
Joint Biofortification of Plants with Selenium and Iodine: New Field of Discoveries



 ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Consumption Levels
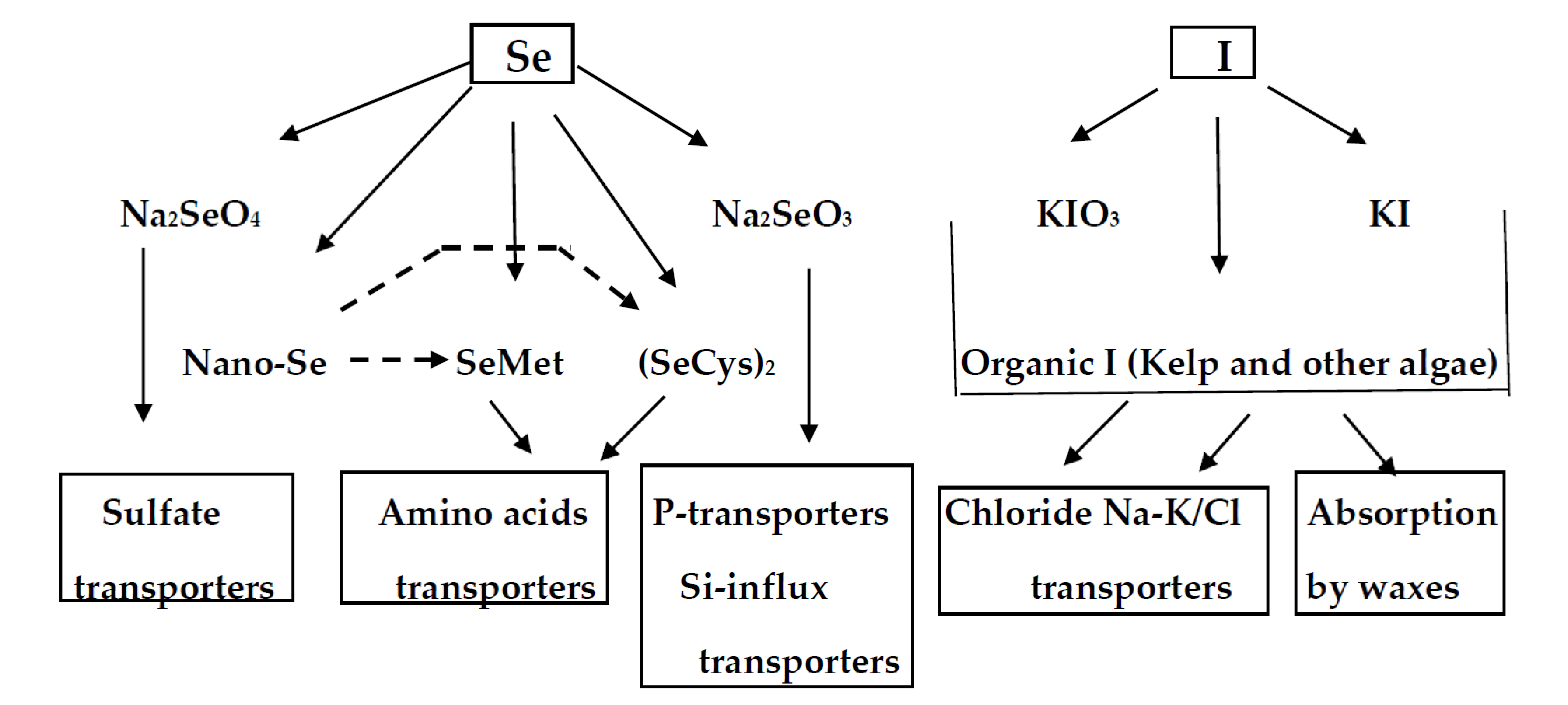
3. Selenium and Iodine Biochemical Characteristics
4. Different Technological Approaches
4.1. Sprouts and Microgreens
4.2. Hydroponics
4.3. Selenium/Iodine Soil Application
4.4. Foliar Biofortification
4.5. The Role of Arbuscular Mycorrhizal Fungi (AMF) and Plant Growth-Promoting Bacteria (GPB)
5. Prospects of Iodine and Selenium Biofortification
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schomburg, L.; Köhrle, J. On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Mol. Nutr. Food Res. 2008, 52, 1235–1246. [Google Scholar] [CrossRef]
- Hetzel, B.S.; Stanbury, J.B. WHO Endemic Goiter and Endemic Cretinism: Iodine Nutrition in Health and Disease; Wiley Medical Publication: Wiley, NY, USA, 1980; 606p. [Google Scholar]
- Lossow, K.; Schwerdtle, T.; Kipp, A. Selenium and iodine–essential trace elements for the thyroid. Ernahr. Umsch. 2019, 66, 175–180. [Google Scholar] [CrossRef]
- Golubkina, N.; Papazyan, T. Selenium in Nutrition: Plants, Animals, Human Being; Pechatny Gorod: Moscow, Russia, 2006. (In Russian) [Google Scholar]
- Golubkina, N.; Kharchenko, V.; Caruso, G. Selenium: Prospects of functional food production with high antioxidant activity. In Plant Antioxidants and Health; Reference Series in Phyto-Chemistry; Ekiert, H.M., Ramawat, K.G., Arora, J., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Lyons, G. Biofortification of cereals with foliar selenium and iodine could reduce hypothyroidism. Front. Plant Sci. 2018, 8, 730. [Google Scholar] [CrossRef] [PubMed]
- White, F.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Bailey, E.H.; Arshad, M.; Ahmed, S.; Watt, M.J.; Stewart, A.G.; Young, S.D. Environmental and human iodine and selenium status: Lessons from Gilgit-Baltistan, North-East Pakistan. Environ. Geochem. Health 2021. [Google Scholar] [CrossRef] [PubMed]
- Izydorczyk, G.; Ligas, B.; Mikula, K.; Witek-Krowiak, A.; Moustakas, K.; Chojnacka, K. Biofortification of edible plants with selenium and iodine—A systematic literature review. Sci. Total Environ. 2020, 754, 141983. [Google Scholar] [CrossRef]
- Dchiavon, M.; Ertani, A.; Parrasia, S.; Vecchia, F.D. Selenium accumulation and metabolism in algae. Aquat. Toxicol. 2017, 189, 1–8. [Google Scholar] [CrossRef]
- Duborská, E.; Urík, M.; Šeda, M. Iodine biofortification of vegetables could improve iodine supplementation status. Agronomy 2020, 10, 1574. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef] [PubMed]
- Kiferle, C.; Martinelli, M.; Salzano, A.M.; Gonzali, S.; Beltrami, S.; Salvadori, P.A.; Hora, K.; Holwerda, H.T.; Scaloni, A.; Perata, P. Evidences for a Nutritional Role of Iodine in Plants. Front. Plant Sci. 2021, 12, 616868. [Google Scholar] [CrossRef]
- Kleine-Kalmer, R.; Profeta, A.; Daum, D.; Enneking, U. Pre-Launch Exploration of Consumer Willingness to Purchase Selenium- and Iodine-Biofortified Apples—A Discrete Choice Analysis of Possible Market Settings. Nutrients 2021, 13, 1625. [Google Scholar] [CrossRef]
- Jóźwiak, W.; Politycka, B. Effect of Selenium on Alleviating Oxidative Stress Caused by a Water Deficit in Cucumber Roots. Plants 2019, 8, 217. [Google Scholar] [CrossRef] [PubMed]
- Antoshkina, M.; Golubkina, N.; Sekara, A.; Tallarita, A.; Сaruso, G. Effects of selenium application on biochemical characteristics and biofortification level of kohlrabi (Brassica oleracea var. gongylodes) produce. Front. Biosci. 2021, in press. [Google Scholar]
- Khalofah, A.; Migdadi, H.; El-Harty, E. Antioxidant Enzymatic Activities and Growth Response of Quinoa (Chenopodium quinoa Willd) to Exogenous Selenium Application. Plants 2021, 10, 719. [Google Scholar] [CrossRef]
- Łukaszewicz, S.; Politycka, B.; Borowiak-Sobkowiak, B. Effect of selenium on alleviating oxidative stress in pea leaves caused by pea aphid feeding. J. Plant Prot. Res. 2021, 61, 83–94. [Google Scholar] [CrossRef]
- Luo, Y.; Wei, Y.; Sun, S.; Wang, J.; Wang, W.; Han, D.; Shao, H.; Jia, H.; Fu, Y. Selenium Modulates the Level of Auxin to Alleviate the Toxicity of Cadmium in Tobacco. Int. J. Mol. Sci. 2019, 20, 3772. [Google Scholar] [CrossRef]
