nair, M.G. Characterization, quantification, and bioactivities of anthocyanins in Cornus species. J. Agric. Food Chem. 2002, 50, 2519–2523. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, Y.; Dong, L.; Gao, Q.; Yin, L.; Quan, H.; Chen, R.; Fu, X.; Lin, D. Ethnopharmacology, phytochemistry, and pharmacology of Cornus officinalis Sieb. et Zucc. J. Ethnopharmacol. 2018, 213, 280–301. [Google Scholar] [CrossRef]
- Zhao, M.; Qian, D.; Shang, E.X.; Jiang, S.; Guo, J.; Liu, P.; Su, S.L.; Duan, J.A.; Du, L.; Tao, J. Comparative pharmacokinetics of the main compounds of Shanzhuyu extract after oral administration in normal and chronic kidney disease rats. J. Ethnopharmacol. 2015, 173, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Li, Q.; Gao, Z.; Wang, L. Antidiabetic effects of Corni Fructus extract in streptozotocin-induced diabetic rats. Yonsei Med. J. 2012, 53, 691–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.H.; Liu, H.K.; Huang, W.J.; Huang, C.C.; Wu, T.H.; Hsu, F.L. Evaluation of the potential hypoglycemic and Beta-cell protective constituents isolated from Corni Fructus to tackle insulin-dependent diabetes mellitus. J. Agric. Food Chem. 2011, 59, 7743–7751. [Google Scholar] [CrossRef]
- Dai, B.; Wu, Q.; Zeng, C.; Zhang, J.; Cao, L.; Xiao, Z.; Yang, M. The effect of Liuwei Dihuang decoction on PI3K/Akt signaling pathway in liver of type 2 diabetes mellitus (T2DM) rats with insulin resistance. J. Ethnopharmacol. 2016, 192, 382–389. [Google Scholar] [CrossRef]
- Han, Y.H.; Jung, H.W.; Park, Y.-K. Selective therapeutic effect of Cornus officinalis fruits on the damage of different organs in STZ-induced diabetic rats. Am. J. Chin. Med. 2014, 42, 1169–1182. [Google Scholar] [CrossRef]
- Ma, W.; Wang, K.J.; Cheng, C.S.; Yan, G.Q.; Lu, W.L.; Ge, J.F.; Cheng, Y.X.; Li, N. Bioactive compounds from Cornus officinalis fruits and their effects on diabetic nephropathy. J. Ethnopharmacol. 2014, 153, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Kim, Y.-C. Antidiabetic and renoprotective effects of Corni Fructus extract in db/db mice. Mol. Cell. Toxicol. 2010, 6, 135–142. [Google Scholar] [CrossRef]
- Liu, H.; Xu, H.; Shen, C.; Wu, C. Effect of the best compatibility of components in Corni Fructus on WT1 expression in glomerular podocytes of type 2 diabetic rats with early nephropathy. Am. J. Chin. Med. 2012, 40, 537–549. [Google Scholar] [CrossRef]
- Liu, J.P.; Feng, L.; Zhang, M.H.; Ma, D.Y.; Wang, S.Y.; Gu, J.; Fu, Q.; Qu, R.; Ma, S.P. Neuroprotective effect of Liuwei Dihuang decoction on cognition deficits of diabetic encephalopathy in streptozotocin-induced diabetic rat. J. Ethnopharmacol. 2013, 150, 371–381. [Google Scholar] [CrossRef]
- Park, C.H.; Noh, J.S.; Tanaka, T.; Yokozawa, T. Effects of morroniside isolated from Corni Fructus on renal lipids and inflammation in type 2 diabetic mice. J. Pharm. Pharmacol. 2010, 62, 374–380. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Song, S.; Zou, Z.; Feng, M.; Wang, D.; Wang, Y.; Li, X.; Ye, X. The hypoglycemic and synergistic effect of loganin, morroniside, and ursolic acid isolated from the fruits of Cornus officinalis. Phytother. Res. 2016, 30, 283–291. [Google Scholar] [CrossRef]
- Yokozawa, T.; Kang, K.S.; Park, C.H.; Noh, J.S.; Yamabe, N.; Shibahara, N.; Tanaka, T. Bioactive constituents of Corni Fructus: The therapeutic use of morroniside, loganin, and 7-O-galloyl-D-sedoheptulose as renoprotective agents in type 2 diabetes. Drug Discov. Ther. 2010, 4, 223–234. [Google Scholar]
- Park, C.H.; Noh, J.S.; Tanaka, T.; Roh, S.S.; Lee, J.C.; Yokozawa, T. Polyphenol isolated from Corni Fructus, 7-O-galloyl-D-sedoheptulose, modulates advanced glycation endproduct-related pathway in type 2 diabetic db/db mice. Arch. Pharm. Res. 2015, 38, 1270–1280. [Google Scholar] [CrossRef] [PubMed]
