
Plant Invasion Research in Russia: Basic Projects and Scientific Fields


 and
and
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. An Inventory of Alien Plant Species
3.2. An Inventory of Invasive Plant Species
3.3. Analysis of Long-Term Dynamics of Alien Flora and Its Invasive Component
3.4. Development of Methods for Alien Species Study
3.5. An Inventory of Polemochores
3.6. Identification of Naturalized Species in Botanical Gardens
- (a)
- presenting noxious species in order to develop effective means of control. A provisional name for this exhibition could be “Our cultivation in the garden is strongly prohibited!” For a better effect, such an exhibition should be located directly in front of the entrance to the botanical garden. Oenothera biennis L. and Conyza canadensis (L.) Cronq. may be planted in the background; Galinsoga parviflora Cav., G. quadriradiata Ruiz & Pav., Matricaria discoidea DC., etc., may be planted in the front zone;
- (b)
- presenting species which are actively turning wild within the garden territory, being potentially invasive. In fact, the species mentioned in the previous paragraph (e.g., Conyza and Galinsoga) escaped from botanical gardens a few centuries ago. A provisional name for such an exhibition could be “Do not let us escape!” Helianthus tuberosus L., Solidago canadensis L., Echinocystis lobata (Michx.) Torr. & A. Gray, and Aster novi-belgii may be planted in the background; Oxalis stricta L. and Bellis perennis L. may be planted in the front zone.
3.7. Microevolution of Alien Species in the Secondary Distribution Range
3.8. Identifying the Biological Characteristics of Invasive Species
3.9. Comparative Analysis of the Invasive Status of Closely Related Alien Species
3.10. Studying Micromorphological Characteristics in Invasive Species and Assessing Their Significance for Taxonomy
3.11. Studying the Phytochemical Characteristics of alien Species
3.12. The Influence of Hybridization Processes on Alien Plant Species Invasion Rate
3.13. Removing the Most Invasive Species for Experimental Purposes
3.14. Invasive Species as Resource Plants
3.15. Glossary Compilation
3.16. Studying the Consortial Relationships between Invasive Species and Pathogens, Phytophages, and Pollinating Insects
3.17. Creating a System to Track Invasive Species Distribution Online
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darwin, C.R. The Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859; pp. 1–502. [Google Scholar]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Methuen: London, UK, 1958; pp. 1–181. [Google Scholar]
- Petrosyan, V.G.; Khlyap, L.A.; Reshetnikov, A.N.; Krivosheina, M.G.; Dergunova, N.N.; Osipov, F.A.; Dgebuadze, Y.Y.; Morozova, O.V. Alien species in the Holarctic. Russ. J. Biol. Invasions 2018, 9, 374–381. [Google Scholar] [CrossRef]
- Nazarov, M.I. 1927 Adventive flora of the middle and northern part of the RSFSR during the war and the revolution. Izv. Main Botan. Gard. 1927, 26, 223–234. (In Russian) [Google Scholar]
- Gusev, Y.D. New and rare adventitious plants of Leningrad and Karelian flora. Bot. Zh. 1968, 53, 267–269. (In Russian) [Google Scholar]
- Gusev, Y.D. Distribution of plants along railways of the North-West part of European Russia. Bot. Zh. 1971, 53, 347–360. (In Russian) [Google Scholar]
- Gusev, Y.D. New additions to adventive flora of north-west regions of European Russia. Bot. Zh. 1973, 58, 904–909. (In Russian) [Google Scholar]
- Gusev, Y.D. New data on adventive flora of Kaliningrad Region. Bot. Zh. 1974, 59, 1458–1460. (In Russian) [Google Scholar]
- Gusev, Y.D. New information on adventive plants distribution in the North-West of the USSR. Bot. Zh. 1975, 60, 380–387. (In Russian) [Google Scholar]
- Gusev, Y.D. Penetration of new adventive plants into Mari autonomous republic along the railway. Bot. Zh. 1977, 62, 429–431. (In Russian) [Google Scholar]
- Gusev, Y.D. New data on the adventive flora in different regions of taiga zone in the European part of the USSR. Bot. Zh. 1980, 65, 249–255. (In Russian) [Google Scholar]
- Tikhomirov, V.N. Contemporary issues of studying adventive and synanthropic plants. In Problems of Studying Adventitious Flora of the USSR: Proceedings of the Meeting, Moscow, 1989; Nauka: Moscow, Russia, 1989; pp. 3–6. (In Russian) [Google Scholar]
- Skvortsov, A.K. New data on the adventive flora of the Moscow region. 1. Bull. Main Botan. Gard. 1973, 87, 3–11. (In Russian) [Google Scholar]
- Skvortsov, A.K. New data on the adventive flora of the Moscow region. 2. Bull. Main Botan. Gard. 1973, 88, 31–35. (In Russian) [Google Scholar]
- Ignatov, M.S.; Makarov, V.V. News of the adventive flora of the Moscow region. Bull. Main Botan. Gard. 1984, 132, 49–51. (In Russian) [Google Scholar]
- Ignatov, M.S.; Makarov, V.V. Supplement to the adventive flora of the Moscow region. Bot. Zh. 1985, 70, 849–854. (In Russian) [Google Scholar]
- Ignatov, M.S.; Makarov, V.V.; Bochkin, V.D. On the naturalization of adventive species in the Moscow Region. Bot. Zh. 1988, 73, 438–442. (In Russian) [Google Scholar]
- Ignatov, M.S.; Makarov, V.V.; Chichev, A.V.; Shvetsov, A.N. Floristic finds in the railways of Moscow region. Bull. Main Botan. Gard. 1983, 129, 43–48. (In Russian) [Google Scholar]
- Makarov, V.V.; Ignatov, M.S. To the adventive flora of Moscow. Bull. Main Botan. Gard. 1983, 127, 38–42. (In Russian) [Google Scholar]
- Maytulina, Y.K. New and rare adventive plants of the Vologda Region. Bull. Main Botan. Gard. 1984, 132, 45–46. (In Russian) [Google Scholar]
- Problems of Studying Adventitious Flora of the USSR: Proceedings of the Meeting, Moscow, 1989; Tikhomirov, V.N.; Ignatov, M.S. (Eds.) Nauka: Moscow, Russia, 1989; pp. 1–138. (In Russian) [Google Scholar]
- Ignatov, M.S.; Makarov, V.V.; Chichev, A.V. Abstract of the flora of adventitious plants in the Moscow region. In Floristic Studies in the Moscow Region; Nauka: Moscow, Russia, 1990; pp. 5–105. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Fomina, O.V. Cenchrus longispinus (Hack.) Fernald—One more American “guest” in Central Black Soil Region. Protect. Quar. Plants 2010, 26–27, 27–28. [Google Scholar]
- Tokhtar, V.K.; Kurskoy, A.Y. Formation of the invasive component of the flora in Belgorod region for 170 years. Bot. Zh. 2020, 105, 854–860. (In Russian) [Google Scholar]
- Abramova, L.M.; Golovanov, Y.M. Invasive plants of the Republic of Bashkortostan: Blacklist, bibliography. RAS Ufa Res. Cent. Bull. 2016, 2, 54–61. (In Russian) [Google Scholar]
- Abramova, L.M.; Golovanov, Y.M.; Muldashev, A.A. Findings of invasive and potentially invasive plant species in the Western regions of the Republic of Bashkortostan. Nikitsky Botan. Gard. Bull. 2020, 137, 29–37. (In Russian) [Google Scholar] [CrossRef]
- Kapitonova, O.A. Alien plant species in aquatic and coastal-aquatic ecosystems of the Vyatka-Kama Urals. Russ. J. Biol. Invasions 2011, 4, 34–43. [Google Scholar]
- Silaeva, T.B. Alien species of the flora in the Sura River basin. Russ. J. Biol. Invasions 2011, 2, 250–255. [Google Scholar] [CrossRef]
- Senator, S.A.; Tokhtar, V.K..; Kurskoy, A.Y. Materials for the flora of railways of the Belgorod region. Udmurt. State Univ. Sci. Bull. Ser. Biol. Earth Sci. 2016, 26, 50–59. (In Russian) [Google Scholar]
- Dronin, G.V. Invasive plant species in the Syzranka River basin. Izv. Sarat. Un-ta Ser. Chem. Biol. Ecol. 2017, 17, 98–102. (In Russian) [Google Scholar] [CrossRef]