- Thuc, L.V.; Sakagami, J.-I.; Hung, L.T.; Huu, T.N.; Khuong, N.Q.; Vi, L.L.V. Foliar selenium application for improving drought tolerance of sesame (Sesamum indicum L.). Open Agric. 2021, 6, 93–101. [Google Scholar] [CrossRef]
- Groth, S.; Budke, C.; Neugart, S.; Ackermann, S.; Kappenstein, F.-S.; Daum, D.; Rohn, S. Influence of a Selenium Biofortification on Antioxidant Properties and Phenolic Compounds of Apples (Malus domestica). Antioxidants 2020, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Macías, J.M.; Caltzontzit, M.G.L.; Martínez, E.N.R.; Ortiz, W.A.N.; Mendoza, A.B.; Lagunes, P.M. Enhancement to Salt Stress Tolerance in Strawberry Plants by Iodine Products Application. Agronomy 2021, 11, 602. [Google Scholar] [CrossRef]
- Halka, M.; Smoleń, S.; Ledwożyw-Smoleń, I.; Ledwożyw-Smoleń, I.; Sady, W. Comparison of Effects of Potassium Iodide and Iodosalicylates on the Antioxidant Potential and Iodine Accumulation in Young Tomato Plants. J. Plant Growth Regul. 2020, 39, 282–295. [Google Scholar] [CrossRef]
- Ledwożyw-Smoleń, I.; Smoleń, S.; Rożek, S.; Sady, W.; Strzetelski, P. Iodine Biofortification of Potato (Solanum tuberosum L.) Grown in Field. Agronomy 2020, 10, 1916. [Google Scholar] [CrossRef]
- Budke, C.; Dierend, W.; Schön, H.-G.; Hora, K.; Mühling, K.H.; Daum, D. Iodine Biofortification of Apples and Pears in an Orchard Using Foliar Sprays of Different Composition. Front. Plant Sci. 2021, 12, 638671. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Scientific opinion on dietary reference values for selenium. EFSA J. 2014, 12, 38467. [Google Scholar]
- European Food Safety Authority. Scientific opinion on dietary reference values for iodine. EFSA J. 2014, 12, 3660. [Google Scholar]
- Shelford, V.E. Some concepts of bioecology. Ecology 1931, 12, 455–467. [Google Scholar] [CrossRef]
- Zhou, C.; Du, Y.; Rashid, A.; Ram, H.; Savasli, E.; Pieterse, P.J.; Ortiz-Monasterio, I.; Yazici, A.; Kaur, C.; Mahmood, K.; et al. Simultaneous biofortification of wheat with zinc, iodine, selenium, and iron through foliar treatment of a micronutrient cocktail in six countries. J. Agric. Food Chem. 2019, 67, 8096–8106. [Google Scholar] [CrossRef]
- Prom-u-thai, C.; Rashid, A.; Ram, H.; Zou, C.; Guilherme, L.R.G.; Corguinha, A.P.B.; Guo, S.; Kaur, C.; Naeem, A.; Yamuangmorn, S.; et al. Simultaneous biofortification of rice with zinc, iodine, iron and selenium through foliar treatment of a micronutrient cocktail in five countries. Front. Plant Sci. 2020, 11, 589835. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, J.; Kronzucker, H.J.; Shi, W. Selenium biofortification and interaction with other elements in plants: A review. Front. Plant Sci. 2020, 11, 586421. [Google Scholar] [CrossRef]
- Hu, T.; Li, H.; Li, J.; Zhao, G.; Wu, W.; Liu, L.; Wang, Q.; Guo, Y. Absorption and bio-transformation of selenium nanoparticles by wheat seedlings (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- Shaw, G.; Scott, L.K.; Kinnersley, R.P. Sorption of cesium, iodine and sulphur in solution to the axial leaf surface of broad bean (Vicia faba L.). Environ. Exp. Bot. 2009, 59, 361–370. [Google Scholar] [CrossRef]
- Germ, M.; Stibilj, V.; Šircelj, H.; Jerše, A.; Kroflič, A.; Golob, A.; Maršić, N.K. Biofortification of common buckwheat microgreens and seeds with different forms of selenium and iodine. J. Sci. Food Agric. 2019, 99, 4353–4362. [Google Scholar] [CrossRef]
- Jerše, A.; Kacjan-Maršić, N.; Šircelj, H.; Germ, M.; Kroflič, A.; Stibilj, V. Seed soaking in I and Se solutions increases concentrations of both elements and changes morphological and some physiological parameters of pea sprouts. Plant Physiol. Biochem. 2017, 118, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Golob, A.; Kroflić, A.; Jerše, A.; Maršić, N.K.; Šircelj, H.; Stibilj, V.; Germ, M. Response of pumpkin to different concentrations and forms of selenium and iodine, and their combinations. Plants 2020, 9, 899. [Google Scholar] [CrossRef]
