
No Effect of Selective Maturation on Fruit Traits for a Bird-Dispersed Species, Sambucus racemosa


Abstract
:1. Introduction
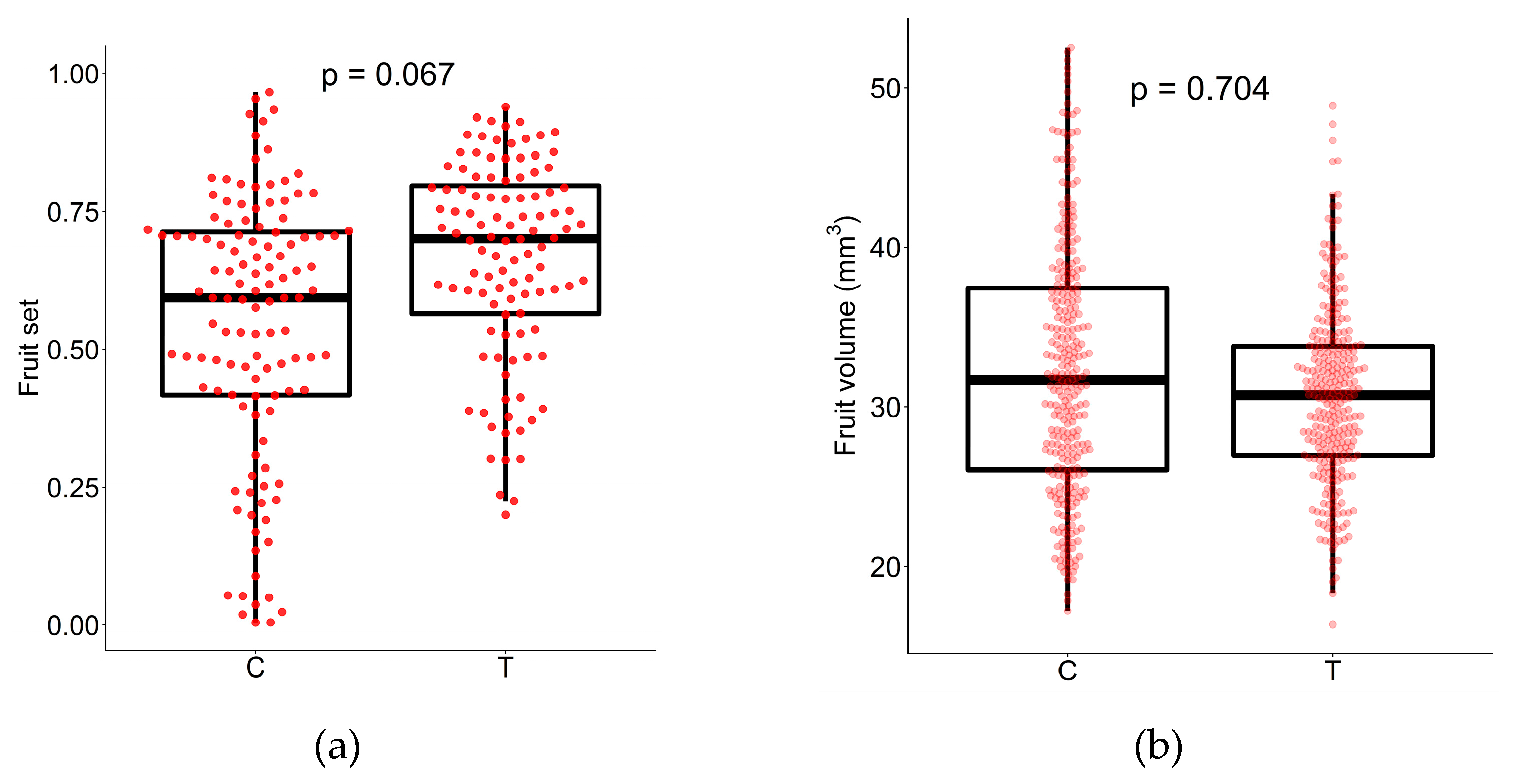
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Species and Sites
4.2. Flower Removal Experiment
4.3. Fruit Set
4.4. Fruit Size and Seed Number per Fruit
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Janzen, D.H. A Note on Optimal Mate Selection by Plants. Am. Nat. 1977, 111, 365–371. [Google Scholar] [CrossRef]
- Stephenson, A.G. Flower and fruit abortion: Proximate causes and ultimate functions. Ann. Rev. Ecol. Syst. 1981, 12, 253–279. [Google Scholar] [CrossRef]
- Lee, T.D.; Bazzaz, F.A. Regulation of fruit and seed production in an annual legume, Cassia fasciculata. Ecology 1982, 63, 1363–1373. [Google Scholar] [CrossRef]
- Collevatti, R.G.; Lopes, F.S.; Amaral, M.E.C. Reproductive success in the tropical weed Triumfetta semitriloba (Tiliaceae): Spatial and temporal variation in seed set. Rev. Biol. Trop. 1997, 1395–1399. [Google Scholar]
- Díaz, M.; Møller, A.P.; Pulido, F.J. Fruit abortion, developmental selection and developmental stability in Quercus ilex. Oecologia 2003, 135, 378–385. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Effect of selective abortion on seed germination and post-germination performance of offspring. Seed Sci. Res. 2019, 29, 210–214. [Google Scholar] [CrossRef]
- Bertin, R.I. Paternity and fruit production in trumpet creeper (Campsis radicans). Am. Nat. 1982, 119, 694–709. [Google Scholar] [CrossRef]
- Wiens, D.; Calvin, C.; Wilson, C.; Davern, C.; Frank, D.; Seavey, S.R. Reproductive success, spontaneous embryo abortion, and genetic load in flowering plants. Oecologia 1987, 71, 501–509. [Google Scholar] [CrossRef]
- Boavida, L.C.; Silva, J.P.; Feijó, J.A. Sexual reproduction in the cork oak (Quercus suber L). II. Crossing intra- and interspecific barriers. Sex. Plant Reprod. 2001, 14, 143–152. [Google Scholar] [CrossRef]
- Tsuruta, M.; Kato, S.; Mukai, Y. Timing of premature acorn abortion in Quercus serrata Thunb. is related to mating pattern, fruit size, and internal fruit development. J. For. Res. 2011, 16, 492–499. [Google Scholar] [CrossRef]
- Ezoe, H. Excessive flower production as an anti-predator strategy: When is random flower abortion favored? Popul. Ecol. 2018, 60, 275–286. [Google Scholar] [CrossRef]
- Niesenbaum, R.A. The effects of pollen load size and donor diversity on pollen performance, selective abortion, and progeny vigor in Mirabilis jalapa (Nyctaginaceae). Am. J. Bot. 1999, 86, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Pflugshaupt, K.; Kollmann, J.; Fischer, M.; Roy, B. Pollen quantity and quality affect fruit abortion in small populations of a rare fleshy-fruited shrub. Basic Appl. Ecol. 2002, 3, 319–327. [Google Scholar] [CrossRef]
