
Hypouricemic Effects of Chrysanthemum indicum L. and Cornus officinalis on Hyperuricemia-Induced HepG2 Cells, Renal Cells, and Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Extract Preparation and HPLC
2.2. Cell Culture and Treatments
2.3. Animals
2.4. Levels of Uric Acid and Creatinine in the Urine and Serum
2.5. Activity of Xanthine Oxidase
2.6. mRNA Expression of Xanthine Dehydrogenase
2.7. Serum Triglycerides, Cholesterols, Aspartate Transaminase (AST), and Alanine Transaminase (ALT)
2.8. Antioxidant Enzyme Activity in the Liver
2.9. Expression Levels of OAT1, OAT3, GLUT9, and URAT1 Proteins
2.10. Statistical Analysis
3. Results
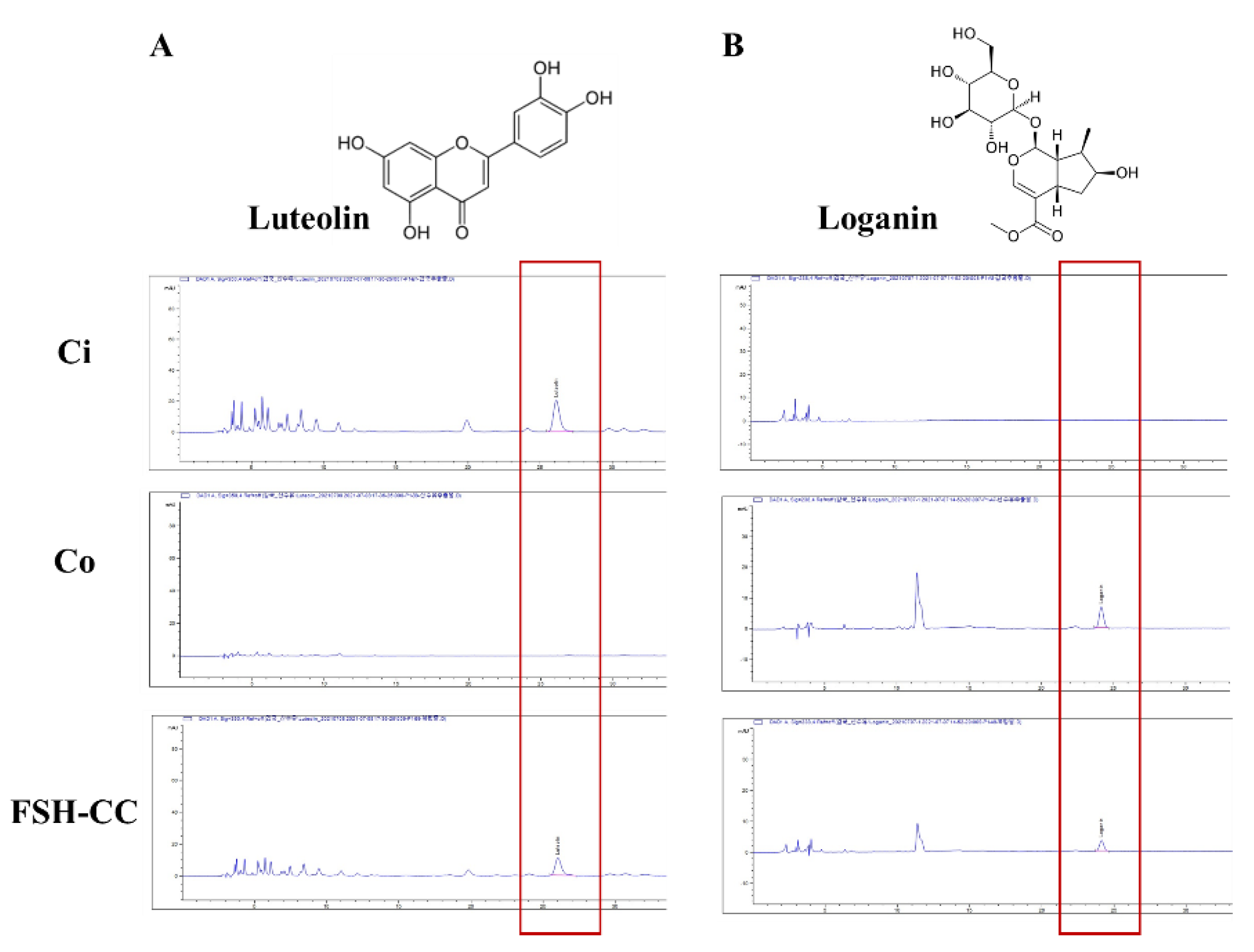
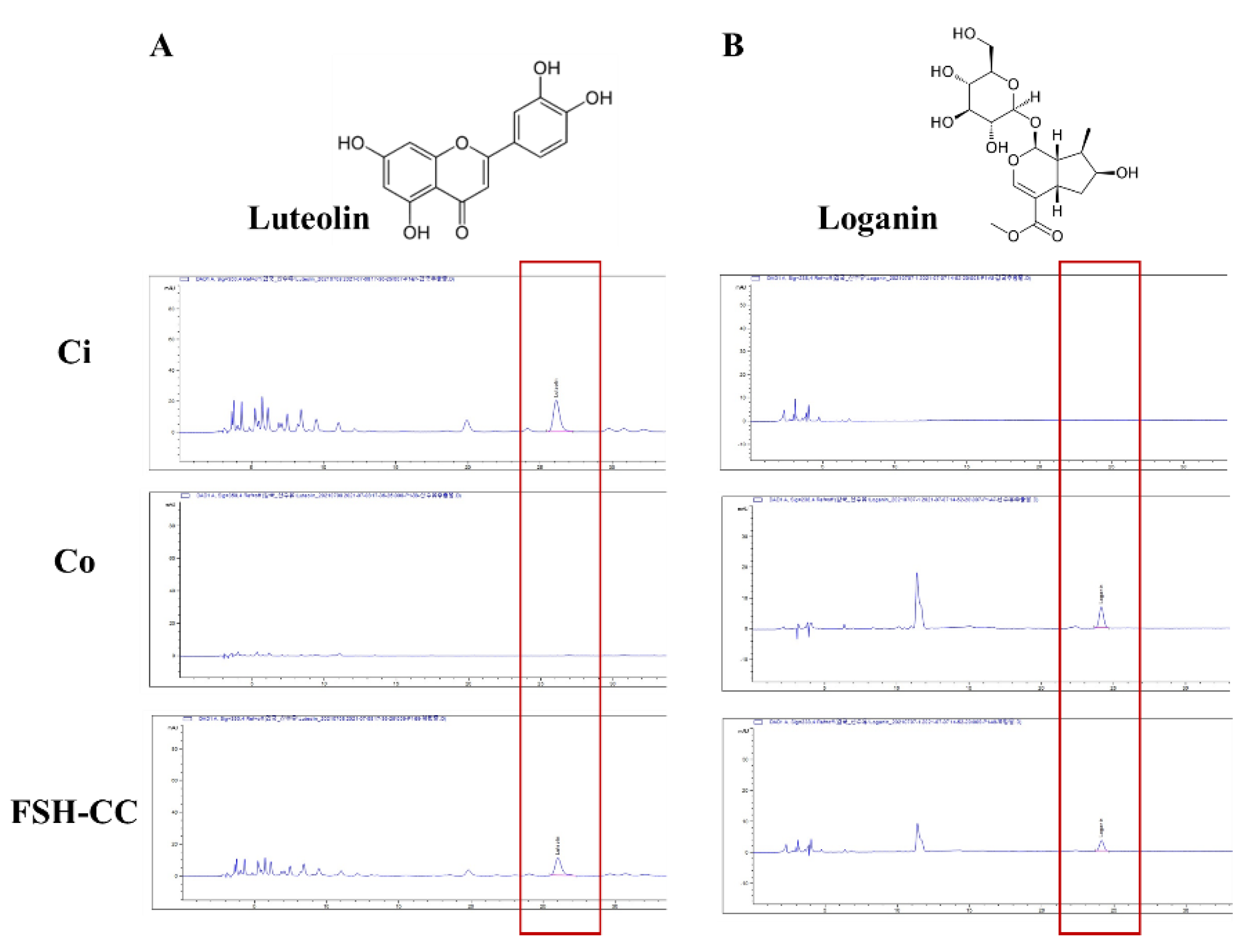
3.1. Luteolin and Loganin of Ci, Co, and FSH-CC
3.2. The Combination of Ci and Co Suppressed the Xanthine Oxidase Activity and Xanthine Dehydrogenase mRNA Expression in Liver Cells More Than Their Individual Treatments
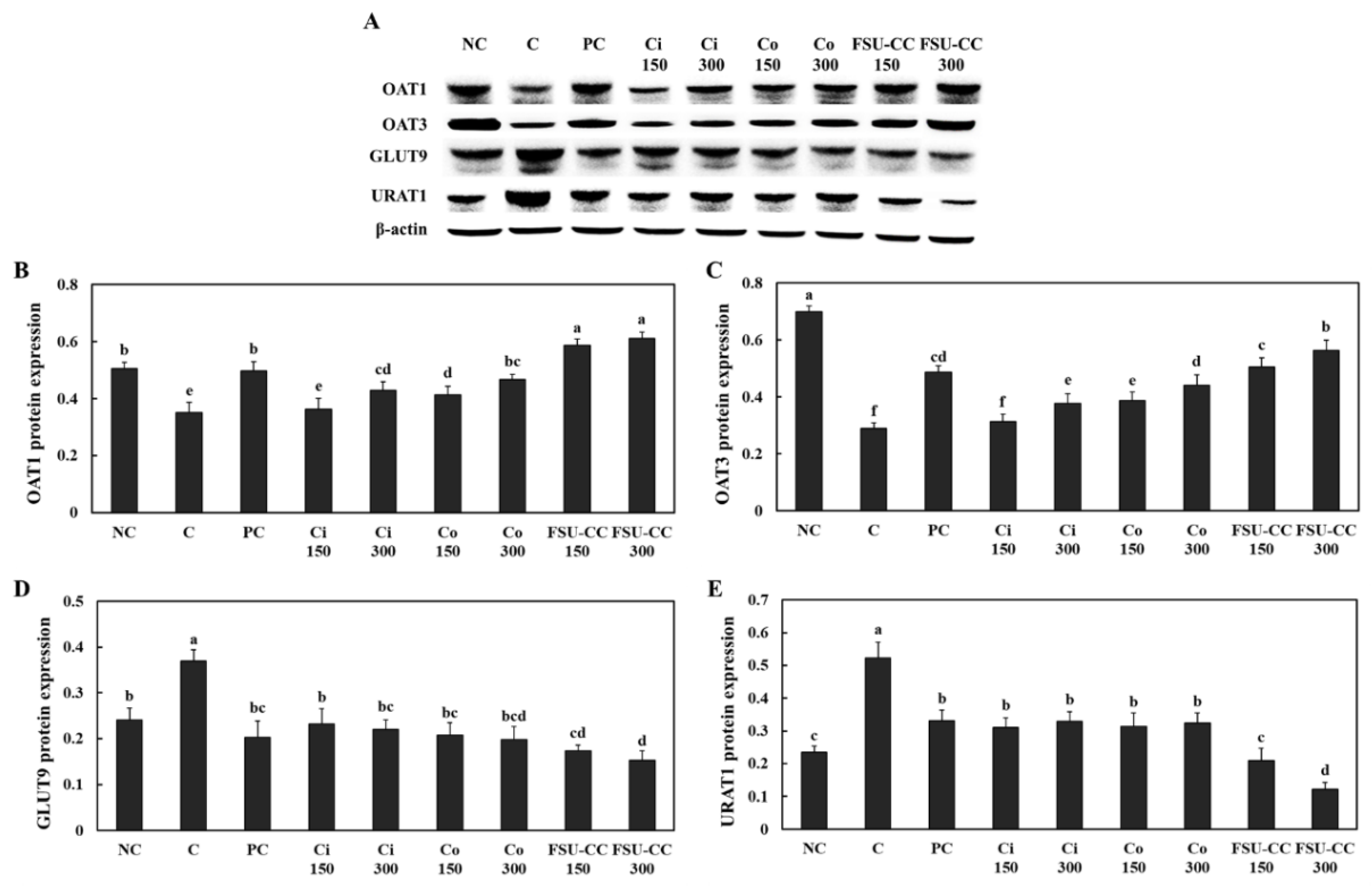
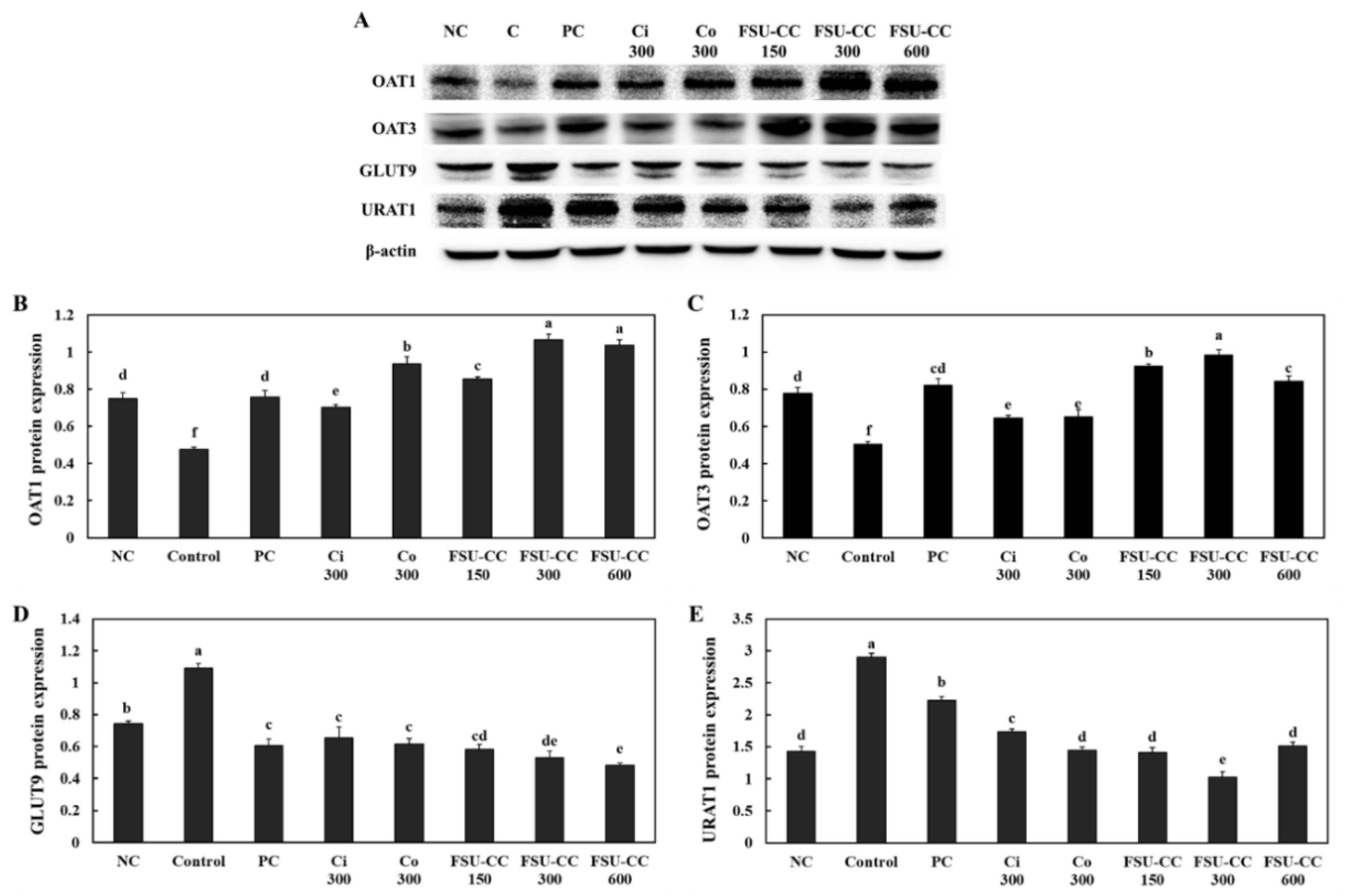
3.3. The Combination of Ci and Co Increased the Expression Levels of OAT1 and OAT3 and Suppressed the Expression Levels of GLUT9 and URAT1 in Renal Proximal Tubule Cells More Than Their Individual Treatments
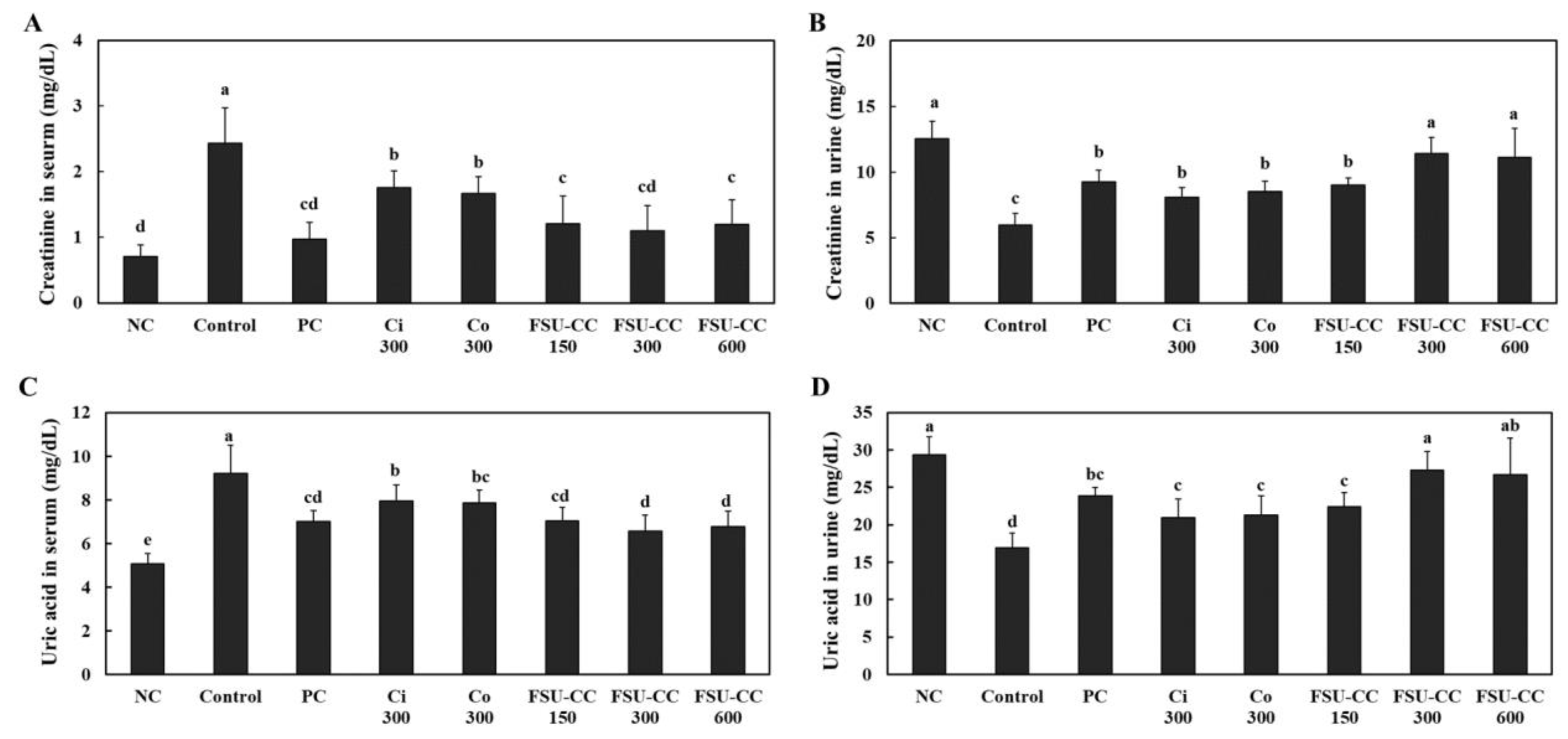