- Moldovan, A.; Kharchenko, V.; Kekina, H.; Golubkina, N.; Poluboyarinov, P. Biofortification of Chervil (Anthriscus Cerefolium) Sprouts with Selenium and Iodine//XII International Biogeochemical School ‘Fundamental Basis of Biogeochemical Technologies and Prospects of Their Application to Agriculture and Medicine’ Devoted to 175 Years of Dokuchaev V.V. Birthday, September; Perelomov, L.V., Ermakov, V.V., Eds.; Tomsk State Pedagogical University: Tomsk, Russia, 2021; in press. [Google Scholar]
- Smoleń, S.; Kowalska, I.; Czernicka, M.; Halka, M.; Kęska, K.; Sady, W. Iodine and selenium biofortification with additional application of salicylic acid affects yield, selected molecular parameters and chemical composition of lettuce plants (Lactuca sativa L. var. capitata). Front. Plant Sci. 2016, 7, 1553. [Google Scholar] [CrossRef]
- Smoleń, S.; Kowalska, I.; Kovacik, P.; Halka, M.; Sady, W. Biofortification of six varieties of lettuce (Lactuca sativa L.) with iodine and selenium in combination with the application of salicylic acid. Front. Plants Sci. 2019, 10, 143. [Google Scholar] [CrossRef]
- Zhu, Y.-G.; Huang, Y.; Hu, Y.; Liu, Y.; Christie, P. Interactions between selenium and iodine uptake by spinach (Spinacia oleracea L.) in solution culture. Plant Soil. 2004, 261, 99–105. [Google Scholar] [CrossRef]
- Smolen, S.; Kowalska, I.; Skoczylas, L.; Liszka-Skoczylas, M.; Grzanka, M.; Halka, M.; Sady, W. The effect of salicylic acid on biofortificaton with iodine and selenium and the quality of potato cultivates in the NFT system. Sci. Hort. 2018, 240, 530–543. [Google Scholar] [CrossRef]
- Smoleń, S.; Barański, R.; Ledwozyw-Smoleń, I.; Skoczylas, Ł.; Sady, W. Combined biofortification of carrot with iodine and selenium. Food Chem. 2019, 300, 125202. [Google Scholar] [CrossRef] [PubMed]
- Skoczylas, Ł.; Tabaszewska, M.; Smoleń, S.; Słupski, J.; Liszka-Skoczylas, M.; Barański, R. Carrots (Daucus carota L.) biofortified with iodine and selenium as a raw material for the production of juice with additional nutritional functions. Agronomy 2020, 10, 1360. [Google Scholar] [CrossRef]
- Smoleń, S.; Skoczylas, L.; Ledwozyw-Smoleń, I.; Rakoczy, R.; Kopeć, A.; Piatkowska, E.; Bieżanowska-Kopeć, R.; Pysz, M.; Koronowicz, A.; Kapusta-Duch, J.; et al. Iodine and selenium biofortification of lettuce (Lactuca sativa L.) by soil fertilization with various compounds of these elements. Acta Sci. Pol. Hort. Cultus 2016, 15, 69–91. [Google Scholar]
- Germ, M.; Kacjan-Maršić, N.; Kroflič, A.; Jerše, A.; Stibilj, V.; Golob, A. Significant accumulation of iodine and selenium in chicory (Cichorium intybus L. var. foliosum Hegi) leaves after foliar spraying. Plants 2020, 9, 1766. [Google Scholar] [CrossRef]
- Jerše, A.; Maršić, N.K.; Kroflič, A.; Germ, M.; Šircelj, H.; Stibilj, V. Is foliar enrichment of pea plants with iodine and selenium appropriate for production of functional food? Food Chem. 2018, 267, 368–375. [Google Scholar] [CrossRef]
- Golob, A.; Novak, T.; Maršić, N.K.; Šircelj, H.; Stibilj, V.; Jerše, A.; Kroflič, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes and increases their contents in tubers. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef]
- Golubkina, N.; Kekina, H.; Caruso, G. Yield, quality and antioxidant properties of Indian mustard (Brassica juncea L.) in response to foliar biofortification with selenium and iodine. Plants 2018, 7, 80. [Google Scholar] [CrossRef]
- Golubkina, N.; Amagova, Z.; Matsadze, V.; Caruso, G. Effects of arbuscular mycorrhizal fungi on yield, biochemical characteristics and elemental composition of garlic and onion under selenium supply. Plants 2020, 9, 84. [Google Scholar] [CrossRef]
- El-Ramady, H.; Alshaal, T.; Abdalla, N.; Prokisch, J.; Sztrik, A.; Fári, M.; Domokos-Szabolcs, É. Selenium and nano-selenium biofortified sprouts using micro-farm systems. In Global Advances in Selenium Research from Theory to Application; Banuelos, G.S., Lin, Z.-Q., Moraes, M.F., Guilherme, L.R.G., dos Reis, A.R., Eds.; Conference Paper;·Taylor & Francis Group: London, UK, 2016; pp. 189–190. ISBN 978-1-138-02731-2. [Google Scholar]
- Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium biofortification impacts the nutritive value, polyphenolic content, and bioactive constitution of variable microgreens genotypes. Antioxidants 2020, 9, 272. [Google Scholar] [CrossRef]