- Starodubtseva, E.A. Species extinction in protected areas (Voronezh reserve, 1935–2015). Russ. J. Ecosyst. Ecol. 2016, 1, 1–22. (In Russian) [Google Scholar] [CrossRef]
- Grigoryevskaya, A.Y.; Starodubtseva, E.A.; Vladimirov, D.R.; Lepeshkina, L.A.; Lisova, O.S. Evaluation of biological contamination of local flora in specially protected natural areas of Voronezh region. VSU Sci. Bull. Ser. Geogr. Geoecol. 2017, 1, 85–92. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Vinogradova, Y.K.; Zelenkova, V.N.; Kurskoy, A.Y. Can invasive plant species “differentiate” colonized ecotopes? Eurasia J. Biosci. 2020, 14, 2285–2292. [Google Scholar]
- Tokhtar, V.K.; Groshenko, S.A. Global alien species invasions: Research problems and perspectives. BSU Sci. Bull. Ser. Natur. Sci. 2008, 7, 50–54. [Google Scholar]
- Malysheva, V.G. Adventive Flora of the Kalinin Region. Cand. Sci. (Biol.) Dissertation, Komarov Botanical Institute, Leningrad, Russia, 1980. (In Russian). [Google Scholar]
- Puzyrev, A.N. Adventive flora of Udmurtia. Cand. Sci. (Biol.) Dissertation, Leningrad State University, Leningrad, Russia, 1986. (In Russian). [Google Scholar]
- Khorun, L.V. Adventive Flora of the Tula Region. Cand. Sci. (Biol.) Dissertation, Moscow State University, Moscow, Russia, 1998. (In Russian). [Google Scholar]
- Barmin, N.A. Adventive Flora of Mordovia Republic. Cand. Sci. (Biol.) Dissertation, Moscow State University, Moscow, Russia, 2000. (In Russian). [Google Scholar]
- Grigoryevskaya, A.Y.; Starodubtseva, E.A.; Khlyzova, N.Y.; Agafonov, V.A. Adventive Flora of Voronezh Region: Historical, Biogeographical, Ecological Aspects; Voronezh State University: Voronezh, Russia, 2004; pp. 1–319. (In Russian) [Google Scholar]
- Markelova, N.R. Dynamics of the Composition and Structure of Adventive Flora of the Tver Region. Cand. Sci. (Biol.) Dissertation, Tsitsin Main Botanical Garden RAS, Moscow, Russia, 2004. (In Russian). [Google Scholar]
- Senator, S.A.; Rakov, N.S.; Saksonov, S.V.; Vasyukov, V.M.; Ivanova, A.V. New and rare alien plants in the Middle Volga Region. Russ. J. Biol. Invasions 2013, 3, 98–104. [Google Scholar]
- Shaulo, D.N.; Zykova, E.Y. Findings of adventive species in the Novosibirsk Region. Plant. World Asian Russ. 2013, 1, 37–43. (In Russian) [Google Scholar]
- Borisova, E.A. Adventive Flora of Ivanovo; Ivanovo State University: Ivanovo, Russia, 2007; pp. 1–187. (In Russian) [Google Scholar]
- Borisova, E.A. Adventive Flora of the Upper Volga Region. (Current State, Dynamic Trends, Orientation of Formation Processes). Doctor Sci. (Biol.) Dissertation, Moscow State University, Moscow, Russia, 2008. (In Russian). [Google Scholar]
- Krylov, A.V. Adventive Component of the Kaluga Region. Flora: Dynamics and Naturalization of Species. Cand. Sci. (Biol.) Dissertation, Moscow State University, Moscow, Russia, 2008. (In Russian). [Google Scholar]
- Notov, A.A. Adventive Component of Tver Regional Flora: Dynamics of Composition and Structure; Tver State University Press: Tver, Russia, 2009; pp. 1–473. (In Russian) [Google Scholar]
- Rakov, N.S. Composition, Structure, and Dynamics of Adventive Flora of the Ulyanovsk Region. Cand. Sci. (Biol.) Dissertation, Institute of Ecology of Volga baisin RAS, Tolyatti, Russia, 2012. (In Russian). [Google Scholar]
- Mayorov, S.R.; Alekseev, Y.E.; Bochkin, V.D.; Nasimovich, Y.A.; Shcherbakov, A.V. Alien Flora of the Moscow Region: The Composition, Origin and the Vectors of Formation; KMK Scientific Press: Moscow, Russia, 2020; pp. 1–576. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Fomina, O.V.; Groshenko, S.A.; Samylovsky, V.A.; Petin, A.N. 2009 Floristic finds of adventive plant species in the Belgorod region. Probl. Region. Ecol. 2009, 1, 121–124. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Kurskoy, A.Y.; Zelenkova, V.N. New data on the flora of the Belgorod Region (based on the materials of 2018). Mosc. Soc. Nat. Bull. Sect. Biol. 2019, 124, 67–69. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Kurskoy, A.Y. Invasive Plants of the South-West. of the Central Russian Upland; Belgorod University: Belgorod, Russia, 2019; pp. 1–120. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Mayorov, S.R.; Khorun, L.V. The Black Book of Flora of Middle Russia; GEOS: Moscow, Russia, 2010; pp. 1–512. (In Russian) [Google Scholar]
- Panasenko, N.N. Blacklist of flora of Bryansk oblast. Russ. J. Biol. Invasions 2014, 5, 203–205. [Google Scholar] [CrossRef]
- Lepeshkina, L.A.; Kalaev, V.N. Invasive flora of the Prof. B.M. Kozo-Polyansky botanical garden Voronezh State University. VSU Sci. Bull. Ser. Geogr. Geoecol. 2012, 2, 32–35. [Google Scholar]
- Lepeshkina, L.A. Ecological and geographical features of plant invasions in the conditions of the Central Russian forest-steppe. Ekosistemy 2020, 21, 59–67. (In Russian) [Google Scholar] [CrossRef]
- Starodubtseva, E.A.; Morozova, O.V.; Grigoryevskaya, A.A. Materials for the Black Book of Voronezh oblast. Russ. J. Biol. Invasions 2014, 5, 206–216. [Google Scholar] [CrossRef]
- Borisova, E.A. Patterns of invasive plant species distribution in the Upper Volga basin. Russ. J. Biol. Invasions 2011, 2, 1–5. [Google Scholar] [CrossRef]
- Reshetnikova, N.M.; Mayorov, S.R.; Krylov, A.B. Black Book of the Kaluga Region. Vascular Plants; Vash Dom: Kaluga, Russia, 2019; pp. 1–342. (In Russian) [Google Scholar]
- Leostrin, A.V. Flora of the Kostroma River Basin. Cand. Sci. (Biol.) Dissertation, Komarov Botanical Institute RAS, St. Petersburg, Russia, 2019. (In Russian). [Google Scholar]
- Poluyanov, A.V. Flora of the Kursk Region; Kursk State University: Kursk, Russia, 2005; pp. 1–263. (In Russian) [Google Scholar]
- Vyukova, N.A. Adventive Flora of Lipetsk and Adjacent Regions. Cand. Sci. (Biol.) Dissertation, Moscow State University, Moscow, Russia, 1982. (In Russian). [Google Scholar]
- Khorun, L.V.; Kazakova, M.V. Floristic structure and naturalization of alien plant species of the Ryazan region flora. Udmurt. State Univ. Bull. Ser. Biol. Earth Sci. 2013, 2, 43–47. (In Russian) [Google Scholar]
- Notov, A.A.; Notov, V.A. Flora of Tver: Dynamics of Composition and Structure for the 200-Year; Tver State University Press: Tver, Russia, 2012; pp. 1–256. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Mayorov, S.R.; Notov, A.A. Black Book of Flora of the Tver Region: Alien Plant. Species in the Ecosystems of Tver Region; KMK Scientific Press: Moscow, Russia, 2011; pp. 1–292. (In Russian) [Google Scholar]
- Khorun, L.V.; Zakharov, V.G.; Sokoloff, D.D. Quantitative estimation of the adventive flora dynamics (an example of the Tula Region). Zh. Obshch. Biol. 2006, 67, 298–310. (In Russian) [Google Scholar]
- Borisova, M.A. Flora of Transport. Routes of the Yaroslavl Region. Cand. Sci. (Biol.) Dissertation, Mordovian State University, Saransk, Russia, 2002. (In Russian). [Google Scholar]
- Tremasova, N.A.; Borisova, E.A.; Borisova, M.A. Invasive plant species of the Yaroslavl Region. Yaroslavl Ped. Bull. Natur. Sci. 2012, 3, 103–111. (In Russian) [Google Scholar]
- Tremasova, N.A.; Borisova, E.A.; Borisova, M.A. Comparative analysis of the invasive components of the flora of the five regions of the Upper Volga Region. Yarosl. Ped. Bull. Natur. Sci. 2013, 4, 171–177. (In Russian) [Google Scholar]