3.4. The Combination of Ci and Co Increased the Excretion of Creatinine and Uric Acid in Hyperuricemia-Induced Mice More Than Their Individual Treatments
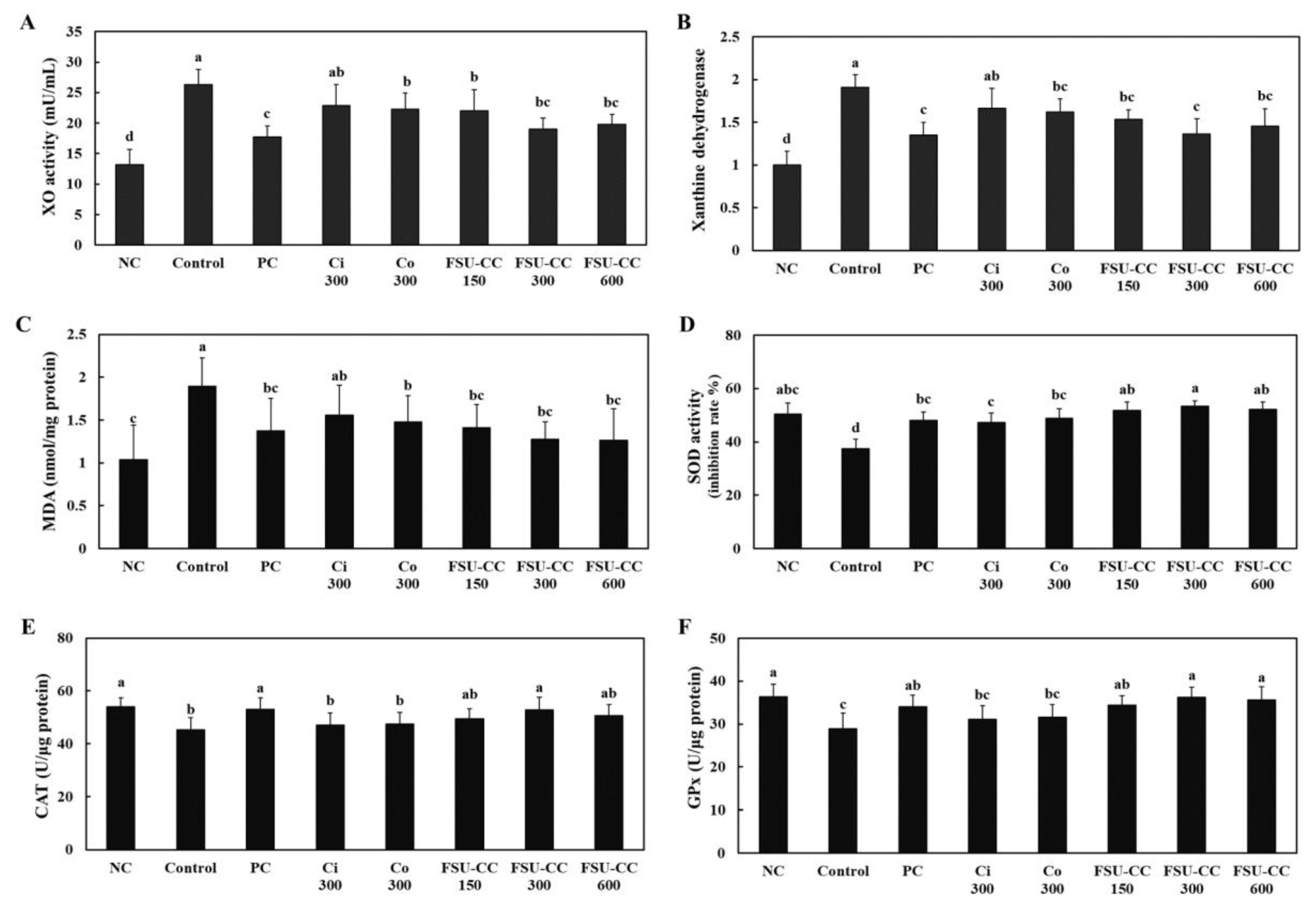
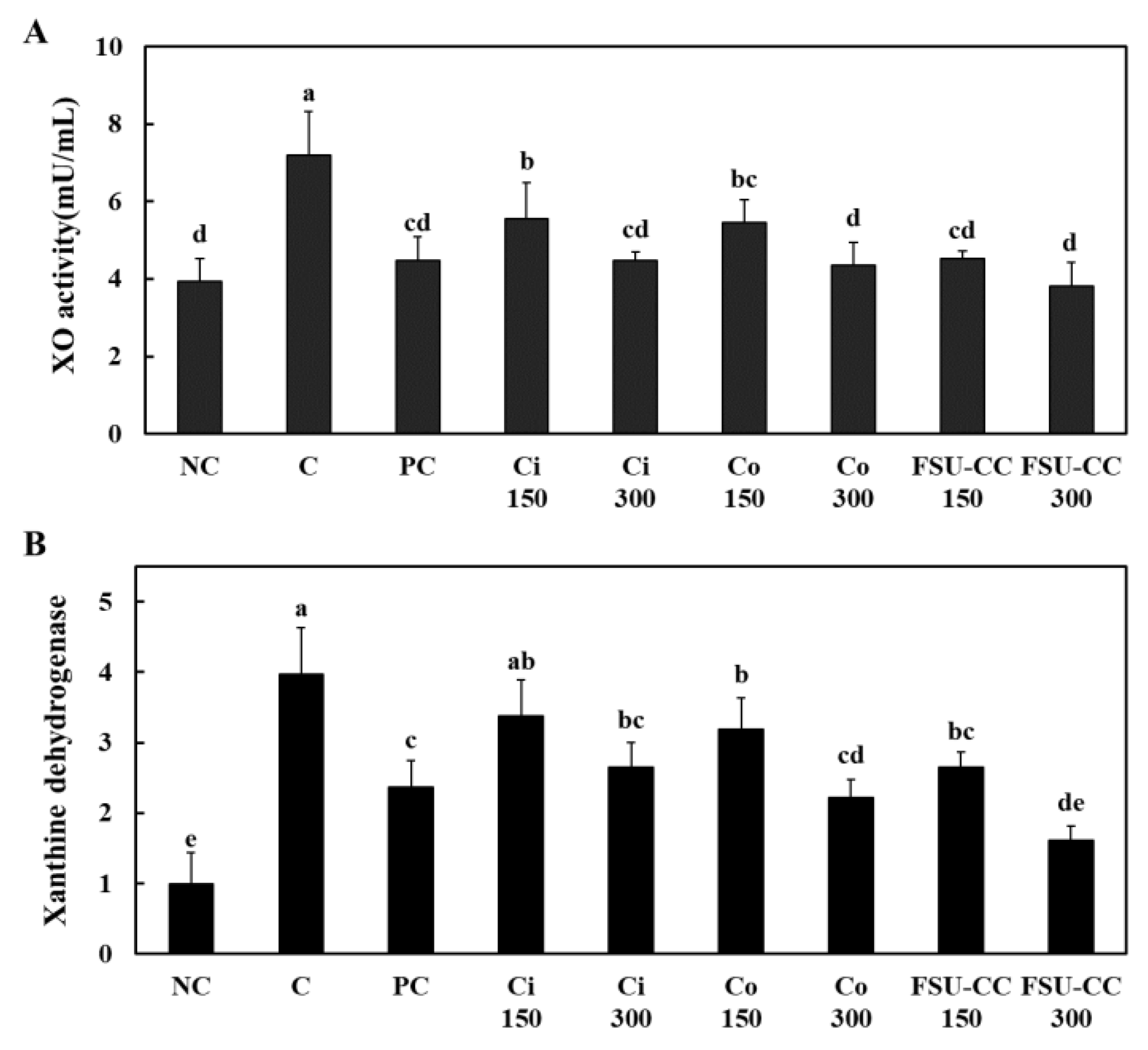
3.5. The Combination of Ci and Co Increased the Xanthine Oxidase Activity and Xanthine Dehydrogenase mRNA Expression and Inhibited the Oxidative Stress in Hyperuricemia-Induced Mice More Than Their Individual Treatments
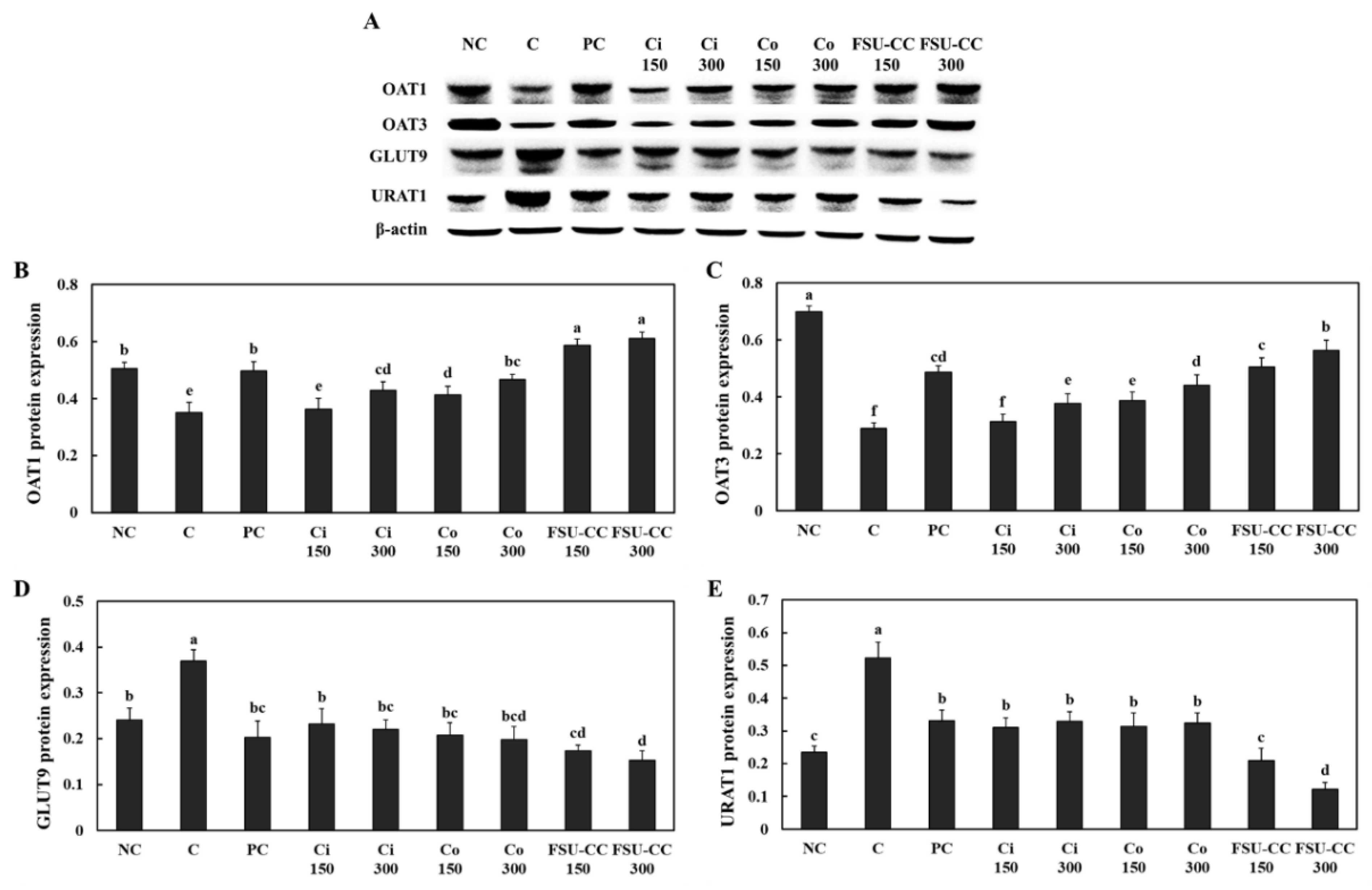
3.6. The Combination of Ci and Co Increased the Expression Levels of OAT1 and OAT3 and Suppressed the Expression Levels of GLUT9 and URAT1 in Hyperuricemia-Induced Mice More Than Their Individual Treatments
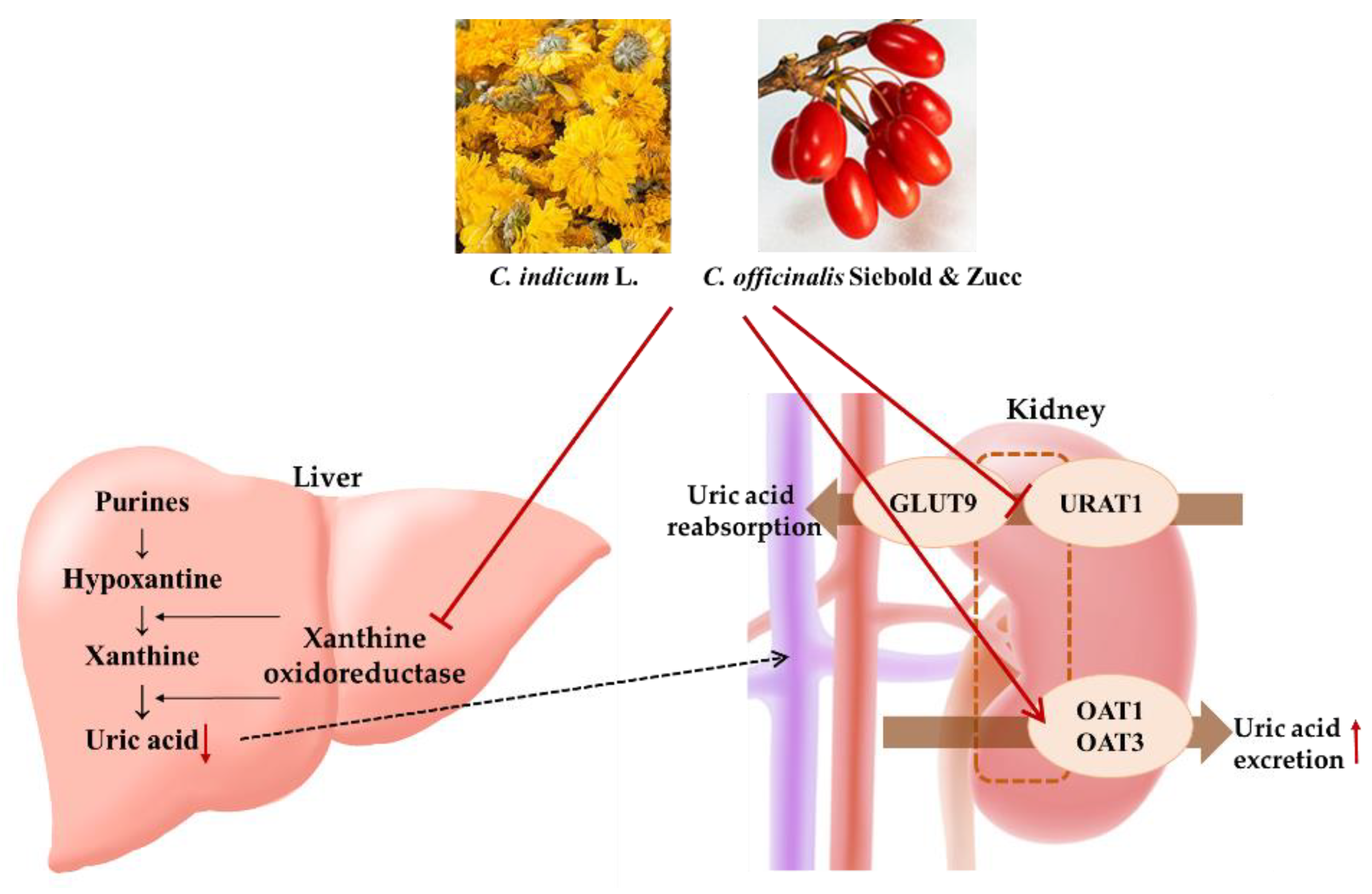
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pedley, A.M.; Benkovic, S.J. A New View into the Regulation of Purine Metabolism: The Purinosome. Trends Biochem. Sci. 2017, 42, 141–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghi, C.; Agabiti-Rosei, E.; Johnson, R.J.; Kielstein, J.T.; Lurbe, E.; Mancia, G.; Redon, J.; Stack, A.G.; Tsioufis, K.P. Hyperuricaemia and gout in cardiovascular, metabolic and kidney disease. Eur. J. Intern. Med. 2020, 80, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Pétrilli, V.; Mayor, A.; Tardivel, A.; Tschopp, J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Dehlin, M.; Jacobsson, L.; Roddy, E. Global epidemiology of gout: Prevalence, incidence, treatment patterns and risk factors. Nat. Rev. Rheumatol. 2020, 16, 380–390. [Google Scholar] [CrossRef]
- Battelli, M.G.; Bortolotti, M.; Polito, L.; Bolognesi, A. The role of xanthine oxidoreductase and uric acid in metabolic syndrome. Biochim. Biophys. Acta. Mol. Basis Dis. 2018, 1864, 2557–2565. [Google Scholar] [CrossRef]
- Furuhashi, M.; Matsumoto, M.; Tanaka, M.; Moniwa, N.; Murase, T.; Nakamura, T.; Ohnishi, H.; Saitoh, S.; Shimamoto, K.; Miura, T. Plasma Xanthine Oxidoreductase Activity as a Novel Biomarker of Metabolic Disorders in a General Population. Circ. J. 2018, 82, 1892–1899. [Google Scholar] [CrossRef] [Green Version]
- Nishino, T.; Okamoto, K.; Eger, B.T.; Pai, E.F.; Nishino, T. Mammalian xanthine oxidoreductase-mechanism of transition from xanthine dehydrogenase to xanthine oxidase. FEBS J. 2008, 275, 3278–3289. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Xu, H.; Sun, Q.; Yu, X.; Chen, W.; Wei, H.; Jiang, J.; Xu, Y.; Lu, W. The Role of Oxidative Stress in Hyperuricemia and Xanthine Oxidoreductase (XOR) Inhibitors. Oxid. Med. Cell Longev. 2021, 26, 1470380. [Google Scholar]