- Kyriacoua, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Galieni, A.; Falcinelli, B.; Stagnari, F.; Benincasa, A.D. Sprouts and microgreens: Trends, opportunities, and horizons for novel research. Agronomy 2020, 10, 1424. [Google Scholar] [CrossRef]
- Liu, H.; Kang, Y.; Zhao, X.; Liu, Y.; Zhang, X.; Zhang, S. Effects of elicitation on bioactive compounds and biological activities of sprouts. J. Funct. Foods 2019, 53, 136–145. [Google Scholar] [CrossRef]
- Golubkina, N. Selenium biorhythms and hormonal regulation. In Selenium. Sources, Functions and Health Effects; Aomori, C., Hokkaido, M., Eds.; Novo Science Publishers: New York, NY, USA, 2012; pp. 33–75. [Google Scholar]
- Golubkina, N.; Folmanis, G.; Tananaev, I.; Krivenkov, L.; Kosheleva, O.; Soldatenko, A. Comparative evaluation of spinach biofortification with selenium nanoparticles and ionic forms of the element. Nanotechnologies Russ. 2017, 12, 569–576. [Google Scholar] [CrossRef]
- Sivaramakrishnan, R.; Incharoensakdi, A. Plant hormone induced enrichment of Chlorella sp. omega-3 fatty acids. Biotechnol. Biofuels 2020, 13, 7. [Google Scholar] [CrossRef]
- Nguyen, T.Q.; Sesin, V.; Kisiala, A.; Emery, R.J.N. Phytohormonal roles of plant responses to heavy metal stress: Implications for using macrophytes in phytoremediation of aquatic ecosystems. Environ. Toxicol. Chem. 2021, 40, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Alfthan, G.; Eurola, M.; Ekholm, P.; Venäläinen, E.R.; Root, T.; Korkalainen, K.; Hartikainen, H.; Salminene, P.; Hietaniemi, V.; Aspila, P.; et al. Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: From deficiency to optimal selenium status of the population. J. Trace Elem. Med. Biol. 2015, 31, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Ligowe, I.S.; Bailey, E.H.; Young, S.D.; Ander, E.L.; Kabambe, V.; Chilimba, A.D.; Lark, R.M.; Nalivata, P.C. Agronomic iodine biofortification of leafy vegetables grown in Vertisols, Oxisols and Alfisols. Environ. Geochem. Health 2021, 43, 361–374. [Google Scholar] [CrossRef]
- Weng, H.-X.; Weng, J.-K.; Yan, A.-L.; Hong, C.-L.; Yong, W.-B.; Qin, Y.-C. Increment of iodine content in vegetable plants by applying iodized fertilizer and the residual characteristics of iodine in soil. Biol. Trace Elem. Res. 2008, 123, 218–228. [Google Scholar] [CrossRef]
- Kharchenko, V.A.; Golubkina, N.A.; Moldovan, A.I.; Caruso, G. Biofortifiction of chervil with selenium. Veg. Crops Russ. 2021, 79–86. [Google Scholar] [CrossRef]
- Golubkina, N.; Gomez, L.; Kekina, H.; Hallam, R.; Tallarita, A.; Cozzolino, E.; Torino, V.; Koshevarov, A.; Cuciniello, A.; Maiello, R.; et al. Joint selenium-iodine supply and arbuscular mycorrhizal fungi inoculation affect yield and quality of chickpea seeds and residual biomass. Plants 2020, 9, 804. [Google Scholar] [CrossRef]
- Kapoor, R.; Singh, N. Arbuscular mycorrhiza and reactive oxygen species. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Wu, Q.-S., Ed.; Springer: Singapore, 2017; Chapter 10. [Google Scholar] [CrossRef]
- Carrino-Kyker, S.R.; Kluber, L.A.; Coyle, K.P.; Burke, D.J. Detection of phosphate transporter genes from arbuscular mycorrhizal fungi in mature tree roots under experimental soil pH manipulation. Symbiosis 2017, 72, 123–133. [Google Scholar] [CrossRef]
- Higo, M.; Tatewaki, Y.; Iida, K.; Yokota, K.; Isobe, K. Amplicon sequencing analysis of arbuscular mycorrhizal fungal communities colonizing maize roots in different cover cropping and tillage systems. Sci. Rep. 2020, 10, 6093. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, W.; Zhou, F.; Du, Z.; Xue, M.; Chen, T.; Liang, D. Effect of phosphate and silicate on selenite uptake and phloem-mediated transport in tomato (Solanum lycopersicum L.). Environ. Sci. Pollut. Res. 2019, 26, 20475–20484. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Qu, J.; Pu, Y.; Rao, S.; Xu, F.; Wu, C. Selenium biofortification of crop food by beneficial microorganisms. J. Fungi 2020, 6, 59. [Google Scholar] [CrossRef]
- Conversa, G.; Lazzizera, C.; Chiaravalle, E.; Elia, A. Selenium fern application and arbuscular mycorrhizal fungi soil inoculation enhance Se content and antioxidant properties of green asparagus (Asparagus officinalis L.) spears. Sci. Hortic. 2019, 252, 176–191. [Google Scholar] [CrossRef]
