
Physiological Characterization of Young ‘Hass’ Avocado Plant Leaves Following Exposure to High Temperatures and Low Light Intensity

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
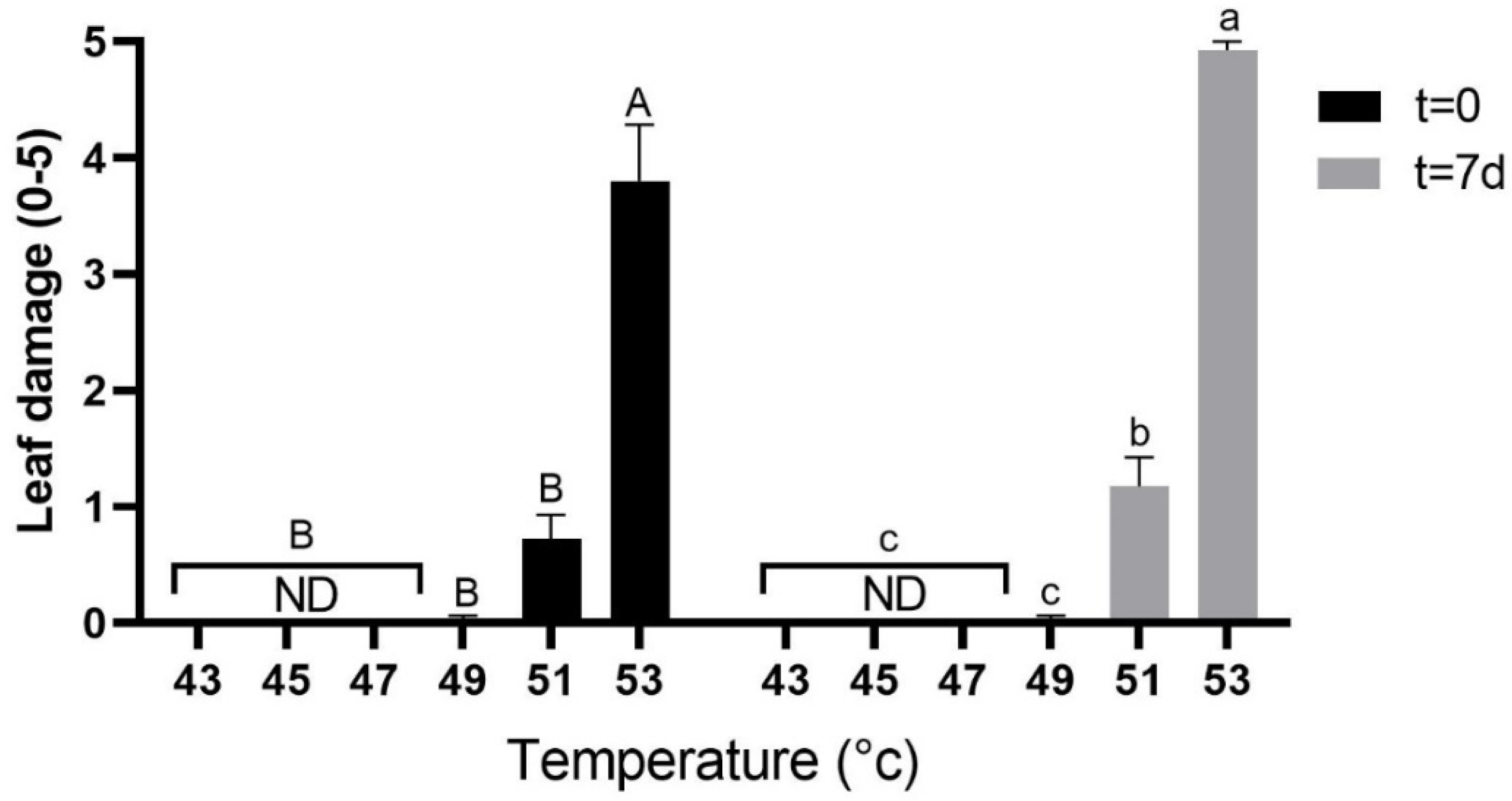
2.1. Effect of Heat Stress on Leaf Damage
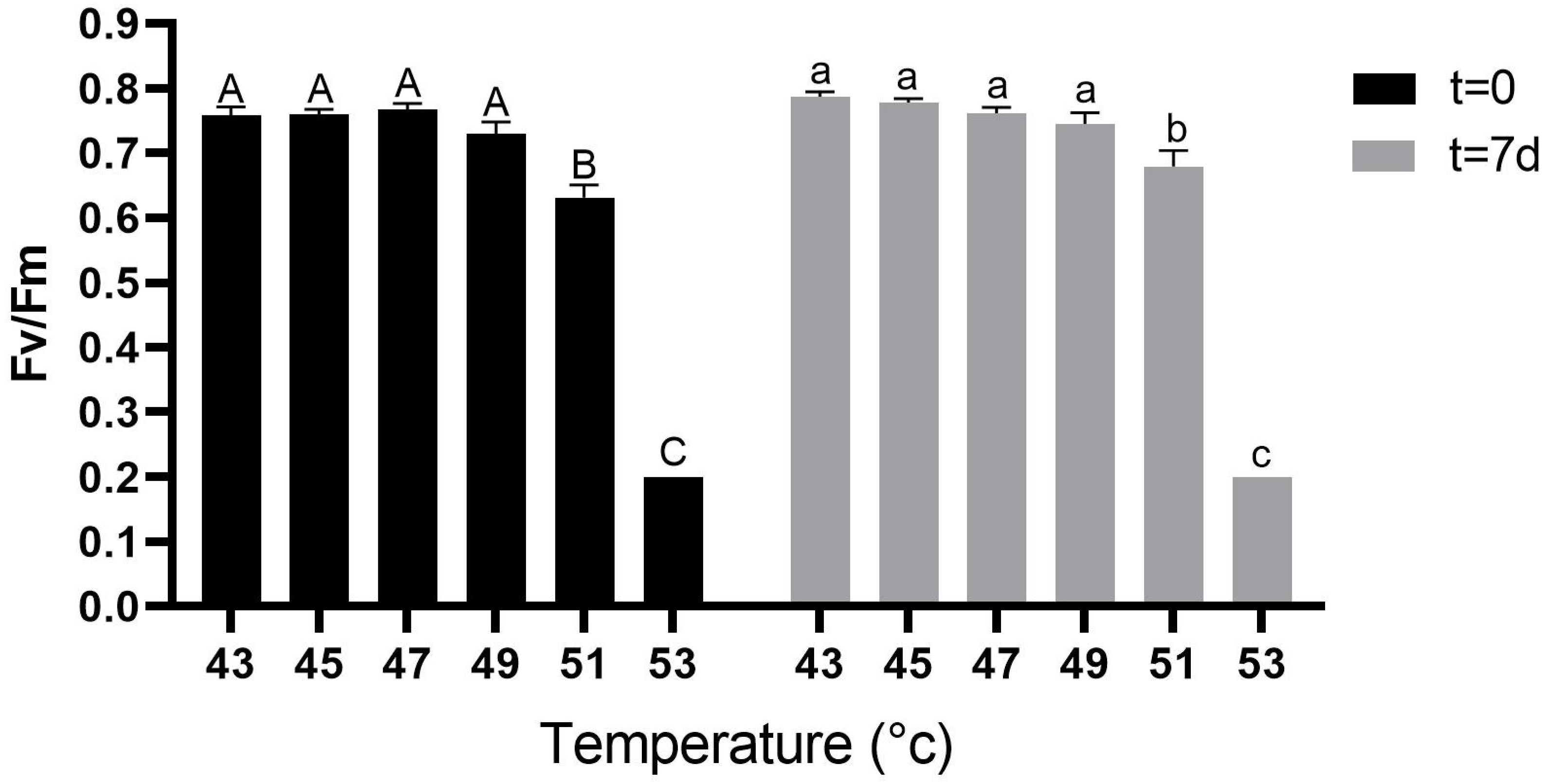
2.2. Effect of Heat Stress on Optimal Quantum Yield of Photosystem II (Fv/Fm)
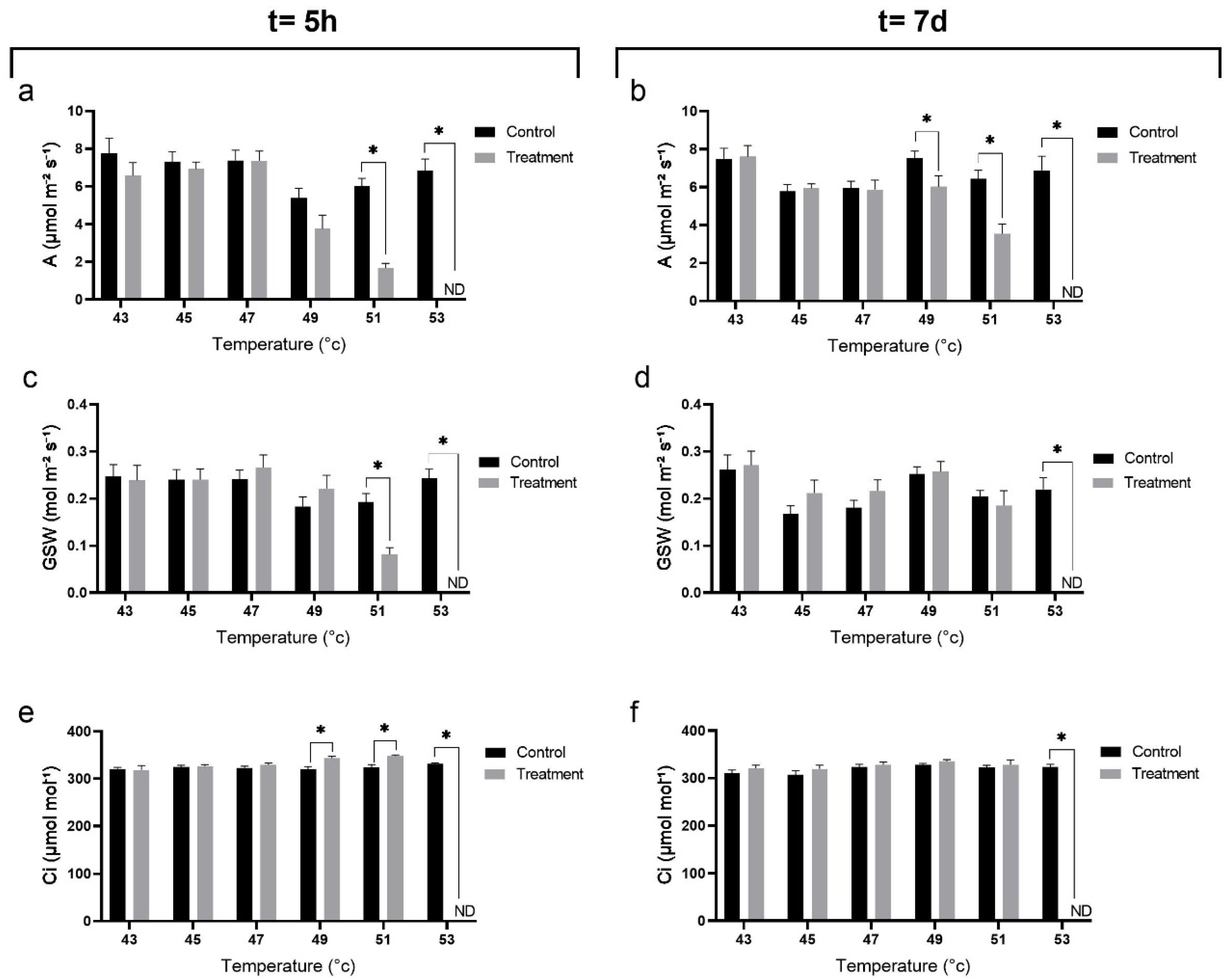
2.3. Effect of Heat Stress on Gas-Exchange Parameters
3. Discussion
4. Materials and Methods
4.1. Plant Material
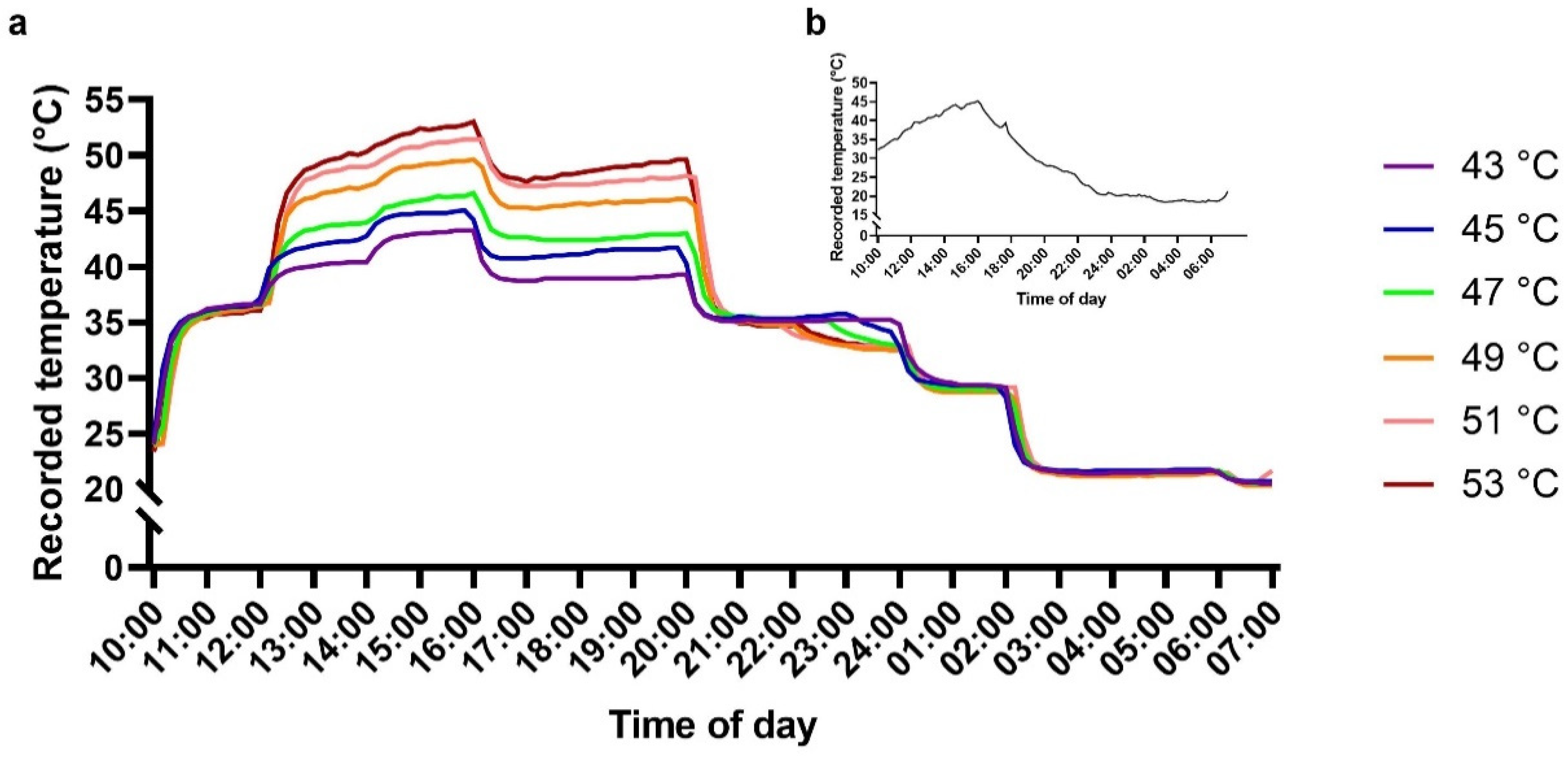
4.2. Heat-Stress Treatments and Temperature Measurements
4.3. Leaf Measurements
4.3.1. Leaf Damage Assessment
4.3.2. Chlorophyll a Fluorescence Analysis
4.3.3. Gas-Exchange Measurements
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zandalinas, S.I.; Fritschi, F.B.; Mittler, R. Global warming, climate change, and environmental pollution: Recipe for a multifactorial stress combination disaster. Trends Plant Sci. 2021, 26, 588–599. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Sharma, S.; Manjeet, M. Heat stress effects in fruit crops. Agric. Rev. 2020, 41, 73–78. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Slattery, R.A.; Ort, D.R. Carbon assimilation in crops at high temperatures. Plant Cell Environ. 2019, 42, 2750–2758. [Google Scholar] [CrossRef] [PubMed]
