
A Standardized Lindera obtusiloba Extract Improves Endothelial Dysfunction and Attenuates Plaque Development in Hyperlipidemic ApoE-Knockout Mice

 , , , and
, , , and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
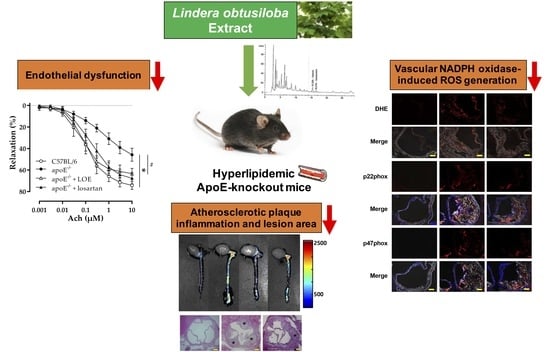
2.1. LOE Improves Endothelial Dysfunction in WD-Fed apoE−/− Mice
2.2. LOE Decreases Excessive Vascular ROS Formation by Inhibiting NADPH Oxidase Subunits in WD-Fed apoE−/− Mice
2.3. LOE Suppresses Inflammation in Murine Aortic Atherosclerosis
2.4. LOE Reduces Atherosclerotic Plaque Burden in apoE−/− Mice in WD-Fed apoE−/− Mice
3. Discussion
4. Materials and Methods
4.1. Plant Extraction and Standardization
4.2. Animals
4.3. Vascular Reactivity
4.4. Determination of Vascular ROS Formation
4.5. Immunohistochemical Analysis of NADPH Oxidase Subunit Expression
4.6. Macroscopic Fluorescence Reflectance Imaging of Plaque Inflammation
4.7. Measurement of H&E Staining and the Aortic Atherosclerotic Plaque Area
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef]
- Quyyumi, A.A. Prognostic value of endothelial function. Am. J. Cardiol. 2003, 91, 19–24. [Google Scholar] [CrossRef]
- Stocker, R.; Keaney, J.F., Jr. Role of oxidative modifications in atherosclerosis. Physiol. Rev. 2004, 84, 1381–1478. [Google Scholar] [CrossRef]
- Miller, F.J.; Gutterman, D.D.; Rios, C.D.; Heistad, D.D.; Davidson, B.L. Superoxide Production in Vascular Smooth Muscle Contributes to Oxidative Stress and Impaired Relaxation in Atherosclerosis. Circ. Res. 1998, 82, 1298–1305. [Google Scholar] [CrossRef]
- Taddei, S.; Virdis, A.; Ghiadoni, L.; Salvetti, G.; Bernini, G.; Magagna, A.; Salvetti, A. Age-Related Reduction of NO Availability and Oxidative Stress in Humans. Hypertension 2001, 38, 274–279. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Tian, R.; Ding, Y.; Peng, Y.Y.; Lu, N. Inhibition of Myeloperoxidase- and Neutrophil-Mediated Hypochlorous Acid Formation in Vitro and Endothelial Cell Injury by (−)-Epigallocatechin Gallate. J. Agric. Food Chem. 2017, 65, 3198–3203. [Google Scholar] [CrossRef]
- Ding, Y.; Tian, R.; Yang, Z.; Chen, J.; Lu, N. NADPH oxidase-dependent degradation of single-walled carbon nanotubes in macrophages. J. Mater. Sci. Mater. Med. 2017, 28, 7. [Google Scholar] [CrossRef]
- Drummond, G.R.; Selemidis, S.; Griendling, K.K.; Sobey, C.G. Combating oxidative stress in vascular disease: NADPH oxidases as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 453–471. [Google Scholar] [CrossRef]
- Lassègue, B.; San Martín, A.; Griendling, K.K. Biochemistry, physiology, and pathophysiology of NADPH oxidases in the cardiovascular system. Circ. Res. 2012, 110, 1364–1390. [Google Scholar] [CrossRef] [PubMed]
- Barry-Lane, P.A.; Patterson, C.; van der Merwe, M.; Hu, Z.; Holland, S.M.; Yeh, E.T.; Runge, M.S. p47phox is required for atherosclerotic lesion progression in ApoE(−/−) mice. J. Clin. Investig. 2001, 108, 1513–1522. [Google Scholar] [CrossRef]