- Popović, B.M.; Štajner, D.; Slavko, K.; Sandra, B. Antioxidant capacity of cornelian cherry (Cornus mas L.)—comparison between permanganate reducing antioxidant capacity and other antioxidant methods. Food Chem. 2012, 134, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Pyrkosz-Biardzka, K.; Kucharska, A.Z.; Sokół-Łętowska, A.; Strugała, P.; Gabrielska, J. A comprehensive study on antioxidant properties of crude extracts from fruits of Berberis vulgaris L., Cornus mas L. and Mahonia aquifolium Nutt. Pol. J. Food Nutr. Sci. 2014, 64, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Ersoy, N.; Bagci, Y.; Gok, V. Antioxidant properties of 12 cornelian cherry fruit types (Cornus mas L.) selected from Turkey. Sci. Res. Essays 2011, 6, 98–102. [Google Scholar] [CrossRef]
- Celep, E.; Aydin, A.; Yesilada, E. A comparative study on the in vitro antioxidant potentials of three edible fruits: Cornelian cherry, Japanese persimmon and cherry laurel. Food Chem. Toxicol. 2012, 50, 3329–3335. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-O.; Kim, S.Y.; Han, S.-M.; Kim, H.-M.; Ham, S.-S.; Kang, I.-J. Corni fructus scavenges hydroxy radicals and decreases oxidative stress in endothelial cells. J. Med. Food 2006, 9, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The role of inflammation in diabetes: Current concepts and future perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Cheng, W.; Zhang, G.; Ma, Q.; Li, X.; Zhang, B.; Hu, T.; Song, G. Protective effects of iridoid glycosides on acute colitis via inhibition of the inflammatory response mediated by the STAT3/NF-кB, CyrillicB pathway. Int. Immunopharmacol. 2020, 81, 106240. [Google Scholar] [CrossRef] [PubMed]
- Świerczewska, A.; Buchholz, T.; Melzig, M.F.; Czerwinska, M.E. In vitro α-amylase and pancreatic lipase inhibitory activity of Cornus mas L. and Cornus alba L. fruit extracts. J. Food Drug Anal. 2019, 27, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreck, K.; Melzig, M.F. Traditionally used plants in the treatment of diabetes mellitus: Screening for uptake inhibition of glucose and fructose in the Caco2-cell model. Front. Pharmacol. 2021, 12, 692566. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.Y.; Manchester, S.R.; Thomas, D.T.; Zhang, W.; Fan, C. Phylogeny, biogeography, and molecular dating of cornelian cherries (Cornus, Cornaceae): Tracking tertiary plant migration. Evolution 2005, 59, 1685–1700. [Google Scholar] [CrossRef] [PubMed]
- Rudrapaul, P.; Kyriakopoulos, A.M.; De, U.C.; Zoumpourlis, V.; Dinda, B. New flavonoids from the fruits of Cornus mas, Cornaceae. Phytochem. Lett. 2015, 11, 292–295. [Google Scholar] [CrossRef]
- Pawlowska, A.M.; Camangi, F.; Braca, A. Quali-quantitative analysis of flavonoids of Cornus mas L. (Cornaceae) fruits. Food Chem. 2010, 119, 1257–1261. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.-Y.; Wang, R.; Shi, Y.-P. Chemical constituents from the fruits of Cornus officinalis. Biochem. Syst. Ecol. 2012, 45, 120–123. [Google Scholar] [CrossRef]
- Cao, G.; Zhang, C.; Zhang, Y.; Cong, X.; Cai, H.; Cai, B. Screening and identification of potential active components in crude Fructus Croni using solid-phase extraction and LC-LTQ-linear ion trap mass spectrometry. Pharm. Biol. 2012, 50, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Zhang, Y.; Feng, J.; Cai, H.; Zhang, C.; Ding, M.; Cong, X.; Cai, B. A rapid and sensitive assay for determining the main components in processed Fructus Corni by UPLC-Q-TOF-MS. Chromatographia 2011, 73, 135–141. [Google Scholar] [CrossRef]
- Gąstoł, M.; Krośniak, M.; Derwisz, M.; Dobrowolska-Iwanek, J. Cornelian cherry (Cornus mas L.) juice as a potential source of biological compounds. J. Med. Food 2013, 16, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Hassanpour, H.; Hamidoghli, Y.; Samizadeh, H. Some fruit characteristics of Iranian cornelian cherries (Cornus mas L.). Not. Bot. Horti. Agrobo. 2012, 40, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012. [Google Scholar] [CrossRef] [Green Version]