- Golubkina, N.; Zamana, S.; Seredin, T.; Poluboyarinov, P.; Sokolov, S.; Baranova, H.; Krivenkov, L.; Pietrantonio, L.; Caruso, G. Effect of selenium biofortification and arbuscular mycorrhizal fungi on yield, quality and antioxidant properties of shallot bulbs. Plants 2019, 8, 102. [Google Scholar] [CrossRef]
- Soldatov, S.A.; Khryanin, V.I. Effect of sodium selenate on growth and development of Cannabis sativa L. Izv. Penza State Univ. 2006, 1, 61–65. (In Russia) [Google Scholar]
- Durán, P.; Acuñab, J.J.; Jorquera, M.A.; Azcónc, R.; Borie, F.; Cornejo, P.; Mora, M.L. Enhanced selenium content in wheat grain by co-inoculation of selenobacteria and arbuscular mycorrhizal fungi: A preliminary study as a potential Se biofortification strategy. J. Cereal Sci. 2013, 57, 275–280. [Google Scholar] [CrossRef]
- Luo, W.; Li, J.; Ma, X.; Niu, H.; Hou, S.; Wu, F. Effect of arbuscular mycorrhizal fungi on uptake of selenate, selenite, and selenomethionine by roots of winter wheat. Plant Soil 2019, 438, 71–83. [Google Scholar] [CrossRef]
- Durán, P.; Acuña, J.J.; Armada, E.; López-Castillo, O.M.; Cornejo, P.; Mora, M.L.; Azcón, R. Inoculation with selenobacteria and arbuscular mycorrhizal fungi to enhance selenium content in lettuce plants and improve tolerance against drought stress. J. Soil Sci. Plant Nutr. 2016, 16. [Google Scholar] [CrossRef][Green Version]
- Souza, M.P.; Chu, D.; Zhao, M.; Zayed, A.M.; Ruzin, S.E.; Schichnes, D.; Terry, N. Rhizosphere bacteria enhance selenium accumulation and volatilization by Indian mustard. Plant Physiol. 1999, 119, 565–574. [Google Scholar] [CrossRef]
- Yasin, M.; El Mehdawi, A.F.; Jahn, C.E.; Anwar, A.; Turner, M.F.; Faisal, M.; Pilon-Smits, E.A. Seleniferous soils as a source for production of selenium-enriched foods and potential of bacteria to enhance plant selenium uptake. Plant Soil 2015, 386, 385–394. [Google Scholar] [CrossRef]
- Grzanka, M.; Smoleń, S.; Kováčik, P. Effect of vanadium on the uptake and distribution of organic and inorganic forms of iodine in sweetcorn plants during early-stage development. Agronomy 2020, 10, 1666. [Google Scholar] [CrossRef]
- Golubkina, N.; Logvinenko, L.; Molchanova, A.; Caruso, G. Genetic and environmental influence on macro- and microelement accumulation in plants of Artemisia species. In Plant Micronutrients; Deficiency and Toxicity Management, ed.; Aftab, T., Haleem, K.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; Chapter 17; pp. 389–416. [Google Scholar] [CrossRef]
- Siason, E.; Willey, N. Inter-taxa differences in iodine uptake by plants: Implications for food quality and contamination. Agronomy 2016, 5, 537–554. [Google Scholar] [CrossRef]
- Panchal, S.K.; Wanyonyi, S.; Brown, L. Selenium, vanadium, and chromium as micronutrients to improve metabolic syndrome. Curr. Hypertens. Rep. 2017, 19, 10. [Google Scholar] [CrossRef]
- Urban, J.; Antonowicz-Juchniewicz, J.; Andrzejak, R.; Nadzieje, W.-Z.I. Wanad–zagrożenia i nadzieje. Med. Pr. 2001, 52, 123–133. [Google Scholar]
- Gruzewska, K.; Michno, A.; Pawelczyk, T.; Bielarczyk, H. Essentiality and toxicity of vanadium supplements in health and pathology. J. Physiol. Pharmacol. 2014, 65, 603–611. [Google Scholar]
- Kaplan, D.I.; Adriano, D.C.; Carlson, C.L.; Sajwan, K.S. Vanadium: Toxicity and accumulation by beans. Water. Air. Soil Pollut. 1990, 49, 81–91. [Google Scholar] [CrossRef]
- Medrano-Macías, J.; Leija-Martínez, P.; González-Morales, S.; Juárez-Maldonado, A.; Benavides-Mendoza, A. Use of iodine to biofortify and promote growth and stress tolerance in crops. Front. Plant. Sci. 2016, 7, 1146. [Google Scholar] [CrossRef]
- Trouvelot, S.; Héloir, M.-C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trdá, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant. Sci. 2014, 5, 592. [Google Scholar] [CrossRef]
- Medrano-Macias, J.; Mendoza-Villarreal, R.; Robledo-Torres, V.; Fuentes-Lara, L.O.; Ramirez-Godina, F.; Prez-Rodriquez, M.A.; Benavides-Mendoza, A. The use of iodine, selenium and silicon in plant nutrition for the increase of antioxidants in fruits and vegetables. In Antioxidants in Foods and Its Applications; Intech Open: London, UK, 2018; Chapter 8; pp. 155–168. [Google Scholar] [CrossRef]