- Geltman, D.V. The concept of “invasive species” and the need to study this phenomenon. In Proceedings of the Scientific Conference “Problems of the Study of Adventive and Synanthropic Flora in the CIS Regions”; Moscow State University: Moscow, Russia, 2003; pp. 35–36. (In Russian) [Google Scholar]
- Bobrov, Y.A.; Lukasheva, T.V.; Kuznetsova, Y.; Pozdeyeva, L.M. Alien Monocotyledons of Komi Republic. Phytodiver. East. Eur. 2017, 9, 75–99. (In Russian) [Google Scholar]
- Gubareva, U.I.; Glukhovsky, E.A. 2013 Impact of plant introduction on the diversity of natural flora and vegetation of the Kaliningrad region. In Proceedings of the Scientific Conference “The Role of Botanical Gardens in the Conservation of Plant Biodiversity”; Batumi Botanical Garden: Batumi, GA, USA, 2013; Volume 1, pp. 66–69. (In Russian). [Google Scholar]
- Popov, V.I. The Adventive Component of the Synanthropic Flora of St. Petersburg. Cand. Sci. (Biol.) Dissertation, Komarov Botanical Institute RAS, St. Petersburg, Russia, 2000. (In Russian). [Google Scholar]
- Eskina, T.G.; Akatov, V.V.; Akatova, T.V. Composition and species richness of fallow phytocenoses with alien species dominance (Belaya River Basin, Western Caucasus). Russ. J. Biol. Invasions 2012, 5, 55–67. [Google Scholar]
- Shhagapsoev, S.H.; Chadaeva, V.A.; Tsepkova, N.L.; Shhagapsoeva, K.A. Materials to the Black List of flora of the Central Caucasus (Within Kabardino-Balkarian Republic). Russ. J. Biol. Invasions 2018, 9, 384–391. [Google Scholar] [CrossRef]
- Komzha, A.L. Floral finds in North Ossetia. Bot. Zh. 2004, 89, 860–865. (In Russian) [Google Scholar]
- Akatov, V.V.; Shadzhe, A.E.; Akatova, T.V. Robinia pseudoacacia L. in the Western Caucasus. Russ. J. Biol. Invasions 2016, 7, 105–118. [Google Scholar] [CrossRef]
- Akatova, T.V.; Akatov, V.V. Elevational distribution of alien plant species in the Western Caucasus. Russ. J. Biol. Invasions 2019, 10, 205–219. [Google Scholar] [CrossRef]
- Afanasyev, V.E. Analysis of alien plants naturalization places in the Astrakhan Region. Astrakhan State Techn. Univ. Bull. 2008, 6, 238–241. (In Russian) [Google Scholar]
- Matveev, D.E. Alien plant species in forest communities of the Volgograd Region. Forest Bull. 2007, 6, 62–63. (In Russian) [Google Scholar]
- Sagalaev, V.A. To the inventory of invasive species of flora of the Volgograd Region. Tver State Univ. Bull. Ser. Biol. Ecol. 2013, 32, 102–105. (In Russian) [Google Scholar]
- Khapugin, A.A.; Vargot, E.V.; Chugunov, G.G.; Dementieva, A.E. Additions and remarks to alien flora of the Mordovian State Nature Reserve. Russ. J. Biol. Invasions 2013, 2, 60–71. [Google Scholar]
- Baranova, O.G.; Bralgina, E.N.; Koldomova, E.A.; Markova, E.M.; Puzyrev, A.S. The Black Book of Flora of the Udmurt Republic; Institute of Comprehensive Research: Izhevsk, Russia, 2016; pp. 1–67. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Vinogradova, Y.K.; Kurskoy, A.Y.; Zelenkova, V.N.; Tretyakov, M.Y. New data on the flora of the Nizhny Novgorod and Kirov Regions (based on the materials of 2020). Byul. MOIP. Otd. Biol. 2020, 125, 46–47. (In Russian) [Google Scholar]
- Mininzon, I.L.; Trostina, O.V. Black Book of the flora of the Nizhny Novgorod Region. In Alien Plant Species, Introduced and Cultivated, Actively Naturalized under the Conditions of the Nizhny Novgorod Region; The Seventh Electronic Version; Dront Ecocentre: Nizhny Novgorod, Russia, 2018; pp. 1–66, (In Russian). Available online: https://dront.ru/wp-content/uploads/2018/02/CHernaya-kniga-NNo-VII.pdf (accessed on 24 June 2021). (In Russian)
- Mininzon, I.L.; Solovyov, A.A.; Trostina, O.V. Black Book of the flora of the Nizhny Novgorod region. In Alien Plant Species, Introduced and Cultivated, Actively Naturalized under the Conditions of the Nizhny Novgorod Region; The Ninth Electronic Version; Dront Ecocentre: Nizhny Novgorod, Russia, 2020; pp. 1–70, (In Russian). Available online: https://dront.ru/wp-content/uploads/2020/03/CHernaya-kniga-NNo-IX.pdf (accessed on 24 June 2021). (In Russian)
- Vasyukov, V.M.; Novikova, L.A. Naturalized alien plants of the Penza region. Samara J. Sci. 2017, 6, 19–22. (In Russian) [Google Scholar] [CrossRef]
- Senator, S.A.; Saksonov, S.V.; Vasyukov, V.M.; Rakov, N.S. Invasive and potentially invasive plants of the Middle Volga region. Rus. J. Biol. Invasions 2017, 1, 57–69. [Google Scholar] [CrossRef]
- Mayevsky, V.V.; Boyakov, M.K.; Trunova, V.M.; Lyashenko, Z.D. Materials for the adventitious flora of the Saratov region. Saratov Univ. Botan. Gard. Bull. 2002, 1, 5–6. (In Russian) [Google Scholar]
- Tretyakova, A.S. Regularities of distribution of alien plants in anthropogenous habitats of Sverdlovsk region. Russ. J. Biol. Invasions 2015, 4, 118–128. [Google Scholar]
- Tretyakova, A.S. Invasive potential of alien plants in the Middle Urals. Russ. J. Biol. Invasions 2011, 2, 281–285. (In Russian) [Google Scholar] [CrossRef]
- Lashchinsky, N.N.; Korolyuk, A.Y.; Lashchinskaya, N.V.; Korolyuk, E.A. Finds of rare and imported species of vascular plants in Omsk, Novosibirsk and Tyumen regions and the Altai Territory. Turczaninowia 2010, 13, 117–123. (In Russian) [Google Scholar]
- Zykova, E.Y. Adventive flora of the Altai Republic. Plant. World Asian Russ. 2015, 3, 72–87. [Google Scholar]
- Ebel, A.L.; Kupriyanov, A.N.; Strelnikova, T.O.; Ankipovich, E.S.; Antipova, E.M.; Antipova, S.V.; Buko, T.E.; Verkhozina, A.V.; Doronkin, V.M.; Efremov, A.N.; et al. The Black Book of Flora of Siberia; GEO: Novosibirsk, Russia, 2016; pp. 1–439. (In Russian) [Google Scholar]
- Shaulo, D.N.; Doduk, A.D.; Molokova, N.I. Floristic finds in the Republic of Tyva. Turczaninowia 2003, 6, 35–42. (In Russian) [Google Scholar]
- Ebel, A.L. Floristic finds in the Republic of Khakassia. Turczaninowia 2002, 5, 44–48. (In Russian) [Google Scholar]
- Terekhina, T.A. Alien plants in the flora of the Altai Territory. In Problems of Studying the Vegetation Cover of Siberia; Tomsk State University Publishing House: Tomsk, Russia, 1995; pp. 60–61. (In Russian) [Google Scholar]
- Mishina, I.A.; Terekhina, T.A. On the behavior of some adventitious plants in the Altai Territory. In Proceedings of the International Conference “Problems of Studying the Adventive and Synanthropic Flora of Russia and Neighboring Countries”; Moscow State University Botan. Garden: Moscow, Russia, 2012; pp. 70–71. (In Russian) [Google Scholar]
- Kiseleva, A.A. Finds of introduced species in the flora of Central Siberia and the Krasnoyarsk Territory. Bot. Zh. 1988, 73, 1760–1761. (In Russian) [Google Scholar]
- Chepinoga, V.V.; Verkhozina, A.V. On the flora of ergasiophytes of the Irkutsk region. In Materials for the Flora of Baikal Siberia; Irkutsk University: Irkutsk, Russia, 2007; Volume 1, pp. 161–172. (In Russian) [Google Scholar]
- Strelnikova, T.A. Alien species in the flora of the Kemerovo region. In Botanical Studies of Siberia and Kazakhstan; KREOO: Kemerovo, Russia, 2013; Volume 19, pp. 114–122. (In Russian) [Google Scholar]
- Filippova, A.V.; Pinigina, Y.A.; Romanova, N.G.; Stepanyuk, G.Y.; Tarasova, I.V. Invasive plant species in Promyshlennovsky district, Kemerovo region. In Proceedings of XVIII International Scientific and Practical Conference, “Problems of Botany in Southern Siberia and Mongolia”; Altai State University: Barnaul, Russia, 2019; pp. 434–438. (In Russian) [Google Scholar]
