
Ectopic Expression of OLEOSIN 1 and Inactivation of GBSS1 Have a Synergistic Effect on Oil Accumulation in Plant Leaves


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
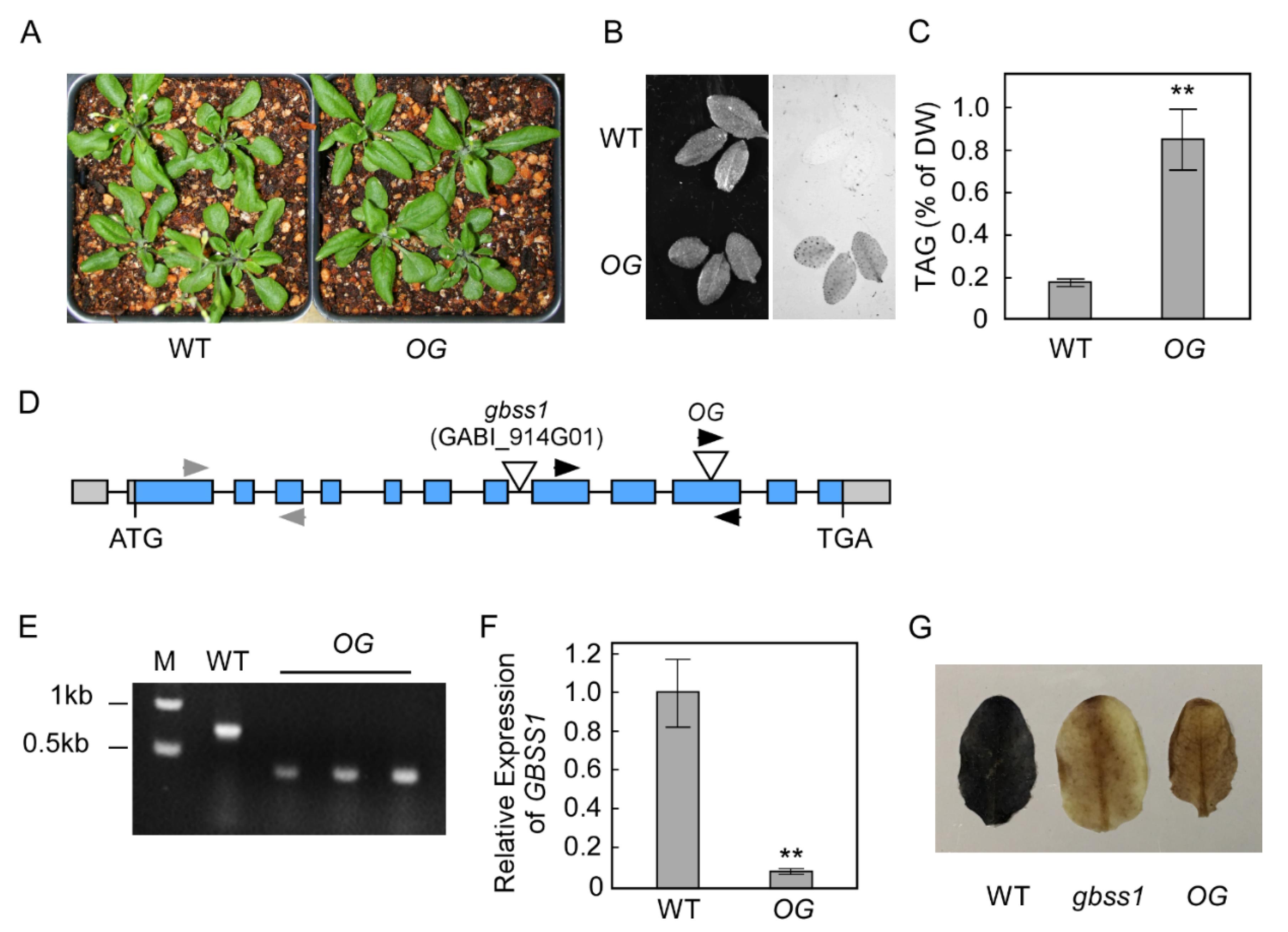
2.1. An Arabidopsis OLE1-GFP Transgenic Line with a T-DNA Insertion in GBSS1 Accumulated Significant Leaf TAG
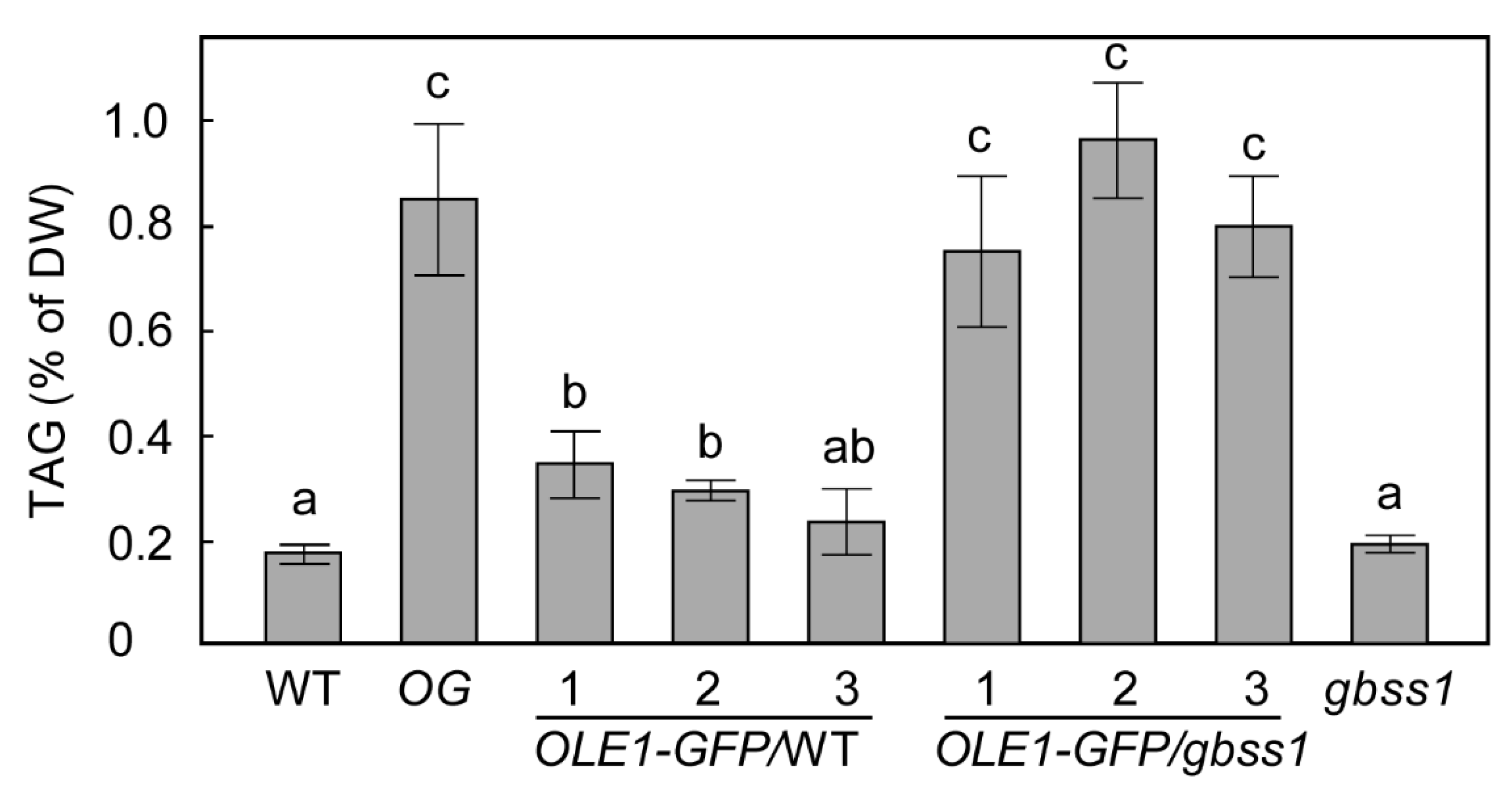
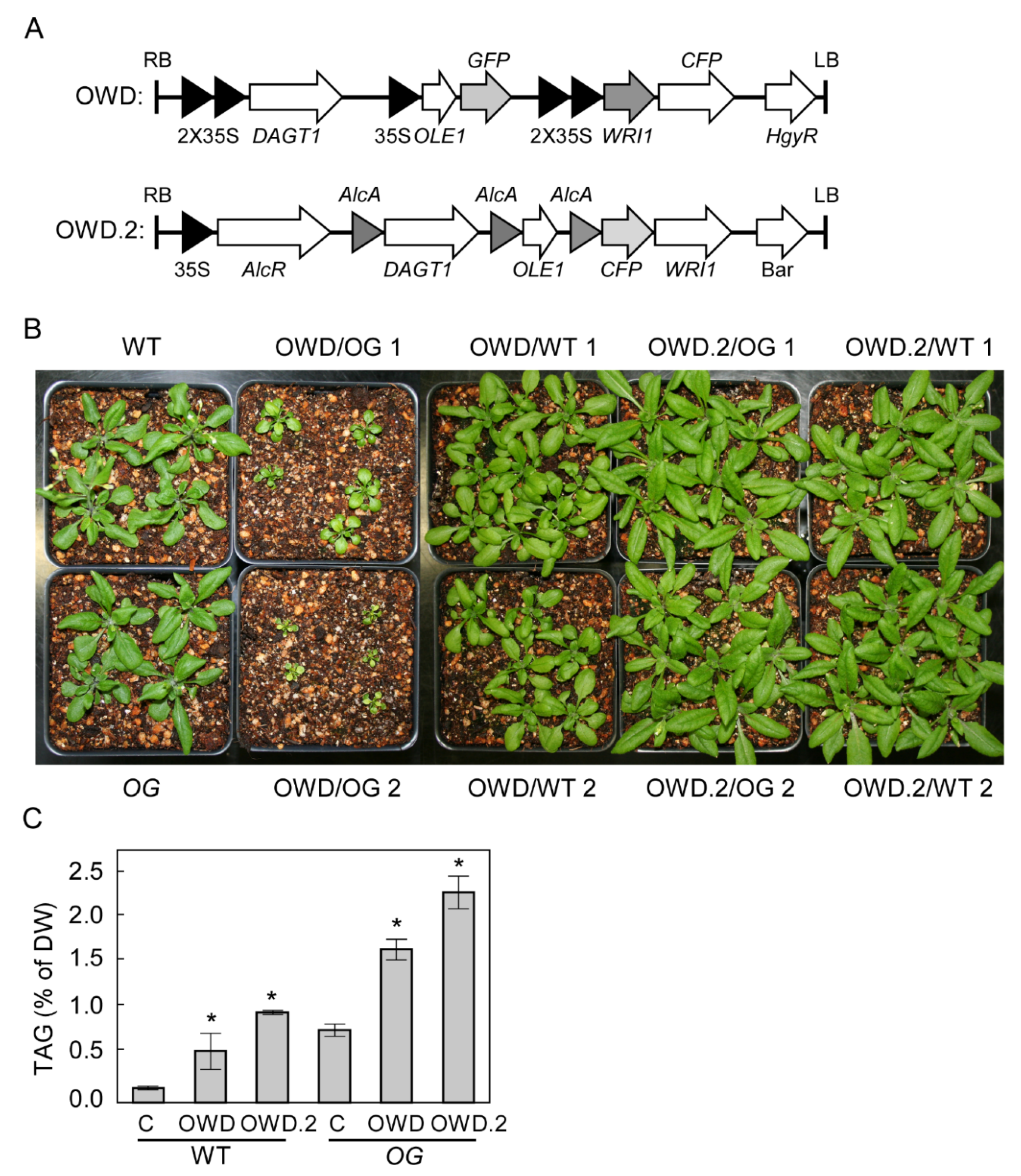
2.2. Overexpression of OLE1 in a gbss1 Background Resulted in Higher TAG Accumulation Than in WT
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Genetic Constructs
4.3. Preparation of Leaf Starch Granules and Iodine Staining
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horn, P.J.; Benning, C. The plant lipidome in human and environmental health. Science 2016, 353, 1228–1232. [Google Scholar] [CrossRef] [PubMed]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957. [Google Scholar]
- Rawsthorne, S. Carbon flux and fatty acid synthesis in plants. Progress Lipid Res. 2002, 41, 182–196. [Google Scholar] [CrossRef]
- Zhou, X.-R.; Bhandari, S.; Johnson, B.S.; Kotapati, H.K.; Allen, D.K.; Vanhercke, T.; Bates, P.D. Reorganization of Acyl Flux through the Lipid Metabolic Network in Oil-Accumulating Tobacco Leaves. Plant Physiol. 2020, 182, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, S.; Kannan, B.; Karan, R.; Sanahuja, G.; Liu, H.; Garcia-Ruiz, E.; Kumar, D.; Singh, V.; Zhao, H.; Long, S.; et al. Towards oilcane: Engineering hyperaccumulation of triacylglycerol into sugarcane stems. GCB Bioenergy 2020, 12, 476–490. [Google Scholar] [CrossRef]
