
Differential Salt Tolerance Strategies in Three Halophytes from the Same Ecological Habitat: Augmentation of Antioxidant Enzymes and Compounds

 , and
, and
Abstract
:1. Introduction
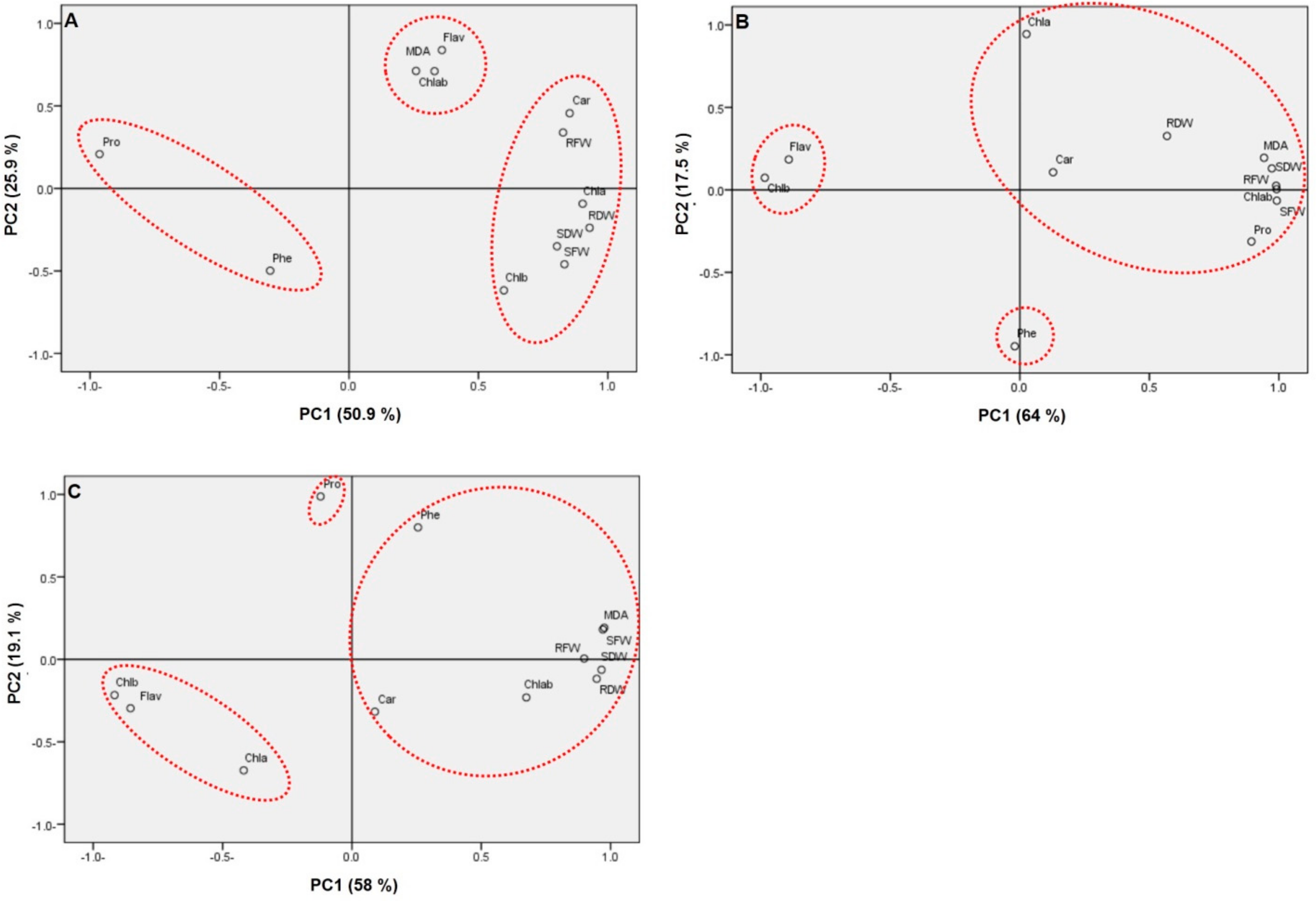
2. Results
2.1. Effect of Salinity on Na+, K+ and Ca2+ Contents
2.2. Effect of Salinity on Growth Parameters
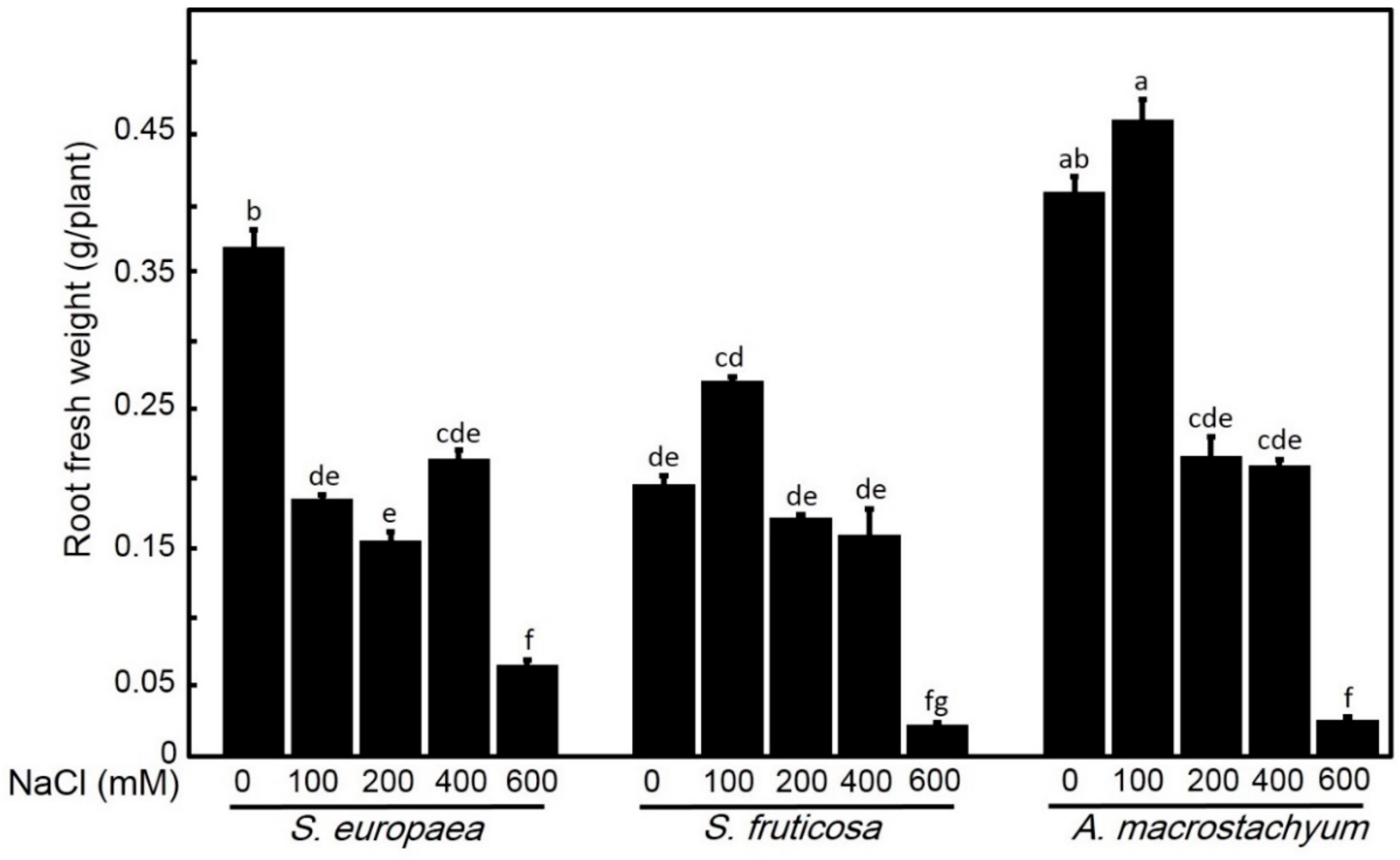
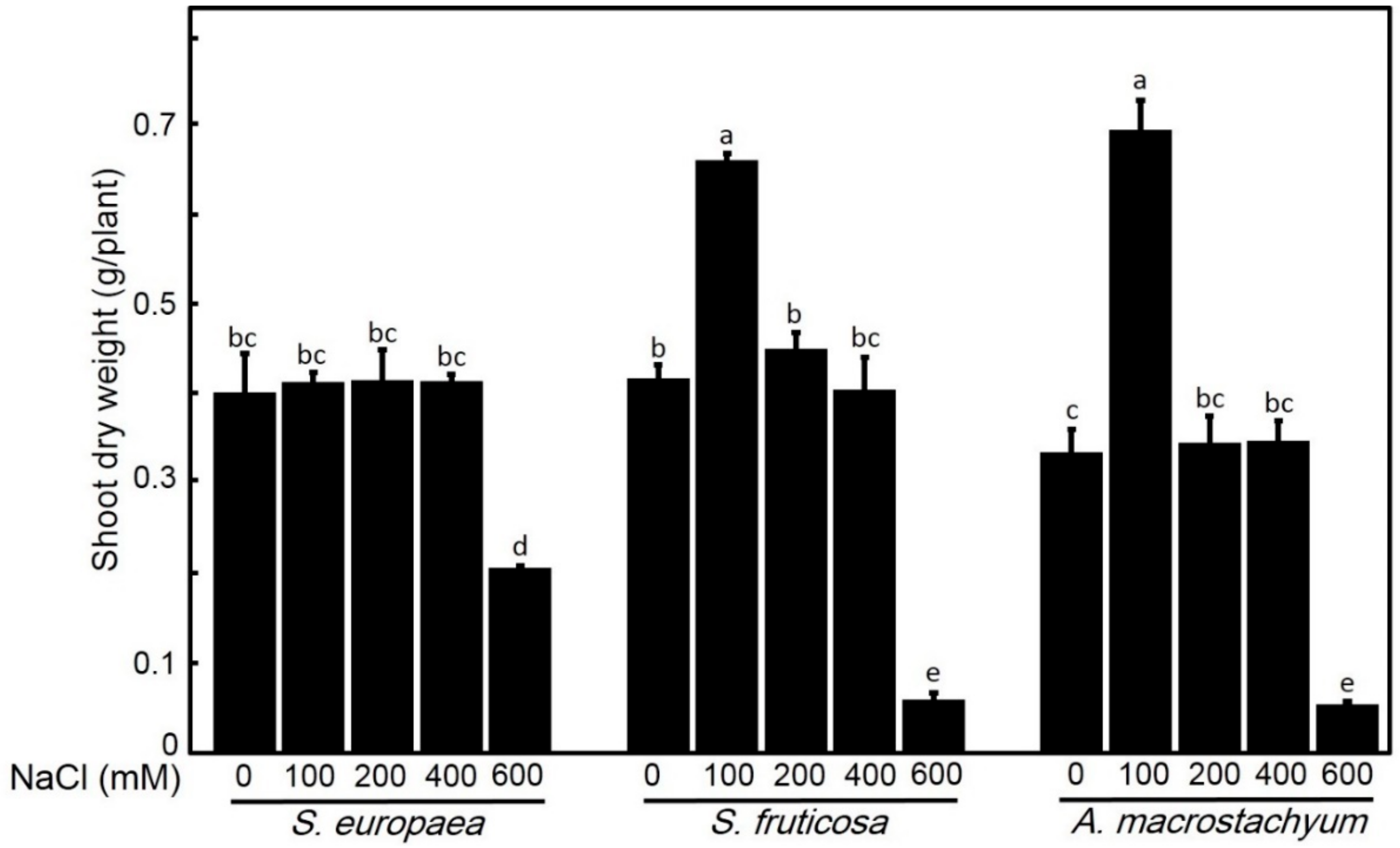
2.2.1. Effect of Salinity on Biomass Production
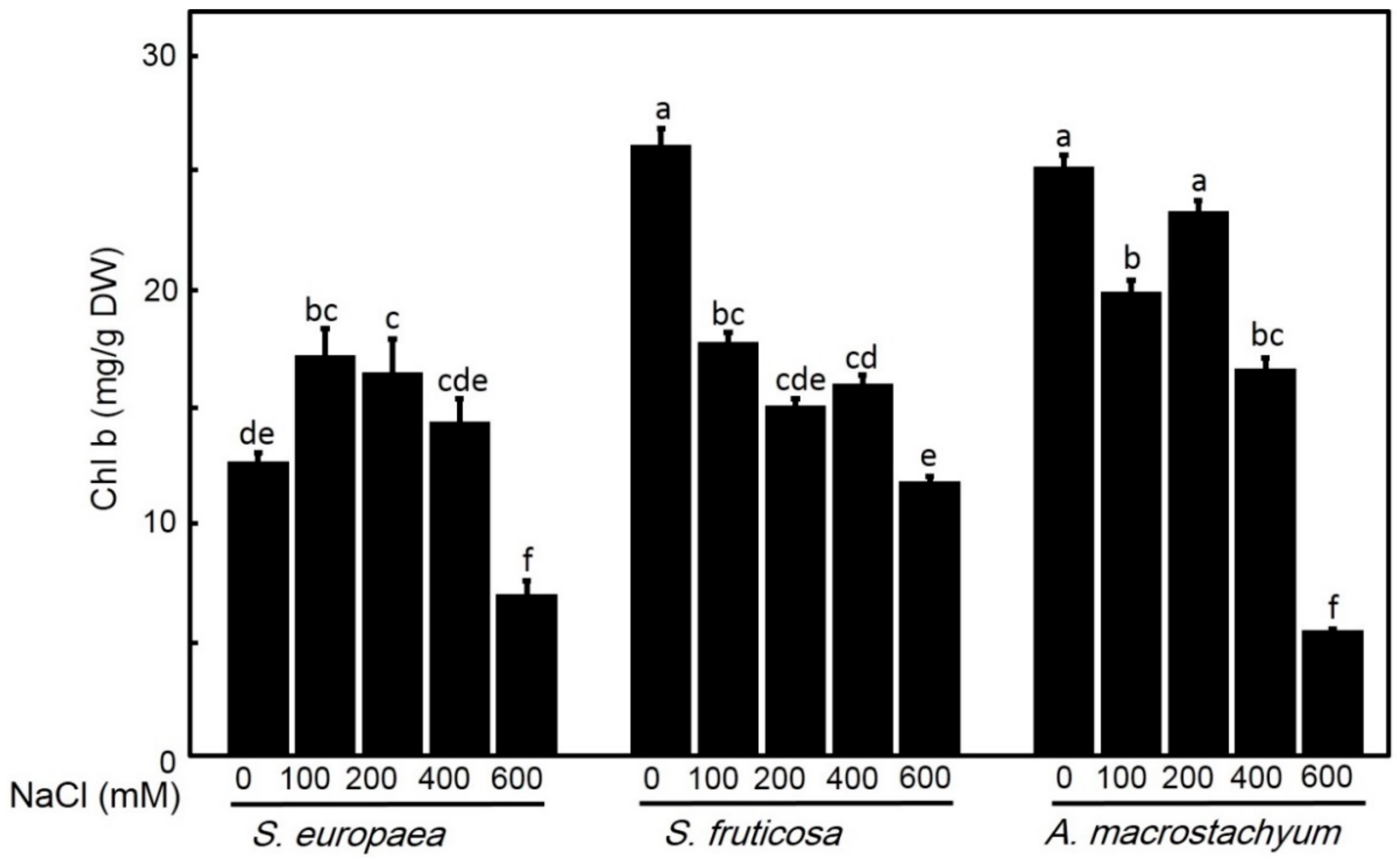
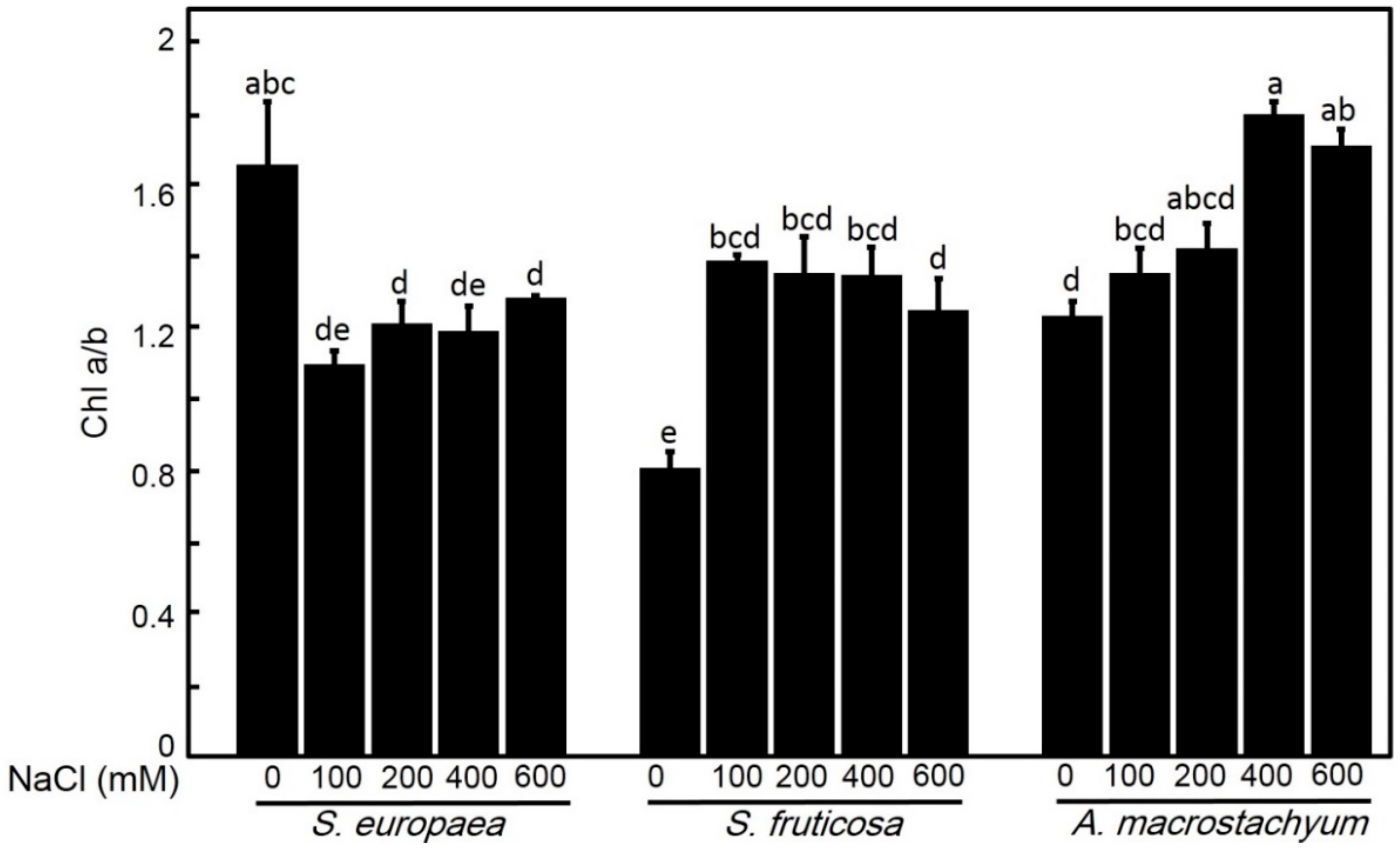
2.2.2. Effect of Salinity on Chlorophyll Contents
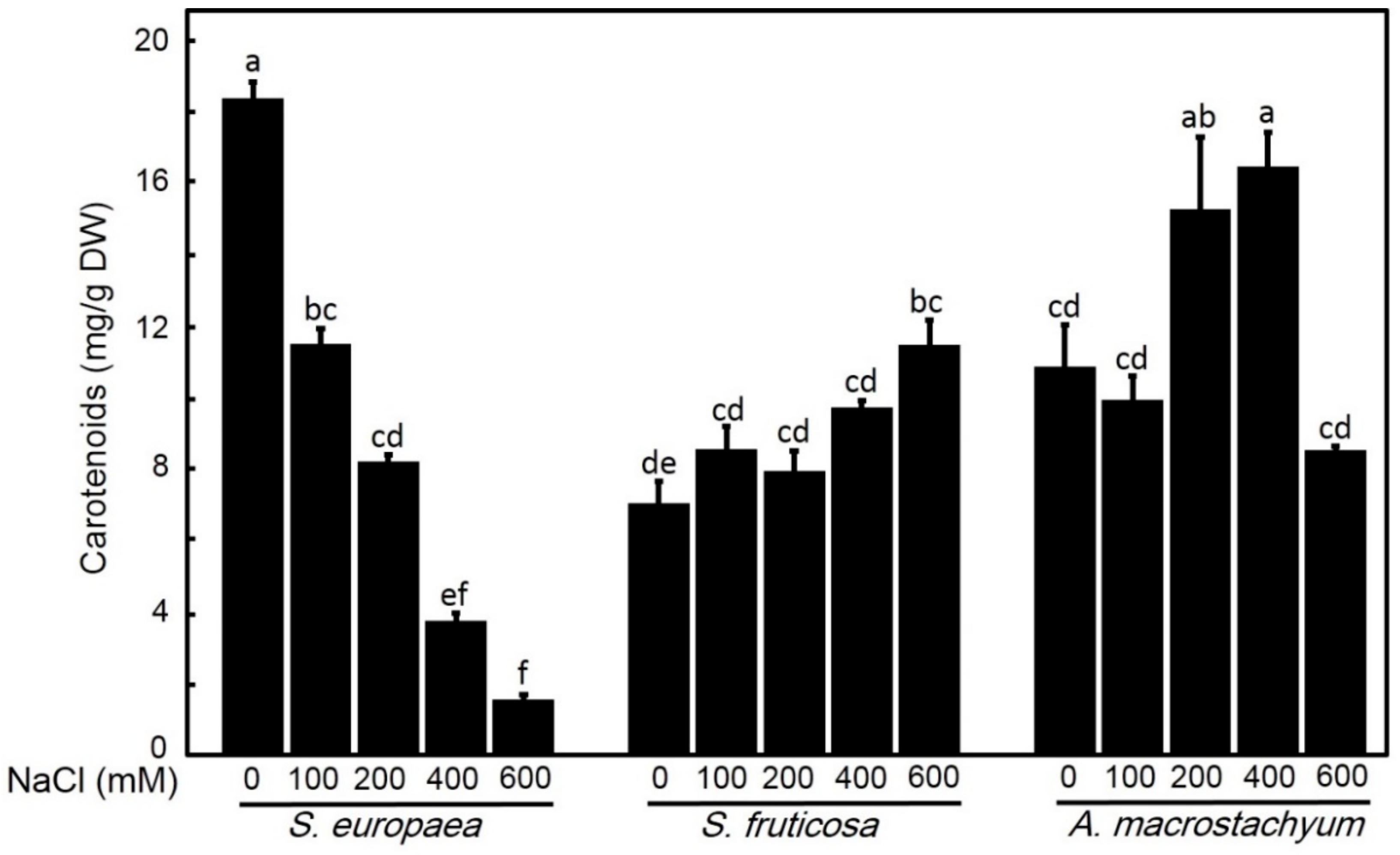
2.2.3. Effect of Salinity on Carotenoids
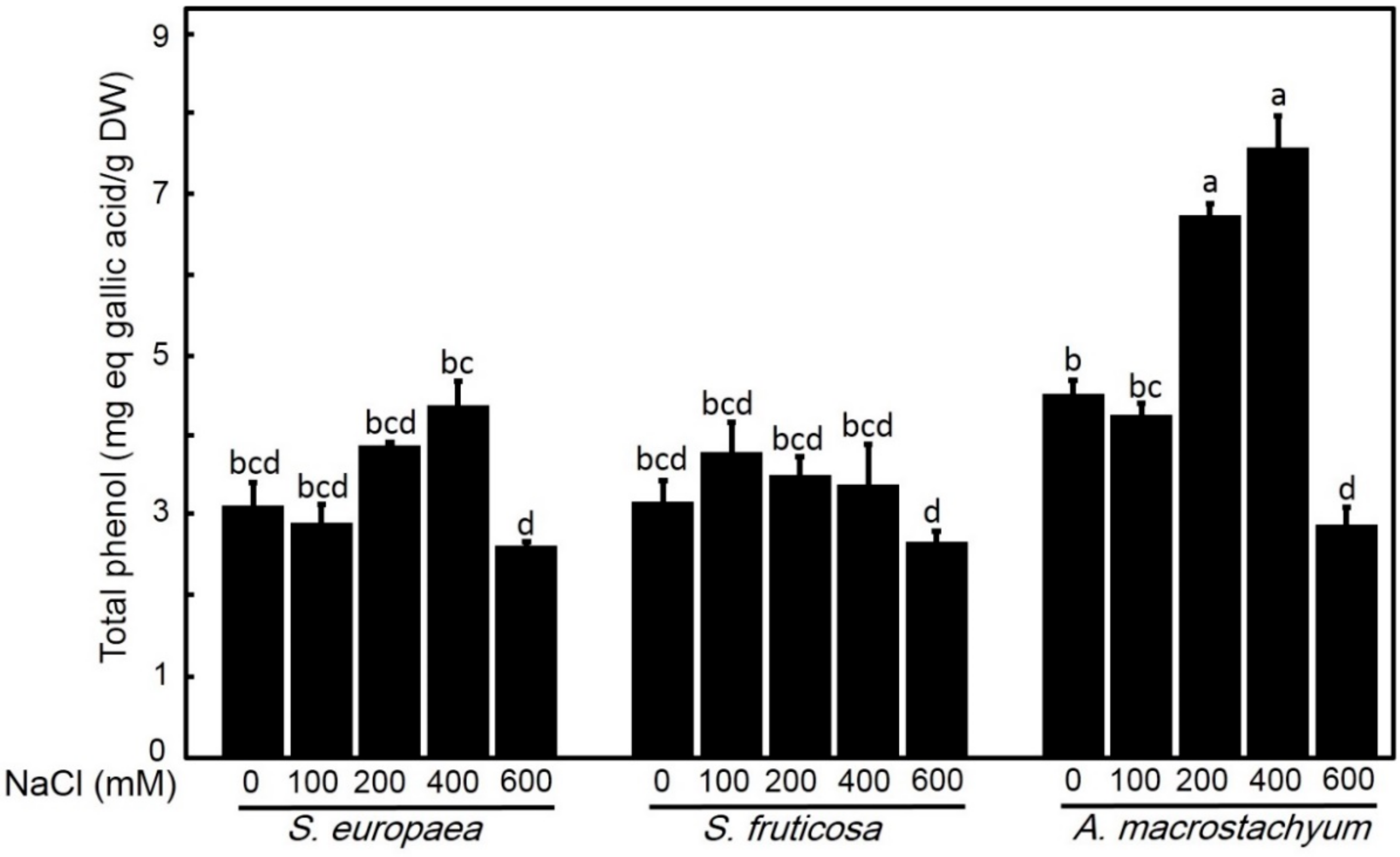
2.2.4. Effect of Salinity on Total Phenolic Contents
2.2.5. Effect of Salinity on Flavonoid Contents
2.2.6. Effect of Salinity on Total Malondialdehyde (MDA) Content
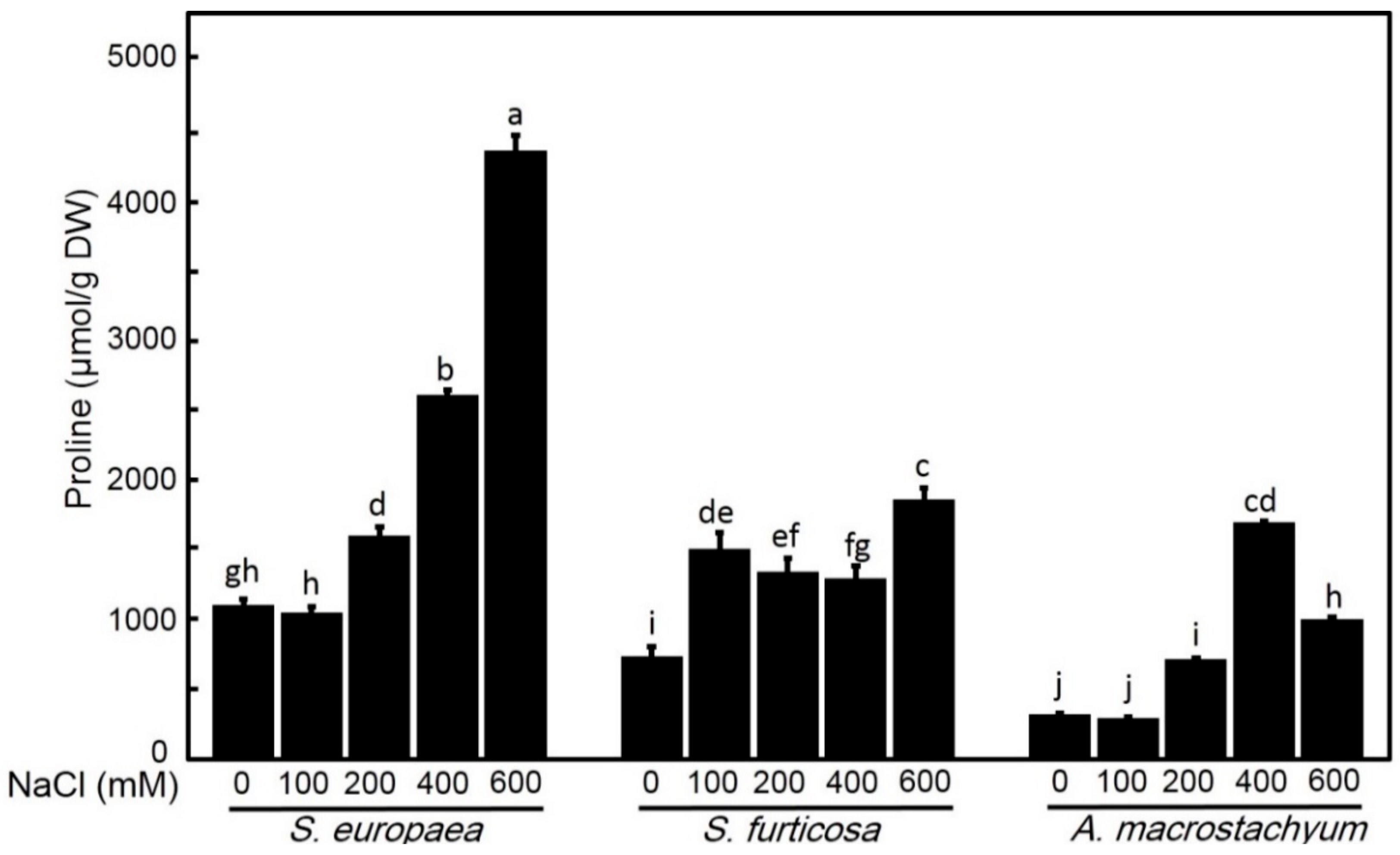
2.2.7. Effect of Salinity on Proline Content
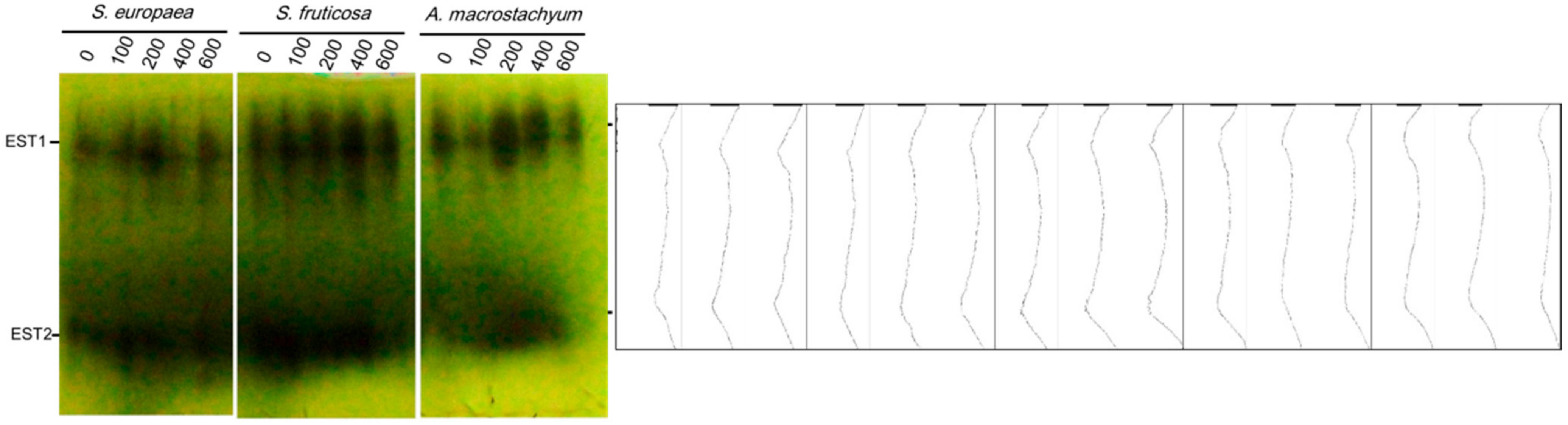
2.3. Isozymes Analysis
2.3.1. Esterases
2.3.2. SOD Isozymes
2.3.3. POD Isozymes
3. Discussion
4. Materials and Methods
4.1. Plant Seeds Collection
4.2. Growth Conditions
4.3. Determination of Na+, K+ and Ca2+
4.4. Growth Parameters
4.4.1. Shoot and Root Fresh and Dry Weight Determination
4.4.2. Determination of Photosynthetic Pigments
4.4.3. Determination of Malondialdehyde (MDA) Content
4.4.4. Determination of Proline Content
4.4.5. Determination of Total Phenolic Compounds and Flavonoids
4.5. Isozymes Analysis
4.5.1. Enzymes Extraction and Detection
4.5.2. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Global Network on Integrated Soil Management for Sustainable Use of Salt-Affected Soils; FAO Land and Plant Nutrition Management Service: Rome, Italy, 2005; Available online: http://www.fao.org/ag/agl/agll/spush (accessed on 29 May 2020).
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 701596. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Plant response to salt stress and role of exogenous protectants to mitigate salt-induced damages. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 25–87. [Google Scholar]
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Herppich, W.B.; Huyskens-keil, S.; Kadau, R. Effects of short-term low-temperature storage on mechanical and chemical properties of white Asparagus cell walls. J. Appl. Bot. Food Qual. 2005, 79, 63–71. [Google Scholar]
- Khan, Z.I.; Hussain, A.; Ashraf, M.; McDowell, L.R. Mineral status of soils and forages in southwestern Punjab-Pakistan: Micro-minerals. Asian Aust. J. Anim. Sci. 2006, 19, 1139–1147. [Google Scholar]
- Zhang, X.; Yin, H.; Chen, S.; He, J.; Guo, S. Changes in antioxidant enzyme activity and transcript levels of related genes in Limonium sinense Kuntze seedlings under NaCl stress. J. Chem. 2014, 749047. [Google Scholar] [CrossRef] [Green Version]
- Poljakoff-Mayber, A. Morphological and anatomical changes in plants as a response to salinity stress. In Plants in Saline Environment; Poljakoff-Mayber, A., Gale, J., Eds.; Springer: New York, NY, USA, 1975; pp. 97–117. [Google Scholar]