- Kang, D.H.; Nakagawa, T. Uric acid and chronic renal disease: Possible implication of hyperuricemia on progression of renal disease. Semin. Nephrol. 2005, 25, 43–49. [Google Scholar] [CrossRef]
- Xu, L.; Shi, Y.; Zhuang, S.; Liu, N. Recent advances on uric acid transporters. Oncotarget 2017, 8, 100852–100862. [Google Scholar] [CrossRef] [Green Version]
- Lipkowitz, M.S. Regulation of uric acid excretion by the kidney. Curr. Rheumatol. Rep. 2012, 14, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Sun, Y.; Li, D.; Chen, Y. Chrysanthemum indicum L.: A Comprehensive Review of its Botany, Phytochemistry and Pharmacology. Am. J. Chin. Med. 2020, 48, 871–897. [Google Scholar] [CrossRef]
- Yu, H.; Yao, S.; Zhou, C.; Fu, F.; Luo, H.; Du, W.; Jin, H.; Tong, P.; Chen, D.; Wu, C.; et al. Morroniside attenuates apoptosis and pyroptosis of chondrocytes and ameliorates osteoarthritic development by inhibiting NF-κB signaling. J. Ethnopharmacol. 2021, 266, 113447. [Google Scholar] [CrossRef]
- Gao, X.; Liu, Y.; An, Z.; Ni, J. Active Components and Pharmacological Effects of Cornus officinalis: Literature Review. Front. Pharmacol. 2021, 12, 633447. [Google Scholar] [CrossRef]