- Vanhercke, T.; El Tahchy, A.; Liu, Q.; Zhou, X.R.; Shrestha, P.; Divi, U.K.; Ral, J.P.; Mansour, M.P.; Nichols, P.D.; James, C.N.; et al. Metabolic engineering of biomass for high energy density: Oilseed-like triacylglycerol yields from plant leaves. Plant Biotechnol. J. 2014, 12, 231–239. [Google Scholar] [CrossRef]
- Focks, N.; Benning, C. wrinkled1: A novel, low-seed-oil mutant of Arabidopsis with a deficiency in the seed-specific regulation of carbohydrate metabolism. Plant Physiol. 1998, 118, 91–101. [Google Scholar] [CrossRef]
- Cernac, A.; Benning, C. WRINKLED1 encodes an AP2/EREB domain protein involved in the control of storage compound biosynthesis in Arabidopsis. Plant J. 2004, 40, 575–585. [Google Scholar] [CrossRef]
- Zou, J.; Wei, Y.; Jako, C.; Kumar, A.; Selvaraj, G.; Taylor, D.C. The Arabidopsis thaliana TAG1 mutant has a mutation in a diacylglycerol acyltransferase gene. Plant J. 1999, 19, 645–653. [Google Scholar] [CrossRef]
- Siloto, R.M.; Findlay, K.; Lopez-Villalobos, A.; Yeung, E.C.; Nykiforuk, C.L.; Moloney, M.M. The accumulation of oleosins determines the size of seed oilbodies in Arabidopsis. Plant Cell 2006, 18, 1961–1974. [Google Scholar] [CrossRef]
- Sanjaya; Durrett, T.P.; Weise, S.E.; Benning, C. Increasing the energy density of vegetative tissues by diverting carbon from starch to oil biosynthesis in transgenic Arabidopsis. Plant Biotechnol. J. 2011, 9, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Zolman, B.K.; Silva, I.D.; Bartel, B. The Arabidopsis pxa1 Mutant Is Defective in an ATP-Binding Cassette Transporter-Like Protein Required for Peroxisomal Fatty Acid β-Oxidation. Plant Physiol. 2001, 127, 1266–1278. [Google Scholar] [CrossRef]
