
Ecological Dynamics and Microbial Treatments against Oomycete Plant Pathogens


Abstract
:1. Introduction
2. Ecological Processes and Their Effects on the Introduction and Success of Microbial Biocontrol Agents
2.1. Community Assembly Processes and Context
| Colonization/Establishment: The ability of an organism to start a new population in a novel or uncolonized habitat. |
| Community invasibility: The ability for a species or population to establish itself and grow in a habitat occupied by a community of different organisms. |
| Dispersal: The movement of organisms among and within habitats and habitat patches. |
| Ecological drift: The change of species abundances within a community over time due to stochastic processes. |
| Environmental filters: The selection of a subset of species that can withstand the abiotic conditions of an environment and determines a community. |
| Metacommunity: An interconnected community of multiple species that is spread across different habitat patches. |
| Metapopulation: An interconnected population of one species that is spread across different habitat patches. |
| Neutral theory in ecology: Simplest model possible for biogeographical patterns of diversity in which all species are first assumed to be equally capable of competing. Neutral drift and random dispersal are main factors that shape community assembly. |
| Niche theory: The distribution of species due to their n-dimensional hypervolume or the space corresponding to species’ requirements (habitat, environmental conditions, food, etc.). This framework is contrary to the neutral theory and suggests certain species are better suited for particular environments and community assembly processes are deterministic. |
| Patch dynamics: The interconnectedness of populations and communities across mosaic landscapes composed of heterogeneous patches or habitats. The distribution, size, and interconnectedness of patches have an effect of biodiversity maintenance. |
| Phenotypic plasticity: The variation of phenotypic traits observed due to differences in environmental conditions. Variation in phenotypic plasticity gives rise to genotype by environment (G×E) interactions. |
| Priority effects: The occurrence of earlier arrivals to a habitat having an advantage for establishment compared to later arrivals during community assembly. |
| Reaction norm: Pattern of a genotype’s trait expression across different environments. The slope of the reaction norm gives information regarding how responsive a trait is to environmental variation. |
| Source-sink dynamics: An aspect of patch dynamics where some high quality patches represent sources of species or populations while other poor-quality patches represent sinks. |
2.1.1. Priority Effects
2.1.2. Community Dynamics, Resource Competition, and Niche Space
2.1.3. Phenotypic Plasticity
2.2. The Effect of Spatial Dynamics on Microbial Biocontrol Agents: Metapopulation and Metacommunity Perspectives
3. Challenges and Limitations of Microbial Manipulations in Agricultural Systems
4. Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229. [Google Scholar] [CrossRef] [Green Version]
- Hacquard, S.; Garrido-Oter, R.; González, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.; Dangl, J.; Knight, R.; Ley, R.; et al. Microbiota and host nutrition across plant and animal kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef] [Green Version]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Ravel, J. The vocabulary of microbiome research: A proposal. Microbiome 2015, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313. [Google Scholar] [CrossRef]
- Bakker, M.G.; Manter, D.K.; Sheflin, A.M.; Weir, T.L.; Vivanco, J.M. Harnessing the rhizosphere microbiome through plant breeding and agricultural management. Plant Soil 2012, 360, 1–13. [Google Scholar] [CrossRef]
- Lozupone, C.A. Unraveling interactions between the microbiome and the host immune system to decipher mechanisms of disease. mSystems 2018, 3, e00183-17. [Google Scholar] [CrossRef] [Green Version]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Christian, N.; Whitaker, B.; Clay, K. Microbiomes: Unifying animal and plant systems through the lens of community ecology theory. Front. Microbiol. 2015, 6, 869. [Google Scholar] [CrossRef] [Green Version]
- Hird, S.M. Evolutionary biology needs wild microbiomes. Front. Microbiol. 2017, 8, 725. [Google Scholar] [CrossRef]
- Busby, P.E.; Soman, C.; Wagner, M.R.; Friesen, M.L.; Kremer, J.; Bennett, A.; Morsy, M.; Eisen, J.A.; Leach, J.E.; Dangl, J.L. Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 2017, 15, e2001793. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiol. Rev. 2016, 41, 109–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.; Raaijmakers, J.M. Cross-kingdom similarities in microbiome functions. ISME J. 2015, 9, 1905–1907. [Google Scholar] [CrossRef] [Green Version]
- Flandroy, L.; Poutahidis, T.; Berg, G.; Clarke, G.; Dao, M.C.; Decaestecker, E.; Furman, E.; Haahtela, T.; Massart, S.; Plovier, H.; et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci. Total Environ. 2018, 627, 1018–1038. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A. Regulation of the immune system by biodiversity from the natural environment: An ecosystem service essential to health. Proc. Natl. Acad. Sci. USA 2013, 110, 18360. [Google Scholar] [CrossRef] [Green Version]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330. [Google Scholar] [CrossRef] [Green Version]
- Costello, E.K.; Stagaman, K.; Dethlefsen, L.; Bohannan, B.J.M.; Relman, D.A. The application of ecological theory toward an understanding of the human microbiome. Science 2012, 336, 1255. [Google Scholar] [CrossRef] [Green Version]
- Relman, D.A. How to build healthy growth-promoting gut communities. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 379. [Google Scholar] [CrossRef]
- Walter, J.; Maldonado-Gómez, M.X.; Martínez, I. To engraft or not to engraft: An ecological framework for gut microbiome modulation with live microbes. Curr. Opin. Biotechnol. 2018, 49, 129–139. [Google Scholar] [CrossRef]