- Collevatti, R.G.; Estolano, R.; Garcia, S.F.; Hay, J.D. Seed abortion in the bat pollinated Neotropical tree species, Caryocar brasiliense (Caryocaraceae). Botany 2009, 87, 1110–1115. [Google Scholar] [CrossRef]
- Collevatti, R.G.; Amara, M.E.C.; Lopes, F.S. Role of pollinators in seed set and a test of pollen limitation hypothesis in the tropical weed Triumfetta semitriloba (Tiliaceae). Rev. Biol. Trop. 1997, 1401–1407. [Google Scholar]
- Díaz, M.; Pulido, F.J.; Møller, A.P. Herbivore effects on developmental instability and fecundity of holm oaks. Oecologia 2004, 139, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, J.T. Developmental selection in vascular plants. Bot. Gaz. 1922, 73, 249–286. [Google Scholar] [CrossRef]
- Stephenson, A.G.; Winsor, J.A. Lotus corniculatus regulates offspring quality through selective fruit abortion. Evolution 1986, 40, 453–458. [Google Scholar] [CrossRef]
- Torres, C.; Eynard, M.C.; Aizen, M.A.; Galetto, L. Selective fruit maturation and seedling performance in Acacia caven (Fabaceae). Int. J. Plant Sci. 2002, 163, 809–813. [Google Scholar] [CrossRef] [Green Version]
- Guitian, J. Why Prunus mahaleb (Rosaceae) produces more flowers than fruits. Am. J. Bot. 1993, 80, 1305–1309. [Google Scholar] [CrossRef]
- Guitian, J. Selective fruit abortion in Prunus mahaleb (Rosaceae). Am. J. Bot. 1994, 81, 1555–1558. [Google Scholar] [CrossRef]
- Thompson, J.D.; Dommée, B. Sequential variation in the components of reproductive success in the distylous Jasminum fruticans (Oleaceae). Oecologia 1993, 94, 480–487. [Google Scholar] [CrossRef]
- Bookman, S.S. Evidence for selective fruit production in Asclepias. Evolution 1984, 38, 72–86. [Google Scholar] [CrossRef]
- Obeso, J.R. Selective fruit and seed maturation in Asphodelus albus Miller (Liliaceae). Oecologia 1993, 93, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Rocha, O.J.; Stephenson, A.G. Effects of nonrandom seed abortion on progeny performance in Phaseolus coccineus L. Evolution 1991, 45, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Menéndez, R.; Obeso, J.R. Effect of ovule position on seed maturation and seed weight in Ulex europaeus and Ulex gallii (Fabaceae). Can. J. Bot. 1996, 74, 848–853. [Google Scholar] [CrossRef]
- Mena-AlÍ, J.I.; Rocha, O.J. Selective seed abortion affects the performance of the offspring in Bauhinia ungulata. Ann. Bot. 2005, 95, 1017–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casper, B.B. Evidence for selective embryo abortion in Cryptantha flava. Am. Nat. 1988, 132, 318–326. [Google Scholar] [CrossRef]
- Korbecka, G.; Klinkhamer, P.G.L.; Vrieling, K. Selective embryo abortion hypothesis revisited—a molecular approach. Plant Biol. 2002, 4, 298–310. [Google Scholar] [CrossRef]
- Nakamura-Yamaguchi, A.; Kitahata, N.; Nishitani, C.; Takada, N.; Terakami, S.; Sawamura, Y.; Matsuyama, T.; Asami, T.; Nakano, T.; Saito, T.; et al. Pattern and trigger of fruit self-thinning in Japanese pears. Hortic. J. 2020, 89, 367–374. [Google Scholar] [CrossRef]