- Zale, J.; Jung, J.H.; Kim, J.Y.; Pathak, B.; Karan, R.; Liu, H.; Chen, X.; Wu, H.; Candreva, J.; Zhai, Z.; et al. Metabolic engineering of sugarcane to accumulate energy-dense triacylglycerols in vegetative biomass. Plant Biotechnol. J. 2016, 14, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.A.; van Erp, H.; Quettier, A.-L.; Shaw, E.; Menard, G.; Kurup, S.; Eastmond, P.J. The Sugar-dependent1 Lipase limits Triacylglycerol accumulation in vegetative tissues of Arabidopsis. Plant Physiol. 2013, 162, 1282–1289. [Google Scholar] [CrossRef]
- Gottwald, J.R.; Krysan, P.J.; Young, J.C.; Evert, R.F.; Sussman, M.R. Genetic evidence for the in planta role of phloem-specific plasma membrane sucrose transporters. Proc. Nat. Acad. Sci. USA 2000, 97, 13979–13984. [Google Scholar] [CrossRef]
- Zhai, Z.; Liu, H.; Xu, C.; Shanklin, J. Sugar Potentiation of Fatty Acid and Triacylglycerol Accumulation. Plant Physiol. 2017, 175, 696–707. [Google Scholar] [CrossRef]
- Martin, C.; Smith, A.M. Starch biosynthesis. Plant Cell 1995, 7, 971. [Google Scholar] [PubMed]
- Buleon, A.; Colonna, P.; Planchot, V.; Ball, S. Starch granules: Structure and biosynthesis. Int. J. Biol. Macromol. 1998, 23, 85–112. [Google Scholar] [CrossRef]
- Galliard, T. Starch: Properties and Potential; John Wiley & Sons Incorporated: Hoboken, NJ, USA, 1987. [Google Scholar]
- Shewry, P.R.; Stobart, K. Seed Storage Compounds: Biosynthesis, Interactions, and Manipulation; Clarendon Press Oxford: Oxford, UK, 1993. [Google Scholar]