- van Nood, E.; Vrieze, A.; Nieuwdorp, M.; Fuentes, S.; Zoetendal, E.G.; de Vos, W.M.; Visser, C.E.; Kuijper, E.J.; Bartelsman, J.F.; Tijssen, J.G.; et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N. Engl. J. Med. 2013, 368, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Shahinas, D.; Silverman, M.; Sittler, T.; Chiu, C.; Kim, P.; Allen-Vercoe, E.; Weese, S.; Wong, A.; Low, D.E.; Pillai, D.R. Toward an understanding of changes in diversity associated with fecal microbiome transplantation based on 16S rRNA gene deep sequencing. mBio 2012, 3, e00338-12. [Google Scholar] [CrossRef] [Green Version]
- Clemente, J.; Ursell, L.; Parfrey, L.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Clarke, G.; Berk, M.; Jacka, F.N. The gut microbiome and diet in psychiatry: Focus on depression. Curr. Opin. Psychiatry 2015, 28, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, e00338-12. [Google Scholar] [CrossRef] [Green Version]
- Thines, M.; Kamoun, S. Oomycete–plant coevolution: Recent advances and future prospects. Curr. Opin. Plant Biol. 2010, 13, 427–433. [Google Scholar] [CrossRef]
- Judelson, H.S. Dynamics and innovations within oomycete genomes: Insights into biology, pathology, and evolution. Eukaryot. Cell 2012, 11, 1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.; Anderson, V.L.; Robertson, E.J.; Secombes, C.J.; van West, P. New insights into animal pathogenic oomycetes. Trends Microbiol. 2008, 16, 13–19. [Google Scholar] [CrossRef]
- Mendoza, L.; Hernandez, F.; Ajello, L. Life cycle of the human and animal oomycete pathogen Pythium insidiosum. J. Clin. Microbiol. 1993, 31, 2967. [Google Scholar] [CrossRef] [Green Version]
- Ploch, S.; Thines, M. Obligate biotrophic pathogens of the genus Albugo are widespread as asymptomatic endophytes in natural populations of Brassicaceae. Mol. Ecol. 2011, 20, 3692–3699. [Google Scholar] [CrossRef] [PubMed]
- Spring, O.; Gomez-Zeledon, J.; Hadziabdic, D.; Trigiano, R.N.; Thines, M.; Lebeda, A. Biological characteristics and assessment of virulence diversity in pathosystems of economically important biotrophic oomycetes. Crit. Rev. Plant Sci. 2018, 37, 439–495. [Google Scholar] [CrossRef]
- Derevnina, L.; Petre, B.; Kellner, R.; Dagdas, Y.F.; Sarowar, M.N.; Giannakopoulou, A.; De la Concepcion, J.C.; Chaparro-Garcia, A.; Pennington, H.G.; van West, P.; et al. Emerging oomycete threats to plants and animals. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2016, 371, 20150459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bebber, D.P.; Gurr, S.J. Crop-destroying fungal and oomycete pathogens challenge food security. Fungal Genet. Biol. 2015, 74, 62–64. [Google Scholar] [CrossRef]
- Tyler, B.M. Genetics and genomics of the oomycete–host interface. Trends Genet. 2001, 17, 611–614. [Google Scholar] [CrossRef]
- Klinter, S.; Bulone, V.; Arvestad, L. Diversity and evolution of chitin synthases in oomycetes (Straminipila: Oomycota). Mol. Phylogenet. Evol. 2019, 139, 106558. [Google Scholar] [CrossRef]
- Mélida, H.; Sandoval-Sierra, J.V.; Diéguez-Uribeondo, J.; Bulone, V. Analyses of extracellular carbohydrates in oomycetes unveil the existence of three different cell wall types. Eukaryot. Cell 2012, 12, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Gaulin, E.; Bottin, A.; Dumas, B. Sterol biosynthesis in oomycete pathogens. Plant Signal. Behav. 2010, 5, 258–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlin, P.; Srivastava, V.; Ekengren, S.; McKee, L.S.; Bulone, V. Comparative analysis of sterol acquisition in the oomycetes Saprolegnia parasitica and Phytophthora infestans. PLoS ONE 2017, 12, e0170873. [Google Scholar] [CrossRef]
- Madoui, M.A.; Bertrand-Michel, J.; Gaulin, E.; Dumas, B. Sterol metabolism in the oomycete Aphanomyces euteiches, a legume root pathogen. New Phytol. 2009, 183, 291–300. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Chen, P.C.; Tsay, T.T. The biocontrol efficacy and antibiotic activity of Streptomyces plicatus on the oomycete Phytophthora capsici. Biol. Control 2016, 98, 34–42. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Michelsen, C.F.; Watrous, J.; Glaring, M.A.; Kersten, R.; Koyama, N.; Dorrestein, P.C.; Stougaard, P. Nonribosomal peptides, key biocontrol components for Pseudomonas fluorescens In5, isolated from a Greenlandic suppressive soil. mBio 2015, 6, e00079-15. [Google Scholar] [CrossRef] [Green Version]
- Ezziyyani, M.; Requena, M.E.; Egea-Gilabert, C.; Candela, M.E. Biological control of Phytophthora root rot of pepper Using Trichoderma harzianum and Streptomyces rochei in combination. J. Phytopathol. 2007, 155, 342–349. [Google Scholar] [CrossRef]
- van Dijk, K.; Nelson, E.B. Fatty acid competition as a mechanism by which Enterobacter cloacae suppresses Pythium ultimum sporangium germination and damping-off. Appl. Environ. Microbiol. 2000, 66, 5340–5347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnell, J.J.; Berka, R.; Young, H.A.; Sturino, J.M.; Kang, Y.; Barnhart, D.M.; DiLeo, M.V. From the lab to the farm: An industrial perspective of plant beneficial microorganisms. Front. Plant Sci. 2016, 7, 1110. [Google Scholar] [CrossRef]
- Dorn, B.; Musa, T.; Krebs, H.; Fried, P.M.; Forrer, H.R. Control of late blight in organic potato production: Evaluation of copper-free preparations under field, growth chamber and laboratory conditions. Eur. J. Plant Pathol. 2007, 119, 217–240. [Google Scholar] [CrossRef]
- Cunniffe, N.J.; Gilligan, C.A. A theoretical framework for biological control of soil-borne plant pathogens: Identifying effective strategies. J. Theor. Biol. 2011, 278, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhl, J.; Postma, J.; Nicot, P.; Ruocco, M.; Blum, B. Stepwise screening of microorganisms for commercial use in biological control of plant–pathogenic fungi and bacteria. Biol. Control 2011, 57, 1–12. [Google Scholar] [CrossRef]