- Muldiyarov, E.Y.; Pyak, A.I.; Ebel, A.L. New species of mossy and vascular plants in the flora of the Tomsk Region. Bot. Zh. 1996, 81, 90–93. (In Russian) [Google Scholar]
- Pyak, A.I. Adventive plants of the Tomsk Region. Bot. Zh. 1994, 79, 45–51. (In Russian) [Google Scholar]
- Karavaev, M.N. Synopsis of the Flora of Yakutia; Publishing House of the USSR Academy of Sciences: Moscow, Leningrad, Russia, 1958; pp. 1–192. (In Russian) [Google Scholar]
- Cherosov, M.M. Synanthropic Vegetation of Yakutia; Publishing house of the YANC SB RAS: Yakutsk, Russia, 2005; pp. 1–160. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Aistova, E.V.; Antonova, L.A.; Chernyagina, O.A.; Chubar, E.A.; Darman, G.F.; Devyatova, E.A.; Khoreva, M.G.; Kotenko, O.V.; Marchuk, E.A.; et al. Invasive plants in flora of the Russian Far East: The checklist and comments. Bot. Pac. 2020, 9, 103–129. [Google Scholar] [CrossRef]
- Nikolin, E.G. The Magadan tract as the main route of invasion by adventitious plants in the Eastern Verkhoyansk region. In Problems of Studying the Adventive and Synanthropic Flora of Russia and Neighboring Countries; IIKI: Izhevsk, Russia, 2012; pp. 151–154. (In Russian) [Google Scholar]
- Nikolin, E.G. The weed and alien plants of Yakutia. Russ. J. Biol Invasions 2014, 5, 86–89. [Google Scholar] [CrossRef]
- Nikolin, E.G. Weeds of Yakutia: The Most Dangerous and Aggressive Elements of the Flora; Nauka: Novosibirsk, Russia, 2016; pp. 1–264. (In Russian) [Google Scholar]
- Shishkin, I.K. Weed Plants of the Southern Part of the Far Eastern Region; Far Eastern Regional State Publishing House: Khabarovsk, Russia, 1936; pp. 1–144. [Google Scholar]
- Vorobyov, D.P. On the issues of drift and weed plants in the Primorsky Territory. In V.L. Komarov Memorial Lectures; Dalnauka: Vladivostok, Russia, 1954; Volume 4, pp. 3–22. (In Russian) [Google Scholar]
- Basargin, D.D. Adventitious species of the genus Euphorbia (Euphorbiaceae) in the Far East. Bot. Zh. 1982, 67, 112–114. (In Russian) [Google Scholar]
- Nechaeva, T.I. To the knowledge of the adventitious flora of Vladivostok. Bull. Main Botan. Gard. 1976, 102, 40–44. (In Russian) [Google Scholar]
- Nechaeva, T.I. New information on the adventitious flora of Vladivostok. Bull. Main Botan. Gard. 1981, 121, 54–55. (In Russian) [Google Scholar]
- Nechaeva, T.I. Adventive flora of the Primorsky Territory. In V.L. Komarov Memorial Lectures; Dalnauka: Vladivostok, Russia, 1984; Volume 31, pp. 46–88. (In Russian) [Google Scholar]
- Nechaeva, T.I. On new adventitious plant species in Vladivostok. Bull. Main Botan. Gard. 1987, 146, 43–44. (In Russian) [Google Scholar]
- Nechaeva, T.I. New and rare adventitious plants for the flora of the Primorsky Territory. Bot. Zh. 1992, 77, 129–131. (In Russian) [Google Scholar]
- Nechaeva, T.I. Adventitious Plants of the Primorsky Territory; Dalnauka: Vladivostok, Russia, 1998; pp. 1–264. (In Russian) [Google Scholar]
- Nechaeva, T.I.; Verkholat, V.P.; Polyychuk, Y.S. On the distribution of some drift plants in the south of Primorye. Bot. Zh. 1978, 63, 387–389. (In Russian) [Google Scholar]
- Ulyanova, T.N. Segetal flora of the Primorsky Territory. Bot. Zh. 1978, 63, 1004–1016. (In Russian) [Google Scholar]
- Ignatov, M.S. Supplement to the adventitious flora of the Far East. Bot. Zh. 1986, 71, 1130–1134. (In Russian) [Google Scholar]
- Probatova, N.S.; Sokolovskaya, A.P.; Rudyka, E.G. Chromosome numbers and distribution of some adventitious and weed plant species in Primorsky Krai and Sakhalin. Izv. Sib. Otd. Acad. Sci. USSR Ser. Biol. 1986, 13, 63–68. (In Russian) [Google Scholar]
- Kozhevnikov, A.E.; Kozhevnikova, Z.V. Alien plant species in the flora of the Russian Far East and regional patterns of their geographical differentiation. RAS Far East. Branch Bull. 2014, 3, 12–19. (In Russian) [Google Scholar]
- Kozhevnikov, A.E.; Kozhevnikova, Z.V. Complex of adventitious plant species as a component of the natural flora of the Russian Far East: Diversity and spatial changes in the taxonomic structure. In V.L. Komarov Memorial Lectures; Dalnauka: Vladivostok, Russia, 2011; Volume 58, pp. 5–36. (In Russian) [Google Scholar]
- Chubar, E.A. Alien species in the small marine island flora: Life-strategic, coenotic activity, adventive index (The Far Eastern Marine Reserve, Primorskii krai). In V.L. Komarov Memorial Lectures; Dalnauka: Vladivostok, Russia, 2015; Volume 63, pp. 128–163. (In Russian) [Google Scholar]
- Gularyants, G.M. Anthropophytes of the Dalnegorsk urban district (the Primorsky Territory). RAS BSI FEB Bull. Sci. J. 2017, 17, 1–20. (In Russian). Available online: http://botsad.ru/media/cms/3765/1-20.pdf. (accessed on 24 June 2021).
- Nedoluzhko, V.A. Distribution of Ambrosia artemisiifolia (Asteraceae) in Primorsky Krai. Bot. Zh. 1984, 69, 527–529. (In Russian) [Google Scholar]
- Kolyada, N.A. Robinia (Robinia pseudoacacia L., Fabaceae Lindl.) as part of a secondary plant community in the Primorsky Territory. Astrakh. Bull. Ecol. Educ. 2020, 3, 190–196. (In Russian) [Google Scholar]
- Shlotgauer, S.D.; Nebaykin, V.D. To the knowledge of the adventitious flora of the southern part of the Khabarovsk Territory. Bull. Main Botan. Gard. 1984, 113, 42–45. (In Russian) [Google Scholar]
- Antonova, L.A. Synopsis of the Adventive Flora of the Khabarovsk Territory; FEB RAS: Khabarovsk, Russia, 2009; pp. 1–93. (In Russian) [Google Scholar]
- Antonova, L.A. Invasive component of flora in Khabarovsk krai. Russ. J. Biol. Invasions 2013, 4, 69–73. [Google Scholar] [CrossRef]
- Antonova, L.A. Adventive component of the flora of the sea coast of the Khabarovsk Territory. In V.L. Komarov Memorial Lectures; Dalnauka: Vladivostok, Russia, 2014; Volume 62, pp. 63–105. (In Russian) [Google Scholar]
- Antonova, L.A. Current state of the alien component in the flora of Khabarovsk Territory. Region. Probl. 2017, 20, 5–12. (In Russian) [Google Scholar]
- Kudrin, S.G. Adventive flora of the Khingansky State Reserve and its environs. Bull. Main Botan. Gard. 1991, 160, 23–27. (In Russian) [Google Scholar]
- Starchenko, V.M. Synopsis of the flora of the Amur region. In V.L. Komarov Memorial Lectures; Dalnauka: Vladivostok, Russia, 2001; Volume 48, pp. 5–55. (In Russian) [Google Scholar]
- Starchenko, V.M. Adventive flora of the Bureya Valley (Amur Region). Sci. Not. ZabGU 2016, 11, 52–58. (In Russian) [Google Scholar]
- Aistova, E.V. Synopsis of the adventive flora of the Amur region. Turczaninowia 2009, 12, 17–40. (In Russian) [Google Scholar]
- Starchenko, V.M.; Timchenko, N.A.; Bobenko, V.F. About the naturalization of the Nirrhae rhamnoides L. in the vicinity of Blagoveshchensk (Amur region). Sci. Not. Trans. Baikal. Univ. Ser. Biol. 2017, 12, 114–120. (In Russian) [Google Scholar]
- Bury, V.V. New types of adventitious plants on the territory of the Bystrinsky Nature Park (Bystrinsky District, Kamchatka Krai). Bot. Zh. 2016, 101, 294–302. (In Russian) [Google Scholar]
- Devyatova, E.A.; Chernyagina, O.A.; Abramova, L.M. Synopsis of the advent flora of the city of Petropavlovsk-Kamchatsky. Bull. North. East. Univ. 2016, 4, 5–16. (In Russian) [Google Scholar]
- Chernyagina, O.A.; Devyatova, E.A. Adventitious plants of the Kamchatka Territory: Distribution and diversity. In Proceedings of the International Conference “Conservation of the Biodiversity of Kamchatka and Adjacent Seas”; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2018; pp. 92–101. (In Russian). [Google Scholar]