- Winichayakul, S.; Scott, R.W.; Roldan, M.; Hatier, J.H.; Livingston, S.; Cookson, R.; Curran, A.C.; Roberts, N.J. In vivo packaging of triacylglycerols enhances Arabidopsis leaf biomass and energy density. Plant Physiol. 2013, 162, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yan, C.; Zhang, X.; Xu, C. Dual role for phospholipid: Diacylglycerol acyltransferase: Enhancing fatty acid synthesis and diverting fatty acids from membrane lipids to triacylglycerol in Arabidopsis leaves. Plant Cell Online 2013, 25, 3506–3518. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, L.; Sun, X.; Tang, K. An oleosin-fusion protein driven by the CaMV35S promoter is accumulated in Arabidopsis (Brassicaceae) seeds and correctly targeted to oil bodies. Genet. Mol. Res. 2012, 11, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Seung, D.; Soyk, S.; Coiro, M.; Maier, B.A.; Eicke, S.; Zeeman, S.C. Protein Targeting to Starch is required for localising granule-bound starch synthase to starch granules and for normal amylose synthesis in Arabidopsis. PLoS Biol. 2015, 13, e1002080. [Google Scholar] [CrossRef]
- Streb, S.; Zeeman, S.C. Starch metabolism in Arabidopsis. Am. Soc. Plant Biol. 2012, 10, e0160. [Google Scholar] [CrossRef] [PubMed]
- Ekman, Å.; Hayden, D.M.; Dehesh, K.; Bülow, L.; Stymne, S. Carbon partitioning between oil and carbohydrates in developing oat (Avena sativa L.) seeds. J. Exp. Bot. 2008, 59, 4247–4257. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Fan, J.; Froehlich, J.E.; Awai, K.; Benning, C. Mutation of the TGD1 chloroplast envelope protein affects phosphatidate metabolism in Arabidopsis. Plant Cell Online 2005, 17, 3094–3110. [Google Scholar] [CrossRef]
- Zhang, M.; Fan, J.; Taylor, D.C.; Ohlrogge, J.B. DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 2009, 21, 3885–3901. [Google Scholar] [CrossRef]