- Mansour, M.M.F. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant. 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Hameed, M.; Ashraf, M.; Naz, N. Anatomical adaptations to salinity in cogon grass [Imperata cylindrica (L.) Raeuschel] from the Salt Range, Pakistan. Plant Soil 2009, 322, 229–238. [Google Scholar] [CrossRef]
- Rivelli, A.R.; James, R.A.; Munns, R.; Condon, A.T. Effect of salinity on water relations and growth of wheat genotypes with contrasting sodium uptake. Sciences 2002, 22, 123–131. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Figueroa, M.E.; Davy, A.J. Salt stimulation of growth and photosynthesis in an extreme halophyte, Arthrocnemum macrostachyum. Plant Biol. 2010, 12, 79–87. [Google Scholar] [CrossRef]
- Koyro, H.W.; Hussain, T.; Huchzermeyer, B.; Khan, M.A. Photosynthetic and growth responses of a perennial halophytic grass Panicum turgidum to increasing NaCl concentrations. Environ. Exp. Bot. 2013, 91, 22–29. [Google Scholar] [CrossRef]
- Koyro, H.W. Effect of salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Aghaleh, M.; Niknam, V.; Ebrahimzadeh, H.; Razavi, K. Salt stress effects on growth, pigments, proteins and lipid peroxidation in Salicornia persica and S. europaea. Biol. Plant. 2009, 53, 243–248. [Google Scholar] [CrossRef]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 27, 442–446. [Google Scholar]
- Esfandiari, E.; Shekari, F.; Shekari, F.; Esfandiari, M. The effect of salt stress on antioxidant enzymes activity and lipid peroxidation on the wheat seedling. Not. Bot. Horti Agrobot. Cluj-Napoca 2007, 35, 48–56. [Google Scholar]
- Ozgur, R.; Uzilday, B.; Sekmen, A.H.; Turkan, I. Reactive oxygen species regulation and antioxidant defence in halophytes. Funct. Plant Biol. 2013, 40, 832–847. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, J.; Zhao, Y. Identification and characterization of maize ACD6-like gene reveal ZmACD6 as the maize orthologue conferring resistance to Ustilago maydis. Plant Signal Behav. 2019, 14, e1651604. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.; Matsuda, R.; El-khatib, A.A.; Takechi, K.; Takano, H.; Takio, S. Characterization of the superoxide dismutase genes of the halophyte Suaeda maritima in Japan and Egypt. Plant Cell Rep. 2015, 34, 2099–2110. [Google Scholar] [CrossRef]
- Mohamed, E.; Matsuda, R.; El-khatib, A.A.; Takechi, K.; Takano, H.; Takio, S. Differential tolerance to high salt with regard to cell growth and superoxide dismutase (SOD) activity in calluses of the halophyte Suaeda maritima from Japan and Egypt. Plant Omics 2016, 9, 81–89. [Google Scholar]