- Judkins, C.P.; Diep, H.; Broughton, B.R.; Mast, A.E.; Hooker, E.U.; Miller, A.A.; Selemidis, S.; Dusting, G.J.; Sobey, C.G.; Drummond, G.R. Direct evidence of a role for Nox2 in superoxide production, reduced nitric oxide bioavailability, and early atherosclerotic plaque formation in ApoE−/− mice. Am. J. Physiol.-Heart Circ. Physiol. 2010, 298, H24–H32. [Google Scholar] [CrossRef]
- Raggi, P.; Genest, J.; Giles, J.T.; Rayner, K.J.; Dwivedi, G.; Beanlands, R.S.; Gupta, M. Role of inflammation in the pathogenesis of atherosclerosis and therapeutic interventions. Atherosclerosis 2018, 276, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Lahera, V.; Goicoechea, M.; de Vinuesa, S.G.; Miana, M.; de las Heras, N.; Cachofeiro, V.; Luño, J. Endothelial dysfunction, oxidative stress and inflammation in atherosclerosis: Beneficial effects of statins. Curr. Med. Chem. 2007, 14, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Kim, P.H.; Lee, W.H.; Hirani, A.A. Interleukin-4, Oxidative Stress, Vascular Inflammation and Atherosclerosis. Biomol. Ther. 2010, 18, 135–144. [Google Scholar] [CrossRef]
- Yook, C. Medical Plants of Korea.; Jinmyeong Publishing Co.: Seoul, Korea, 1989; p. 184. [Google Scholar]
- Freise, C.; Querfeld, U. The lignan (+)-episesamin interferes with TNF-α-induced activation of VSMC via diminished activation of NF-ĸB, ERK1/2 and AKT and decreased activity of gelatinases. Acta Physiol. 2015, 213, 642–652. [Google Scholar] [CrossRef]
- Hong, C.O.; Lee, H.A.; Rhee, C.H.; Choung, S.Y.; Lee, K.W. Separation of the antioxidant compound quercitrin from Lindera obtusiloba Blume and its antimelanogenic effect on B16F10 melanoma cells. Biosci. Biotechnol. Biochem. 2013, 77, 58–64. [Google Scholar] [CrossRef]
- Jung, S.H.; Han, J.H.; Park, H.S.; Lee, J.J.; Yang, S.Y.; Kim, Y.H.; Heo, K.S.; Myung, C.S. Inhibition of Collagen-Induced Platelet Aggregation by the Secobutanolide Secolincomolide A from Lindera obtusiloba Blume. Front. Pharmacol. 2017, 8, 560. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.W.; Ha, J.H.; Shin, H.G.; Jeong, S.H.; Kim, J.H.; Lee, J.; Park, J.Y.; Kwon, H.J.; Jung, K.; Lee, W.S.; et al. Lindera obtusiloba Attenuates Oxidative Stress and Airway Inflammation in a Murine Model of Ovalbumin-Challenged Asthma. Antioxidants 2020, 9, 563. [Google Scholar] [CrossRef]
- Freise, C.; Erben, U.; Neuman, U.; Kim, K.; Zeitz, M.; Somasundaram, R.; Ruehl, M. An active extract of Lindera obtusiloba inhibits adipogenesis via sustained Wnt signaling and exerts anti-inflammatory effects in the 3T3-L1 preadipocytes. J. Nutr. Biochem. 2010, 21, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Oak, M.H.; Jung, S.H.; Park, D.H.; Auger, C.; Kim, K.R.; Lee, S.W.; Schini-Kerth, V.B. An ethanolic extract of Lindera obtusiloba stems causes NO-mediated endothelium-dependent relaxations in rat aortic rings and prevents angiotensin II-induced hypertension and endothelial dysfunction in rats. Naunyn Schmiedebergs Arch. Pharmacol. 2011, 383, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Auger, C.; Park, D.H.; Kang, M.; Oak, M.H.; Kim, K.R.; Schini-Kerth, V.B. An ethanolic extract of Lindera obtusiloba stems, YJP-14, improves endothelial dysfunction, metabolic parameters and physical performance in diabetic db/db mice. PLoS ONE 2013, 8, e65227. [Google Scholar] [CrossRef]