- Smits, W.K.; Kuipers, O.P.; Veening, J.W. Phenotypic variation in bacteria: The role of feedback regulation. Nat. Rev. Microbiol. 2006, 4, 259–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Kim, Y.S.; Balaraju, K.; Seo, Y.S.; Park, J.; Ryu, C.M.; Park, S.H.; Kim, J.F.; Kang, S.; Jeon, Y. Molecular changes associated with spontaneous phenotypic variation of Paenibacillus polymyxa, a commonly used biocontrol agent, and temperature-dependent control of variation. Sci. Rep. 2020, 10, 16586. [Google Scholar] [CrossRef] [PubMed]
- Ambrico, A.; Trupo, M.; Magarelli, R.A. Influence of phenotypic dissociation in Bacillus subtilis strain ET-1 on Iturin A production. Curr. Microbiol. 2019, 76, 1487–1494. [Google Scholar] [CrossRef]
- Stone, B.W.G.; Weingarten, E.A.; Jackson, C.R. The role of the phyllosphere microbiome in plant health and function. Annu. Plant Rev. Online 2018, 1, 533–556. [Google Scholar] [CrossRef]
- Griggs, R.G.; Steenwerth, K.L.; Mills, D.A.; Cantu, D.; Bokulich, N.A. Sources and assembly of microbial communities in vineyards as a functional component of winegrowing. Front. Microbiol. 2021, 12, 673810. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wei, Z.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Rhizosphere microbiome functional diversity and pathogen invasion resistance build up during plant development. Environ. Microbiol. 2020, 22, 5005–5018. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Tao, J.; Liu, T.; Liu, Y.; Xiao, N.; Li, T.; Gu, Y.; Yin, H.; Meng, D. Responses of phyllosphere microbiota and plant health to application of two different biocontrol agents. AMB Express 2019, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jousset, A.; Becker, J.; Chatterjee, S.; Karlovsky, P.; Scheu, S.; Eisenhauer, N. Biodiversity and species identity shape the antifungal activity of bacterial communities. Ecology 2014, 95, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Köberl, M.; Rybakova, D.; Müller, H.; Grosch, R.; Smalla, K. Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 2017, 93, fix050. [Google Scholar] [CrossRef] [PubMed]
- Arif, I.; Batool, M.; Schenk, P.M. Plant microbiome engineering: Expected benefits for improved crop growth and resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Jin, K.; Raaijmakers, J.M. Designing a home for beneficial plant microbiomes. Curr. Opin. Plant Biol. 2021, 62, 102025. [Google Scholar] [CrossRef] [PubMed]
- Adam, E.; Groenenboom, A.E.; Kurm, V.; Rajewska, M.; Schmidt, R.; Tyc, O.; Weidner, S.; Berg, G.; de Boer, W.; Falcão Salles, J. Controlling the microbiome: Microhabitat adjustments for successful biocontrol strategies in soil and human gut. Front. Microbiol. 2016, 7, 1079. [Google Scholar] [CrossRef] [PubMed]
- Vassileva, M.; Flor-Peregrin, E.; Malusá, E.; Vassilev, N. Towards better understanding of the interactions and efficient application of plant beneficial prebiotics, probiotics, postbiotics and synbiotics. Front. Plant Sci. 2020, 11, 1068. [Google Scholar] [CrossRef] [PubMed]
- Kehe, J.; Kulesa, A.; Ortiz, A.; Ackerman, C.M.; Thakku, S.G.; Sellers, D.; Kuehn, S.; Gore, J.; Friedman, J.; Blainey, P.C. Massively parallel screening of synthetic microbial communities. Proc. Natl. Acad. Sci. USA 2019, 116, 12804–12809. [Google Scholar] [CrossRef] [Green Version]
- Johns, N.I.; Blazejewski, T.; Gomes, A.L.C.; Wang, H.H. Principles for designing synthetic microbial communities. Curr. Opin. Microbiol. 2016, 31, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Yang, T.; Friman, V.P.; Kowalchuk, G.A.; Hautier, Y.; Li, M.; Wei, Z.; Xu, Y.; Shen, Q.; Jousset, A. Introduction of probiotic bacterial consortia promotes plant growth via impacts on the resident rhizosphere microbiome. Proc. R. Soc. B Biol. Sci. 2021, 288, 20211396. [Google Scholar] [CrossRef]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography, Monographs in Population Biology; Princeton Univ. Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Carlström, C.I.; Field, C.M.; Bortfeld-Miller, M.; Müller, B.; Sunagawa, S.; Vorholt, J.A. Synthetic microbiota reveal priority effects and keystone strains in the Arabidopsis phyllosphere. Nat. Ecol. Evol. 2019, 3, 1445–1454. [Google Scholar] [CrossRef]
- Leopold, D.R.; Busby, P.E. Host genotype and colonist arrival order jointly govern plant microbiome composition and function. Curr. Biol. 2020, 30, 3260–3266.e5. [Google Scholar] [CrossRef] [PubMed]
- Halliday, F.W.; Umbanhowar, J.; Mitchell, C.E. Interactions among symbionts operate across scales to influence parasite epidemics. Ecol. Lett. 2017, 20, 1285–1294. [Google Scholar] [CrossRef]
- Tkacz, A.; Bestion, E.; Bo, Z.; Hortala, M.; Poole, P.S. Influence of plant fraction, soil, and plant species on microbiota: A multikingdom comparison. mBio 2020, 11, e02785-19. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Gu, Y.; Friman, V.P.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Initial soil microbiome composition and functioning predetermine future plant health. Sci. Adv. 2019, 5, eaaw0759. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.M.; Becker-Uncapher, I.; Carlson, M.; Fierer, N. Variable influences of soil and seed-associated bacterial communities on the assembly of seedling microbiomes. ISME J. 2021, 15, 2748–2762. [Google Scholar] [CrossRef] [PubMed]
- Amend, A.S.; Cobian, G.M.; Laruson, A.J.; Remple, K.; Tucker, S.J.; Poff, K.E.; Antaky, C.; Boraks, A.; Jones, C.A.; Kuehu, D.; et al. Phytobiomes are compositionally nested from the ground up. PeerJ 2019, 7, e6609. [Google Scholar] [CrossRef] [Green Version]
- Jack, A.L.H.; Nelson, E.B. A seed-recruited microbiome protects developing seedlings from disease by altering homing responses of Pythium aphanidermatum zoospores. Plant Soil 2018, 422, 209–222. [Google Scholar] [CrossRef]
- Löbmann, M.T.; Vetukuri, R.R.; de Zinger, L.; Alsanius, B.W.; Grenville-Briggs, L.J.; Walter, A.J. The occurrence of pathogen suppressive soils in Sweden in relation to soil biota, soil properties, and farming practices. Appl. Soil Ecol. 2016, 107, 57–65. [Google Scholar] [CrossRef]