- Sieprawska, A.; Kornaś, A.; Filek, M. Involvement of selenium in protective mechanism of plants under environmental stress conditions–review. Acta Biol. Crac., Ser. Bot. 2015, 57, 9–20. [Google Scholar] [CrossRef]
- Schiavon, M.; Lima, L.W.; Jiang, Y.; Hawkesford, M. Effects of selenium on plant metabolism and implications for crops and consumers. In Selenium in plants Molecular, Physiological, Ecological and Evolutionary Aspects, 1st ed.; Pilon-Smits, E.A.H., Winkel, L.H.E., Lin, Z.-Q., Eds.; Springer International Publishing: Basel, Switzerland, 2017; Chapter 15. [Google Scholar] [CrossRef]
- Kiferle, C.; Gonzali, S.; Holwerda, H.T.; Ibaceta, R.R.; Perata, P. Tomato fruits: A good target for iodine biofortification. Front. Plant. Sci. 2013, 4, 205. [Google Scholar] [CrossRef]
- Li, R.; Li, D.-W.; Liu, H.-P.; Hong, C.-L.; Song, M.-Y.; Dai, Z.-X.; Liu, J.-W.; Zhou, J.; Weng, H. Enhancing iodine content and fruit quality of pepper (Capsicum annuum L.) through biofortification. Sci. Hortic. 2017, 214, 165–173. [Google Scholar] [CrossRef]
- Golubkina, N.; Krivenkov, L.; Sekara, A.; Vasileva, V.; Tallarita, A.; Caruso, G. Prospects of arbuscular mycorrhizal fungi utilization in production of Allium Plants. Plants 2020, 9, 279. [Google Scholar] [CrossRef]
- Mora, M.L.; Durán, P.; Acuña, A.J.; Cartes, P.; Demanet, R.; Gianfreda, L. Improving selenium status in plant nutrition and quality. J. Soil Sci. Plant. Nutr. 2015, 15, 486–503. [Google Scholar] [CrossRef]
- El-Ramady, H.; Faizy, S.E.-D.; Abdalla, N.; Taha, H.; Domokos-Szabolcsy, É.; Fari, M.; Elsakhawy, T.; El-Dein Omara, A.; Shalaby, T.; Bayoumi, Y.; et al. Selenium and nano-selenium biofortification for human health: Opportunities and challenges. Soil Syst. 2020, 4, 57. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H. On the ecology of selenium accumulation in plants. Plants 2019, 8, 197. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Dose and Method of Application | Beneficial Effect | References |
|---|---|---|---|
| Selenium | |||
| Cucumber | 1–5 µM Na2SeO3 Seedling exposure | Increase of seedling tolerance to water deficiency, by increasing the activities of the antioxidant enzymes and decrease of plasma membranes damage | [15] |
| Kohlrabi | Foliar supply Na2SeO4 50–100 mg L−1 | Increase of stem weight (by 1.35–1.61 times), yield (1.37–1.66 times), monosaccharide (1.59–2.24 times), ascorbic acid (1.54–2.01 times) and total phenolic (by 1.23–1.37 times) levels | [16] |
| Quinoa | 2.5 and 5 mg L−1 soil application at early plant growth stage | Growth parameters, relative water content, photosynthetic pigments, proline, total soluble sugars, and antioxidant enzyme activities (superoxide dismutase, catalase, peroxidase, ascorbate peroxidase, glutathione reductase) increase, and decrease of malondialdehyde and H2O2 content. | [17] |
| Pea | 10 µM Na2SeO3 or 20 µM Na2SeO4 in nutrient seedlings exposure | Protection against pea aphid Acyrthosiphon pisum | [18] |
| Tobacco | 10 μM Na2Se03 Seedling’s exposure | Increase of biomass and antioxidant capacity and reduced uptake of Cd. Higher auxin concentrations at Cd uptake compared with lack of Se supply | [19] |
| Sesame | Foliar supply, 5 mg L−1, pot experiment | Maintaining the number of leaves and increase proline accumulation, plant biomass, and grain weight per plant | [20] |
| Apple | Foliar 0.15 kg Se ha−1 (Na2SeO4) | Increase in TP, TAA and polyphenol oxidase activity | [21] |
| Iodine | |||
| Strawberry | 100 µM KIO3 bi-weekly | Enhancement of salt stress tolerance, increase in GSH and APX activity, P, K and Ca, Mn, ascorbic acid and I accumulation | [22] |
| Tomato | 5–10 µM KI, 5-iodo salicylic acid nutrient solution | Reduction of ascorbic acid and increase of dehydroascorbic acid content in leaves. Significant increase of ascorbate peroxidase activity only with 10 µM of KI and 5-iodo salicylic acid | [23] |
| Potato | Soil application of KI and foliar application of KIO3 in doses up to 2.0 kg I ha−1 | Increased content of I with no decrease of starch or sugar content. The highest efficiency of iodine biofortification was noted with KIO3 foliar spraying at 2.0 kg I ha−1 | [24] |