- Buday, A.; Őrsy, P.; Godó, M.; Mózes, M.; Kökény, G.; Lacza, Z.; Koller, Á.; Ungvári, Z.; Gross, M.-L.; Benyó, Z.; et al. Elevated systemic TGF-β impairs aortic vasomotor function through activation of NADPH oxidase-driven superoxide production and leads to hypertension, myocardial remodeling, and increased plaque formation in apoE−/− mice. Am. J. Physiol.-Heart Circ. Physiol. 2010, 299, H386–H395. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Williams, K.J.; Borén, J. Subendothelial Lipoprotein Retention as the Initiating Process in Atherosclerosis. Circulation 2007, 116, 1832–1844. [Google Scholar] [CrossRef]
- Antoniades, C.; Cunnington, C.; Antonopoulos, A.; Neville, M.; Margaritis, M.; Demosthenous, M.; Bendall, J.; Hale, A.; Cerrato, R.; Tousoulis, D.; et al. Induction of vascular GTP-cyclohydrolase I and endogenous tetrahydrobiopterin synthesis protect against inflammation-induced endothelial dysfunction in human atherosclerosis. Circulation 2011, 124, 1860–1870. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat. Rev. Immunol. 2010, 10, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.; Ganz, P. Role of Endothelial Dysfunction in Atherosclerosis. Circulation 2004, 109, III-27–III-32. [Google Scholar] [CrossRef]
- Sarr, M.; Chataigneau, M.; Martins, S.; Schott, C.; El Bedoui, J.; Oak, M.H.; Muller, B.; Chataigneau, T.; Schini-Kerth, V.B. Red wine polyphenols prevent angiotensin II-induced hypertension and endothelial dysfunction in rats: Role of NADPH oxidase. Cardiovasc. Res. 2006, 71, 794–802. [Google Scholar] [CrossRef]
- Zanetti, M.; Gortan Cappellari, G.; Burekovic, I.; Barazzoni, R.; Stebel, M.; Guarnieri, G. Caloric restriction improves endothelial dysfunction during vascular aging: Effects on nitric oxide synthase isoforms and oxidative stress in rat aorta. Exp. Gerontol. 2010, 45, 848–855. [Google Scholar] [CrossRef]
- Zhang, H.; Morgan, B.; Potter, B.J.; Ma, L.; Dellsperger, K.C.; Ungvari, Z.; Zhang, C. Resveratrol improves left ventricular diastolic relaxation in type 2 diabetes by inhibiting oxidative/nitrative stress: In vivo demonstration with magnetic resonance imaging. Am. J. Physiol.-Heart Circ. Physiol. 2010, 299, H985–H994. [Google Scholar] [CrossRef]
- Keidar, S.; Attias, J.; Smith, J.; Breslow, J.L.; Hayek, T. The angiotensin-II receptor antagonist, losartan, inhibits LDL lipid peroxidation and atherosclerosis in apolipoprotein E-deficient mice. Biochem. Biophys. Res. Commun. 1997, 236, 622–625. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.S.; Choi, J.Y.; Kim, J.Y.; Han, S.H.; Park, J.E. Simvastatin and losartan differentially and synergistically inhibit atherosclerosis in apolipoprotein e(−/−) mice. Korean Circ. J. 2012, 42, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; West, N.E.; Black, E.; McDonald, D.; Ratnatunga, C.; Pillai, R.; Channon, K.M. Functional effect of the C242T polymorphism in the NAD(P)H oxidase p22phox gene on vascular superoxide production in atherosclerosis. Circulation 2000, 102, 1744–1747. [Google Scholar] [CrossRef]