- Lu, X.; Zeng, R.; Lin, J.; Hu, J.; Rong, Z.; Xu, W.; Liu, Z.; Zeng, W. Pharmacological basis for use of madecassoside in gouty arthritis: Anti-inflammatory, anti-hyperuricemic, and NLRP3 inhibition. Immunopharmacol. Immunotoxicol. 2019, 41, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Pontremoli, R.; Bocale, R.; Ferri, C.; Desideri, G. Therapeutic approaches to chronic hyperuricemia and gout. High Blood Press. Cardiovasc. Prev. 2014, 21, 243–250. [Google Scholar] [CrossRef]
- Zhang, K.H.; Wang, M.Q.; Wei, L.L.; Feng, C.J.; Zhang, Y.S.; Teng, J.B. Investigation of the Effects and Mechanisms of Dendrobium loddigesii Rolfe Extract on the Treatment of Gout. Evid. Based Complement. Alternat. Med. 2020, 30, 4367347. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.G.; Song, H.; Kim, J.H.; Oh, J.M.; Park, J.Y.; Ko, H.C.; Hur, S.P.; Kim, S.J. Prevention of Hyperuricemia by Clerodendrum trichotomum Leaf Extract in Potassium Oxonate-Induced Mice. Dev. Reprod. 2020, 24, 89–100. [Google Scholar] [CrossRef]
- Jeong, S.C.; Kim, S.M.; Jeong, Y.T.; Song, C.H. Hepatoprotective effect of water extract from Chrysanthemum indicum L. flower. Chin. Med. 2013, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Feng, Z.L.; Chen, H.B.; Wang, F.S.; Lu, J.H. Corni Fructus: A review of chemical constituents and pharmacological activities. Chin. Med. 2018, 13, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, Y. Relation