
The Effect of Abiotic Stresses on the Protein Composition of Four Hungarian Wheat Varieties


Abstract
:1. Introduction
2. Results and Discussion
- Analytical data are compressed in [% of the whole meal sample] dimension, by combining raw protein content size exclusion- and reverse-phase high performance liquid chromatography (SE- and RP-HPLC) data as described in the Method section.
- Alterations caused by the abiotic treatments are illustrated by comparing the analytical data to the percentage of the corresponding control data.
2.1. Effect of Abiotic Stress Treatments on the Physiological Parameters and Yield
2.1.1. Drought Stress
2.1.2. Heat Stress
2.1.3. Combined Stress
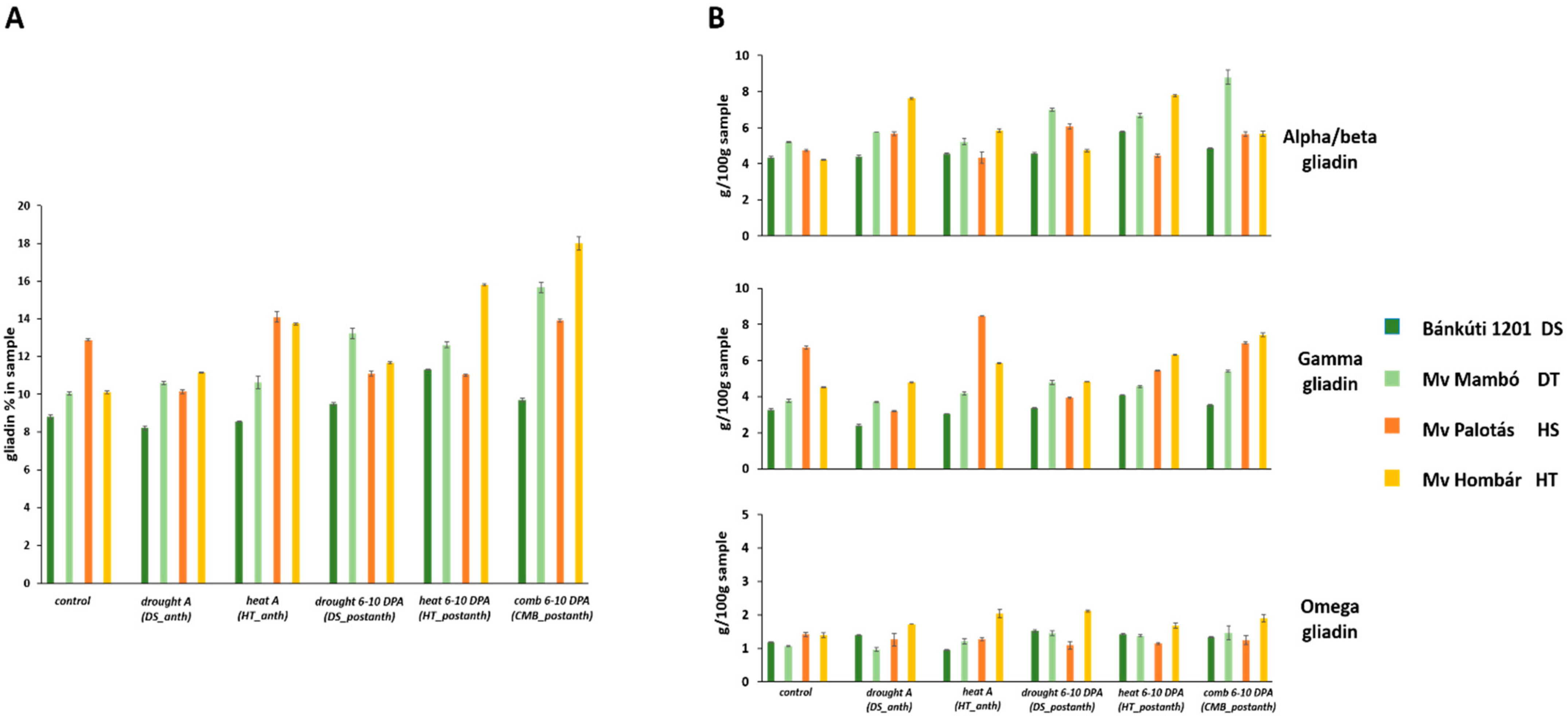
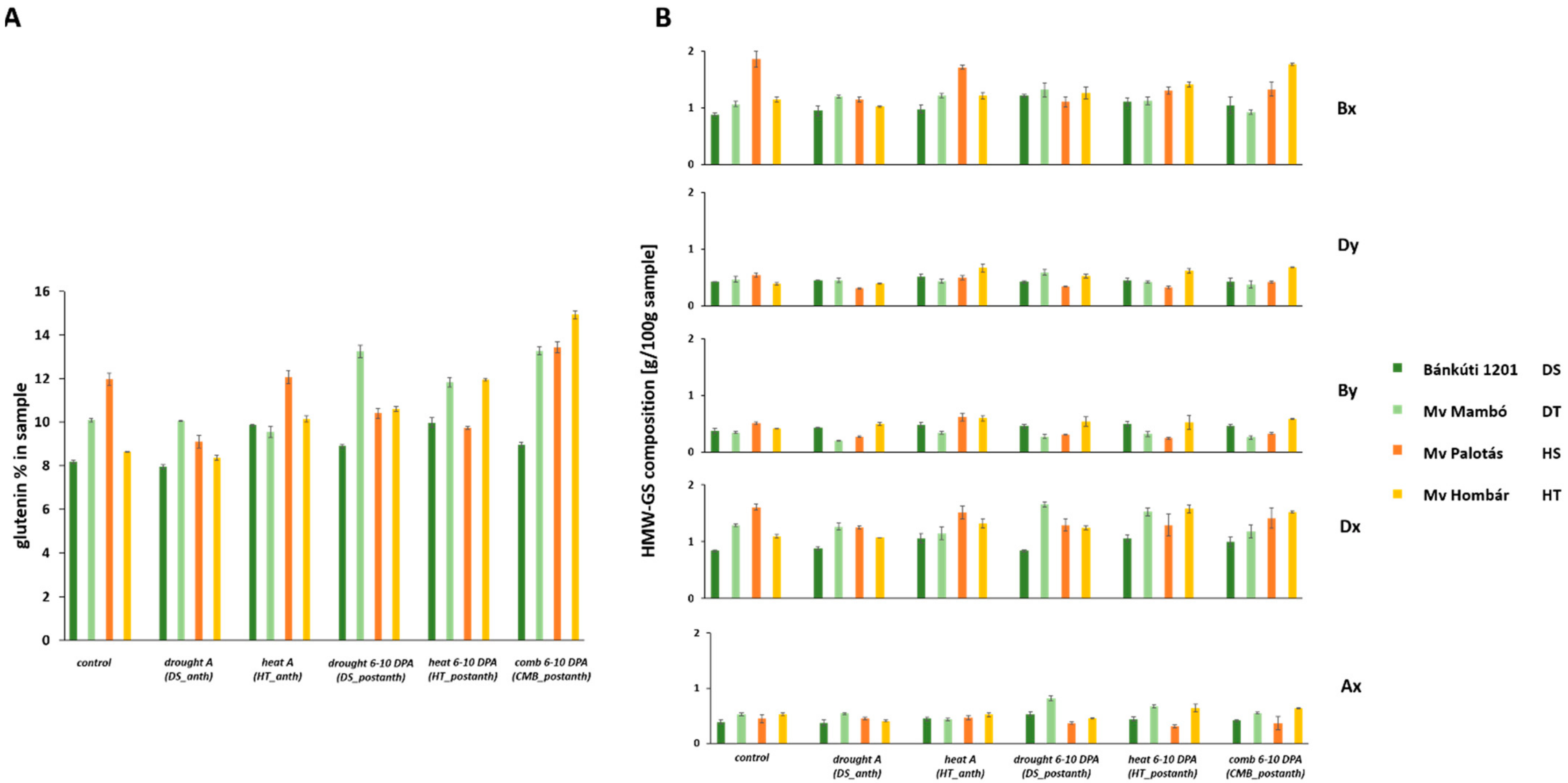
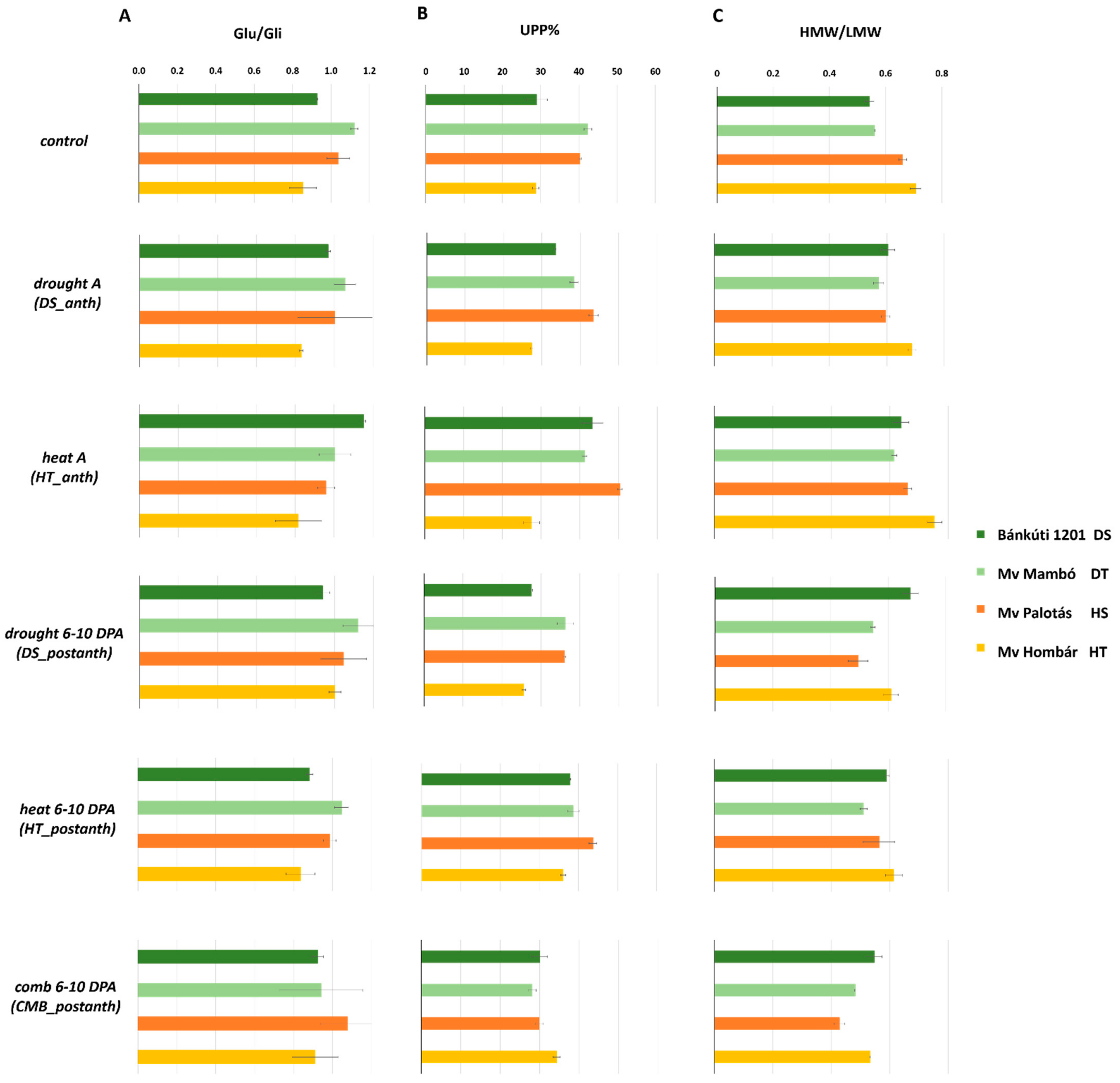
2.2. Effect of Abiotic Stress on Protein Content and Protein Composition
2.2.1. Drought Stress
2.2.2. Heat Stress
2.2.3. Combined Stress
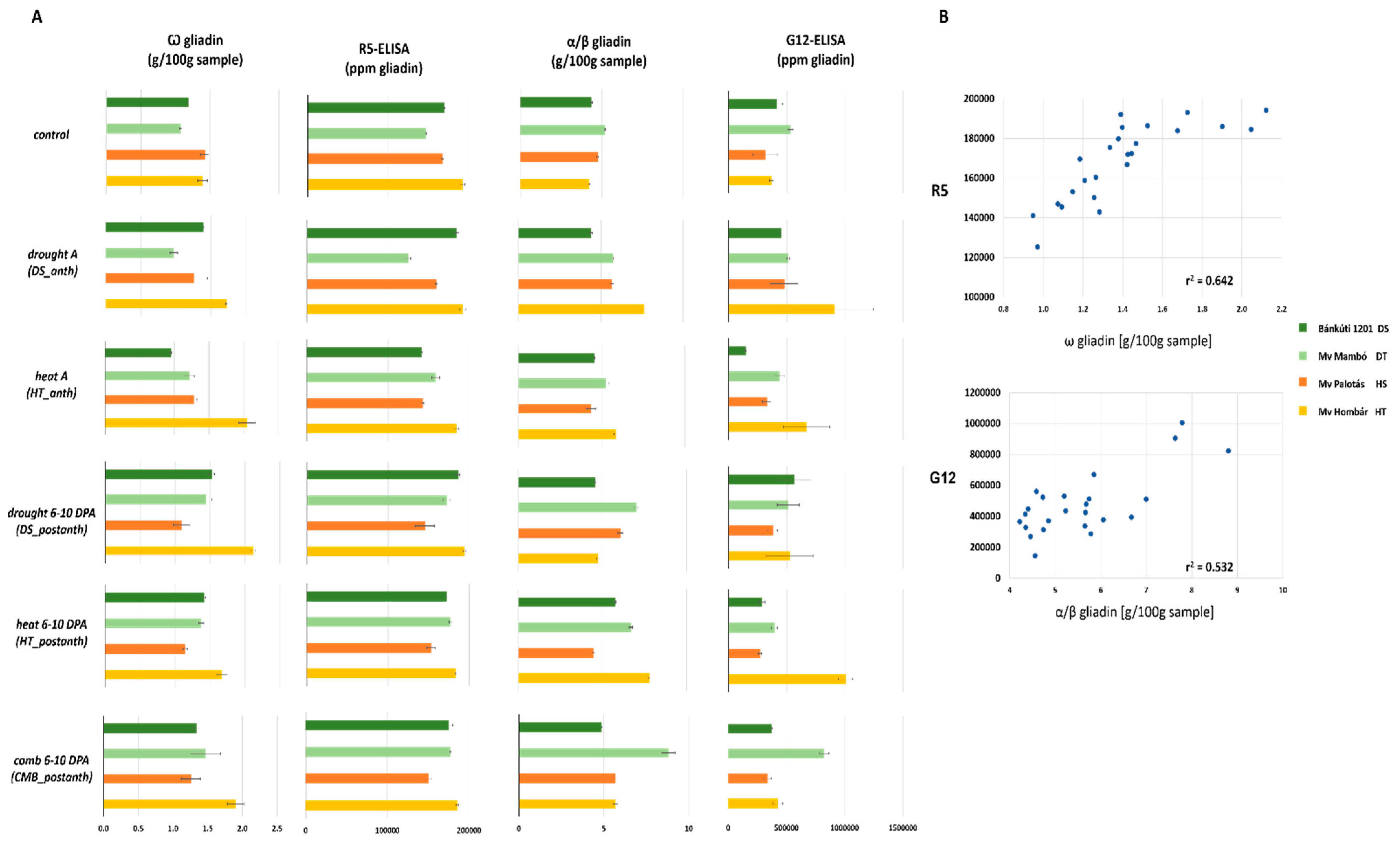
2.3. Effect of Abiotic Stress to Immunoreactive R5 and G12 Peptide Content
3. Materials and Methods
3.1. Plant Material
3.2. Growing Conditions and Abiotic Stress Treatments
3.3. Protein Content
3.4. RIDASCREEN R5 and ROMER G12 Sandwich ELISA Tests
3.5. Size Exclusion High Performance Liquid Chromatography (SE-HPLC)
3.6. Reversed-Phase High Performance Liquid Chromatography (RP-HPLC)
3.7. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Tatham, A.S.; Halford, N.G.; Barker, J.H.A.; Hannappel, U.; Gallois, P.; Thomas, M.; Kreis, M. Opportunities for manipulating the seed protein composition of wheat and barley in order to improve quality. Transgenic Res. 1994, 3, 3–12. [Google Scholar] [CrossRef]
- Sapirstein, H.D.; Fu, B.X. Intercultivar Variation in the Quantity of Monomeric Proteins, Soluble and Insoluble Glutenin, and Residue Protein in Wheat Flour and Relationships to Breadmaking Quality. Cereal Chem. 1998, 75, 500–507. [Google Scholar] [CrossRef]
- Khatkar, B.S.; Schofield, J.D. Molecular and physico-chemical basis of breadmaking-properties of wheat gluten proteins: A critical appraisal. J. Food Sci. Technol. 1997, 34, 85–102. [Google Scholar]
- Johansson, E.; Prieto-Linde, M.L.; Jönsson, J.Ö. Effects of Wheat Cultivar and Nitrogen Application on Storage Protein Composition and Breadmaking Quality. Cereal Chem. 2001, 78, 19–25. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Huang, X.W.; Wang, L.L.; Wei, L.; Wu, Z.H.; You, M.S.; Li, B.Y. Proteomic Analysis of Wheat Seed in Response to Drought Stress. J. Integr. Agric. 2014, 13, 919–925. [Google Scholar] [CrossRef]