- Haworth, M.; Marino, G.; Brunetti, C.; Killi, D.; De Carlo, A.; Centritto, M. The impact of heat stress and water deficit on the photosynthetic and stomatal physiology of olive (Olea europaea L.)—A case study of the 2017 heat wave. Plants 2018, 7, 76. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.O.; Rosenqvist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant 2015, 153, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Balfagón, D.; Sengupta, S.; Gómez-Cadenas, A.; Fritschi, F.B.; Azad, R.K.; Mittler, R.; Zandalinasc, S.I. Jasmonic acid is required for plant acclimation to a combination of high light and heat stress. Plant Physiol. 2019, 181, 1668–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filewod, B.; Thomas, S.C. Impacts of a spring heat wave on canopy processes in a northern hardwood forest. Glob. Chang. Biol. 2014, 20, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Alcaraz, M.L.; Thorp, T.G.; Hormaza, J.I. Phenological growth stages of avocado (Persea americana) according to the BBCH scale. Sci. Hortic. Amst. 2013, 164, 434–439. [Google Scholar] [CrossRef]
- Ayala Silva, T.; Ledesma, N. Avocado history, biodiversity and production. In Sustainable Horticultural Systems; Springer International Publishing: Berlin/Heidelberg, Germany, 2014; pp. 157–205. ISBN 978-3-319-06903-6. [Google Scholar]