- Gómez Expósito, R.; de Bruijn, I.; Postma, J.; Raaijmakers, J.M. Current insights into the role of rhizosphere bacteria in disease suppressive soils. Front. Microbiol. 2017, 8, 2529. [Google Scholar] [CrossRef] [PubMed]
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144. [Google Scholar] [CrossRef]
- Fukami, T. Historical contingency in community assembly: Integrating niches, species pools, and priority effects. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Bazghaleh, N.; Prashar, P.; Woo, S.; Vandenberg, A. Effects of lentil genotype on the colonization of beneficial Trichoderma species and biocontrol of Aphanomyces root rot. Microorganisms 2020, 8, 1290. [Google Scholar] [CrossRef]
- Singh, P.; Santoni, S.; This, P.; Péros, J.P. Genotype-environment interaction shapes the microbial assemblage in grapevine’s phyllosphere and carposphere: An NGS aproach. Microorganisms 2018, 6, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, K.; Alcalá-Briseño, R.; Andersen, K.; Buddenhagen, C.; Choudhury, R.; Fulton, J.; Nopsa, J.H.; Poudel, R.; Xing, Y. Network analysis: A systems framework to address grand challenges in plant pathology. Annu. Rev. Phytopathol. 2018, 56, 559–580. [Google Scholar] [CrossRef]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.T.; Weigel, D.; Kemen, E.M. Microbial hub taxa link host and abiotic factors to plant microbiome variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [Green Version]
- Larousse, M.; Rancurel, C.; Syska, C.; Palero, F.; Etienne, C.; Industri, B.; Nesme, X.; Bardin, M.; Galiana, E. Tomato root microbiota and Phytophthora parasitica-associated disease. Microbiome 2017, 5, 56. [Google Scholar] [CrossRef]
- Hol, W.H.G.; Garbeva, P.; Hordijk, C.; Hundscheid, M.P.J.; Gunnewiek, P.J.A.K.; van Agtmaal, M.; Kuramae, E.E.; de Boer, W. Non-random species loss in bacterial communities reduces antifungal volatile production. Ecology 2015, 96, 2042–2048. [Google Scholar] [CrossRef] [Green Version]
- Hennessy, R.C.; Glaring, M.A.; Olsson, S.; Stougaard, P. Transcriptomic profiling of microbe–microbe interactions reveals the specific response of the biocontrol strain P. fluorescens In5 to the phytopathogen Rhizoctonia solani. BMC Res. Notes 2017, 10, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Z.; Yang, T.; Friman, V.P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic network architecture of root-associated bacterial communities determines pathogen invasion and plant health. Nat. Commun. 2015, 6, 8413. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Müller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Münch, P.C.; Spaepen, S.; Remus-Emsermann, M.; et al. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 2015, 528, 364. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Salas Gonzalez, I.; Mittelviefhaus, M.; Clingenpeel, S.; Herrera Paredes, S.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic features of bacterial adaptation to plants. Nat. Genet. 2018, 50, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Kong, P.; Hong, C. Soil bacteria as sources of virulence signal providers promoting plant infection by Phytophthora pathogens. Sci. Rep. 2016, 6, 33239. [Google Scholar] [CrossRef] [Green Version]
- McMullan, M.; Gardiner, A.; Bailey, K.; Kemen, E.; Ward, B.J.; Cevik, V.; Robert-Seilaniantz, A.; Schultz-Larsen, T.; Balmuth, A.; Holub, E.; et al. Evidence for suppression of immunity as a driver for genomic introgressions and host range expansion in races of Albugo candida, a generalist parasite. eLife 2015, 4, e04550. [Google Scholar] [CrossRef]
- Wagner, M.R.; Lundberg, D.S.; Del Rio, T.G.; Tringe, S.G.; Dangl, J.L.; Mitchell-Olds, T. Host genotype and age shape the leaf and root microbiomes of a wild perennial plant. Nat. Commun. 2016, 7, 12151. [Google Scholar] [CrossRef]
- Yergeau, E.; Bell, T.H.; Champagne, J.; Maynard, C.; Tardif, S.; Tremblay, J.; Greer, C.W. Transplanting soil microbiomes leads to lasting effects on willow growth, but not on the rhizosphere microbiome. Front. Microbiol. 2015, 6, 1436. [Google Scholar] [CrossRef] [Green Version]
- Henry, L.P.; Bruijning, M.; Forsberg, S.K.G.; Ayroles, J.F. The microbiome extends host evolutionary potential. Nat. Commun. 2021, 12, 5141. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.P.; Handelsman, J.; Goodman, R.M. Genetic basis in plants for interactions with disease-suppressive bacteria. Proc. Natl. Acad. Sci. USA 1999, 96, 4786–4790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottesen, A.R.; Gorham, S.; Reed, E.; Newell, M.J.; Ramachandran, P.; Canida, T.; Allard, M.; Evans, P.; Brown, E.; White, J.R. Using a control to better understand phyllosphere microbiota. PLoS ONE 2016, 11, e0163482. [Google Scholar] [CrossRef] [Green Version]
- Ostfeld, R.S.; Glass, G.E.; Keesing, F. Spatial epidemiology: An emerging (or re-emerging) discipline. Trends Ecol. Evol. 2005, 20, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Otten, W.; Gilligan, C.A. Soil structure and soil-borne diseases: Using epidemiological concepts to scale from fungal spread to plant epidemics. Eur. J. Soil Sci. 2006, 57, 26–37. [Google Scholar] [CrossRef]
- Campbell, C.; Madden, L. Introduction to Plant Disease Epidemiology; Wiley: New York, NY, USA, 1990. [Google Scholar]
- Neupane, K.; Baysal-Gurel, F. Automatic identification and monitoring of plant diseases using unmanned aerial vehicles: A review. Remote Sens. 2021, 13, 3841. [Google Scholar] [CrossRef]
- Rodríguez, J.; Lizarazo, I.; Prieto, F.; Angulo-Morales, V. Assessment of potato late blight from UAV-based multispectral imagery. Comput. Electron. Agric. 2021, 184, 106061. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Perspectives for remote sensing with unmanned aerial vehicles in precision agriculture. Trends Plant Sci. 2019, 24, 152–164. [Google Scholar] [CrossRef]
- Calderón, R.; Montes-Borrego, M.; Landa, B.B.; Navas-Cortés, J.A.; Zarco-Tejada, P.J. Detection of downy mildew of opium poppy using high-resolution multi-spectral and thermal imagery acquired with an unmanned aerial vehicle. Precis. Agric. 2014, 15, 639–661. [Google Scholar] [CrossRef]