- Juhász, A.; Haraszi, R.; Békés, F. Effects of Environmental Changes on the Allergen Content of Wheat Grain. In Wheat Quality for Improving Processing and Human Health; Igrejas, G., Ikeda, T.M., Guzmán, C., Eds.; Springer: Cham, Switzerland, 2020; ISBN 978-3-030-34163-3. [Google Scholar]
- Porter, J.L.; Xie, L.; Challinor, A.J.; Cochrane, K.; Howden, S.M.; Iqubal, M.M.; Lobell, D.B.; Travasso, M.I. Food security and food production systems. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 485–533. [Google Scholar]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Barnabás, B.; Jager, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Su, Y.H.; Dunham, S.J.; Rakszegi, M.; Bedo, Z.; McGrath, S.P.; Shewry, P.R. Variation in mineral micronutrient concentrations in grain of wheatlines of diverse origin. J. Cereal Sci. 2009, 49, 290–295. [Google Scholar] [CrossRef]
- Vensel, W.H.; Tanaka, C.K.; Cai, N.; Wong, J.H.; Buchanan, B.B.; Hurkman, W.J. Developmental changes in the metabolic protein profiles of wheat endosperm. Proteomics 2005, 5, 1594–1611. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B. Proteomics of Wheat Flour. In Proteomics in Food Science; Colgrave, M.L., Ed.; Academic Press: London, UK, 2017; pp. 57–73. ISBN 978-0-12-804007-2. [Google Scholar]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrigley, C.W.; Békés, F.; Bushuk, W. Chapter 1. Gluten: A balance of gliadin and glutenin. In Gliadin and Glutenin. The Unique Balance of Wheat Quality; Wrigley, C.W., Békés, F., Bushuk, W., Eds.; AACCI Press: St. Paul, MN, USA, 2006; pp. 3–33. ISBN 10-1-891127-51-9. [Google Scholar]
- Rakszegi, M.; Lovegrove, A.; Balla, K.; Lang, L.; Bedo, Z.; Shewry, P.R. Effect of heat and drought stress on the structure and composition of arabinoxylan and β-glucan in wheat grain. Carbohydr. Polym. 2014, 102, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Juhász, A.; Belova, T.; Florides, C.G.; Maulis, C.S.; Fischer, I.; Gell, G.Y.; Birinyi, Z.S.; Ong, J.; Keeble-Gagnére, G.; Maharajan, A.; et al. Genome mapping of seed-borne allergens and immunoresponsive proteins in wheat. Sci. Adv. 2018, 4, 8602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altenbach, S.B. New insights into the effects of high temperature, drought and post-anthesis fertilizer on wheat grain development. J. Cereal Sci. 2012, 56, 39–50. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, J.; Huang, X.; Guo, D.; Lou, H.; Hou, Z.; Su, M.; Liang, R.; Xie, C.; You, M.; et al. Differential effects of a post-anthesis heat stress on wheat (Triticum aestivum L.) grain proteome determined by iTRAQ. Sci. Rep. 2017, 7, 3468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, A.D.; Barmore, M.A. The effects of soil moisture stress on the yields and quality of flour from wheat. Agron. J. 1971, 63, 115–117. [Google Scholar] [CrossRef]