- Azumi, H.; Inoue, N.; Takeshita, S.; Rikitake, Y.; Kawashima, S.; Hayashi, Y.; Itoh, H.; Yokoyama, M. Expression of NADH/NADPH oxidase p22phox in human coronary arteries. Circulation 1999, 100, 1494–1498. [Google Scholar] [CrossRef]
- Ying, C.J.; Xu, J.W.; Ikeda, K.; Takahashi, K.; Nara, Y.; Yamori, Y. Tea polyphenols regulate nicotinamide adenine dinucleotide phosphate oxidase subunit expression and ameliorate angiotensin II-induced hyperpermeability in endothelial cells. Hypertens. Res. 2003, 26, 823–828. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Loke, W.M.; Proudfoot, J.M.; Hodgson, J.M.; McKinley, A.J.; Hime, N.; Magat, M.; Stocker, R.; Croft, K.D. Specific dietary polyphenols attenuate atherosclerosis in apolipoprotein E-knockout mice by alleviating inflammation and endothelial dysfunction. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 749–757. [Google Scholar] [CrossRef]
- Loke, W.M.; Proudfoot, J.M.; Stewart, S.; McKinley, A.J.; Needs, P.W.; Kroon, P.A.; Hodgson, J.M.; Croft, K.D. Metabolic transformation has a profound effect on anti-inflammatory activity of flavonoids such as quercetin: Lack of association between antioxidant and lipoxygenase inhibitory activity. Biochem. Pharmacol. 2008, 75, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Hong, H.-Y.; Moon, H.J.; Lee, B.-H.; Kim, I.-S.; Kwon, I.C.; Rhee, K. A new atherosclerotic lesion probe based on hydrophobically modified chitosan nanoparticles functionalized by the atherosclerotic plaque targeted peptides. J. Control Release 2008, 128, 217–223. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ihm, S.-H.; Park, S.-H.; Lee, J.-O.; Kim, O.-R.; Park, E.-H.; Kim, K.-R.; Kim, J.-H.; Hwang, B.-H.; Youn, H.-J.; Oak, M.-H.; et al. A Standardized Lindera obtusiloba Extract Improves Endothelial Dysfunction and Attenuates Plaque Development in Hyperlipidemic ApoE-Knockout Mice. Plants 2021, 10, 2493. https://doi.org/10.3390/plants10112493
Ihm S-H, Park S-H, Lee J-O, Kim O-R, Park E-H, Kim K-R, Kim J-H, Hwang B-H, Youn H-J, Oak M-H, et al. A Standardized Lindera obtusiloba Extract Improves Endothelial Dysfunction and Attenuates Plaque Development in Hyperlipidemic ApoE-Knockout Mice. Plants. 2021; 10(11):2493. https://doi.org/10.3390/plants10112493
Chicago/Turabian StyleIhm, Sang-Hyun, Sin-Hee Park, Jung-Ok Lee, Ok-Ran Kim, Eun-Hye Park, Kyoung-Rak Kim, Jong-Hoon Kim, Byung-Hee Hwang, Ho-Joong Youn, Min-Ho Oak, and et al. 2021. "A Standardized Lindera obtusiloba Extract Improves Endothelial Dysfunction and Attenuates Plaque Development in Hyperlipidemic ApoE-Knockout Mice" Plants 10, no. 11: 2493. https://doi.org/10.3390/plants10112493
APA StyleIhm, S.-H., Park, S.-H., Lee, J.-O., Kim, O.-R., Park, E.-H., Kim, K.-R., Kim, J.-H., Hwang, B.-H., Youn, H.-J., Oak, M.-H., & Chang, K. (2021). A Standardized Lindera obtusiloba Extract Improves Endothelial Dysfunction and Attenuates Plaque Development in Hyperlipidemic ApoE-Knockout Mice. Plants, 10(11), 2493. https://doi.org/10.3390/plants10112493