
A qPCR Assay for the Fast Detection and Quantification of Colletotrichum lupini


 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Quantification of C. lupini DNA from Pure Cultures
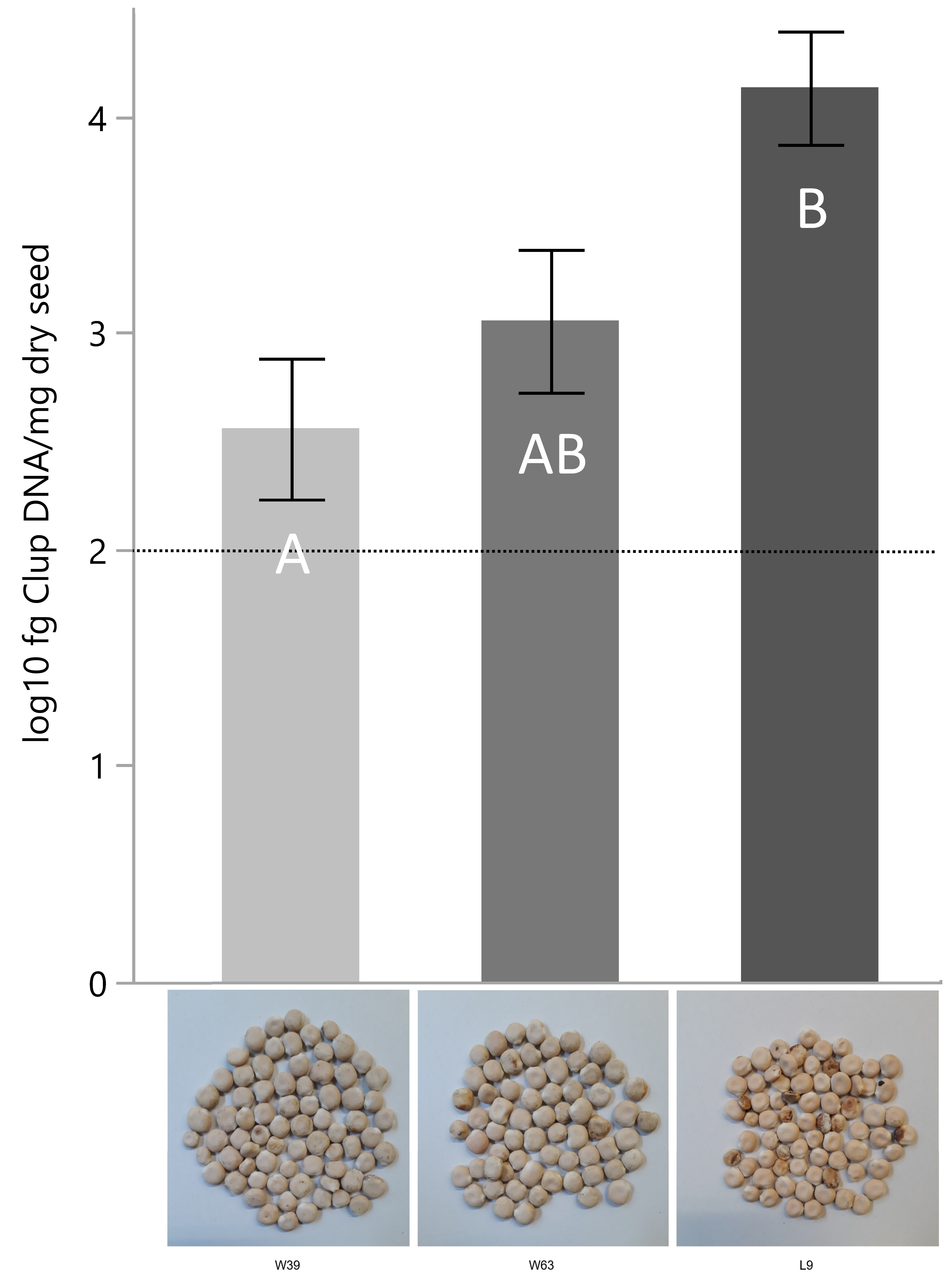
2.2. Quantification of C. lupini DNA from Seeds
2.3. Quantification of C. lupini DNA from Stem, Leaves, and Roots
3. Discussion
4. Materials and Methods
4.1. Fungal Strains
4.2. Seed Material
4.3. DNA Extraction from Seeds and Fungal Cultures
4.4. Primer Design and Amplification Protocol
4.5. Quantification of C. lupini DNA in Lupin Plant Material
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Statistics Database of the Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2018. [Google Scholar]
- Fernández-Pascual, M.; PueyoMaría, J.; Lucas, M.M. Singular features of the Bradyrhizobium-Lupinus symbiosis. Dyn. Soil Dyn. Plant 2007, 1, 1–16. [Google Scholar]
- Schulze, J.; Temple, G.; Temple, S.J.; Beschow, H.; Vance, C.P. Nitrogen fixation by white lupin under phosphorus deficiency. Ann. Bot. 2006, 98, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Goergen, E.; Chambers, J.C.; Blank, R. Effects of water and nitrogen availability on nitrogen contribution by the legume, Lupinus argenteus Pursh. Appl. Soil Ecol. 2009, 42, 200–208. [Google Scholar] [CrossRef]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frias, J.; Martinez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Nirenberg, H.I.; Feiler, U.; Hagedorn, G. Description of Colletotrichum lupini comb. nov in modern terms. Mycologia 2002, 94, 307–320. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talhinhas, P.; Baroncelli, R.; Le Floch, G. Anthracnose of lupins caused by Colletotrichum lupini: A recent disease and a successful worldwide pathogen. J. Plant Pathol. 2016, 98, 5–14. [Google Scholar]
- Alkemade, J.A.; Messmer, M.M.; Voegele, R.T.; Finckh, M.R.; Hohmann, P. Genetic diversity of Colletotrichum lupini and its virulence on white and Andean lupin. Sci. Rep. 2021, 11, 13547. [Google Scholar] [CrossRef] [PubMed]
- White, P.; French, R.; McLarty, A. Producing Lupins; Department of Agriculture and Food: Perth, Australia, 2008. [Google Scholar]
- PGRO. Website of the Processors and Growers Research Organisation. Available online: https://www.pgro.org/ (accessed on 12 December 2020).
- Thomas, G.J.; Sweetingham, M.W. Cultivar and environment influence the development of lupin anthracnose caused by Colletotrichum lupini. Australas. Plant Pathol. 2004, 33, 571–577. [Google Scholar] [CrossRef]
- Thomas, G.J.; Sweetingham, M.W.; Yang, H.A.; Speijers, J. Effect of temperature on growth of Colletotrichum lupini and on anthracnose infection and resistance in lupins. Australas. Plant Pathol. 2008, 37, 35–39. [Google Scholar] [CrossRef]