- Jiang, D.Y.; Yu, Z.W. Effects of soil water on yield and grain quality of wheat. J. Nucl. Agric. Sci. 2007, 21, 641–645. [Google Scholar]
- Zhou, J.; Liu, D.; Deng, X.; Zhen, S.; Wang, Z.; Yan, Y. Effects of water deficit on breadmaking quality and storage protein compositions in bread wheat (Triticum aestivum L.). J. Sci. Food Agric. 2018, 98, 4357–4368. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Jiang, D.; Xie, Z.J.; Dai, Y.B.; Jing, Q.; Cao, W.X. Effects of post-anthesis drought and waterlogging on grain quality traits in different specialty wheat varieties. J. Soil Water Conserv. 2004, 18, 193–196. [Google Scholar]
- Dai, Y.B.; Zhao, H.; Jing, Q.; Jiang, D.; Cao, W.X. Effects of high temperature and water stress during grain filling on grain protein and starch formation in winter wheat. Acta Ecol. Sin. 2006, 26, 3670–3676. [Google Scholar]
- Begcy, K.; Walia, H. Drought stress delays endosperm development and misregulates genes associated with cytoskeleton organization and grain quality proteins in developing wheat seeds. Plant Sci. 2015, 240, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.S.; Liang, X.N.; Li, X.; Wang, S.L.; Lv, D.W.; Ma, C.Y.; Li, X.H.; Ma, W.J.; Yan, Y.M. Wheat drought-responsive grain proteome analysis by linear and nonlinear 2-DE and MALDI-TOF mass spectrometry. Int. J. Mol. Sci. 2012, 13, 16065–16083. [Google Scholar] [CrossRef] [Green Version]
- Phakela, K.; van Biljon, A.; Wentzel, B.; Guzman, C.; Labuschagne, M.T. Gluten protein response to heat and drought stress in durum wheat as measured by reverse phase—High performance liquid chromatography. J. Cereal Sci. 2021, 100, 103267. [Google Scholar] [CrossRef]
- Iqbal, M.; Raja, N.I.; Yasmeen, F.; Hussain, M.; Ejaz, M.; Shah, M.A. Impacts of Heat Stress on Wheat: A Critical Review. Adv. Crop Sci. Technol. 2017, 5, 251. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, A.A.; Shakoor, A.; Goheer, A.; Qadir, T.; Khan, M.M.; Hussain, Z. Effect of cold and heat stress on different stages of wheat: A review. J. Glob. Innov. Agric. Soc. Sci. 2018, 6, 123–128. [Google Scholar]
- Wrigley, C.W.; Blumenthal, C.; Gras, P.W.; Barlow, E.W.R. Temperature variation during grain filling and changes in wheat-grain quality and chemistry. Aust. J. Plant Physiol. 1994, 21, 875–885. [Google Scholar] [CrossRef]