- Chernyagina, O.A.; Shtreker, L. Invasive species in the flora of Kamchatka. Natur. Techn. Sci. 2012, 6, 150–152. (In Russian) [Google Scholar]
- Abramova, L.M.; Chernyagina, O.A.; Devyatova, E.A. Invasive species in Kamchatka: Distribution and communities. Bot. Pac. 2017, 6, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Ulyanova, T.N. New drift plants of the Magadan region. Bot. Zh. 1977, 62, 694–696. (In Russian) [Google Scholar]
- Khokhryakov, A.P. Flora of the Magadan Region; Nauka: Moscow, Russia, 1985; pp. 1–395. (In Russian) [Google Scholar]
- Lysenko, D.S. Alien plants. In Flora and Vegetation of the Magadan region (Synopsis of Vascular Plants and an Outline of Vegetation); IBPS FEB RAS: Magadan, Russia, 2010; pp. 235–289. (In Russian) [Google Scholar]
- Lysenko, D.S. Synanthropic Flora of the Magadan Region; SVNTs FEB RAS: Magadan, Russia, 2012; pp. 1–111. (In Russian) [Google Scholar]
- Khoreva, M.G. Floristic finds in the south of the Magadan region. Byul. MOIP. Otd. Biol. 2018, 123, 71–74. (In Russian) [Google Scholar]
- Ulyanova, T.N. New drift plants in the flora of Sakhalin island. Byul. VIR 1976, 65, 78–90. (In Russian) [Google Scholar]
- Fukuda, T.; Taran, A.; Sato, H.; Kato, Y.; Takahashi, H. Alien plants collected or confirmed on the islands of Shikotan, Kunashir and Iturup on the 2009–2012 Botanical Expeditions. Bull. Hokkaido Univ. Mus. 2014, 7, 8–26. [Google Scholar]
- Fukuda, T.; Kato, Y.; Sato, H.; Taran, A.A.; Barkalov, V.Y.; Takahashi, H. Naturalization of Cakile edentula (Brassicaceae) on the beaches of Kunashiri and Etorofu Islands—The first record for species from the Kuril Islands. J. Jpn. Bot. 2013, 88, 124–128. [Google Scholar]
- Rubtsova, T.A. Adventive element of the flora of the Small Khingan. In Proceedings of the Symposium “The Human Dimension in Regional Development”; ICARP FEB RAS: Birobidzhan, Russia, 2000; pp. 194–196. (In Russian) [Google Scholar]
- Rubtsova, T.A.; Antonova, L.A. Adventive component of the flora of the Jewish Autonomous Region: Current state and additions. Region. Probl. 2020, 23, 12–22. (In Russian) [Google Scholar] [CrossRef]
- Yurtsev, B.A.; Koroleva, T.M.; Petrovsky, V.V.; Polozova, T.G.; Zhukova, P.G.; Katenin, A.E. Summary of the Flora of the Chukchi tundra; VVM: St. Petersburg, Russia, 2010; pp. 1–628. (In Russian) [Google Scholar]
- Fomina, Z.V. On new types of weed plants in the Ulan-Ude district of the Buryat ASSR. Sci. Not. He Buryat Agr. Inst. Agron. 1965, 18, 62–66. (In Russian) [Google Scholar]
- Sutkin, A.V. Urban. Flora of the City of Ulan-Ude; Publishing house of the BNC SB RAS: Ulan-Ude, Russia, 2010; pp. 1–142. (In Russian) [Google Scholar]
- Anenkhonov, O.A.; Pykhalova, T.D. Check-List of Vascular Plants Flora of Zabaikalskyi National Park; BNC SB RAS: Ulan-Ude, Russia, 2010; pp. 1–228. (In Russian) [Google Scholar]
- Pazdnikova, N.M.; Chepinoga, V.V. Synopsis of the flora of vascular plants of the central part of Dauria Ononskaya (South-Eastern Transbaikalia, Russia). Izv. Irkut. Univ. Ser. Biol. Ecol. 2013, 6, 32–60. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Tokhtar, V.K.; Zelenkova, V.N.; Galkina, M.A.; Tretyakov, M.Y.; Kurskoy, A.Y.; Stogova, A.V. Flora of the Trans-Siberian railway and its correlation with the characteristics of natural biomes on the territory of the East European Plain. Tver State Univ. Bull. Ser. Biol. Ecol. 2020, 4, 61–82. [Google Scholar]
- Tokhtar, V.K.; Vinogradova, Y.K.; Kurskoy, A.Y.; Zelenkova, V.N.; Tretyakov, M.Y. Flora of railway stations of the Trans-Siberian railway within the Nizhny Novgorod Region. Tver State Univ. Bull. Ser. Biol. Ecol. 2020, 3, 102–114. (In Russian) [Google Scholar] [CrossRef]
- Notov, A.A.; Vinogradova, Y.K.; Mayorov, S.R. On the problem of development and management of regional black books. Russ. J. Biol. Invasions 2011, 2, 35–45. [Google Scholar] [CrossRef]
- Vinogradova, Y.K. “Black hundred” of invasive plants of Russia. In Newsletter of the Council of Botanical Gardens of the CIS Countries at the International Association of Academies of Sciences; Nauchtekhlitizdat: Moscow, Russia, 2015; Volume 4, pp. 85–89. (In Russian) [Google Scholar]
- Vinogradova, Y.; Pergl, J.; Hejda, M.; Pyšek, P.; Essl, F.; Regional Contributors; van Kleunen, M. Invasive alien plants of Russia: Insights from regional inventories. Biol. Invasions 2018, 20, 1931–1943. [Google Scholar] [CrossRef] [Green Version]
- Petrosyan, V.; Dgebuadze, Y.; Khlyap, L.; Vinogradova, Y.; Krivosheina, M.; Feniova, I.; Bashinskiy, I.; Reshetnikov, A.; Omelchenko, A.; Goryaynova, Z.; et al. Global Register of Introduced and Invasive Species-Russian Federation. Version 2.7. Invasive Species Specialist Group ISSG. Checklist Dataset. GRIIS IPT. 2020. Available online: https://doi.org/10.15468/f6joyb (accessed on 24 June 2021).
- The Most Dangerous Invasive Species of Russia (TOP-100); Dgebuadze, Y.Y.; Petrosyan, V.G.; Klyap, L.A. (Eds.) KMK Scientific Press: Moscow, Russia, 2018; pp. 1–688. (In Russian) [Google Scholar]
- Tokhtar, V.K. Advanced approach to the visualization of data characterizing distribution features of alien plant species. Russ. J. Biol. Invasions 2018, 9, 263–269. [Google Scholar] [CrossRef]
- Tokhtar, V.K.; Groshenko, S.A. Differentiation of the climatic niches of the invasive Oenothera l. (Subsect. Oenothera, Onagraceae) Species in the Eastern Europe. Adv. Environ. Biol. 2014, 8, 529–531. [Google Scholar]
- Egoshin, A.V. Modeling of the spatial distribution of alien plant species using remote sensing data on the example of Paulownia tomentosa. Voronezh State University Bull. Ser. Geogr. Geoecol. 2020, 1, 39–47. (In Russian) [Google Scholar]
- Egoshin, A.V. Bioclimatic and ecological-geographical requirements for alien plant species of Asian origin naturalized in the south of the Russian Black Sea Region. In Plants in the Monsoon Climate: Anthropogenic and Climatogenic Transformation of Flora and Vegetation: Proceedings of VIII All-Russian Scientific Conference, Blagoveshchensk, 2018; Far Eastern State Agrarian University: Blagoveshchensk, Russia, 2018; pp. 59–64. (In Russian) [Google Scholar]
- Morozova, O.V. Naturalized alien species in the floras of the Middle part of European Russia: Homogenization or differentiation? Russ. J. Biol. Invasions 2018, 3, 88–98. [Google Scholar] [CrossRef]
- Sennikov, A.N. Memories of war times: War-time plant immigrants (polemochores) in East Fennoscandia and North-West Russia. In Problems of Studying the Adventive and Synanthropic Flora of Russia and Neighboring Countries; IIKI: Izhevsk, Russia, 2012; pp. 182–185. (In Russian) [Google Scholar]
- Reshetnikova, N.M. The way of emergence of some Western European plant species in Kaluga oblast—The pathway of the German army in 1941–1943. Russ. J. Biol. Invasions 2016, 7, 62–68. [Google Scholar] [CrossRef]
- Reshetnikova, N.M.; Shcherbakov, A.V.; Fadeeva, I.A. On the flora of “Krasny Bor”, a unique protected area of the Smolensk Region. Tver State Univ. Bull. Ser. Biol. Ecol. 2019, 4, 160–190. (In Russian) [Google Scholar] [CrossRef]
- Reshetnikova, N.M.; Shcherbakov, A.V.; Korolkova, E.O. Central European species in vicinity of Kobelevo village (Kaluga Region) as the traces of the Great Patriotic War. Bot. Zh. 2019, 104, 1122–1134. (In Russian) [Google Scholar]