- Bouvier-Nave, P.; Benveniste, P.; Noiriel, A.; Schaller, H. Expression in yeast of an acyl-CoA:diacylglycerol acyltransferase cDNA from Caenorhabditis elegans. Biochem. Soc. Trans. 2000, 28, 692–695. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vanhercke, T.; El Tahchy, A.; Shrestha, P.; Zhou, X.R.; Singh, S.P.; Petrie, J.R. Synergistic effect of WRI1 and DGAT1 coexpression on triacylglycerol biosynthesis in plants. FEBS Lett. 2013, 587, 364–369. [Google Scholar] [CrossRef]
- Roslan, H.A.; Salter, M.G.; Wood, C.D.; White, M.R.; Croft, K.P.; Robson, F.; Coupland, G.; Doonan, J.; Laufs, P.; Tomsett, A.B. Characterization of the ethanol-inducible alc gene-expression system in Arabidopsis thaliana. Plant J. 2001, 28, 225–235. [Google Scholar] [CrossRef]
- Liu, Y.-G.; Whittier, R.F. Thermal asymmetric interlaced PCR: Automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking. Genomics 1995, 25, 674–681. [Google Scholar] [CrossRef]
- Zhong, S.; Lin, Z.; Fray, R.G.; Grierson, D. Improved plant transformation vectors for fluorescent protein tagging. Transgenic Res. 2008, 17, 985–989. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nakagawa, T.; Suzuki, T.; Murata, S.; Nakamura, S.; Hino, T.; Maeo, K.; Tabata, R.; Kawai, T.; Tanaka, K.; Niwa, Y. Improved Gateway binary vectors: High-performance vectors for creation of fusion constructs in transgenic analysis of plants. Biosci. Biotechnol. Biochem. 2007, 71, 2095–2100. [Google Scholar] [CrossRef]