- Burd, M. Offspring quality in relation to excess flowers in Pultenaea gunnii (Fabaceae). Evolution 2004, 58, 2371–2376. [Google Scholar] [CrossRef] [PubMed]
- Burd, M. “Excess” flower production and selective fruit abortion: A model of potential benefits. Ecology 1998, 79, 2123–2132. [Google Scholar] [CrossRef]
- Lee, T.D. Patterns of fruit maturation: A gametophyte competition hypothesis. Am. Nat. 1984, 123, 427–432. [Google Scholar] [CrossRef]
- Bawa, K.S.; Webb, C.J. Flower, fruit and seed abortion in tropical forest trees: Implications for the evolution of paternal and maternal reproductive patterns. Am. J. Bot. 1984, 71, 736–751. [Google Scholar] [CrossRef]
- Howe, H.F.; Kerckhove, G.A.V. Nutmeg dispersal by tropical birds. Science 1980, 210, 925. [Google Scholar] [CrossRef]
- Sobral, M.; Larrinaga, A.R.; Guitián, J. Do seed-dispersing birds exert selection on optimal plant trait combinations? Correlated phenotypic selection on the fruit and seed size of hawthorn (Crataegus monogyna). Evol. Ecol. 2010, 24, 1277–1290. [Google Scholar] [CrossRef]
- Valenta, K.; Nevo, O. The dispersal syndrome hypothesis: How animals shaped fruit traits, and how they did not. Funct. Ecol. 2020, 34, 1158–1169. [Google Scholar] [CrossRef]
- Alcántara, J.M.; Rey, P.J. Conflicting selection pressures on seed size: Evolutionary ecology of fruit size in a bird-dispersed tree, Olea europaea. J. Evol. Biol. 2003, 16, 1168–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, C.M.; Jordano, P. Prunus mahaleb and birds: The high-efficiency seed dispersal system of a temperate fruiting tree. Ecol. Monogr. 1981, 51, 203–218. [Google Scholar] [CrossRef]
- Jordano, P.; Schupp, E.W. Seed disperser effectiveness: The quantity component and patterns of seed rain for Prunus mahaleb. Ecol. Monogr. 2000, 70, 591–615. [Google Scholar] [CrossRef]
- Guitián, J.; Guitián, P.; Medrano, M. Floral biology of the distylous Mediterranean shrub Jasminum fruticans (Oleaceae). Nord. J. Bot. 1998, 18, 195–201. [Google Scholar] [CrossRef]
- De Araujo, F.D. A review of Caryocar brasiliense (caryocaraceae)—an economically valuable species of the central brazilian cerrados. Econ. Bot. 1995, 49, 40–48. [Google Scholar] [CrossRef]
- Gómez, J.M. Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography 2003, 26, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Pulido, F.J.; Díaz, M. Regeneration of a Mediterranean oak: A whole-cycle approach. Écoscience 2016, 12, 92–102. [Google Scholar] [CrossRef]
- Masaki, T.; Nakashizuka, T.; Niiyama, K.; Tanaka, H.; Iida, S.; Bullock, J.M.; Naoe, S. Impact of the spatial uncertainty of seed dispersal on tree colonization dynamics in a temperate forest. Oikos 2019, 128, 1816–1828. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.; Swihart, R.K.; Zhao, Y.; Si, X.; Ding, P. Cascading effects of forested area and isolation on seed dispersal effectiveness of rodents on subtropical islands. J. Ecol. 2019, 107, 1506–1517. [Google Scholar] [CrossRef]