- Corbellini, M.; Canevar, M.G.; Mazza, L.; Ciaffi, M.; Lafiandra, D.; Borghi, B. Effect of the duration and intensity of heat shock during grain filling on dry matter and protein accumulation, technological quality and protein composition in bread and durum wheat. Aust. J. Plant Physiol. 1997, 24, 245–260. [Google Scholar] [CrossRef]
- Daniel, C.; Triboi, E. Effects of Temperature and Nitrogen Nutrition on the Grain Composition of Winter Wheat: Effects on Gliadin Content and Composition. J. Cereal Sci. 2000, 32, 45–56. [Google Scholar] [CrossRef]
- Bencze, S.; Veisz, O.; Bedő, Z. Effects of high atmospheric CO2 and heat stress on phytomass, yield and grain quality of winter wheat. Cereal Res. Commun. 2004, 32, 75–82. [Google Scholar] [CrossRef]
- Castro, M.; Peterson, C.J.; Dalla Rizza, M.; DÍaz Dellavalle, P.; Vázquez, D.; IbáÑez, V.; Ross, A. Influence of Heat Stress on Wheat Grain Characteristics and Protein Molecular Weight Distribution. In Wheat Production in Stressed Environments. Developments in Plant Breeding; Buck, H.T., Nisi, J.E., Salomón, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 12, pp. 365–371. ISBN 978-1-4020-5497-6. [Google Scholar] [CrossRef]
- Labuschagne, M.T.; Elago, O.; Koen, E. The influence of temperature extremes on some quality and starch characteristics in bread, biscuit and durum wheat. J. Cereal Sci. 2009, 49, 184–189. [Google Scholar] [CrossRef]
- Hrušková, M.; Švec, I. Wheat hardness in relation to other quality factors. Czech J. Food Sci. 2009, 27, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Li, F.; Cao, S.; Zhang, K. Genomic and functional genomics analyses of gluten proteins and prospect for simultaneous improvement of end-use and health-related traits in wheat. Theor. Appl. Genet. 2020, 133, 1521–1539. [Google Scholar] [CrossRef] [Green Version]
- Dupont, F.M.; Hurkman, W.J.; Vensel, W.H.; Chan, R.; Lopez, R.; Tanaka, C.K.; Altenbach, S.B. Differential accumulation of sulfur-rich and sulfur-poor wheat flour proteins is affected by temperature and mineral nutrition during grain development. J. Cereal Sci. 2006, 44, 101–112. [Google Scholar] [CrossRef]
- Hurkman, W.J.; Tanaka, C.K.; Vensel, W.H.; Thilmony, R.; Altenbach, S.B. Comparative proteomic analysis of the effect of temperature and fertilizer on gliadin and glutenin accumulation in the developing endosperm and flour from Triticum aestivum L. cv. Butte 86. Proteome Sci. 2013, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Jørgensen, A.D.; Li, H.; Søndergaard, I.; Finnie, C.; Svensson, B.; Jiang, D.; Wollenweber, B.; Jacobsen, S. Implications of High-Temperature Events and Water Deficits on Protein Profiles in Wheat (Triticum aestivum L. cv. Vinjett) Grain. Proteomics 2011, 11, 1684–1695. [Google Scholar] [CrossRef]