- Shcherbakov, A.V.; Reshetnikova, N.M. Where to look for polemochorous plants in the Smolensk region? In The Study of the Adventive and Synanthropic Flora of Russia and Neighboring Countries: Results, Problems, and Prospects: Proceedings of the International Scientific Conference 2017; Institute of comprehensive research: Moscow, Izhevsk, Russia, 2017; pp. 134–137. (In Russian) [Google Scholar]
- Reshetnikova, N.M.; Notov, A.A.; Mayorov, S.R.; Shcherbakov, A.V. The Great Patriotic War as a factor of florogenesis: Results of the search for polemochores in Central Russia. Zh. Obshch. Biol. 2021, 82, 1–25. (In Russian) [Google Scholar]
- Notov, A.A.; Notov, V.A.; Zueva, L.V.; Ivanova, S.A. Succession dynamics of phytocenoses with polemohora-species. Field Biol. J. 2020, 2, 260–271. (In Russian) [Google Scholar]
- Notov, A.A.; Notov, V.A.; Zueva, L.V.; Andreeva, E.A. Polemochores of the Tver Region and the problem of biological invasions. Diver. Plant. World 2019, 3, 22–27. (In Russian) [Google Scholar] [CrossRef]
- Vinogradova, Y.K.; Kuklina, A.G.; Mukhina, L.N. Entomo-phytopathological status of golden currant (Ribes aureum Pursh) populations in secondary distribution range. In Tree Plants: Fundamental and Applied Research; Main Botanical Garden RAS: Moscow, Russia, 2011; pp. 120–127. (In Russian) [Google Scholar]
- Vinogradova, Y.K. Code of Conduct for Botanic Gardens on Invasive Alien Species; Main Botanical Garden RAS: Moscow, Russia, 2015; pp. 1–68. (In Russian) [Google Scholar]
- Notov, A.A.; Notov, V.A. Some results of analysis of invasive activity of species in the Botanical garden of Tver State University. In Problems of the Study of Adventive and Synanthropic Flora of Russia and Neighboring Countries: Proceedings of the International Scientific Conference 2012; Institute of comprehensive research: Izhevsk, Russia, 2012; pp. 154–157. (In Russian) [Google Scholar]
- Lepeshkina, L.A.; Mukovnina, Z.P. Adventive flora of the Botanical Garden and the extent of its naturalization. In Proceedings of the Scientific Conference “Botanical Gardens as Centers for the Conservation of Biodiversity and Rational Use of Plant Resources”; Tsitsin Main Botanical Garden RAS: Moscow, Russia, 2005; pp. 300–302. (In Russian) [Google Scholar]
- Shmarayeva, A.N.; Shishlova, Z.N.; Kuzmenko, I.P. Adventive grassy flora of the Botanical garden of Southern Federal University. In Proceedings of the XVIII International Scientific and Practical Conference “ Problems of Botany in Southern Siberia and Mongolia”; Altai State University: Barnaul, Russia, 2019; pp. 644–649. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Mayorov, S.R.; Yatsenko, I.O. Spontaneous Flora of the Main Botanical Garden as a Reflection of the Dynamics of Alien Species Invasion into Natural Ecosystems; KMK Scientific Press: Moscow, Russia, 2020; pp. 1–540. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Mayorov, S.R.; Bochkin, V.D. Effect of alien plant species on flora dynamics in the Main Botanical Garden of the Russian Academy of Sciences. Russ. J. Biol. Invasions 2016, 7, 12–25. [Google Scholar] [CrossRef]
- Maytulina, Y.K. Intraspecific Variability of Some Plants Introduced from North America. Cand. Sci. (Biol.) Dissertation, Tsitsin Main Botanical Garden RAS, Moscow, Russia, 1980. (In Russian). [Google Scholar]
- Vinogradova, Y.K. Processes of Microevolution in Adventitious and Introduced Plants. Doctor Sci. (Biol.) Dissertation, Tsitsin Main Botanical Garden RAS, Moscow, Russia, 1992. (In Russian). [Google Scholar]
- Vinogradova, Y.K.; Kuklina, A.G.; Galkina, M.A. The Dynamics of Clonal Dispersal and Regenerative Activity of Veronica filiformis J.E. Smith. Russ. J. Biol. Invasions 2017, 8, 197–205. [Google Scholar] [CrossRef]
- Dietrich, W.; Wagner, W.L.; Raven, P.H. Systematics of Oenothera section Oenothera subsection Oenothera (Onagraceae). Syst. Bot. Monogr. 1997, 50, 1–234. [Google Scholar] [CrossRef]
- Tokhtar, V.K.; Vinogradova, Y.K.; Groshenko, A.S. Microevolution and Invasiveness of Oenothera L. Species (Subsect. Oenothera, Onagraceae) in Europe. Russ. J. Biol. Invasions 2011, 2, 273–280. [Google Scholar] [CrossRef]
- Galkina, M.A.; Vinogradova, Y.K. Analysis of biomorphological traits and competitive ability of two invadere species in the genus Conyza Less. Bull. Main Botan. Gard. 2011, 197, 3–15. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Mazur, N.V. Morphological features study in Conyza canadensis (L.) Cronq. populations under the southwest of Central Russian Upland conditions. BSU Sci. Bull. Ser. Natur. Sci. 2011, 15, 249–253. (In Russian) [Google Scholar]
- Tokhtar, V.K.; Mazur, N.V. Variability of correlation structures of morphological features in populations of Conyza canadensis (L.) Cronq. BSU Sci. Bull. Ser. Natur. Sci. 2011, 15, 254–258. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Kuklina, A.G.; Tkacheva, E.V. Invasive Species of Fabaceae Family. Lupinus, Galega, Robinia, Amorpha, Caragana; ABF: Moscow, Russia, 2014; pp. 1–304. (In Russian) [Google Scholar]
- Tkacheva, E.V.; Vinogradova, Y.K.; Pavlova, I.V. Variability of morphometric characteristics of Galega orientalis Lam. in some populations of natural and secondary range. Russ. J. Biol. Invasions 2011, 2, 268–272. [Google Scholar] [CrossRef]
- Vinogradova, Y.K.; Kuklina, A.G.; Tkacheva, E.V.; Ryabchenko, A.S.; Khomutovskiy, M.I.; Shelepova, O.V. Comparative floral and pollen morphology of some invasive and native Impatiens species. Rev. Univ. Zulia 2020, 11, 315–335. Available online: https://www.produccioncientificaluz.org/index.php/rluz/article/view/32820. (accessed on 24 June 2021).
- Vinogradova, Y.K. Bio-morphological characters of alien legume species, influencing their invasion in natural plant communities. Am. J. Plant. Sci. 2016, 7, 2390–2398. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, Y.K.; Ryabchenko, A.S.; Galkina, M.A. The taxonomic relevance of achenial trichomes in invasive European Asteraceae. Ann. Mo. Bot. Gard. 2017, 102, 730–741. [Google Scholar] [CrossRef]
- Vinogradova, Y.K.; Grygorieva, O.V.; Vergun, E.N. Stomatal structure in Symphyotrichum nees species as an additional index of invasiveness. Russ. J. Biol. Invasions 2021, 12, 27–35. [Google Scholar] [CrossRef]
- Vinogradova, Y.; Shelepova, O.; Vergun, O.; Grygorieva, O.; Kuklina, A.; Brindza, J. Differences between Aronia Medik. taxa on the morphological and biochemical characters. Environ. Res., Engin. Manag. 2018, 74, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Shelepova, O.V.; Vinogradova, Y.K. Chemical composition of oil in the seed of Echinocystis lobata (Mich.) Torr. et Gray. Skvortsovia 2017, 3, 88–90. (In Russian) [Google Scholar]
- Vinogradova, Y.; Vergun, E.; Grygorieva, O.; Brindza, J. Antioxidant (free radical scavenging) activity of Solidago L. complex. Acta Hortic. 2021, in press. [Google Scholar]
- Galkina, M.; Vinogradova, Y. Hybridogenic activity of invasive species of Asteraceae. In Invasive Species—Introduction Pathways, Economic Impact, and Possible Management Options; El-Shafie, H., Ed.; InTechOpen Limited: London, UK, 2020; pp. 1–253. Available online: https://www.intechopen.com/books/invasive-species-introduction-pathways-economic-impact-and-possible-management-options/hybridogenic-activity-of-invasive-species-of-asteraceae (accessed on 14 June 2021).