- Packer, A.; Clay, K. Soil pathogens and spatial patterns of seedling mortality in a temperate tree. Nature 2000, 404, 278–281. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Larkin, R.P.; Campbell, C.L. Spatial and temporal dynamics of Phytophthora epidemics in commercial bell pepper fields. Phytopathology 1993, 83, 1312–1320. [Google Scholar] [CrossRef]
- Skelsey, P.; Rossing, W.A.H.; Kessel, G.J.T.; van der Werf, W. Invasion of Phytophthora infestans at the landscape level: How do spatial scale and weather modulate the consequences of spatial heterogeneity in host resistance? Phytopathology 2010, 100, 1146–1161. [Google Scholar] [CrossRef] [Green Version]
- Leibold, M.; Holyoak, M.; Mouquet, N. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Plantegenest, M.; May, C.L.; Fabre, F. Landscape epidemiology of plant diseases. J. R. Soc. Interface 2007, 4, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Park, A.W.; Gubbins, S.; Gilligan, C.A. Invasion and persistence of plant parasites in a spatially structured host population. Oikos 2001, 94, 162–174. [Google Scholar] [CrossRef]
- Thrall, P.H.; Burdon, J.J. Host-pathogen dynamics in a metapopulation context: The ecological and evolutionary consequences of being spatial. J. Ecol. 1997, 85, 743–753. [Google Scholar] [CrossRef]
- Papaïx, J.; Burdon, J.J.; Lannou, C.; Thrall, P.H. Evolution of pathogen specialisation in a host metapopulation: Joint effects of host and pathogen dispersal. PLoS Comput. Biol. 2014, 10, e1003633. [Google Scholar] [CrossRef] [PubMed]
- Holyoak, M.; Leibold, M.; Holt, R. Metacommunities: Spatial Dynamics and Ecological Communities; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Urban, M.C.; Skelly, D.K. Evolving metacommunities: Toward an evolutionary perspective on metacommunities. Ecology 2006, 87, 1616–1626. [Google Scholar] [CrossRef] [Green Version]
- Borer, E.T.; Laine, A.L.; Seabloom, E.W. A multiscale approach to plant disease using the metacommunity concept. Annu. Rev. Phytopathol. 2016, 54, 397–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, B.; Levine, J.M. Ecological drift and the distribution of species diversity. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170507. [Google Scholar] [CrossRef] [PubMed]
- Richards, T.A.; Soanes, D.M.; Jones, M.D.M.; Vasieva, O.; Leonard, G.; Paszkiewicz, K.; Foster, P.G.; Hall, N.; Talbot, N.J. Horizontal gene transfer facilitated the evolution of plant parasitic mechanisms in the oomycetes. Proc. Natl. Acad. Sci. USA 2011, 108, 15258–15263. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Mihaljevic, J.R. Linking metacommunity theory and symbiont evolutionary ecology. Trends Ecol. Evol. 2012, 27, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, R.; Nicolaisen, M. Cropping history shapes fungal, oomycete and nematode communities in arable soils and affects cavity spot in carrot. Agric. Ecosyst. Environ. 2018, 257, 120–131. [Google Scholar] [CrossRef]
- Li, X.; Jousset, A.; de Boer, W.; Carrión, V.J.; Zhang, T.; Wang, X.; Kuramae, E.E. Legacy of land use history determines reprogramming of plant physiology by soil microbiome. ISME J. 2019, 13, 738–751. [Google Scholar] [CrossRef] [Green Version]
- Pillai, P.; Gouhier, T.C.; Vollmer, S.V. The cryptic role of biodiversity in the emergence of host–microbial mutualisms. Ecol. Lett. 2014, 17, 1437–1446. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Gao, D.; Zhou, X.; Chen, S.; Li, C.; Wu, F. Intercropping with potato-onion enhanced the soil microbial diversity of tomato. Microorganisms 2020, 8, 834. [Google Scholar] [CrossRef]
- Wang, G.; Bei, S.; Li, J.; Bao, X.; Zhang, J.; Schultz, P.A.; Li, H.; Li, L.; Zhang, F.; Bever, J.D.; et al. Soil microbial legacy drives crop diversity advantage: Linking ecological plant–soil feedback with agricultural intercropping. J. Appl. Ecol. 2021, 58, 496–506. [Google Scholar] [CrossRef]
- Blumenstein, K.; Albrectsen, B.R.; Martín, J.A.; Hultberg, M.; Sieber, T.N.; Helander, M.; Witzell, J. Nutritional niche overlap potentiates the use of endophytes in biocontrol of a tree disease. BioControl 2015, 60, 655–667. [Google Scholar] [CrossRef]
- Zhu, S.; Morel, J.B. Molecular mechanisms underlying microbial disease control in intercropping. Mol. Plant-Microbe Interact. 2018, 32, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Bouws, H.; Finckh, M.R. Effects of strip intercropping of potatoes with non-hosts on late blight severity and tuber yield in organic production. Plant Pathol. 2008, 57, 916–927. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, Y.; Qi, L.; Mei, X.; Liao, J.; Ding, X.; Deng, W.; Fan, L.; He, X.; Vivanco, J.M.; et al. Plant-plant-microbe mechanisms involved in soil-borne disease suppression on a maize and pepper intercropping system. PLoS ONE 2015, 9, e115052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wille, L.; Messmer, M.M.; Bodenhausen, N.; Studer, B.; Hohmann, P. Heritable variation in pea for resistance against a root rot complex and its characterization by amplicon sequencing. Front. Plant Sci. 2020, 11, 542153. [Google Scholar] [CrossRef]
- Noel, Z.A.; Chang, H.X.; Chilvers, M.I. Variation in soybean rhizosphere oomycete communities from Michigan fields with contrasting disease pressures. Appl. Soil Ecol. 2020, 150, 103435. [Google Scholar] [CrossRef]
- Somera, T.S.; Freilich, S.; Mazzola, M. Comprehensive analysis of the apple rhizobiome as influenced by different Brassica seed meals and rootstocks in the same soil/plant system. Appl. Soil Ecol. 2021, 157, 103766. [Google Scholar] [CrossRef]
- Lakkis, S.; Trotel-Aziz, P.; Rabenoelina, F.; Schwarzenberg, A.; Nguema-Ona, E.; Clément, C.; Aziz, A. Strengthening grapevine resistance by Pseudomonas fluorescens PTA-CT2 relies on distinct defense pathways in susceptible and partially resistant genotypes to downy mildew and gray mold diseases. Front. Plant Sci. 2019, 10, 1112. [Google Scholar] [CrossRef] [PubMed]