- Thomas, G.J.; Sweetingham, M.W. Fungicide seed treatments reduce seed transmission and severity of lupin anthracnose caused by Colletotrichum gloeosporioides. Australas. Plant Pathol. 2003, 32, 39–46. [Google Scholar] [CrossRef]
- Falconi, C.E.; Yanez-Mendizabal, V. Dry heat treatment of Andean lupin seed to reduce anthracnose infection. Crop. Prot. 2016, 89, 178–183. [Google Scholar] [CrossRef]
- Tinivella, F.; Hirata, L.; Celan, M.; Wright, S.; Amein, T.; Schmitt, A.; Koch, E.; Wolf, J.; Groot, S.; Stephan, D.; et al. Control of seed-borne pathogens on legumes by microbial and other alternative seed treatments. Eur. J. Plant Pathol. 2009, 123, 139–151. [Google Scholar] [CrossRef]
- Yamagishi, N.; Fujinaga, M.; Ishiyama, Y.; Ogiso, H.; Sato, T.; Tosa, Y. Life cycle and control of Colletotrichum nymphaeae, the causal agent of celery stunt anthracnose. J. Gen. Plant Pathol. 2015, 81, 279–286. [Google Scholar] [CrossRef]
- Feiler, U.; Nirenberg, H.I. Eine neue klassische Methode zur Bestimmung des Colletotrichum-Befalls an Saatgut von Lupinus spp. Nachr. Dtsch. Pflanzenschutzd. 1998, 50, 259–262. [Google Scholar]
- Pecchia, S.; Caggiano, B.; Da Lio, D.; Cafa, G.; Le Floch, G.; Baroncelli, R. Molecular detection of the seed-borne pathogen Colletotrichum lupini targeting the hyper-variable IGS region of the ribosomal cluster. Plants 2019, 8, 222. [Google Scholar] [CrossRef] [Green Version]
- Kulik, T.; Jestoi, M.; Okorski, A. Development of TaqMan assays for the quantitative detection of Fusarium avenaceum/Fusarium tricinctum and Fusarium poae esyn1 genotypes from cereal grain. FEMS Microbiol. Lett. 2011, 314, 49–56. [Google Scholar] [CrossRef]
- Tao, G.; Hyde, K.D.; Cai, L. Species-specific real-time PCR detection of Colletotrichum kahawae. J. Appl. Microbiol. 2013, 114, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, D.; Da Lio, D.; Bartolini, L.; Cappellini, G.; Bruscoli, T.; Bracalini, M.; Benigno, A.; Salemi, C.; Del Nista, D.; Aronadio, A.; et al. A duplex real-time PCR with probe for simultaneous detection of Geosmithia morbida and its vector Pityophthorus juglandis. PLoS ONE 2020, 15, e0241109. [Google Scholar] [CrossRef]
- Dubrulle, G.; Pensec, F.; Picot, A.; Rigalma, K.; Pawtowski, A.; Nicolleau, S.; Harzic, N.; Nodet, P.; Baroncelli, R.; Le Floch, G. Phylogenetic diversity and effect of temperature on pathogenicity of Colletotrichum lupini. Plant Dis. 2020, 104, 938–950. [Google Scholar] [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Alkemade, J.A.; Messmer, M.M.; Arncken, C.; Leska, A.; Annicchiarico, P.; Nazzicari, N.; Ksiazkiewicz, M.; Voegele, R.T.; Finckh, M.R.; Hohmann, P. A high-throughput phenotyping tool to identify field-relevant anthracnose resistance in white lupin. Plant Dis. 2021, 105. [Google Scholar] [CrossRef]