between creatinine and uric acid excretion. Ann. Rheum. Dis. 1992, 51, 101–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novikov, A.; Fu, Y.; Huang, W.; Freeman, B.; Patel, R.; van Ginkel, C.; Koepsell, H.; Busslinger, M.; Onishi, A.; Nespoux, J.; et al. SGLT2 inhibition and renal urate excretion: Role of luminal glucose, GLUT9, and URAT1. Am. J. Physiol. Ren. Physiol. 2019, 316, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Bush, K.T.; Nigam, S.K. Key Role for the Organic Anion Transporters, OAT1 and OAT3, in the in vivo Handling of Uremic Toxins and Solutes. Sci. Rep. 2017, 7, 4939. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Morikawa, T.; Toguchida, I.; Harima, S.; Yoshikawa, M. Medicinal flowers. VI. Absolute stereostructures of two new flavanone glycosides and a phenylbutanoid glycoside from the flowers of Chrysanthemum indicum L.: Their inhibitory activities for rat lens aldose reductase. Chem. Pharm. Bull. 2002, 50, 972–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, A.; Zhao, L.; Wang, C.; Nadeem, M.; Raza, A.; Ali, N.; Shah, A.A. Management of hyperuricemia through dietary polyphenols as a natural medicament: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1433–1455. [Google Scholar] [CrossRef]