- Mohamed, E.; Kasem, A.M.M.A.; Gobouri, A.A.; Elkelish, A.; Azab, E. Influence of Maternal Habitat on Salinity Tolerance of Zygophyllum coccineum with Regard to Seed Germination and Growth Parameters. Plants 2020, 9, 1504. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, E.; Kasem, A.M.M.A.; Farghali, K.A. Seed germination of Egyptian Pancratium maritimum under salinity with regard to cytology, antioxidant and reserve mobilization enzymes, and seed anatomy. Flora 2018, 242, 120–127. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, C.Y.; Song, W.K.; Park, D.S.; Kwon, S.Y.; Lee, H.S.; Bang, J.W.; Kwak, S.S. Overexpression of sweet potato swpa4 peroxidase results in increased hydrogen peroxide production and enhances stress tolerance in tobacco. Planta 2008, 227, 867–881. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and non-enzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Caillet, S.; Salmiéri, S.; Lacroix, M. Evaluation of free radical-scavenging properties of commercial grape phenol extracts by a fast-colorimetric method. Food Chem. 2006, 95, 1–8. [Google Scholar] [CrossRef]
- Amarowicz, R.; Weidner, S. Biological activity of grapevine phenolic compounds. In Grapevine Molecular Physiology and Biotechnology, 2nd ed.; Roubelakis-Angelakis, K.A., Ed.; Springer: New York, NY, USA, 2009; pp. 389–405. [Google Scholar]
- Amarowicz, R.; Weidner, S.; Wojtowicz, I.; Karmac, M.; Kosinska, A.; Rybarczyk, A. Influence of low-temperature stress on changes in the composition of grapevine leaf phenolic compounds and their antioxidant properties. Funct. Plant Sci. Biotechnol. 2010, 4, 90–96. [Google Scholar]
- Lattanzio, V. Phenolic compounds: Introduction. In Natural Products; Ramawat, K.G., Merillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Pereira, A. Plant abiotic stress challenges from the changing environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [Green Version]
- Di Ferdinando, M.; Brunetti, C.; Fini, A.; Tattini, M. Flavonoids as Antioxidants in plants under Abiotic Stresses. In Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 159–179. [Google Scholar]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savouré, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plant species in the world and its annual increase. Phylotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Breckle, S.W. Studies on halophytes from Iran and Afganistan. II Ecology of halophytes along salt gradients. Proceeding R. Soc. Edinb. 1986, 89, 203–215. [Google Scholar]
- Aronson, J. Halophytes. In A Database of Salt Tolerant Plant of the World; Office of Arid Lands Studies, University Arizona Tucson: Tucson, AZ, USA, 1989. [Google Scholar]
- Akhani, P.; Ghorbanli, M. A contribution to the halophytic vegetation and flora of Iran. In Towards the Rational Use of High Salinity Tolerant Plants; Lieth, H., Al-Masoom, A., Eds.; Kluwer Academic Publishers: Dodrecht, The Netherlands, 1993; Volume 1, pp. 35–44. [Google Scholar]
- Öztürk, M.; Altay, V.; Altundağ, E.; Gücel, S. Halophytic plant diversity of unique habitats in Turkey: Salt mine caves of Çankırı and Iğdır. In Halophytes for Food Security in Dry Lands; Khan, M.A., Ozturk, M., Gul, B., Ahmed, M.Z., Eds.; Elsevier Academic Publisher: Oxford, GB, USA, 2016; pp. 291–315. [Google Scholar]