- Bellini, A.; Ferrocino, I.; Cucu, M.A.; Pugliese, M.; Garibaldi, A.; Gullino, M.L. A compost treatment acts as a suppressive agent in Phytophthora capsici–Cucurbita pepo pathosystem by modifying the rhizosphere microbiota. Front. Plant Sci. 2020, 11, 885. [Google Scholar] [CrossRef]
- Wang, L.; Mazzola, M. Effect of soil physical conditions on emission of allyl isothiocyanate and subsequent microbial inhibition in response to Brassicaceae seed meal amendment. Plant Dis. 2018, 103, 846–852. [Google Scholar] [CrossRef]
- Soonvald, L.; Loit, K.; Runno-Paurson, E.; Astover, A.; Tedersoo, L. The role of long-term mineral and organic fertilisation treatment in changing pathogen and symbiont community composition in soil. Appl. Soil Ecol. 2019, 141, 45–53. [Google Scholar] [CrossRef]
- Asiloglu, R.; Samuel, S.O.; Sevilir, B.; Akca, M.O.; Acar Bozkurt, P.; Suzuki, K.; Murase, J.; Turgay, O.C.; Harada, N. Biochar affects taxonomic and functional community composition of protists. Biol. Fertil. Soils 2021, 57, 15–29. [Google Scholar] [CrossRef]
- Lee, S.; An, R.; Grewal, P.; Yu, Z.; Borherova, Z.; Lee, J. High-performing windowfarm hydroponic system: Transcriptomes of fresh produce and microbial communities in response to beneficial bacterial treatment. Mol. Plant-Microbe Interact. 2016, 29, 965–976. [Google Scholar] [CrossRef]
- McGehee, C.S.; Raudales, R.E.; Elmer, W.H.; McAvoy, R.J. Efficacy of biofungicides against root rot and damping-off of microgreens caused by Pythium spp. Crop Prot. 2019, 121, 96–102. [Google Scholar] [CrossRef]
- De Vrieze, M.; Pandey, P.; Bucheli, T.D.; Varadarajan, A.R.; Ahrens, C.H.; Weisskopf, L.; Bailly, A. Volatile organic compounds from native potato-associated Pseudomonas as potential anti-oomycete agents. Front. Microbiol. 2015, 6, 1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinchilla, D.; Bruisson, S.; Meyer, S.; Zühlke, D.; Hirschfeld, C.; Joller, C.; L’Haridon, F.; Mène-Saffrané, L.; Riedel, K.; Weisskopf, L. A sulfur-containing volatile emitted by potato-associated bacteria confers protection against late blight through direct anti-oomycete activity. Sci. Rep. 2019, 9, 18778. [Google Scholar] [CrossRef] [Green Version]
- Bruisson, S.; Zufferey, M.; L’Haridon, F.; Trutmann, E.; Anand, A.; Dutartre, A.; De Vrieze, M.; Weisskopf, L. Endophytes and epiphytes from the grapevine leaf microbiome as potential biocontrol agents against phytopathogens. Front. Microbiol. 2019, 10, 2726. [Google Scholar] [CrossRef] [Green Version]
- Agisha, V.N.; Kumar, A.; Eapen, S.J.; Sheoran, N.; Suseelabhai, R. Broad-spectrum antimicrobial activity of volatile organic compounds from endophytic Pseudomonas putida BP25 against diverse plant pathogens. Biocontrol Sci. Technol. 2019, 29, 1069–1089. [Google Scholar] [CrossRef]
- Khalaf, E.M.; Raizada, M.N. Bacterial seed endophytes of domesticated cucurbits antagonize Fungal and Oomycete pathogens including powdery mildew. Front. Microbiol. 2018, 9, 42. [Google Scholar] [CrossRef]
- Cucu, M.A.; Gilardi, G.; Pugliese, M.; Ferrocino, I.; Gullino, M.L. Effects of biocontrol agents and compost against the Phytophthora capsici of zucchini and their impact on the rhizosphere microbiota. Appl. Soil Ecol. 2020, 154, 103659. [Google Scholar] [CrossRef]
- Aleandri, M.P.; Chilosi, G.; Bruni, N.; Tomassini, A.; Vettraino, A.M.; Vannini, A. Use of nursery potting mixes amended with local Trichoderma strains with multiple complementary mechanisms to control soil-borne diseases. Crop Prot. 2015, 67, 269–278. [Google Scholar] [CrossRef]
- Granada, S.D.; Ramírez-Restrepo, S.; López-Luján, L.; Peláez-Jaramillo, C.A.; Bedoya-Pérez, J.C. Screening of a biological control bacterium to fight avocado diseases: From agroecosystem to bioreactor. Biocatal. Agric. Biotechnol. 2018, 14, 109–115. [Google Scholar] [CrossRef]
- Velázquez-Becerra, C.; Macías-Rodríguez, L.I.; López-Bucio, J.; Flores-Cortez, I.; Santoyo, G.; Hernández-Soberano, C.; Valencia-Cantero, E. The rhizobacterium Arthrobacter agilis produces dimethylhexadecylamine, a compound that inhibits growth of phytopathogenic fungi in vitro. Protoplasma 2013, 250, 1251–1262. [Google Scholar] [CrossRef]
- Méndez-Bravo, A.; Cortazar-Murillo, E.M.; Guevara-Avendaño, E.; Ceballos-Luna, O.; Rodríguez-Haas, B.; Kiel-Martínez, A.L.; Hernández-Cristóbal, O.; Guerrero-Analco, J.A.; Reverchon, F. Plant growth-promoting rhizobacteria associated with avocado display antagonistic activity against Phytophthora cinnamomi through volatile emissions. PLoS ONE 2018, 13, e0194665. [Google Scholar] [CrossRef] [Green Version]
- Cheffi, M.; Bouket, A.C.; Alenezi, F.N.; Luptakova, L.; Belka, M.; Vallat, A.; Rateb, M.E.; Tounsi, S.; Triki, M.A.; Belbahri, L. Olea europaea L. root endophyte Bacillus velezensis OEE1 counteracts oomycete and fungal harmful pathogens and harbours a large repertoire of secreted and volatile metabolites and beneficial functional genes. Microorganisms 2019, 7, 314. [Google Scholar] [CrossRef] [Green Version]
- Nandhini, M.; Rajini, S.B.; Udayashankar, A.C.; Niranjana, S.R.; Lund, O.S.; Shetty, H.S.; Prakash, H.S. Diversity, plant growth promoting and downy mildew disease suppression potential of cultivable endophytic fungal communities associated with pearl millet. Biol. Control 2018, 127, 127–138. [Google Scholar] [CrossRef]
- Vetukuri, R.R.; Masini, L.; McDougal, R.; Panda, P.; de Zinger, L.; Brus-Szkalej, M.; Lankinen, A.; Grenville-Briggs, L.J. The presence of Phytophthora infestans in the rhizosphere of a wild Solanum species may contribute to off-season survival and pathogenicity. Appl. Soil Ecol. 2020, 148, 103475. [Google Scholar] [CrossRef]
- Fiore-Donno, A.M.; Bonkowski, M. Different community compositions between obligate and facultative oomycete plant parasites in a landscape-scale metabarcoding survey. Biol. Fertil. Soils 2021, 57, 245–256. [Google Scholar] [CrossRef]