- Mehmood, A.; Ishaq, M.; Zhao, L.; Safdar, B.; Rehman, A.U.; Munir, M.; Raza, A.; Nadeem, M.; Iqbal, W.; Wang, C. Natural compounds with xanthine oxidase inhibitory activity: A review. Chem. Biol. Drug Des. 2019, 93, 387–418. [Google Scholar] [CrossRef]
- Ishaq, M.; Mehmood, A.; Ur Rehman, A.; Dounya Zad, O.; Li, J.; Zhao, L.; Wang, C.; Hossen, I.; Naveed, M.; Lian, Y. Antihyperuricemic effect of dietary polyphenol sinapic acid commonly present in various edible food plants. J. Food Biochem. 2020, 44, 13111. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AST (mU/mL) | ALT (mU/mL) | Triglycerides (μg/mL) | Total cholesterol (μg/mL) | HDL (μg/mL) | LDL (μg/mL) | |
|---|---|---|---|---|---|---|
| NC | 22.79 ± 3.34ns | 3.37 ± 0.41ns | 2.60 ± 0.39ns | 7.58 ± 0.72ns | 1.72 ± 0.24ns | 1.63 ± 0.89ns |
| C | 23.02 ± 2.26 | 2.91 ± 0.64 | 2.26 ± 0.14 | 8.06 ± 0.81 | 1.89 ± 0.29 | 1.74 ± 0.84 |
| PC | 21.83 ± 2.49 | 2.91 ± 0.35 | 2.62 ± 0.26 | 7.75 ± 0.83 | 1.70 ± 0.34 | 1.30 ± 0.91 |
| Ci 300 | 23.66 ± 2.13 | 3.06 ± 0.77 | 2.63 ± 0.37 | 7.56 ± 0.97 | 1.74 ± 0.32 | 1.61 ± 0.75 |
| Co 300 | 23.39 ± 2.18 | 3.13 ± 0.48 | 2.44 ± 0.33 | 8.33 ± 0.56 | 1.75 ± 0.19 | 1.55 ± 092 |
| FSU-CC 150 | 22.77 ± 1.29 | 2.79 ± 0.32 | 2.47 ± 0.42 | 7.60 ± 0.76 | 1.80 ± 0.33 | 1.30 ± 1.55 |