- Winter, K.; Troughton, J.H.; Evenari, M.; Läuchli, A.; Lüttge, U. Mineral ion composition and occurrence of CAM-like diurnal malate fluctuations in plants of coastal and desert habitats of Israel and the Sinai. Oecologia 1976, 25, 125–143. [Google Scholar] [CrossRef]
- Boulos, L. Flora of Egypt; Al Hadara Publication: Cairo, Egypt, 1999; Volume 1, p. 417. [Google Scholar]
- Singh, D.; Buhmann, A.K.; Flowers, T.J.; Seal, C.E.; Papenbrock, J. Salicornia as a crop plant in temperate regions: Selection of genetically characterized ecotypes and optimization of their cultivation conditions. AoB Plants 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Said, O.B.; DaSilva, M.M.; Hannier, F.; Beyrem, H.; Chícharo, L. Using Sarcocornia fruticosa and Saccharomyces cerevisiae to remediate metal contaminated sediments of the Ria Formosa lagoon (SE Portugal). Ecohydrol. Hydrobiol. 2019, 19, 588–597. [Google Scholar] [CrossRef]
- El Naker, N.A.; Yousef, A.F.; Yousef, L.F. A review of Arthrocnemum (Arthrocaulon) macrostachyum chemical content and bioactivity. Phytochem. Rev. 2020, 19, 1427–1448. [Google Scholar] [CrossRef]
- Xiong, L.; Zhu, J.K. Salt tolerance. Arab. Book 2002, 1, e0048. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, J.; Daliakopoulos, I.N.; del Moral, F.; Hueso, J.J.; Tsanis, I.K. A Review of Soil-Improving Cropping Systems for Soil Salinization. Agronomy 2019, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New Insights on Plant Salt Tolerance Mechanisms and Their Potential Use for Breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Gul, B. Arthrocnemum macrostachyum: A potential case for agriculture using above seawater salinity. In Prospects for Saline Agriculture; Springer: Dordrecht, The Netherlands, 2002; pp. 353–364. [Google Scholar]
- Redondo-Gómez, S.; Wharmby, C.; Castillo, J.M.; Mateos-Naranjo, E.; Luque, C.J.; DeCires, A.; Luque, T.; Davy, A.J.; Figueroa, E.M. Growth and photosynthetic responses to salinity in an extreme halophyte, Sarcocornia fruticosa. Physiol. Plant. 2006, 128, 116–124. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Llanderal, A.; Pestana, M.; Correia, P.J.; Lao, M.T. Nutritional and physiological responses of the dicotyledonous halophyte Sarcocornia fruticosa to salinity. Aust. J. Bot. 2017, 65, 573–581. [Google Scholar] [CrossRef]
- Ungar, I.A.; Benner, D.K.; McGraw, D.C. Distribution and growth of Salicornia europaea on an inland salt pan. Ecology 1979, 60, 329–336. [Google Scholar] [CrossRef]
- Cai, Z.Q.; Gao, Q. Comparative physiological and biochemical mechanisms of salt tolerance in five contrasting highland quinoa cultivars. BMC Plant Biol. 2020, 20, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Flowers, T.J.; Munns, R.; Colmer, T.D. Sodium chloride toxicity and the cellular basis of salt tolerance in halophytes. Ann. Bot. 2015, 115, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Ungar, I.A.; Showalter, A.M. Salt stimulation and tolerance in an intertidal stem succulent halophyte. J. Plant Nutr. 2005, 28, 1365–1374. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt tolerance and crop potential. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Shabala, S.; Mackay, A. Ion transport in halophytes. Adv. Bot. Res. 2011, 57, 151–199. [Google Scholar]