- Dupont, F.M.; Hurkman, W.J.; Vensel, W.H.; Tanaka, C.; Kothari, K.M.; Chung, O.K.; Altenbach, S.B. Protein accumulation and composition in wheat grains: Effects of mineral nutrients and high temperature. Eur. J. Agron. 2006, 25, 96–107. [Google Scholar] [CrossRef]
- Majoul, T.; Bancel, E.; Triboï, E.; Hamida, J.B.; Branlard, G. Proteomic analysis of the effect of heat stress on hexaploid wheat grain: Characterization of heat-responsive proteins from total endosperm. Proteomics 2003, 3, 175–183. [Google Scholar] [CrossRef]
- Gu, A.; Hao, P.; Lv, D.; Zhen, S.; Bian, Y.; Ma, C.; Xu, Y.; Zhang, W.; Yan, Y. Integrated Proteome Analysis of the Wheat Embryo and Endosperm Reveals Central Metabolic Changes Involved in the Water Deficit Response during Grain Development. J. Agric. Food Chem. 2015, 63, 8478–8487. [Google Scholar] [CrossRef]
- Hajheidari, M.; Eivazi, A.; Buchanan, B.B.; Wong, J.H.; Majidi, I.; Salekdeh, G.H. Proteomics uncovers a role for redox in drought tolerance in wheat. J. Proteome Res. 2007, 6, 1451–1460. [Google Scholar] [CrossRef]
- Balla, K.; Rakszegi, M.; Bencze, S.; Karsai, I.; Veisz, O. Effect of high temperature and drought on the composition of gluten proteins in Martonvásár wheat varieties. Acta Agron. Hung. 2010, 58, 343–353. [Google Scholar] [CrossRef]
- Dubois, B.; Bertin, P.; Hautier, L.; Muhovski, Y.; Escarnot, E.; Mingeot, D. Genetic and environmental factors affecting the expression of α-gliadin canonical epitopes involved in celiac disease in a wide collection of spelt (Triticum aestivum ssp. spelta) cultivars and landraces. BMC Plant Biol. 2018, 18, 262. [Google Scholar] [CrossRef] [Green Version]
- Hidy, D.; Barcza, Z.; Marjanović, H.; Ostrogović Sever, M.Z.; Dobor, L.; Gelybó, G.Y.; Fodor, N.; Pintér, K.; Churkina, G.; Running, S.; et al. Terrestrial Ecosystem Process Model Biome.BGCMuSo: Summary of improvements and new modeling possibilities. Geosci. Model Dev. 2016, 9, 4405–4437. [Google Scholar] [CrossRef] [Green Version]
- Micskei, G.Y.; Fodor, N.; Marton, C.S.; Bónis, P.; Árendás, T. Using long-term field experiment data to prepare a crop simulation model for climate impact studies. Appl. Ecol. Environ. Res. 2016, 14, 263–280. [Google Scholar] [CrossRef]
- Osman, A.A.; Uhlig, H.H.; Valdes, I.; Amin, M.; Mendez, E.; Mothes, T. A monoclonal antibody that recognizes a potential coeliac-toxic repetitive pentapeptide epitope in gliadins. Eur. J. Gastroenterol. Hepatol. 2001, 13, 1189–1193. [Google Scholar] [CrossRef]