- Minas, K.; McEwan, N.R.; Newbold, C.J.; Scott, K.P. Optimization of a high-throughput CTAB-based protocol for the extraction of qPCR-grade DNA from rumen fluid, plant and bacterial pure cultures. FEMS Microbiol. Lett. 2011, 325, 162–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Forootan, A.; Sjoback, R.; Bjorkman, J.; Sjogreen, B.; Linz, L.; Kubista, M. Methods to determine limit of detection and limit of quantification in quantitative real-time PCR (qPCR). Biomol. Detect. Quantif. 2017, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Clade | Species | Isolate | Cq Value | R2 Value | Efficiency | LoD/LoQ | |||
|---|---|---|---|---|---|---|---|---|---|
| 10 ng/µL | 1 ng/µL | 0.1 ng/µL | 0.01 ng/µL | ||||||
| 1 | C. lupini | JA01 | 24.6 +/−0.41 | 28.5 +/−0.51 | 31.9 +/−0.42 | 35.5 +/−0.72 | 0.97 | 1.06 | 0.01 ng/µL |
| 1 | C. tamarilloi | CBS 129814 | ND | ND | ND | ND | |||
| 2 | C. nymphaeae | CBS 130239 | ND | ND | ND | ND | |||
| 4 | C. acutatum | CBS 369.73 | ND | ND | ND | ND | |||
| - | C. coccodes | CBS 641.97 | ND | ND | ND | ND | |||
| - | C. trifolii | CTR3 | ND | ND | ND | ND | |||
| Plant Organ | Inoculated Plants | Control Plants |
|---|---|---|
 | C. lupini DNA/mg dry plant material | |
| ND | ND | |
| ND | ND | |
| 1.7 × 104 fg | ND | |
| 2.0 × 104 fg | ND | |
| ND | ND | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamber, T.; Malpica-López, N.; Messmer, M.M.; Oberhänsli, T.; Arncken, C.; Alkemade, J.A.; Hohmann, P. A qPCR Assay for the Fast Detection and Quantification of Colletotrichum lupini. Plants 2021, 10, 1548. https://doi.org/10.3390/plants10081548
Kamber T, Malpica-López N, Messmer MM, Oberhänsli T, Arncken C, Alkemade JA, Hohmann P. A qPCR Assay for the Fast Detection and Quantification of Colletotrichum lupini. Plants. 2021; 10(8):1548. https://doi.org/10.3390/plants10081548
Chicago/Turabian StyleKamber, Tim, Nachelli Malpica-López, Monika M. Messmer, Thomas Oberhänsli, Christine Arncken, Joris A. Alkemade, and Pierre Hohmann. 2021. "A qPCR Assay for the Fast Detection and Quantification of Colletotrichum lupini" Plants 10, no. 8: 1548. https://doi.org/10.3390/plants10081548
APA StyleKamber, T., Malpica-López, N., Messmer, M. M., Oberhänsli, T., Arncken, C., Alkemade, J. A., & Hohmann, P. (2021). A qPCR Assay for the Fast Detection and Quantification of Colletotrichum lupini. Plants, 10(8), 1548. https://doi.org/10.3390/plants10081548