- Marcum, K.B.; Murdoch, C.L. Salt tolerance of the coastal salt marsh grass, Sporobolus virginicus (L.) Kunth. New Phytol. 1992, 120, 281–288. [Google Scholar] [CrossRef]
- Hadi, M.R.; Karimi, N. The role of calcium in plants’ salt tolerance. J. Plant Nutr. 2012, 35, 2037–2054. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Sulpice, R.; Gibon, Y.; Fernie, A.R. The enigmatic contribution of mitochondrial function in photosynthesis. J. Exp. Bot. 2008, 59, 1675–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burman, U.; Garg, B.; Kathju, S. Interactive Effects of Thiourea and Phosphorus on Cluster bean Under Water Stress. Biol. Plant. 2004, 48, 61–65. [Google Scholar] [CrossRef]
- Akcin, A.; Yalcin, E. Effect of salinity stress on chlorophyll, carotenoid content, and proline in Salicornia prostrata Pall. and Suaeda prostrata Pall. subsp. prostrata (Amaranthaceae). Braz. J. Bot. 2016, 39, 101–106. [Google Scholar] [CrossRef]
- Rabhi, M.; Castagna, A.; Remorini, D.; Scattino, C.; Smaoui, A.; Ranieri, A.; Abdelly, C. Photosynthetic responses to salinity in two obligate halophytes: Sesuvium portulacastrum and Tecticornia indica. S. Afr. J. Bot. 2012, 79, 39–47. [Google Scholar] [CrossRef]
- Boughalleb, F.; Denden, M. Physiological and biochemical changes of two halophytes, Nitraria retusa (Forssk.) and Atriplex halimus (L.) under increasing salinity. Agric. J. 2011, 6, 327–339. [Google Scholar] [CrossRef]
- Morsy, A.A.; Youssef, A.M.; Mosallam, H.A.; Hashem, A.M. Assessment of selected species along Alamein-Wadi El-Natrun Desert Road. Egypt J. Appl. Sci. Res. 2008, 4, 1276–1284. [Google Scholar]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B.; Sanada, Y.; Mohanty, P. Effects of salinity on biochemical components of the mangrove, Aegiceras corniculatum. Aquat. Bot. 2004, 80, 77–87. [Google Scholar] [CrossRef]
- Król, A.; Amarowicz, R.; Weidner, S. Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous of long-term drought stress. Acta Physiol. Plant. 2014, 6, 1491–1499. [Google Scholar]
- Caliskan, O.; Bayazıt, S.; Oktem, M.; Ergul, A. Evaluation of the genetic diversity of pomegranate accessions from Turkey using new microsatellite markers. Turk. J. Agric. For. 2017, 41, 142–153. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Sood, P.; Citovsky, V. The roles of plant phenolics in defence and communication during agrobacterium and rhizobium infection. Mol. Plant Pathol. 2010, 11, 705–719. [Google Scholar] [CrossRef]