- Winsor, J.A.; Davis, L.E.; Stephenson, A.G. The relationship between pollen load and fruit maturation and the effect of pollen load on offspring vigor in Cucurbita pepo. Am. Nat. 1987, 129, 643–656. [Google Scholar] [CrossRef]
- Kistler, L.; Newsom, L.A.; Ryan, T.M.; Clarke, A.C.; Smith, B.D.; Perry, G.H. Gourds and squashes (Cucurbita spp.) adapted to megafaunal extinction and ecological anachronism through domestication. Proc. Natl. Acad. Sci. USA 2015, 112, 15107–15112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, S.B.; Tarwater, C.E. Functional traits of avian frugivores have shifted following species extinction and introduction in the Hawaiian Islands. Funct. Ecol. 2020, 34, 2467–2476. [Google Scholar] [CrossRef]
- Gómez, J.M. Bigger is not always better: Conflicting selective pressures on seed size in Quercus ilex. Evolution 2004, 58, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Botton, A.; Vizzotto, G. Fruit thinning: Advances and trends. In Hortic. Rev.; Warrington, I., Ed.; Wiley: Hboken, NJ, USA, 2018; Volume 46, pp. 185–226. [Google Scholar]
- Einhorn, T.C.; Arrington, M. ABA and Shading Induce ‘Bartlett’ Pear Abscission and Inhibit Photosynthesis but Are Not Additive. J. Plant Growth Regul. 2018, 37, 300–308. [Google Scholar] [CrossRef]
- Kundu, A.; Vadassery, J. Chlorogenic acid-mediated chemical defence of plants against insect herbivores. Plant Biol. 2019, 21, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Valtueña, F.J.; Ortega-Olivencia, A.; Rodríguez-Riaño, T. Germination and seed bank biology in some Iberian populations of Anagyris foetida L. (Leguminosae). Plant Syst. Evol. 2008, 275, 231. [Google Scholar] [CrossRef]
- Valtueña, F.J. Personal Communication; Universidad de Extremadura: Badajoz, Extremadura, Spain, 2020. [Google Scholar]
- Valtueña, F.J.; Ortega-Olivencia, A.; Rodríguez-Riaño, T. Regulation of fruit and seed set in Anagyris foetida L. (Fabaceae): The role of intrinsic factors. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2012, 146, 190–200. [Google Scholar] [CrossRef]
- Hodgson, W. Agavaceae Agave Family: Part One: Agave L. Century Pant, Maguey. J. Ariz.-Nev. Acad. Sci. 1999, 32, 1–21. [Google Scholar]
- Sutherland, S.D. Why hermaphroditic plants produce many more flowers than fruits: Experimental tests with Agave mckelveyana. Evolution 1987, 41, 750–759. [Google Scholar] [CrossRef]
- Andersson, S. No evidence for selective seed maturation in Anchusa officinalis (Boraginaceae). Oikos 1990, 57, 88–93. [Google Scholar] [CrossRef]
- Andersson, S. Personal Communication; Lund University: Lund, Sweden, 2020. [Google Scholar]
- Andersson, S. The cost of floral attractants in Achillea ptarmica (Asteraceae): Evidence from a ray removal experiment. Plant Biol. 1999, 1, 569–572. [Google Scholar] [CrossRef]