- Valdés, I.; García, E.; Llorente, M.; Méndez, E. Innovative approach to low-level gluten determination in foods using a novel sandwich enzyme-linked immunosorbent assay protocol. Eur. J. Gastroenterol. 2003, 15, 465–474. [Google Scholar] [CrossRef]
- Morón, B.; Cebolla, Á.; Manyani, H.; Alvarez-Maqueda, M.; Megías, M.; Thomas, M.C.; López, M.C.; Sousa, C. Sensitive detection of cereal fractions that are toxic to celiac disease patients by using monoclonal antibodies to a main immunogenic wheat peptide. Am. J. Clin. Nutr. 2008, 87, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Tischner, T.; Kőszegi, B.; Veisz, O. Climatic programmes used on Martonvásár phytotron most effectly in recent years. Acta Agron. Hung. 1997, 45, 85–104. [Google Scholar]
- Lee, M.H. Association of Official Analytical Chemists. In Official Methods of Analysis of AOAC International, 16th ed.; Cunniff, P.A., Ed.; AOAC International: Washington, DC, USA, 1995. [Google Scholar]
- Batey, I.L.; Gupta, R.B.; MacRitchie, F. Use of size-exclusion high performance liquid chromatography in the study of wheat flour proteins: An improved chromatographic procedure. Cereal Chem. 1991, 68, 207–209. [Google Scholar] [CrossRef]
- Rakszegi, M.; Molnár, I.; Lovegrove, A.; Darkó, É.; Farkas, A.; Láng, L.; Bedő, Z.; Doležel, J.; Molnár-Láng, M.; Shewry, P. Addition of Aegilops U and M chromosomes affects protein and dietary fiber content of wholemeal wheat flour. Front. Plant Sci. 2017, 8, 1529. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.B.; Khan, K.; MacRitchie, F. Biochemical basis of flour properties in bread wheats. 1. Effects of variation in the quantity and size distribution of polymeric protein. J. Cereal Sci. 1993, 18, 23–41. [Google Scholar] [CrossRef]
- Larroque, O.R.; Bekes, F.; Wrigley, C.W.; Rathmell, W.G. Analysis of gluten proteins in grain and flour blends by RP-HPLC. In Wheat Gluten, Proceedings of the 7th 838 International Workshop Gluten, Bristol, UK, 2–6 April 2000; Shewry, P.R., Tatham, A.S., Eds.; Royal Society of Chemistry: Cambridge, UK, 2000; pp. 136–139. [Google Scholar]
- Marchylo, B.A.; Hatcher, D.W.; Kruger, J.E. Identification of wheat cultivars by reversed-phase high-performance liquid chromatography of storage proteins. Cereal Chem. 1988, 65, 28–40. [Google Scholar]
- Marchylo, B.A.; Kruger, J.E.; Hatcher, D.W. Quantitative reversed-phase high-performance liquid chromatographic analysis of wheat storage proteins as a potential quality prediction tool. J. Cereal Sci. 1989, 9, 113–130. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Height (cm) | Number of Spikelets Per Main Spike | Number of Grains Per Main Spike | Weight of Grains Per Main Spike (g) | Summarized Weight of Grains (g) | Thousand Kernel Weight | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | 38.21 | 17.79 | 33.30 | 0.69 | 1.61 | 20.37 | ||||||