| Apple, pear | 0.5 kg KIO3 ha−1 foliar application | Increase of total soluble solids content of fruits up to 1.0 Brix | [25] |
| Daily Se Requirement (µg) | Daily I Requirement (µg) | |
|---|---|---|
| Infants | 10–15 | 40–80 |
| Children (1–10 years) | 15–30 | 100–140 |
| Adolescents | 45–70 | 180–200 |
| Adults | 60–70 | 200 |
| Pregnant women | 60 | 230 |
| Breastfeeding women | 75 | 260 |
| Upper limit | 300 | 600 |
| Object | Chemical Forms of Elements | Doses | Results (Se and I Content) | Se–I Interaction | Ref. |
|---|---|---|---|---|---|
| Foliar application of Zn, I, Se, Fe Cocktail | |||||
| Wheat 10 cultivars | ZnSO4+ KIO3+ Na2SeO4+Fe, EDTA | 0.05% KIO3 0.001% Na2SeO4 | No significant effect on grain yield (338 μg Se kg−1; 249 μg I kg−1) | Decreased I levels via cocktail supply compared to single I application | [29] |
| Rice 7 cultivars | ZnSO4 KIO3 Na2SeO4 FeEDTA | 0.05% KIO3 0.001% Na2SeO4 (5 countries) | No effect on grain yield (90-584 μg Se kg−1; 101-335 μg I kg−1) | No data | [30] |
| Sprouts | |||||
| Common buckwheat (microgreens) | SeO32- SeO4 2-; I−; IO3− | 10 mg Se L−1 1000 mg I L−1 | Under Se–I combined treatment, microgreens yield was 50–70% higher than with Se and I singly (Se and I reached the contents of 0.24 μg g−1 DW and 216 μg g−1 DW, respectively) | Se decreased I by 50%, and I increased Se by 50% | [34] |
| Pea | KI, KIO3 + Na2SeO3, Na2SeO4 | 1000 mg I L−1 10 mg Se L−1 | No effect on chlorophyll accumulation and a slight decrease of biomass (3.9–14.1 µg Se g–1 DW; 152–247 µg I g–1 DW) | No significant relationship between elements | [35] |
| Pumpkin | Seed soaking + foliar application in the field | 10 mg Se L−1, 1000 mg I L−1 | Enhanced germination, no effect on yield (0.8–2.3 μg Se g−1 DW; 288–323 μg I g−1 DW) | Synergism in sprouts; I increased seed Se accumulation | [36] |
| Chervil | Na2SeO4 + KIO3 KI+ (SeCys)2 | 5 µM | Growth stimulation and TAA/TP increase only for KIO3+ Na2SeO4 (0.89–0.90 μg Se g−1 DW; 0.29–0.46 μg I g−1 DW) | No significant relationship between elements | [37] |
| Hydroponics | |||||
| Lettuce | Na2SeO4, KIO3 Salicylic acid | 30 mg I m−3; 8.5 mg Se dm−3, | SeMet and sugar increase, no effect on biomass; root Р increase and Mg decrease; the effect is dose-dependent (7.8–10.4 mg Se kg−1 DW; about 250 mg I kg−1 DW) | No data | [38] |
| Lettuce 6 varieties | KIO3 Na2SeO3 Salicylic acid | 5 mg I L−1 0.5 mg Se L−1 | High varietal differences (7.5-13.7 μg Se g−1 DW; 75.1–304.7 μg I g−1 DW) | No data | [39] |
| Spinach | KIO3 Na2SeO4 | 10 µM I 50 µM Se | I-Se transfer factor: 3.5 to 13.4 (3–13 mg Se kg−1 FW; 10–25 mg I kg−1 FW) | I did not influence Se accumulation and vice versa | [40] |
| Potato | KIO3 Na2SeO3 Salicylic acid (SA) | 39.4 µM I 6.3 µM Se | I, Se, SA did not affect tubers yield; 1 mg SA L−1 + (I+Se) resulted in the highest I tuber content; SA did not affect Se; N, K, Na increased and Mn, Zn decreased (100 g of fresh tubers provide 444–489% RDA Se and 47–71% RDA I) | No data | [41] |
| Soil application | |||||
| Carrot | KI Na2SeO4 | 4 kg I ha−1 + 0.25 kg Se ha−1 | Low effect of Se and I on biochemical characteristics of roots; 100 g of biofortified carrots substantially cover the RDA for I and Se | No data | [42] |
| Carrot | KI Na2SeO4 | 4 kg I ha−1 and 0.25 kg Se ha−1 | Fertilization had no effect on yield (7.24 mg Se kg−1 DW; 1.47 mg I kg−1 DW) (juice) | No data | [43] |
| Lettuce | Na2SeO3 Na2SeO4 KI KIO3 | 2.5 kg I·ha−1 + 0.5 kg Se·ha−1 | SeMet and SeCys2 increase; higher biofortification level for KI and Na2SeO4 (9.4–86.7 mg Se kg−1; 4.2–4.7 mg I kg−1) | Decrease of I and Se accumulation under joint application | [44] |
| Foliar application | |||||
| Chicory | KI, KIO3, Na2SeO3, Na2SeO4 | 10 mg Se salt L−1; 1000 mg I salt L−1 | No effect on plant biomass (73–85 μg Se kg−1 DW; 75 μg I kg−1 DW) | I increased Se+4 accumulation but decreased that of Se+6 | [45] |
| Pea | KI KIO3 Na2SeO3 Na2SeO4 | 1000 mg I L−1 (KI or KIO3) 10 mg Se L−1 (Na2SeO3 or Na2SeO4) | No growth depression (up to 0.18–0.19μg Se kg−1 DW; >2% RDA for I) | Se+4 increased I in pea leaves, roots and pods; Se+6 increased seed I− | [46] |