- Fournier, B.M.; Parkos, C.A. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012, 5, 354–366. [Google Scholar] [CrossRef]
- Wéra, O.; Lancellotti, P.; Oury, C. The dual role of neutrophils in inflammatory bowel diseases. J. Clin. Med. 2016, 5, 118. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, J.; Liu, Z.; Jiang, A.; Li, S.; Wu, D.; Zhang, Y.; Zhu, X.; Zhou, E.; Wei, Z.; et al. Sodium butyrate alleviates lipopolysaccharide-induced inflammatory responses by down-regulation of NF-kappaB, NLRP3 signaling pathway, and activating histone acetylation in bovine macrophages. Front. Vet. Sci. 2020, 7, 579674. [Google Scholar] [CrossRef]
- Russo, I.; Luciani, A.; De Cicco, P.; Troncone, E.; Ciacci, C. Lipopolysaccharide-induced inflammation in intestinal cells and Crohn’s mucosa through modulation of antioxidant defense machinery. PLoS ONE 2012, 7, e32841. [Google Scholar] [CrossRef] [Green Version]
- Bates, J.M.; Akerlund, J.; Mittge, E.; Guillemin, K. Intestinal alkaline phosphatase detoxifies lipopolysaccharide and prevents inflammation in zebrafish in response to the gut microbiota. Cell Host Microbe 2007, 2, 371–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.; McGee, D.W. ROCK activity affects IL-1-induced signaling possibly through MKK4 and p38 MAPK in Caco-2 cells. Vitr. Cell. Dev. Biol. Anim. 2016, 52, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Eckmann, L.; Jung, H.C.; Schurer-Maly, C.; Panja, A.; Morzycka-Wroblewska, E.; Kagnoff, M.F. Differential cytokine expression by human intestinal epithelial cell lines: Regulated expression of interleukin 8. Gastroenterology 1993, 105, 1689–1697. [Google Scholar] [CrossRef]
- Van De Walle, J.; Hendrickx, A.; Romier, B.; Larondelle, Y.; Schneider, Y.J. Inflammatory parameters in Caco-2 cells: Effect of stimuli nature, concentration, combination and cell differentiation. Toxicol. Vitr. 2010, 24, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, B.; Filip, A.; Clichici, S.; Suharoschi, R.; Bolfa, P.; David, L. Antioxidant activity of Cornelian cherry (Cornus mas L.) fruits extract and the in vivo evaluation of its anti-inflammatory effects. J. Funct. Food 2016, 26, 77–87. [Google Scholar] [CrossRef]
- Yin, X.; You, Q.; Jiang, Z.; Zhou, X. Optimization for ultrasonic-microwave synergistic extraction of polysaccharides from Cornus officinalis and characterization of polysaccharides. Int. J. Biol. Macromol. 2016, 83, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, X.; Shen, B.; Kang, L.; Fan, E. Extraction, structure and bioactivities of the polysaccharides from Fructus corni. Recent Pat. Food Nutr. Agric. 2013, 5, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Z.; Huang, L. Isolation and structural characterization of a polysaccharide FCAP1 from the fruit of Cornus officinalis. Carbohydr. Res. 2010, 345, 1909–1913. [Google Scholar] [CrossRef] [PubMed]
- Bijelić, S.M.; Gološin, B.R.; Ninić Todorović, J.I.; Cerović, S.B.; Popović, B.M. Physicochemical fruit characteristics of cornelian cherry (Cornus mas L.) genotypes from Serbia. HortScience 2011, 46, 849–853. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Liu, Q.; Fu, Y.; Luo, X.; Hu, M.; Ma, F.; Wang, Q.; Lai, X.; Zhou, L. Effects of Lycium barbarum polysaccharides with different molecular weights on function of RAW264.7 macrophages. Food Agric. Immunol. 2018, 29, 808–820. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Xiao, X.; Liu, J.; Wang, J.; Zhang, N.; Bing, T.; Liu, X.; Zhang, Z.; Shangguan, D. Immunomodulatory effects of Lycium barbarum polysaccharide extract and its uptake behaviors at the cellular level. Molecules 2020, 25, 1351. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.E.; Jeong, J.J.; Hyam, S.R.; Han, M.J.; Kim, D.H. Ursolic acid isolated from the seed of Cornus officinalis ameliorates colitis in mice by inhibiting the binding of lipopolysaccharide to Toll-like receptor 4 on macrophages. J. Agric. Food Chem. 2014, 62, 9711–9721. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Cha, H.-J.; Lee, H.; Kim, G.-Y.; Choi, Y.-H. The regulation of the TLR4/NF-κB and Nrf2/HO-1 signaling pathways is involved in the inhibition of lipopolysaccharide-induced inflammation and oxidative reactions by morroniside in RAW 264.7 macrophages. Arch. Biochem. Biophys. 2021, 706, 108926. [Google Scholar] [CrossRef]