| F for VARIETIES | 8081.42 | *** | 11.97 | *** | 3051.50 | *** | 929.52 | *** | 767.17 | *** | 848.35 | *** |
| F for TREATMENTS | 366.53 | *** | 1.10 | 18.29 | *** | 9.29 | *** | 5.33 | *** | 7.31 | *** | |
| F for VAR*TRE | 7.60 | *** | 6.14 | *** | 3.59 | *** | 17.83 | *** | 33.64 | *** | 25.74 | *** |
| LSD (cal by t) | 3.14 | 0.81 | 2.95 | 0.17 | 0.25 | 3.27 | ||||||
| Bánkúti 1201 (DS) | 63.9 | C | 16.8 | A | 28.4 | A | 0.87 | C | 2.11 | C | 30.61 | C |
| Mv Mambó (DT) | 29.3 | A | 19.1 | B | 33.0 | B | 0.62 | A | 1.31 | A | 17.87 | AB |
| Mv Palotás (HS) | 30.5 | A | 17.1 | A | 31.9 | B | 0.51 | A | 1.40 | A | 15.45 | A |
| Mv Hombár (HT) | 33.1 | B | 18.1 | B | 39.3 | C | 0.81 | B | 1.74 | B | 19.40 | B |
| LSD (cal by t) | 3.27 | 0.74 | 3.14 | 0.17 | 3.74 | 6.14 | ||||||
| Control | 41.8 | D | 18.2 | A | 35.5 | D | 0.80 | D | 1.96 | C | 23.21 | C |
| Drought A (DS_anth) | 37.2 | B | 17.6 | A | 31.4 | B | 0.61 | C | 1.10 | B | 18.23 | B |
| Heat A (HT_anth) | 34.8 | B | 18.5 | A | 37.8 | D | 1.11 | E | 3.01 | D | 29.35 | D |
| Comb A (CMB_anth) | 30.5 | A | 17.6 | A | 26.5 | A | 0.16 | A | 0.23 | A | 6.04 | A |
| Drought 6–10 DPA (DS_postanth) | 40.0 | C | 18.0 | A | 33.9 | C | 0.69 | C | 1.45 | B | 21.00 | C |
| Heat 6–10 DPA (HT_postanth) | 39.2 | C | 17.3 | A | 36.4 | D | 0.97 | D | 2.63 | D | 27.98 | D |
| Comb 6–10 DPA (CMB_postanth) | 42.5 | D | 17.5 | A | 30.5 | B | 0.39 | B | 0.61 | B | 13.88 | B |
| Protein% | Glu% | Gli% | GLU/GLI | UPP | HMW/LMW | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | 25.77 | 10.55 | 11.77 | 0.98 | 35.41 | 0.59 | ||||||
| F for VARIETIES | 1254.30 | *** | 133.54 | *** | 2098.38 | *** | 611.10 | *** | 236.10 | *** | 72.00 | *** |
| F for TREATMENTS | 4677.13 | *** | 60.92 | *** | 1152.01 | *** | 64.71 | *** | 116.30 | *** | 80.59 | *** |
| F for VAR × TRE | 946.18 | *** | 54.41 | *** | 312.58 | *** | 100.01 | *** | 39.78 | *** | 13.90 | *** |
| LSD (cal by t) | 0.13 | 0.12 | 0.11 | 0.01 | 0.86 | 0.015 | ||||||
| Bánkúti 1201 (DS) | 23.89 | A | 10.05 | A | 9.35 | A | 0.97 | B | 33.53 | B | 0.60 | C |
| Mv Mambó (DT) | 26.93 | C | 11.10 | D | 12.14 | B | 1.05 | D | 37.51 | C | 0.55 | A |
| Mv Palotás (HS) | 25.05 | B | 10.76 | C | 12.19 | B | 1.02 | C | 40.65 | D | 0.57 | B |
| Mv Hombár (HT) | 27.22 | D | 10.29 | B | 13.41 | C | 0.87 | A | 29.94 | A | 0.65 | D |