- Bakker, M.G.; Moorman, T.B.; Kaspar, T.C.; Manter, D.K. Isolation of cultivation-resistant oomycetes, first detected as amplicon sequences, from roots of herbicide-terminated winter rye. Phytobiomes J. 2017, 1, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Ebadzadsahrai, G.; Higgins Keppler, E.A.; Soby, S.D.; Bean, H.D. Inhibition of fungal growth and induction of a novel volatilome in response to Chromobacterium vaccinii volatile organic compounds. Front. Microbiol. 2020, 11, 1035. [Google Scholar] [CrossRef]
- Weinhold, A.; Karimi Dorcheh, E.; Li, R.; Rameshkumar, N.; Baldwin, I.T. Antimicrobial peptide expression in a wild tobacco plant reveals the limits of host-microbe-manipulations in the field. eLife 2018, 7, e28715. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, I.M.B.; Hockenhull, J.; Jensen, D.F.; Gerhardson, B.; Hökeberg, M.; Tahvonen, R.; Teperi, E.; Sundheim, L.; Henriksen, B. Selection of biological control agents for controlling soil and seed-borne diseases in the field. Eur. J. Plant Pathol. 1997, 103, 775–784. [Google Scholar] [CrossRef]
- Poorter, H.; Fiorani, F.; Pieruschka, R.; Wojciechowski, T.; van der Putten, W.H.; Kleyer, M.; Schurr, U.; Postma, J. Pampered inside, pestered outside? Differences and similarities between plants growing in controlled conditions and in the field. New Phytol. 2016, 212, 838–855. [Google Scholar] [CrossRef]
- Bardin, M.; Ajouz, S.; Comby, M.; Lopez-Ferber, M.; Graillot, B.; Siegwart, M.; Nicot, P.C. Is the efficacy of biological control against plant diseases likely to be more durable than that of chemical pesticides? Front. Plant Sci. 2015, 6, 566. [Google Scholar] [CrossRef]
- Cai, G.; Myers, K.; Fry, W.E.; Hillman, B.I. Phytophthora infestans RNA virus 2, a novel RNA virus from Phytophthora infestans, does not belong to any known virus group. Arch. Virol. 2019, 164, 567–572. [Google Scholar] [CrossRef]
- Cai, G.; Fry, W.E.; Hillman, B.I. PiRV-2 stimulates sporulation in Phytophthora infestans. Virus Res. 2019, 271, 197674. [Google Scholar] [CrossRef]
- Kwak, M.J.; Kong, H.G.; Choi, K.; Kwon, S.K.; Song, J.Y.; Lee, J.; Lee, P.A.; Choi, S.Y.; Seo, M.; Lee, H.J.; et al. Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat. Biotechnol. 2018, 36, 1100. [Google Scholar] [CrossRef] [PubMed]
- Zahn, G.; Amend, A.S. Foliar microbiome transplants confer disease resistance in a critically-endangered plant. PeerJ 2017, 5, e4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clay, K.; Kover, P.X. The red queen hypothesis and plant/pathogen interactions. Annu. Rev. Phytopathol. 1996, 34, 29–50. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Jousset, A. Plant breeding goes microbial. Trends Plant Sci. 2017, 22, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, I.; Bokulich, N.A.; Caporaso, J.G.; Enders, L.S.; Ghanem, W.; Ingerslew, K.S. Phylogenetic farming: Can evolutionary history predict crop rotation via the soil microbiome? Evol. Appl. 2020, 13, 1984–1999. [Google Scholar] [CrossRef] [Green Version]
- Panke-Buisse, K.; Poole, A.C.; Goodrich, J.K.; Ley, R.E.; Kao-Kniffin, J. Selection on soil microbiomes reveals reproducible impacts on plant function. Isme J. 2014, 9, 980. [Google Scholar] [CrossRef]
- Syed Ab Rahman, S.F.; Singh, E.; Pieterse, C.M.J.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Jeffries, P.; Pautasso, M.; Jeger, M.J. Combined use of biocontrol agents to manage plant diseases in theory and practice. Phytopathology 2011, 101, 1024–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, C.; Moënne-Loccoz, Y.; McCarthy, J.; Higgins, P.; Powell, J.; Dowling, D.N.; O’Gara, F. Combining proteolytic and phloroglucinol-producing bacteria for improved biocontrol of Pythium-mediated damping-off of sugar beet. Plant Pathol. 1998, 47, 299–307. [Google Scholar] [CrossRef]
- Velivelli, S.L.S.; De Vos, P.; Kromann, P.; Declerck, S.; Prestwich, B.D. Biological control agents: From field to market, problems, and challenges. Trends Biotechnol. 2014, 32, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Kinnunen, M.; Dechesne, A.; Proctor, C.; Hammes, F.; Johnson, D.; Quintela-Baluja, M.; Graham, D.; Daffonchio, D.; Fodelianakis, S.; Hahn, N.; et al. A conceptual framework for invasion in microbial communities. ISME J. 2016, 10, 2773–2779. [Google Scholar] [CrossRef]
- Hultberg, M.; Alsberg, T.; Khalil, S.; Alsanius, B. Suppression of disease in tomato infected by Pythium ultimum with a biosurfactant produced by Pseudomonas koreensis. BioControl 2010, 55, 435–444. [Google Scholar] [CrossRef]
- D’aes, J.; De Maeyer, K.; Pauwelyn, E.; Höfte, M. Biosurfactants in plant–Pseudomonas interactions and their importance to biocontrol. Environ. Microbiol. Rep. 2010, 2, 359–372. [Google Scholar] [CrossRef]
- Sang, M.K.; Kim, K.D. Biocontrol activity and root colonization by Pseudomonas corrugata strains CCR04 and CCR80 against Phytophthora blight of pepper. BioControl 2014, 59, 437–448. [Google Scholar] [CrossRef]
- Pandin, C.; Le Coq, D.; Canette, A.; Aymerich, S.; Briandet, R. Should the biofilm mode of life be taken into consideration for microbial biocontrol agents? Microb. Biotechnol. 2017, 10, 719–734. [Google Scholar] [CrossRef] [Green Version]
- Larousse, M.; Galiana, E. Microbial partnerships of pathogenic oomycetes. PLoS Pathog. 2017, 13, e1006028. [Google Scholar] [CrossRef]
- Lemanceau, P.; Blouin, M.; Muller, D.; Moënne-Loccoz, Y. Let the core microbiota be functional. Trends Plant Sci. 2017, 22, 583–595. [Google Scholar] [CrossRef]



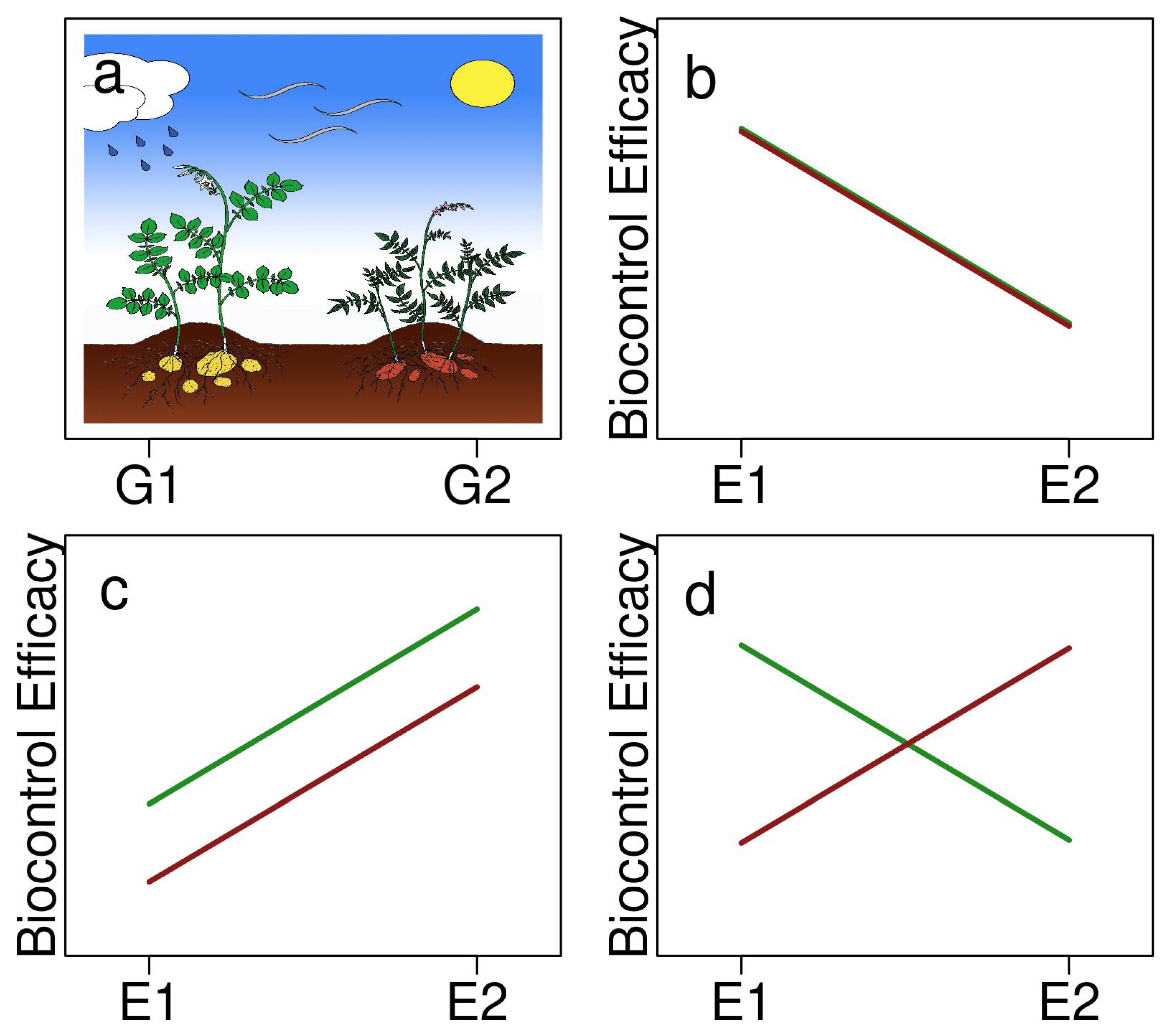{kind=link}
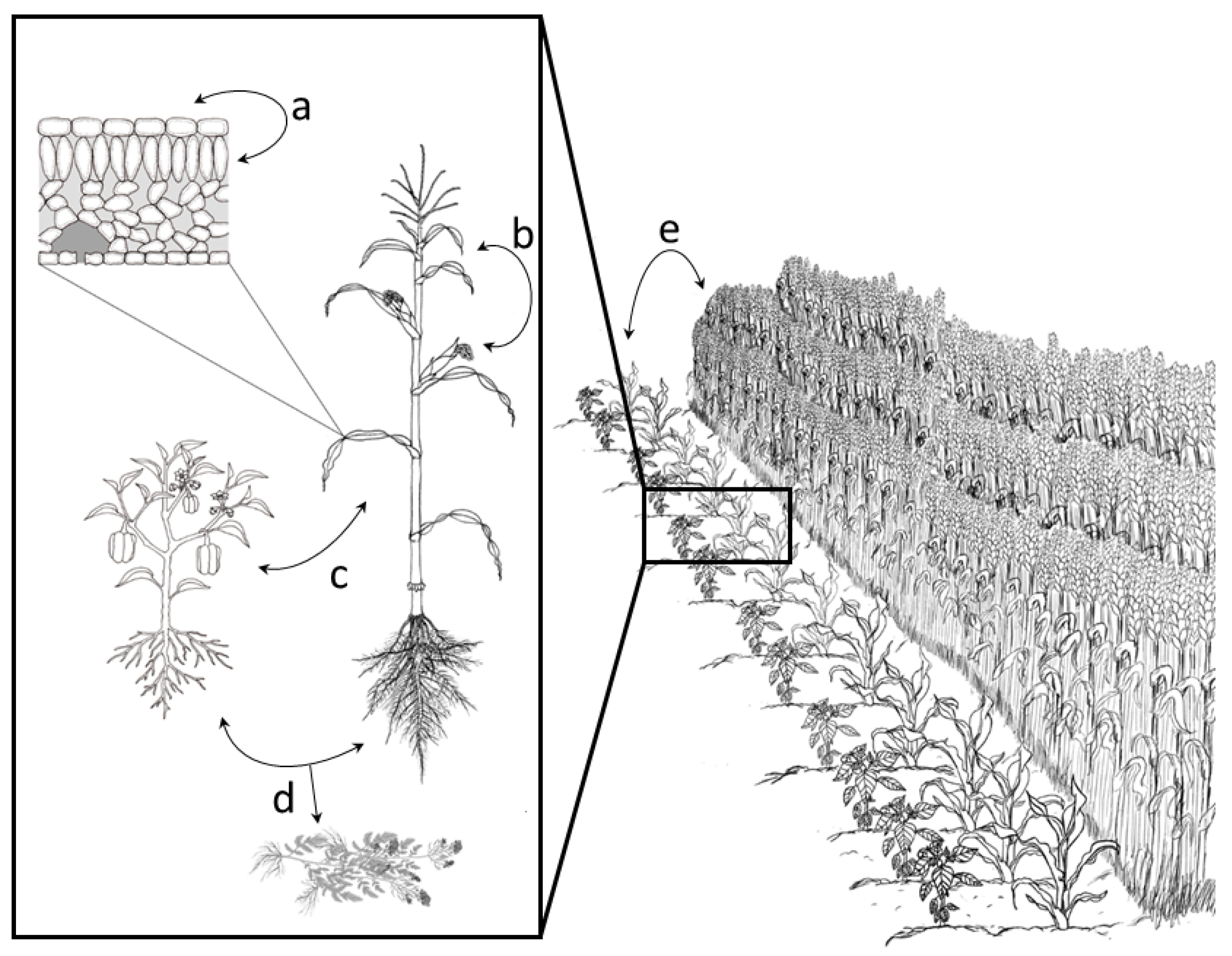{kind=link}
| Ecological Concept and/or Mechanism of Control | Disease or Disease Taxon | Crop | Experimental Context | Main Experimental Focus | Ref. |
|---|---|---|---|---|---|
| Influence of genotype × environment interactions | Root rot (Aphanomyces & Pythium spp.) | Pea (Pisum sativum L.) | In planta (growth chamber), on-farm | Screened resistance of pea genotypes in sterile or infected soil | [125] |
| Lentil (Lens spp.) | In vitro, In planta (growth chamber) | Assessed commercially available products containing Trichoderma spp. on root colonization and plant growth promotion | [77] | ||