- Weidner, S.; Karolak, M.; Karamać, M.; Kosińska, A.; Amarowicz, R. Phenolic compounds and properties of antioxidants in grapevine roots (Vitis vinifera L.) under drought stress followed by recovery. Acta Soc. Bot. Pol. 2009, 78, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Bettaieb, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and regulation of plants phenolics in abiotic stress tolerance: An overview. In Plant Signaling Molecules: Role and Regulation Under Stressful Environments; Iqbal, M., Ferrante, A., Redd, P.S., Khan, N.A., Eds.; Elsevier Publications: Amsterdam, The Netherlands, 2019; pp. 157–168. [Google Scholar]
- Abd El-Maboud, M.M. Seasonal variations effect on antioxidant compounds and their role in the adaptation of some halophytes at Wadi Gharandal, Southwest Sinai. Ann. Agric. Sci. 2019, 64, 161–166. [Google Scholar] [CrossRef]
- Brown, D.E.; Rashotte, A.M.; Murphy, A.S.; Normanly, J.; Tague, B.W.; Peer, W.A.; Taiz, L.; Muday, G.K. Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis. Plant Physiol. 2001, 126, 524–535. [Google Scholar] [CrossRef] [Green Version]
- Jithesh, M.N.; Prashanth, S.R.; Sivaprakash, K.R.; Parida, A.K. Antioxidative response mechanisms in halophytes: Their role in stress defense. J. Genet. 2006, 85, 237–254. [Google Scholar] [CrossRef]
- Prasad, T.K.; Anderson, M.D.; Stewart, C.R. Localization and characterization of peroxidases in the mitochondria of chilling acclimated maize seedlings. Plant Physiol. 1995, 108, 1597–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mhamdi, A.; Hager, J.; Chaouch, S.; Queval, G.; Han, Y.; Taconnat, L.; Saindrenan, P.; Goua, H.; Issakidis Bourguet, E.; Renou, J.P.; et al. Arabidopsis Glutathione Reductase 1 plays a crucial role in leaf responses to intracellular hydrogen peroxide and in ensuring appropriate gene expression through both salicylic acid and jasmonic acid signaling pathways. Plant Physiol. 2010, 153, 1144–1160. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Nelson, D.E.; Jensen, R.G. Adaptations to environmental stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef]
- Ahmad, P.; John, R.; Sarwat, M.; Umar, S. Responses of proline, lipid peroxidation and antioxidative enzymes in two varieties of Pisum sativum L. under salt stress. Int. J. Plant Prod. 2012, 2, 353–366. [Google Scholar]
- Mohamed, E.; Ansari, N.; Yadav, D.S.; Agrawal, M.; Agrawal, S.B. Salinity alleviates the toxicity level of ozone in a halophyte Mesembryanthemum crystallinum L. Ecotoxicology 2021. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Mann, A.; Kumar, A.; Saha, M.; Lata, C.; Kumar, A. Stress induced changes in osmoprotectants, ionic relations, antioxidants activities and protein profiling characterize Sporobolus marginatus Hochst. ex A. Rich. salt tolerance mechanism. Indian J. Exp. Biol. 2019, 57, 672–679. [Google Scholar]
- Ce’cile, T.; Francoise, L.; Pierre, V.C. Polymorphism and modulation of cell wall esterase enzyme activities in the chicory root during the growing season. J. Exp. Bot. 2006, 57, 81–89. [Google Scholar]