- Migliore, G.; Farina, V.; Tinervia, S.; Matranga, G.; Schifani, G. Consumer interest towards tropical fruit: Factors affecting avocado fruit consumption in Italy. Agric. Food Econ. 2017, 5, 24. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Lomas, J. An agrometeorological model for assessing the effect of heat stress during the flowering and early fruit set on avocado yields. J. Am. Soc. Hortic. Sci. 1988, 113, 172–176. [Google Scholar]
- Lomas, J. Analysis of the effect of heat stress during flowering on the yield of avocado under mediterranean climatic conditions. Agric. For. Meteorol. 1992, 59, 207–216. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Cohen, I.; Zandalinas, S.I.; Fritschi, F.B.; Sengupta, S.; Fichman, Y.; Azad, R.K.; Mittler, R. The impact of water deficit and heat stress combination on the molecular response, physiology, and seed production of soybean. Physiol. Plant 2021, 172, 41–52. [Google Scholar] [CrossRef]
- Balfagón, D.; Terán, F.; de Oliveira, T.D.R.; Santa, C.; Gómez-Cadenas, A. Citrus rootstocks modify scion antioxidant system under drought and heat stress combination. Plant Cell Rep. 2021. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Joshi, N.C.; Yadav, D.; Ratner, K.; Kamara, I.; Aviv-Sharon, E.; Irihimovitch, V.; Charuvi, D. Sodium hydrosulfide priming improves the response of photosynthesis to overnight frost and day high light in avocado (Persea americana Mill, cv. ‘Hass’). Physiol. Plant. 2019, 168. [Google Scholar] [CrossRef] [Green Version]