| Seedling rot (Pythium ultimum var. ultimum) | Soybean (Glycine max L. Merr) | Field experiments | Conducted trials with four soybean genotypes across different treatment regimes in high and low disease pressure sites | [126] | |
| Apple replant disease (Phytophthora & Pythium spp.) | Apple (Malus domestica) | In planta (greenhouse) | Assessed different Brassica seed meals and rootstocks | [127] | |
| Genotype effect on induced resistance | Downy mildew of grapevine (Plasmopara viticola) | Grapevine (Vitis vinifera L.) | In planta (greenhouse) | Tested Pseudomonas fluorescens strain in two cultivars | [128] |
| Influence of environmental conditions on the microbiome and/or pathogen control measures | Foliar and crown rot (Phytophthora capsici) | Summer squash (Cucurbita pepo var. cylindrica L.) | In planta (greenhouse) | Evaluated different composts, including one Trichoderma-enriched compost | [129] |
| Apple replant disease (Phytophthora & Pythium spp.) | Apple (Malus domestica) | Sampling from orchards | Evaluated the effect of soil physical properties with seed meal amendments | [130] | |
| Various soil-borne plant oomycete pathogens | Potato (Solanum tuberosum L.) | Field experiments | Evaluated the effect of soil fertilization on fungal and oomycete pathogen- and mycorrhizal communities | [131] | |
| Aphanomycetes & Pythium spp. | Rice (Oryza sativa L.) | In planta (microcosms in 50 mL centrifuge tubes) | Assessed effects of different biochar soil amendments on relative abundance of oomycetes | [132] | |
| Induced resistance | Pythium ultimum | Romaine lettuce (Lactuca sativa L. var. longifolia) | In planta (hydroponic systems) | Evaluated treatments with Pseudomonas chlororaphis, UV irradiation and different media | [133] |
| Interaction of pathogen and biocontrol treatments | Root rot and damping-off (Pythium spp.) | Brassica microgreens: Arugula (Eruca sativa Mill.), kale (Brassica oleracea var. sabellica L.), radish (Raphanus raphanistrum subsp. sativus L.), and mustard (Brassica juncea L. Czern) microgreens | In planta (hydroponic and tray experiments) | Tested commercial products: Companion® (Bacillus subtilis GB03), Triathlon BA® (Bacillus amyloliquefaciens D747), and RootShield Plus® (Trichoderma harzianum KRL-AG2 and Trichoderma virens G-41) | [134] |
| Direct antagonism from members of microbiome | Potato late blight (Phytophthora infestans) | Potato (Solanum tuberosum L.) | In vitro, In planta (leaf disks) | Tested VOCs | [135] |
| Potato (Solanum tuberosum L.) | In planta (leaf disks) | Tested effect of sulfur-containing VOCs | [136] | ||
| Grapevine (Vitis vinifera L.) | In vitro | Screened microbial isolates for biocontrol properties | [137] | ||
| Foot rot (Phytophthora capsici) & Pythium myriotylum | Black pepper (Piper nigrum L.) | In vitro, In planta (shoot cuttings) | Used chemically synthesized volatiles from a Pseudomonas putida strain | [138] | |
| Root rot (Pythium aphanidermatum & Phytophthora capsici) | Diverse cucurbits | In vitro | Screened microbial isolates from seed endophytes | [139] | |
| Crown rot (Phytophthora capsici) | Zucchini (Cucurbita pepo L.) | Field experiments | Tested commercially available and experimental biocontrol agents and composts | [140] | |
| Root rot (Phytophthora cinnamomi & Phytophthora nicotianae) | Lavender (Lavandula angustifolia var. Hidcote), Olive (Olea europaea L) | In vitro, In planta (greenhouse trials) | Screened Trichoderma species isolated from rhizospheres (also for induced resistance) | [141] | |
| Root and/or crown rot (Phytophthora cinnamomi) | Avocado (Persea americana Mill.) | In vitro | Screened microbial isolates | [142] | |
| Strawberry (Fragaria × ananassa) plants | In vitro | Tested volitales from an Arthrobacter agilis strain | [143] | ||
| Avocado (Persea americana Mill.) | In vitro | Screening of microbial isolates from rhizosphere | [144] | ||
| Phytophthora & Pythium spp., Phytopythium vexans | Olive (Olea europaea L) | In vitro | Screened microbial isolates from root endophytes | [145] | |
| Damping off/root rot (Pythium aphanidermatum) | Cucumber (Cucumis sativus) | In vitro, In planta (seedlings) | Tested recruited microbiome from vermicomposted dairy manure | [72] | |
| Downy mildew (Sclerospora graminicola) | Pearl Millet (Cenchrus americanus L.) | In planta (germination tests and greenhouse trials) | Screened microbial isolates | [146] | |
| Alternative hosts/reservoirs of pathogens (metapopulation dynamics) | Potato late blight (Phytophthora infestans), root rot (Pythium spp.) | Potato (Solanum tuberosum L.) | In planta (detached leaf, root infection assays) | Collected oomycete communities from wild Solanum species | [147] |
| Pythium & Phytophthora spp. | Not applicable | Sampling from semi-natural and natural ecosystems | Sampled diversity and distribution of oomycetes across landscapes | [148] | |
| Spatial structure and/or microbial community dynamics | Pythium volutum, Pythium sp. F86, and Lagena radicicola | Corn (Zea mays L.) | Field experiments, In planta (pathogenicity assays with isolates of P. volutum) | Evaluated the effect of rye cover crop termination on fungal and oomycete communities | [149] |
| Root rot (Phytophthora cinnamomi) | Cranberries (Vaccinium macrocarpon Ait.) | In vitro | Screened microbial isolates for VOCs in monocultures and bacterial-fungal co-cultures | [150] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullam, K.E.; Musa, T. Ecological Dynamics and Microbial Treatments against Oomycete Plant Pathogens. Plants 2021, 10, 2697. https://doi.org/10.3390/plants10122697
Sullam KE, Musa T. Ecological Dynamics and Microbial Treatments against Oomycete Plant Pathogens. Plants. 2021; 10(12):2697. https://doi.org/10.3390/plants10122697
Chicago/Turabian StyleSullam, Karen E., and Tomke Musa. 2021. "Ecological Dynamics and Microbial Treatments against Oomycete Plant Pathogens" Plants 10, no. 12: 2697. https://doi.org/10.3390/plants10122697
APA StyleSullam, K. E., & Musa, T. (2021). Ecological Dynamics and Microbial Treatments against Oomycete Plant Pathogens. Plants, 10(12), 2697. https://doi.org/10.3390/plants10122697