| FSU-CC 300 | 24.08 ± 6.82 | 3.25 ± 0.58 | 2.43 ± 0.14 | 7.72 ± 1.46 | 1.57 ± 0.22 | 1.38 ± 1.32 |
| FSU-CC 600 | 22.85 ± 1.36 | 3.24 ± 0.78 | 2.61 ± 0.19 | 7.11 ± 0.56 | 1.64 ± 0.27 | 1.24 ± 0.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, O.-K.; Yun, J.-M.; Lee, M.; Kim, D.; Lee, J. Hypouricemic Effects of Chrysanthemum indicum L. and Cornus officinalis on Hyperuricemia-Induced HepG2 Cells, Renal Cells, and Mice. Plants 2021, 10, 1668. https://doi.org/10.3390/plants10081668
Kim O-K, Yun J-M, Lee M, Kim D, Lee J. Hypouricemic Effects of Chrysanthemum indicum L. and Cornus officinalis on Hyperuricemia-Induced HepG2 Cells, Renal Cells, and Mice. Plants. 2021; 10(8):1668. https://doi.org/10.3390/plants10081668
Chicago/Turabian StyleKim, Ok-Kyung, Jeong-Moon Yun, Minhee Lee, Dakyung Kim, and Jeongmin Lee. 2021. "Hypouricemic Effects of Chrysanthemum indicum L. and Cornus officinalis on Hyperuricemia-Induced HepG2 Cells, Renal Cells, and Mice" Plants 10, no. 8: 1668. https://doi.org/10.3390/plants10081668
APA StyleKim, O.-K., Yun, J.-M., Lee, M., Kim, D., & Lee, J. (2021). Hypouricemic Effects of Chrysanthemum indicum L. and Cornus officinalis on Hyperuricemia-Induced HepG2 Cells, Renal Cells, and Mice. Plants, 10(8), 1668. https://doi.org/10.3390/plants10081668