- Yan, P.; Shen, W.; Gao, X.; Li, X.; Zhou, P.; Duan, J. High-throughput construction of intron-containing hairpin RNA vectors for RNAi in plants. PLoS ONE 2012, 7, e38186. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Liu, H.; Shanklin, J. Phosphorylation of WRINKLED1 by KIN10 Results in Its Proteasomal Degradation, Providing a Link between Energy Homeostasis and Lipid Biosynthesis. Plant Cell 2017, 29, 871–889. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Northrop, F.; Smith, A.M.; Rees, T. A starch-accumulating mutant of Arabidopsis thaliana deficient in a chloroplastic starch-hydrolysing enzyme. Plant J. 1998, 15, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Schütze, K.; Harter, K.; Chaban, C. Bimolecular fluorescence complementation (BiFC) to study protein-protein interactions in living plant cells. In Plant Signal Transduction; Springer: Berlin/Heidelberg, Germany, 2009; pp. 189–202. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, Z.; Liu, H.; Shanklin, J. Ectopic Expression of OLEOSIN 1 and Inactivation of GBSS1 Have a Synergistic Effect on Oil Accumulation in Plant Leaves. Plants 2021, 10, 513. https://doi.org/10.3390/plants10030513
Zhai Z, Liu H, Shanklin J. Ectopic Expression of OLEOSIN 1 and Inactivation of GBSS1 Have a Synergistic Effect on Oil Accumulation in Plant Leaves. Plants. 2021; 10(3):513. https://doi.org/10.3390/plants10030513
Chicago/Turabian StyleZhai, Zhiyang, Hui Liu, and John Shanklin. 2021. "Ectopic Expression of OLEOSIN 1 and Inactivation of GBSS1 Have a Synergistic Effect on Oil Accumulation in Plant Leaves" Plants 10, no. 3: 513. https://doi.org/10.3390/plants10030513
APA StyleZhai, Z., Liu, H., & Shanklin, J. (2021). Ectopic Expression of OLEOSIN 1 and Inactivation of GBSS1 Have a Synergistic Effect on Oil Accumulation in Plant Leaves. Plants, 10(3), 513. https://doi.org/10.3390/plants10030513