- Vinogradova, Y.K. Experiments on reduction of abundance of invasive species (Impatiens glandulifera, Solidago gigantea, Adenocaulon adhaerescens). In Bridging the Gap between Scientific Knowledge and Management Practice, Proceedings of the 11th International Conference on the Ecology and Management of Alien Plant Invasions (EMAPi), 2011; Cultural and Youth Centre of Vas County: Szombathely, Hungary, 2011; p. 111. [Google Scholar]
- Vinogradova, Y.K.; Kuklina, A.G. Resource Potential of Invasive Plant. Species; GEOS: Moscow, Russia, 2012; pp. 1–185. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Kuklina, A.G.; Brindza, J. Invasive Plant. Species Used for Economic Purposes and Human Health; Slovak University of Agriculture in Nitra: Nitra, Slovakia, 2019; pp. 1–163. (In Russian) [Google Scholar]
- Dalke, I.V.; Chadin, I.F.; Zakhozhiy, I.G. Control of Sosnowskyi’s Hogweed (Heracleum sosnowskyi Manden.) Invasion on the Territory of the Russian Federation. Russ. J. Biol. Invasions 2018, 9, 331–344. [Google Scholar] [CrossRef]
- Baranova, O.G.; Shcherbakov, A.V.; Senator, S.A.; Panasenko, N.N.; Sagalaev, V.A.; Saksonov, S.V. The main terms and concepts used in the study of alien and synanthropic flora. Phytodiver. East. Eur. 2018, 12, 4–22. (In Russian) [Google Scholar]
- Vinogradova, Y.K. Adenocaulon adhaerescens Maxim. In Biology and Ecology of Plants in the Russian Far East; Far Eastern Federal University: Ussuriysk, Russia, 2013; pp. 99–106. (In Russian) [Google Scholar]
- Mayorov, S.; Serebryanyi, M.; Vinogradova, Y. Expansion of invasive Adenocaulon adhaerescens Maxim. (Asteraceae) in Moscow Region. Bioinvas. Rec. 2021, in press. [Google Scholar]
- Puchałka, R.; Wyborska, D.; Rutkowski, L.; Piwczyński, M. Pilosella bauhinii (Schult.) Arv-Touv. and P. cymosa subsp. vaillantii (Tausch) S. Braut. and Greuter (Asteraceae) from new localities in north-central Poland. Acta Soc. Bot. Pol. 2015, 84, 449–451. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, Y.K.; Notov, A.A.; Bochkin, V.D.; Barinov, A.V.; Notov, V.A. Rare and endangered plant species on the railways of the Moscow city and Tver region. Tver State Univ. Bull. Ser. Biol. Ecol. 2018, 3, 130–164. (In Russian) [Google Scholar] [CrossRef]
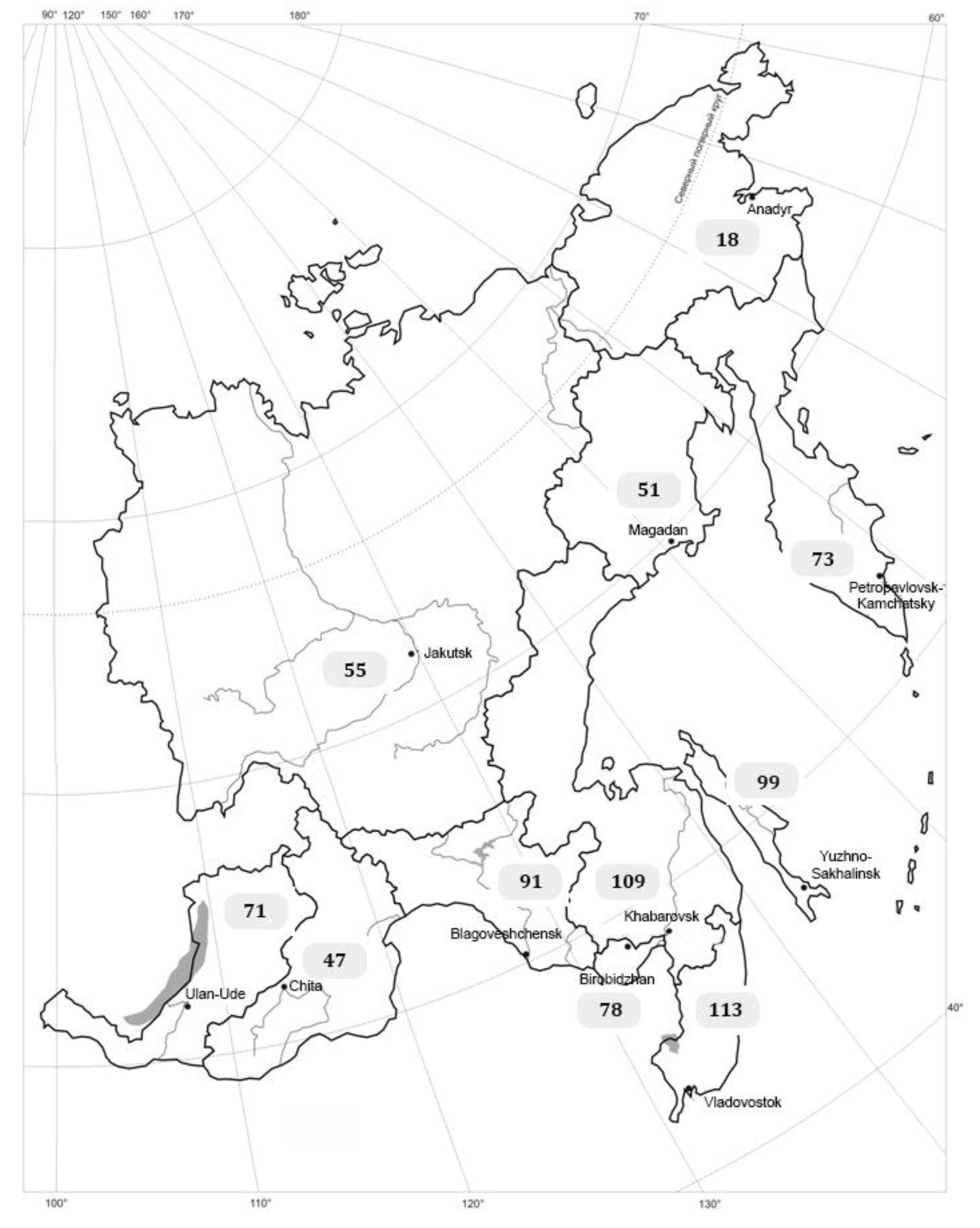

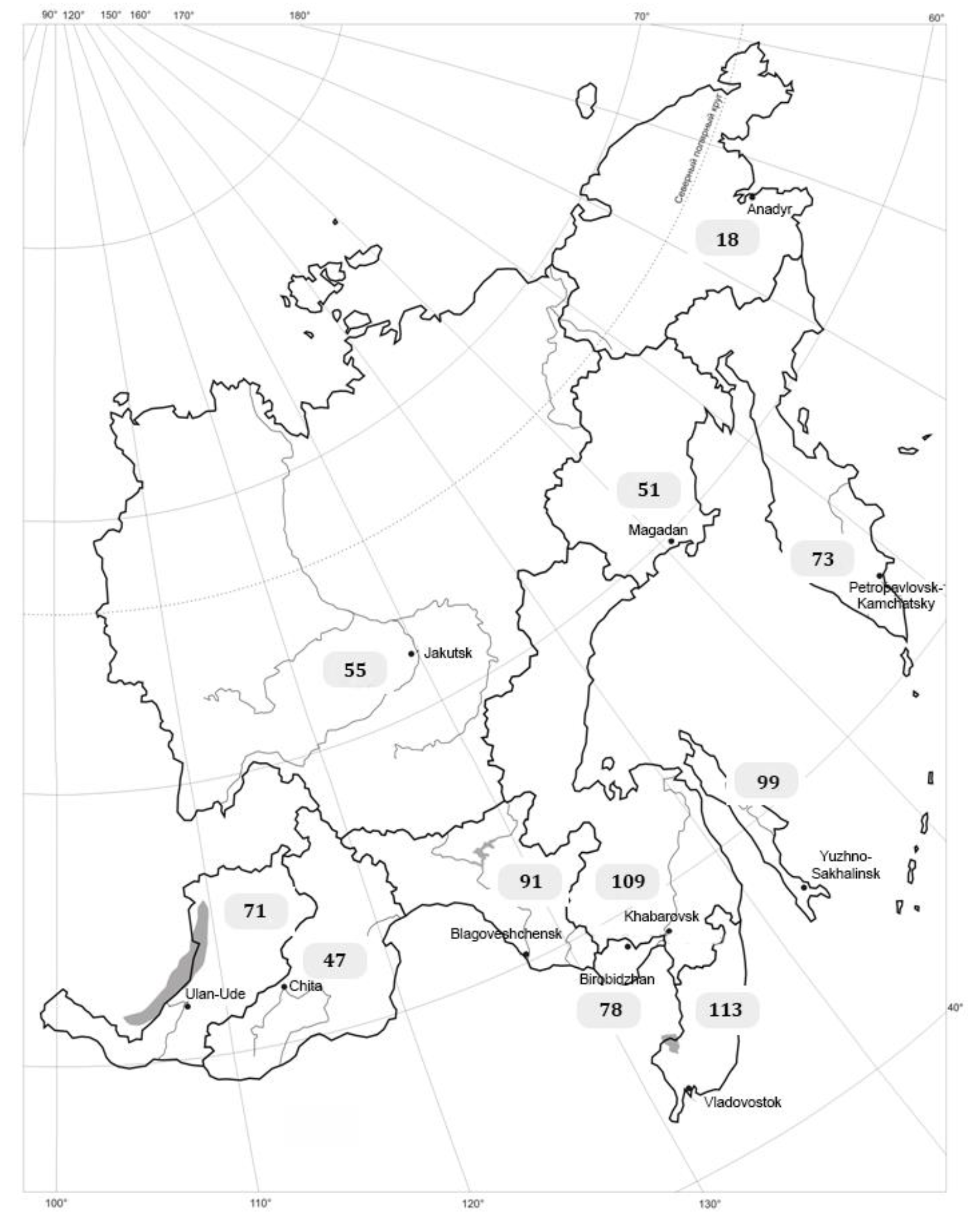{kind=link}
| Administrative Subjects of Russia | Area, ths. km2 | Area, % of Country | Papers on Alien Plant Species | Papers on Invasive Plant Species |
|---|---|---|---|---|
| Central Federal District | ||||
| Belgorod Region | 27,134 | 0.16 | [33,49,50] | [24,51,52] |
| Bryansk Region | 34,857 | 0.20 | Data are fragmentary | [52,53] |
| Vladimir Region | 29,084 | 0.17 | [44] | [52] |
| Voronezh Region | 52,216 | 0.30 | [39] | [52,54,55,56] |
| Ivanovo Region | 21,437 | 0.13 | [43,44] | [52,57] |
| Kaluga Region | 29,777 | 0.17 | [45] | [52,58] |
| Kostroma Region | 60,211 | 0.35 | [59] | [52] |
| Kursk Region | 29,997 | 0.18 | [60] | [52] |
| Lipetsk Region | 24,047 | 0.14 | [61] | [52] |
| Moscow Region | 44,329 | 0.26 | [13,14,15,16,22,48] | [17,48] |
| Orel Region | 24,652 | 0.14 | Data are fragmentary | [52] |
| Ryazan Region | 39,605 | 0.23 | [62] | [52] |
| Smolensk Region | 49,790 | 0.29 | Data are fragmentary | [52] |
| Tambov Region | 34,462 | 0.20 | Data are fragmentary | [52] |
| Tver Region | 84,201 | 0.49 | [35,40,44,46,63] | [52,63,64] |
| Tula Region | 25,679 | 0.15 | [37,65] | [52] |
| Yaroslavl Region | 36,177 | 0.21 | [44,66] | [52,67,68] |
| The City of Moscow | 2,561 | 0.01 | [19,48] | [48] |
| Total | 650,205 | 3.78 | All the regions have been explored sufficiently | |
| The North-West Federal District | ||||
| Republic of Karelia | 180,520 | 1.05 | [5] | [69] |
| Republic of Komi | 416,774 | 2.3 | [70] | No data available |
| Arkhangelsk Region | 589,913 | 3.44 | No data available | No data available |
| Vologda Region | 144,527 | 0.84 | No data available | No data available |
| Kaliningrad Region | 15,125 | 0.09 | [8] | [71] |
| Leningrad Region | 83,908 | 0.49 | [5,6,7] | [69] |
| Murmansk Region | 144,902 | 0.85 | No data available | No data available |
| Novgorod Region | 54,501 | 0.32 | Data are fragmentary | [69] |
| Pskov Region | 55,399 | 0.32 | Data are fragmentary | [69] |
| The City of Saint Petersburg | 1403 | 0.01 | [72] | No data available |
| Total | 1,686,972 | 9.71 | 7.44% of the territory remain underexplored | |
| The South Federal District | ||||
| Republic of Adygeya | 7792 | 0.05 | [73] | Data are fragmentary |
| Republic of Daghestan | 50,270 | 0.29 | No data available | No data available |
| Republic of Ingushetia | 3123 | 0.02 | No data available | No data available |
| Kabardino-Balkarian Republic | 12,470 | 0.07 | No data available | [74] |
| Republic of Kalmykia | 74,731 | 0.44 | No data available | No data available |