- Andersson, S. The potential for selective seed maturation in Achillea ptarmica (Asteraceae). Oikos 1993, 66, 36–42. [Google Scholar] [CrossRef]
- Bookman, S.S. Costs and benefits of flower abscission and fruit abortion in Asclepias speciosa. Am. J. Bot. 1983, 70, 897–905. [Google Scholar] [CrossRef]
- Collevatti, R.G.; Grattapaglia, D.; Hay, J.D. High resolution microsatellite based analysis of the mating system allows the detection of significant biparental inbreeding in Caryocar brasiliense, an endangered tropical tree species. Heredity 2001, 86, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.D. Effects of seed number per fruit on seed dispersal in Cassia fasciculata (Caesalpiniaceae). Bot. Gaz. 1984, 145, 136–139. [Google Scholar] [CrossRef]
- Chachalis, D.; Reddy, K.N. Factors affecting Campsis radicans seed germination and seedling emergence. Weed Sci. 2000, 48, 212–216. [Google Scholar] [CrossRef]
- Bawa, K.S.; Frankie, G.W. Cochlospermum vitifolium (Poro-poro, Cochlospermum, Silk tree, cotton tree). In Costa Rican natural history; Janzen, D.H., Ed.; University of Chicago Press: Chicago, IL, USA, 1983; pp. 215–216. [Google Scholar]
- Berardi, A.E.; Frey, F.M.; Denton, E.M.; Wells, J.H. Betalain color morphs exhibit differential growth, defensive ability, and pollen tube growth rates in Mirabilis jalapa (Nyctaginaceae). Int. J. Plant Sci. 2013, 174, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Casper, B.B. Spatial patterns of seed dispersal and postdispersal seed predation of Cryptantha flava (Boraginaceae). Am. J. Bot. 1987, 74, 1646–1655. [Google Scholar] [CrossRef]
- Cortés-Flores, J.; Andresen, E.; Cornejo-Tenorio, G.; Ibarra-Manríquez, G. Fruiting phenology of seed dispersal syndromes in a Mexican Neotropical temperate forest. For. Ecol. Manag. 2013, 289, 445–454. [Google Scholar] [CrossRef]
- de Kok, R.P.J.; West, J.G. A revision of the genus Pultenaea (Fabaceae). 3. The eastern species with recurved leaves. Aust. Syst. Bot. 2004, 17, 273–326. [Google Scholar] [CrossRef]
- Morimoto, J.; Kominami, R.; Koike, T. Distribution and characteristics of the soil seed bank of the black locust (Robinia pseudoacacia) in a headwater basin in northern Japan. Landsc. Ecol. Eng. 2010, 6, 193–199. [Google Scholar] [CrossRef]
- Susko, D.J. Effect of ovule position on patterns of seed maturation and abortion in Robinia pseudoacacia (Fabaceae). Can. J. Bot. 2006, 84, 1259–1265. [Google Scholar] [CrossRef]
- Tybirk, K. Pollination, breeding system and seed abortion in some African acacias. Bot. J. Linn. Soc. 1993, 112, 107–137. [Google Scholar] [CrossRef]
- Jara Guerrero, A.K. Ecología de la dispersión de plantas en los bosques secos del suroccidente Ecuatoriano. PhD Dissertation, Universidad Politécnica de Madrid, Madrid, Spain, 2014. [Google Scholar]
- Francis, J.K. Wildland Shrubs of the United States and Its Territories: Thamnic Descriptions, Volume 1.; U.S. Department of Agriculture, Forest Service: Fort Collins, CO, USA, 2004.