- Choi, Y.H.; Jin, G.Y.; Li, G.Z.; Yan, G.H. Cornuside suppresses lipopolysaccharide-induced inflammatory mediators by inhibiting nuclear factor-kappa B activation in RAW 264.7 macrophages. Biol. Pharm. Bull. 2011, 34, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Dzydzan, O.; Brodyak, I.; Sokol-Letowska, A.; Kucharska, A.Z.; Sybirna, N. Loganic Acid, an iridoid glycoside extracted from Cornus mas L. fruits, reduces of carbonyl/oxidative stress biomarkers in plasma and restores antioxidant balance in leukocytes of rats with streptozotocin-induced diabetes mellitus. Life 2020, 10, 349. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Suzuki, K.; Okutsu, M.; Yamazaki, K.; Shinkai, S. The association of elevated reactive oxygen species levels from neutrophils with low-grade inflammation in the elderly. Immun. Ageing 2008, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [Green Version]
- Li, H.Q.; Wei, J.J.; Xia, W.; Li, J.H.; Liu, A.J.; Yin, S.B.; Wang, C.; Song, L.; Wang, Y.; Zheng, G.Q.; et al. Promoting blood circulation for removing blood stasis therapy for acute intracerebral hemorrhage: A systematic review and meta-analysis. Acta Pharmacol. Sin. 2015, 36, 659–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, K.A.; Hwang, Y.J.; Song, J. Antioxidant activities and oxidative stress inhibitory effects of ethanol extracts from Cornus officinalis on RAW 264.7 cells. BMC Complement. Altern. Med. 2016, 16, 196. [Google Scholar] [CrossRef] [Green Version]
- Sengul, M.; Eser, Z.; Ercisli, S. Chemical properties and antioxidant capacity of cornelian cherry genotypes grown in Coruh valley of Turkey. Acta Sci. Pol. Hortorum Cultus 2014, 13, 73–82. [Google Scholar]
- Ahn, J.H.; Mo, E.J.; Jo, Y.H.; Kim, S.B.; Hwang, B.Y.; Lee, M.K. Variation of loganin content in Cornus officinalis fruits at different extraction conditions and maturation stages. Biosci. Biotechnol. Biochem. 2017, 81, 1973–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzamaloukas, A.H.; Ing, T.S.; Siamopoulos, K.C.; Raj, D.S.; Elisaf, M.S.; Rohrscheib, M.; Murata, G.H. Pathophysiology and management of fluid and electrolyte disturbances in patients on chronic dialysis with severe hyperglycemia. Semin. Dial. 2008, 21, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, A.; Świderski, F.; Laskowski, W. Osmolality of components and their Application in the design of functional recovery drinks. Appl. Sci. 2020, 10, 7663. [Google Scholar] [CrossRef]
- Asgary, S.; Kelishadi, R.; Rafieian-Kopaei, M.; Najafi, S.; Najafi, M.; Sahebkar, A. Investigation of the lipid-modifying and antiinflammatory effects of Cornus mas L. supplementation on dyslipidemic children and adolescents. Pediatr. Cardiol. 2013, 34, 1729–1735. [Google Scholar] [CrossRef]
- Asgary, S.; Rafieian-Kopaei, M.; Shamsi, F.; Najafi, S.; Sahebkar, A. Biochemical and histopathological study of the anti-hyperglycemic and anti-hyperlipidemic effects of cornelian cherry (Cornus mas L.) in alloxan-induced diabetic rats. J. Complement. Integr. Med. 2014, 11, 63–69. [Google Scholar] [CrossRef]
- Sozanski, T.; Kucharska, A.Z.; Szumny, A.; Magdalan, J.; Bielska, K.; Merwid-Lad, A.; Wozniak, A.; Dzimira, S.; Piorecki, N.; Trocha, M. The protective effect of the Cornus mas fruits (cornelian cherry) on hypertriglyceridemia and atherosclerosis through PPARalpha activation in hypercholesterolemic rabbits. Phytomedicine Int. J. Phytother. Phytopharm. 2014, 21, 1774–1784. [Google Scholar] [CrossRef]