| Karachayevo-Circassian Republic | 14,277 | 0.08 | No data available | No data available |
| Republic of North Ossetia-Alania | 7987 | 0.05 | No data available | [75] |
| Chechen Republic | 16,171 | 0.09 | No data available | No data available |
| Krasnodar Territory | 75,485 | 0.44 | No data available | [76,77] |
| Stavropol Territory | 66,160 | 0.39 | No data available | No data available |
| Astrakhan Region | 49,024 | 0.29 | [78] | Data are fragmentary |
| Volgograd Region | 112,877 | 0.66 | [79] | [80] |
| Rostov Region | 100,967 | 0.59 | No data available | No data available |
| Total | 447,821 | 3.46 | 1.95% of the territory remain underexplored | |
| The Privolzhsky (Volga) Federal District | ||||
| Republic of Bashkortastan | 142,947 | 0.83 | No data available | [25,26] |
| Republic of Mariy El | 23,375 | 0.14 | [10] | |
| Republic of Mordovia | 26,128 | 0.15 | [28,38] | [81] |
| Republic of Tatarstan | 67,847 | 0.40 | No data available | No data available |
| Udmurt Republic | 42,061 | 0.25 | [36] | [82] |
| Chuvash Republic | 18,343 | 0.11 | No data available | No data available |
| Kirov Region | 120,374 | 0.70 | [27,83] | No data available |
| Nizhny Novgorod Region | 76,624 | 0.45 | [83] | [84,85] |
| Orenburg Region | 123,702 | 0.72 | No data available | No data available |
| Penza Region | 43,352 | 0,25 | Data are fragmentary | [86] |
| Perm Region | 160,236 | 0.94 | No data available | No data available |
| Samara Region | 53,565 | 0.31 | Data are fragmentary | [30,87] |
| Saratov Region | 101,240 | 0.59 | [88] | No data available |
| Ulyanovsk Region | 37,181 | 0.22 | [47] | [30,87] |
| Total | 1,036,975 | 6.06 | 3.60% of the territory remain underexplored | |
| The Ural Federal District | ||||
| Kurgan Region | 71,488 | 0.42 | No data available | No data available |
| Sverdlovsk Region | 194,307 | 1.13 | [89] | [90] |
| Tyumen Region (without autonomousareas) | 160,122 | 0.94 | [91] | |
| Khanty-Mansi autonomous area—Yugra | 534,801 | 3.12 | No data available | No data available |
| Yamalo-Nenets autonomous area | 769,250 | 4.49 | No data available | No data available |
| Chelyabinsk Region | 88,529 | 0.52 | No data available | No data available |
| Total | 1,818,497 | 10.62 | 9.49% of the territory remain underexplored | |
| The Siberian Federal District | ||||
| Republic of Altai | 92,903 | 0.54 | [92] | [93] |
| Republic of Tyva | 168,604 | 0.98 | [94] | [93] |
| Republic of Khakassia | 61,569 | 0.36 | [95] | [93] |
| Altai Territory | 167,996 | 0.98 | [91,96,97] | [93] |
| Krasnoyarsk Territory | 2,366,797 | 13.82 | [98] | [93] |
| Irkutsk Region | 774,846 | 4.52 | [99] | [93] |
| Kemerovo Region | 95,725 | 0.56 | [100] | [93,101] |
| Novosibirsk Region | 177,756 | 1.04 | [42,91] | [93] |
| Omsk Region | 141,140 | 0.82 | [91] | [93] |
| Tomsk Region | 314,391 | 1.84 | [102,103] | [93] |
| Total | 4,361,727 | 25.46 | All the regions have been explored sufficiently | |
| The Far Eastern Federal District | ||||
| Republic of Sakha (Yakutia) | 3,083,523 | 18.01 | [104,105,106,107,108,109] | [106] |
| Primorsky Territory | 164,673 | 0.96 | [110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126] | [105,127,128] |
| Khabarovsk Territory | 787,633 | 4.60 | [129,130,131,132,133] | [105,130,131,132] |
| Amur Region | 361,908 | 2.11 | [134,135,136,137] | [105,138] |
| Kamchatka Territory | 464,275 | 2.71 | [139,140,141] | [105,142,143] |
| Magadan Region | 462,464 | 2.70 | [144,145,146,147,148] | [105] |
| Sakhalin Region | 87,101 | 0.51 | [149,150] | [105,151] |
| Jewish Autonomous Region | 36,271 | 0.21 | [152,153] | [105] |
| Chukotka autonomous area | 721,481 | 4.21 | [154] | [105] |
| Republic of Buryatia | 351,334 | 2.05 | [155,156,157] | [93,105] |
| Trans-Baikal Territory | 431,892 | 2.52 | [158] | [93,105] |
| Total | 6,952,555 | 40.59 | All the regions have been explored sufficiently | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinogradova, Y.K.; Tokhtar, V.K.; Notov, A.A.; Mayorov, S.R.; Danilova, E.S. Plant Invasion Research in Russia: Basic Projects and Scientific Fields. Plants 2021, 10, 1477. https://doi.org/10.3390/plants10071477
Vinogradova YK, Tokhtar VK, Notov AA, Mayorov SR, Danilova ES. Plant Invasion Research in Russia: Basic Projects and Scientific Fields. Plants. 2021; 10(7):1477. https://doi.org/10.3390/plants10071477
Chicago/Turabian StyleVinogradova, Yulia K., Valeriy K. Tokhtar, Alexander A. Notov, Sergey R. Mayorov, and Elena S. Danilova. 2021. "Plant Invasion Research in Russia: Basic Projects and Scientific Fields" Plants 10, no. 7: 1477. https://doi.org/10.3390/plants10071477
APA StyleVinogradova, Y. K., Tokhtar, V. K., Notov, A. A., Mayorov, S. R., & Danilova, E. S. (2021). Plant Invasion Research in Russia: Basic Projects and Scientific Fields. Plants, 10(7), 1477. https://doi.org/10.3390/plants10071477