- Couturier, E. Folded isometric deformations and banana-shaped seedpod. Proc. R. Soc. A Math. Phys. Eng. Sci. 2016, 472, 20150760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Ann. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Inoue, T.; Kato, M.; Kakutani, T.; Suka, T.; Itino, T. Insect-flower relationship in the temperate deciduous forest of Kibune, Kyoto: An overview of the flowering phenology and the seasonal pattern of insect visits. Contrib. Biol. Lab. Kyoto Univ. 1990, 27, 377–463. [Google Scholar]
- Traveset, A.; Willson, M.F. Effect of birds and bears on seed germination of fleshy-fruited plants in temperate rainforests of southeast Alaska. Oikos 1997, 80, 89–95. [Google Scholar] [CrossRef]
- Burns, K.C. A simple null model predicts fruit-frugivore interactions in a temperate rainforest. Oikos 2006, 115, 427–432. [Google Scholar] [CrossRef]
- Fujimaki, Y. Food habit of hazel grouse in Hokkaido, Japan. J. Yamashina Inst. Ornithol. 2002, 34, 73–79. [Google Scholar] [CrossRef]
- Japan Meteorological Agency. Available online: https://www.jma.go.jp (accessed on 2 March 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Clarke, E.; Sherrill-Mix, S. Ggbeeswarm: Categorical Scatter (Violin Point) Plots. 2017. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Koyama, K.; Shirakawa, H.; Kikuzawa, K. Redeployment of shoots into better-lit positions within the crowns of saplings of five species with different growth patterns. Forests 2020, 11, 1301. [Google Scholar] [CrossRef]
- Barr, D.J.; Levy, R.; Scheepers, C.; Tily, H.J. Random effects structure for confirmatory hypothesis testing: Keep it maximal. J. Mem. Lang. 2013, 68, 255–278. [Google Scholar] [CrossRef] [Green Version]
- Deguchi, R.; Koyama, K. Photosynthetic and morphological acclimation to high and low light environments in Petasites japonicus subsp. giganteus. Forests 2020, 11, 1365. [Google Scholar] [CrossRef]


{kind=link}
| Number of Seeds Within Each Fruit | Fruit Counts | Relative Frequency (%) | ||
|---|---|---|---|---|
| Control | Treatment | Control | Treatment | |
| 2 | 40 | 10 | 3.44% | 0.93% |
| 3 | 1116 | 1066 | 96.04% | 98.70% |
| 4 | 6 | 4 | 0.52% | 0.37% |
| Total | 1162 | 1080 | 100% | 100% |
| Mean seed number per fruit | 2.97 | 2.99 | ||
| Sp. | Fruit Type | Type of Primary Dispersal 1 | Hand- Removal Performed? | Wider Choice 3 | Seed Num. per FRUIT | Fruit Size | Seed Size 2 | Offspring Vigor | Ref. 4 |
|---|---|---|---|---|---|---|---|---|---|
| Aa | Capsule | Self 5 | Flowers and fruits | + | [24] | ||||
| Af | Legume [54] | Self [54,55] | Flowers | Mixed results | ± | ± | [56] | ||
| Am | Capsule [57] | Flowers | No | 0 | [58] | ||||
| Ao | Nutlet [59] | Self [60] | Ovules | No | – | [59] | |||
| Ap | Achene [61] | Self [60] | Flowers | No | 0 Fixed 6 [61] | 0 | [62] | ||
| As | Legume | Wind [63] | No | + | Increased germination rate | [23] | |||
| Bu | Legume | Self [27] | Fruits | + | + | Increased germination rate | [27] | ||
| Cb | Drupe [42] | Gravity, with possible dispersal by large birds [64] | No | [14] | |||||