- Whiley, A.W.; Searle, C.; Schaffer, B.; Wolstenholme, B.N. Cool orchard temperatures or growing trees in containers can inhibit leaf gas exchange of avocado and mango. J. Am. Soc. Hortic. Sci. 1999, 124, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, K.; Tsonev, T.; Velikova, V.; Yordanov, I. Photosynthetic activity during high temperature treatment of pea plants. J. Plant Physiol. 2000, 157, 169–176. [Google Scholar] [CrossRef]
- Law, R.D.; Crafts-Brandner, S.J. Inhibition and acclimation of photosynthesis to heat stress is closely correlated with activation of ribulose-1,5-bisphosphate carboxylase/oxygenase. Plant Physiol. 1999, 120, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Ameye, M.; Wertin, T.M.; Bauweraerts, I.; McGuire, M.A.; Teskey, R.O.; Steppe, K. The effect of induced heat waves on Pinus taeda and quercus rubra seedlings in ambient and elevated CO2 atmospheres. New Phytol. 2012, 196, 448–461. [Google Scholar] [CrossRef]
- Abd El-Naby, S.K.M.; Esmail, A.M.A.M.; Baiea, M.H.M.; Amin, O.A.E.F.; Mohamed, A.A.A. Mitigation of heat stress effects by using shade net on Washington navel orange trees grown in Al-Nubaria region, Egypt. Acta Sci. Pol. Hortorum Cultus 2020, 19, 15–24. [Google Scholar] [CrossRef]