| Kohlrabi | Na2SeO3, Na2SeO4 KI KIO3 | 1 g I L−1, 10 mg Se L−1 | Se increased chlorophyll and carotene content; I increased anthocyanins; (100 g of fresh tubers provide 1.38–8.5% RDA Se and 0.79–2.01% RDA I) | Se had antagonistic effects on accumulation of I in leaves. | [47] |
| Indian mustard | Field experiment KI Na2SeO4 | 50 mg Na2SeO4 L−1 100 mg KI L−1 | Al, B increased; Cd, Sr decreased; NO3− decreased especially under joint Se–I application; 8.6 mg Se kg−1 DW; 2.8 mg I kg−1 DW | I and Se synergism under separate supply and no effect under joint Se–I application | [48] |
| Chickpea | Na2SeO4 KI AMF inoculation | 100 mg KI L−1 50 mg Na2SeO4 L−1 | Improvement of yield; 3305 μg Se kg−1 DW; 15 μg I kg−1 DW | Increase of Se and I fortification level by AMF; Se–I synergism | [49] |
| Apple, pear | KIO3, Na2SeO4 | 0.5 kg KIO3 ha−1 0.05 kg Na2SeO4 ha−1 | 51% and 75% of the biofortified I was localized in the apple and pear peel, respectively; 20–30 μg Se kg−1 FW, 500–600 μg I kg−1 FW | No effect of Se on I accumulation | [25] |
| Treatment | Se µg kg−1 d.w | I µg kg−1 d.w. | TAA mg GAE g−1 d.w. | TP mg GAE g−1 d.w. |
|---|---|---|---|---|
| Control (water) | 81 ± 8b | traces | 14.89 ± 1.7а | 8.12 ± 1.1а |
| Na2SeO4 | 850 ± 84а | traces | 12.9 ± 1.4a | 7.4 ± 0.6a |
| KI | 86 ± 9a | 443 ± 115a | 14.5 ± 1.5a | 8.2 ± 0.9a |
| Na2SeO4 + KI | 890 ± 91a | 288 ± 75a | 12.9 ± 1.4a | 7.3 ± 0.6a |
| KIO3 | 85 ± 8b | 327 ± 85a | 19.9 ± 2.0b | 12.6 ± 1.5b |
| Na2SeO4 + KIO3 | 900 ± 92a | 460 ± 120a | 17.8 ± 1.9b | 11.7 ± 1.9b |
| Basic Points | Promising Directions | |
|---|---|---|
| Known Facts | References | |
| Close relationship of Se with N, P, S, Si accumulation | [86] | Effect of N, P, S, Si on the efficiency of Se–I biofortification |
| Protective role of Se against biotic and abiotic stresses including heavy metals | [65,87] | Se–I biofortification under oxidant stress |
| Close relationship of Se, sugar and antioxidants accumulation | [5,88] | Effect of Se–I biofortification on sugar and antioxidants accumulation |
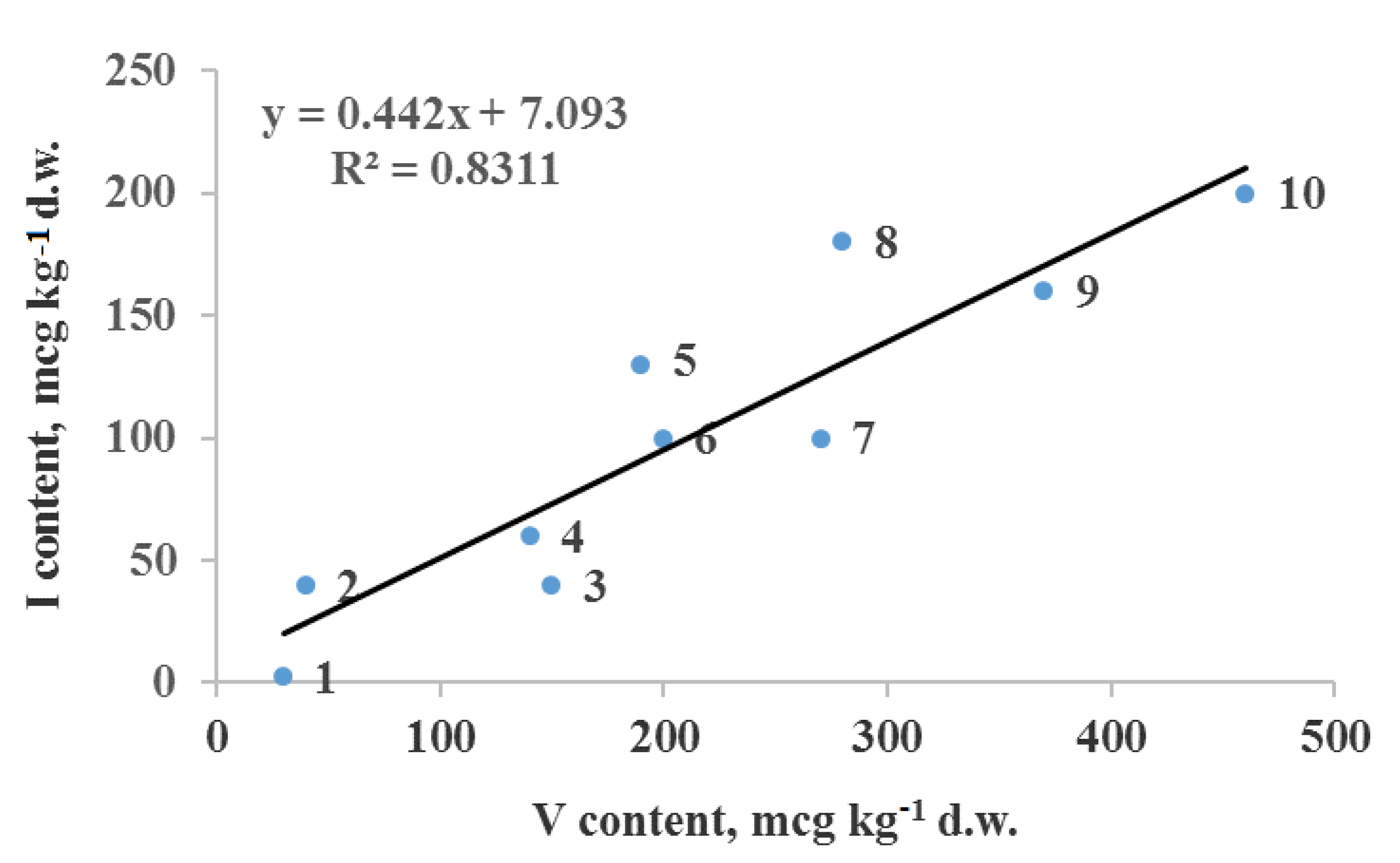
| Stimulation of I accumulation by vanadium (sweetcorn) | [77] | Effect of V on the efficiency of Se–I biofortification |
| Separate biofortification of tomato, pepper and onion with Se and I | [11,24,48,89,90] | Joint Se–I biofortification of tomato, onion, pepper and garlic (vegetables widely used by the population) |
| Increase in Se accumulation by AMF and growth promoting bacteria; a single example of Se–I biofortification of chickpea under AMF supply | [63,91,92] | Efficiency of AMF and growth promoting bacteria application on joint Se–I biofortification of different agricultural crops |
| High efficiency of plant biofortification with organic selenium (SeCys)2 and Se nanoparticles | [32,56,70,93] | Efficiency of (SeCys)2 and Se NP utilization in joint Se–I biofortification of plants |
| Division of plants to hyperaccumulators, indicators and non-accumulators of Se; with low and high iodine accumulation capacity | [7,79,94] | Efficiency of joint Se–I biofortification of hyperaccumulators, for Se hyperaccumulators and I accumulators in particular |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golubkina, N.; Moldovan, A.; Kekina, H.; Kharchenko, V.; Sekara, A.; Vasileva, V.; Skrypnik, L.; Tallarita, A.; Caruso, G. Joint Biofortification of Plants with Selenium and Iodine: New Field of Discoveries. Plants 2021, 10, 1352. https://doi.org/10.3390/plants10071352
Golubkina N, Moldovan A, Kekina H, Kharchenko V, Sekara A, Vasileva V, Skrypnik L, Tallarita A, Caruso G. Joint Biofortification of Plants with Selenium and Iodine: New Field of Discoveries. Plants. 2021; 10(7):1352. https://doi.org/10.3390/plants10071352
Chicago/Turabian StyleGolubkina, Nadezhda, Anastasia Moldovan, Helene Kekina, Victor Kharchenko, Agnieszka Sekara, Viliana Vasileva, Liubov Skrypnik, Alessio Tallarita, and Gianluca Caruso. 2021. "Joint Biofortification of Plants with Selenium and Iodine: New Field of Discoveries" Plants 10, no. 7: 1352. https://doi.org/10.3390/plants10071352
APA StyleGolubkina, N., Moldovan, A., Kekina, H., Kharchenko, V., Sekara, A., Vasileva, V., Skrypnik, L., Tallarita, A., & Caruso, G. (2021). Joint Biofortification of Plants with Selenium and Iodine: New Field of Discoveries. Plants, 10(7), 1352. https://doi.org/10.3390/plants10071352