- Dasgupta, N.; Nandy, P.; Tiwari, C.; Das, S. Salinity-imposed changes of some isozymes and total leaf protein expression in five mangroves from two different habitats. J. Plant Interact. 2010, 5, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Bowler, C.; Van Montagu, M.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Mohamed, E.; Kasem, A.M.M.A.; El-khatib, A. Allelopathic potential of Egyptian halophytes Arthrocnemum macrostachyum and Halocnemum strobilaceum from two coastal areas. Allelopath. J. 2020, 50, 225–241. [Google Scholar] [CrossRef]
- Sekmen, A.H.; Turkan, I.; Tanyolac, Z.O.; Ozfidan, C.; Dinc, A. Different antioxidant defense responses to salt stress during germination and vegetative stages of endemic halophyte Gypsophila oblanceolata Bark. Environ. Exp. Bot. 2012, 77, 63–76. [Google Scholar] [CrossRef]
- Nisar, F.; Gul, B.; Khan, M.A.; Hameed, A. Heteromorphic seeds of coastal halophytes Arthrocnemum macrostachyum and A. indicum display differential patterns of hydrogen peroxide accumulation, lipid peroxidation and antioxidant activities under increasing salinity. Plant Physiol. Biochem. 2019, 144, 58–63. [Google Scholar] [CrossRef]
- Aghaleh, M.; Niknam, V.; Ebrahimzadeh, H.; Razavi, K. Effect of salt stress on physiological and antioxidative responses in two species of Salicornia (S. persica and S. europaea). Acta Physiol. Plant. 2011, 33, 1261–1270. [Google Scholar] [CrossRef]
- Cevahir, G.; Yentur, S.; Yazgan, M.; Unal, M.; Yilmazer, N. Peroxidase activity in relation to anthocyanin and chlorophyll content in juvenile and adult leaves of “mini-star” Gazanla splendens. Pak. J. Bot. 2004, 36, 603–609. [Google Scholar]
- Graskova, I.A.; Zhivetyev, M.A.; Putalina, T.E.; Krasnobaev, V.A.; Voinikov, V.N. Activity and izoenzyme spectrum of peroxidase of some herbaceous plants from the bank of lake Baikal, growing under abiotic stress. Electron. Sci. J. Investig. Russ. (Issledovano v Rosii) 2010, 2010, 293–303. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; New Delhi Prentice Hall of India: New Delhi, India, 1973. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, W.R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Carmak, I.; Horst, J.H. Effects of aluminum on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiolgia Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for wate-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bray, H.G.; Thorpe, W.V. Analysis of phenolic compounds of interest in metabolism. Methods Biochem. Anal. 1954, 1, 27–52. [Google Scholar]
- Miliauskas, G.; Venskutonis, P.R.; Van Beek, T.A. Screening of radical scavenging activity of some medicinal and aromatic plant extracts. Food Chem. 2004, 85, 231–237. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.L. Protein Measurements with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.H.; Hamill, D.E.; Weaver, E.A.; Thompson, K.H. Multiple molecular forms of peroxidases and esterases among Nicotiana species and amphidiploids. J. Hered. 1970, 61, 203–212. [Google Scholar] [CrossRef]
- Seevers, P.M.; Daly, J.M.; Catedral, F.F. The role of peroxidase isozymes in resistance to wheat stem rust disease. Plant Physiol. 1971, 48, 353–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to imageJ: 25 years of image analysis. Nature Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| species | NaCl (mM) | Shoot | Root | ||||