- Pelletier, V.; Pepin, S.; Gallichand, J.; Caron, J. Reducing cranberry heat stress and midday depression with evaporative cooling. Sci. Hortic. (Amst.) 2016, 198, 445–453. [Google Scholar] [CrossRef]
- Weil, A.; Sofer-Arad, C.; Bar-Noy, Y.; Liran, O.; Rubinovich, L. Comparative study of leaf antioxidant activity as a possible mechanism for frost tolerance in ‘Hass’ and ‘Ettinger’ avocado cultivars. J. Agric. Sci. 2019, 157, 342–349. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shapira, O.; Chernoivanov, S.; Neuberger, I.; Levy, S.; Rubinovich, L. Physiological Characterization of Young ‘Hass’ Avocado Plant Leaves Following Exposure to High Temperatures and Low Light Intensity. Plants 2021, 10, 1562. https://doi.org/10.3390/plants10081562
Shapira O, Chernoivanov S, Neuberger I, Levy S, Rubinovich L. Physiological Characterization of Young ‘Hass’ Avocado Plant Leaves Following Exposure to High Temperatures and Low Light Intensity. Plants. 2021; 10(8):1562. https://doi.org/10.3390/plants10081562
Chicago/Turabian StyleShapira, Or, Simon Chernoivanov, Itamar Neuberger, Shay Levy, and Lior Rubinovich. 2021. "Physiological Characterization of Young ‘Hass’ Avocado Plant Leaves Following Exposure to High Temperatures and Low Light Intensity" Plants 10, no. 8: 1562. https://doi.org/10.3390/plants10081562