| Cf | Legume [65] | Self [65] | Flowers | + | + | [3] | |||
| Cp | Pepo [48] | May not exist in the wild [48] | No | + | Rapid germination, increased seedling biomass | [47] | |||
| Cr | Capsule [66] | Wind [7,66] | No | + | + | [7] | |||
| Cv | Capsule | Wind [67] | No | + | [34] | ||||
| Jf | Berry [41] | Fruits | Mixed results | [22] | |||||
| Lc | Legume | Self | Flowers | Yes | + | Increased growth rate | [18] | ||
| Mj | Capsule [68] | No | 0 Fixed 6 | [12] | |||||
| Of | Nutlet [69] | Wind [69] | Ovules | Increased germination rate, but no change in survival rate | [28] | ||||
| Pc | Legume | Self [70] | Ovules | Rapid germination, increased growth, survival, and reproductive output | [25] | ||||
| Pg | Legume [71] | Self (primary) | No | 0 Fixed 6 | [31] | ||||
| Pm | Drupe | Birds and other animals [40] | Flowers | 0 Fixed 6 | + | + | [20,21] | ||
| Qi | Acorn | Gravity (primary), animals (secondary) [43,44] | No | 0 Acorn | Change neither in germination nor growth rates, but a difference in resource allocation | [5] | |||
| Qs | Acorn | Gravity (primary) [45] animals (secondary) [46] | No | 0 Acorn | [10] | ||||
| Rs | Legume | Wind, water [72] | No | + | Increased seedling size | [73] | |||
| Sa | Legume | Wind | [74] | ||||||
| Sp | Legume | Wind | [74] | ||||||
| Sr | Drupe | Birds and other animals | Flowers | Yes | 0 | 0 | This study | ||
| Ss | Legume | Wind | [74] | ||||||
| Ts | Capsule [75] | Epizoochory [76] | + | [15] | |||||
| Ue | Legume | Self (primary) | No | [26] | |||||
| Ug | Legume | Self (primary) | No | [26] | |||||
| Vc | Legume [77] | Self [77] | Flowers | 0 | 0 | 0 | Increased germination rate and seedling survival rate | [19] |
| Treatment (Inflorescences 50% Thinned) | Control | |
|---|---|---|
| Total number of trees investigated | 12 | 12 |
| Median tree height (m) | 2.98 | 2.69 |
| Range of tree height (m) | 1.70–3.83 | 1.75–3.85 |
| Total number of infructescences sampled | 108 | 120 |
| Total number of fruits counted for the calculation of the fruit set | 16,998 | 15,036 |
| Total number of fruit scars on the infructescences counted for the calculation of the fruit set | 7544 | 11,626 |
| Total number of fruits investigated for counting seed number per fruit | 1080 | 1162 |
| Total number of trees for which fruit sizes were measured (only for the Forest site) | 3 | 3 |
| Total number of infructescences used for size measurement | 30 | 30 |
| Total number of fruits used for size measurement | 300 | 300 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koyama, K.; Tashiro, M. No Effect of Selective Maturation on Fruit Traits for a Bird-Dispersed Species, Sambucus racemosa. Plants 2021, 10, 376. https://doi.org/10.3390/plants10020376
Koyama K, Tashiro M. No Effect of Selective Maturation on Fruit Traits for a Bird-Dispersed Species, Sambucus racemosa. Plants. 2021; 10(2):376. https://doi.org/10.3390/plants10020376
Chicago/Turabian StyleKoyama, Kohei, and Mayu Tashiro. 2021. "No Effect of Selective Maturation on Fruit Traits for a Bird-Dispersed Species, Sambucus racemosa" Plants 10, no. 2: 376. https://doi.org/10.3390/plants10020376