- Rutkowski, L. Klucz do Oznaczania Roślin Naczyniowych Polski Niżowej, 2nd ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2011. [Google Scholar]
- Buchholz, T.; Melzig, M.F. Medicinal plants traditionally used for treatment of obesity and diabetes mellitus—Screening for pancreatic lipase and α-amylase inhibition. Phytother. Res. 2016, 30, 260–266. [Google Scholar] [CrossRef]
- Boyum, A. A one-stage procedure for isolation of granulocytes and lymphocytes from human blood. General sedimentation properties of white blood cells in a 1g gravity field. Scand. J. Clin. Lab. Investig. Suppl. 1968, 97, 51–76. [Google Scholar]
- Czerwińska, M.E.; Świerczewska, A.; Granica, S. Bioactive constituents of Lamium album L. as inhibitors of cytokines secretion in human neutrophils. Molecules 2018, 23, 2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerwińska, M.; Kiss, A.K.; Naruszewicz, M. A comparison of antioxidant activities of oleuropein and its dialdehydic derivative from olive oil, oleacein. Food Chem. 2012, 131, 940–947. [Google Scholar] [CrossRef]
- Orchel, A.; Dzierzewicz, Z.; Parfiniewicz, B.; Weglarz, L.; Wilczok, T. Butyrate-induced differentiation of colon cancer cells is PKC and JNK dependent. Dig. Dis. Sci. 2005, 50, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Olejnik, A.; Lewandowska, M.; Grajek, W.; Czaczyk, K. New rapid method of Caco-2 cell differentiation. Pol. J. Food Nutr. Sci. 2003, 12, 60–64. [Google Scholar]
- Faqar-Uz-Zaman, W.F.; Schmidt, K.G.; Thomas, D.; Pfeilschifter, J.M.; Radeke, H.H.; Schwiebs, A. S1P lyase siRNA dampens malignancy of DLD-1 colorectal cancer cells. Lipids 2021, 56, 155–166. [Google Scholar] [CrossRef]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- ICH. Harmonised Tripartite Guideline. Q2(R1) Validation of analytical procedures: Text and methodology. In International Conference on Harmonization of Technical Requirements for Registration of Pharmaceuticals for Human Use; ICH: Geneva, Switzerland, 2005. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Regression Equation | r | R2 | Test F | LOQ [µg/mL] | LOD [µg/mL] |
|---|---|---|---|---|---|---|
| Loganic acid (1) | y = 1.6791 × 107 x − 2.0096 × 105 | 0.9999 | 0.9997 | 42,771.41 | 45.92 | 15.15 |
| Loganin (2) | y = 1.6791 × 107 x − 2.0096 × 105 | 0.9999 | 0.9997 | 46,182.75 | 62.59 | 20.65 |
| Sweroside (3) | y = 1.4243 × 107 x − 50,927.8208 | 0.9852 | 0.9707 | 364.37 | 17.06 | 5.63 |
| Extract | Loganic Acid (1) | Loganin (2) | Sweroside (3) |
|---|---|---|---|
| CM_et | 16.97 ± 0.10 | n.d. | 0.65 ± 0.02 |
| CO_met | 3.27 ± 0.04 | 11.38 ± 0.08 | 1.77 ± 0.03 |
| CO_aq | 3.40 ± 0.06 | 10.55 ± 0.13 | 1.72 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czerwińska, M.E.; Bobińska, A.; Cichocka, K.; Buchholz, T.; Woliński, K.; Melzig, M.F. Cornus mas and Cornus officinalis—A Comparison of Antioxidant and Immunomodulatory Activities of Standardized Fruit Extracts in Human Neutrophils and Caco-2 Models. Plants 2021, 10, 2347. https://doi.org/10.3390/plants10112347
Czerwińska ME, Bobińska A, Cichocka K, Buchholz T, Woliński K, Melzig MF. Cornus mas and Cornus officinalis—A Comparison of Antioxidant and Immunomodulatory Activities of Standardized Fruit Extracts in Human Neutrophils and Caco-2 Models. Plants. 2021; 10(11):2347. https://doi.org/10.3390/plants10112347
Chicago/Turabian StyleCzerwińska, Monika E., Agata Bobińska, Katarzyna Cichocka, Tina Buchholz, Konrad Woliński, and Matthias F. Melzig. 2021. "Cornus mas and Cornus officinalis—A Comparison of Antioxidant and Immunomodulatory Activities of Standardized Fruit Extracts in Human Neutrophils and Caco-2 Models" Plants 10, no. 11: 2347. https://doi.org/10.3390/plants10112347