|---|---|---|---|---|---|---|---|
| Na+ (µmol g−1 DW) | K+ (µmol g−1 DW) | Ca2+ (µmol g−1 DW) | Na+ (µmol g−1 DW) | K+ (µmol g−1 DW) | Ca2+ (µmol g−1 DW) | ||
| S. europaea | 0 | 2217 ± 5 m | 387 ± 4 ef | 293 ± 8 j | 406 ± 19 k | 493 ± 10 c | 58.75 ± 6 bc |
| 100 | 8239 ± 12 cd | 607 ± 11 a | 2166 ± 22 b | 985± 9 h | 301 ± 13 g | 39 ± 3 ef | |
| 200 | 8637 ± 10 c | 543 ± 11 b | 2300± 14 a | 1239 ± 12 g | 338 ± 11 f | 51.5 ± 4 cd | |
| 400 | 8680 ± 8 c | 362± 12 fg | 2150 ± 16 b | 1474 ± 25 f | 370 ± 7 e | 58 ± 5 bc | |
| 600 | 7969 ± 10 d | 372 ± 8 f | 1925± 28 d | 1670± 20 d | 375 ± 8 e | 66.5 ± 4 b | |
| S. fruticosa | 0 | 6420 ± 10 f | 438± 11 d | 1578 ± 4 e | 330± 16 l | 337 ± 11 f | 13.5 ± 0.75 i |
| 100 | 5760 ± 11 g | 428± 11 d | 1291 ± 35 f | 811 ± 38 i | 294 ± 11 g | 31 ± 3 fg | |
| 200 | 5028 ± 15 j | 264 ± 9 h | 1127 ± 28 h | 1006 ± 19 h | 468 ± 9 cd | 40 ± 5 ef | |
| 400 | 4217 ± 15 k | 211 ± 8 i | 866± 35 i | 1560 ± 22 e | 397 ± 7 e | 55 ± 3 bcd | |
| 600 | 8913 ± 12 b | 262 ± 9 h | 2075 ± 28 c | 1782 ± 22 c | 296 ± 8 g | 90 ± 2.5 a | |
| A. macrostachyum | 0 | 3130 ±22 l | 414 ± 13 de | 85 ± 7 k | 537± 22 j | 454 ± 11 d | 16.5 ± 1 hi |
| 100 | 5057 ± 8 i | 494 ± 9 c | 1162 ± 7 g | 1202 ± 15 g | 671 ± 11 a | 25.75 ± 0.75 gh | |
| 200 | 5736 ± 17 h | 336 ± 8 g | 1239 ± 7 g | 1570± 7 e | 496 ± 7 c | 46.5 ± 2.75 de | |
| 400 | 7760 ± 12 de | 367 ± 8 f | 1664± 10 d | 2338 ± 17 b | 607 ± 11 b | 56 ± 1.25 bcd | |
| 600 | 10652 ± 25 a | 276 ± 11 h | 2187 ± 7 b | 2693 ± 13 a | 620 ± 9 b | 64 ± 3 b | |
| Parameters | Species | Species × Salinity | Salinity |
|---|---|---|---|
| shoot Fresh weight | *** | *** | *** |
| shoot dry weight | *** | *** | *** |
| Root fresh weight | *** | *** | *** |
| Root Dry weight | *** | *** | *** |
| Chl a | *** | *** | *** |
| Chl b | *** | *** | *** |
| Carotenoids | *** | *** | *** |
| Chl a/b | *** | *** | ** |
| MDA | *** | *** | *** |
| Proline | *** | *** | *** |
| phenol | *** | *** | *** |
| Flavonoids | *** | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghanem, A.-M.F.M.; Mohamed, E.; Kasem, A.M.M.A.; El-Ghamery, A.A. Differential Salt Tolerance Strategies in Three Halophytes from the Same Ecological Habitat: Augmentation of Antioxidant Enzymes and Compounds. Plants 2021, 10, 1100. https://doi.org/10.3390/plants10061100
Ghanem A-MFM, Mohamed E, Kasem AMMA, El-Ghamery AA. Differential Salt Tolerance Strategies in Three Halophytes from the Same Ecological Habitat: Augmentation of Antioxidant Enzymes and Compounds. Plants. 2021; 10(6):1100. https://doi.org/10.3390/plants10061100
Chicago/Turabian StyleGhanem, AbdEl-Mageed F. M., Elsayed Mohamed, Ahmed M. M. A. Kasem, and Abbas A. El-Ghamery. 2021. "Differential Salt Tolerance Strategies in Three Halophytes from the Same Ecological Habitat: Augmentation of Antioxidant Enzymes and Compounds" Plants 10, no. 6: 1100. https://doi.org/10.3390/plants10061100