| LSD (cal by t) | 0.17 | 0.14 | 0.13 | 0.01 | 1.06 | 0.018 | ||||||
| Control | 22.90 | B | 10.43 | B | 10.47 | B | 0.98 | C | 35.10 | B | 0.62 | C |
| Drought A (DS_anth) | 20.83 | A | 10.47 | B | 10.03 | A | 0.96 | B | 35.68 | B | 0.62 | C |
| Heat A (HT_anth) | 25.19 | D | 10.21 | A | 11.76 | D | 0.98 | C | 40.62 | D | 0.67 | D |
| Comb A (CMB_anth) | 32.98 | G | ||||||||||
| Drought 6–10 DPA (DS_postanth) | 23.65 | C | 11.20 | D | 11.37 | C | 1.03 | D | 31.36 | A | 0.58 | B |
| Heat 6–10 DPA (HT_postanth) | 26.28 | E | 10.20 | A | 12.69 | E | 0.94 | A | 39.09 | C | 0.57 | B |
| Comb 6–10 DPA (CMB_postanth) | 28.59 | F | 10.81 | C | 14.31 | F | 0.96 | B | 30.60 | A | 0.50 | A |
| Omega | Alpha/Beta | Gamma | Bx | Dy | By | Dx | Ax | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gliadin% | HMW-GS% | |||||||||||||||
| Mean | 1.892 | 5.883 | 3.996 | 1.22 | 0.46 | 0.41 | 1.25 | 83.62 | ||||||||
| F for VARIETIES | 2834.29 | *** | 949.75 | *** | 1265.57 | *** | 92.62 | *** | 47.74 | *** | 103.44 | *** | 104.60 | *** | 9.80 | *** |
| F for TREATMENTS | 456.71 | *** | 284.66 | *** | 816.08 | *** | 11.10 | *** | 15.96 | *** | 18.44 | *** | 11.38 | *** | 17.88 | *** |
| F for VAR*TRE | 108.55 | *** | 105.20 | *** | 252.15 | *** | 31.14 | *** | 20.30 | *** | 11.84 | *** | 12.62 | *** | 11.8 | *** |
| LSD (cal by t) | 0.06 | 0.06 | 0.04 | 0.05 | 0.02 | 0.03 | 0.06 | 0.03 | ||||||||
| Bánkúti 1201 (DS) | 1.17 | A | 4.75 | A | 3.43 | A | 1.03 | A | 0.45 | B | 0.45 | C | 0.94 | A | 0.43 | B |
| Mv Mambó (DT) | 1.46 | B | 6.36 | C | 4.32 | B | 1.14 | B | 0.46 | B | 0.29 | A | 1.34 | CB | 0.59 | D |
| Mv Palotás (HS) | 1.41 | B | 6.95 | C | 3.83 | A | 1.41 | D | 0.41 | A | 0.38 | B | 1.39 | C | 0.40 | A |
| Mv Hombár (HT) | 3.53 | C | 5.48 | B | 4.41 | B | 1.30 | C | 0.54 | C | 0.52 | D | 1.30 | B | 0.53 | C |
| LSD (cal by t) | 0.07 | 0.07 | 0.04 | 0.06 | 0.03 | 0.03 | 0.07 | 0.03 | ||||||||
| Control | 1.47 | A | 5.31 | A | 3.70 | A | 1.24 | B | 0.45 | B | 0.41 | B | 1.21 | B | 0.47 | AB |
| Drought A (DS_anth) | 1.54 | B | 5.11 | A | 3.38 | A | 1.08 | A | 0.40 | A | 0.35 | A | 1.11 | A | 0.44 | A |
| Heat A (HT_anth) | 1.75 | C | 6.12 | B | 3.89 | A | 1.28 | B | 0.53 | C | 0.51 | C | 1.26 | CB | 0.47 | AB |
| Comb A (CMB_anth) | ||||||||||||||||
| Drought 6–10 DPA (DS_postanth) | 1.75 | C | 5.59 | AB | 4.03 | B | 1.22 | B | 0.47 | B | 0.39 | B | 1.26 | CB | 0.55 | D |
| Heat 6–10 DPA (HT_postanth) | 1.91 | D | 6.42 | B | 4.36 | C | 1.24 | B | 0.45 | B | 0.39 | B | 1.36 | D | 0.52 | C |
| Comb 6–10 DPA (CMB_postanth) | 2.95 | E | 6.74 | C | 4.63 | D | 1.26 | B | 0.48 | B | 0.41 | B | 1.28 | C | 0.49 | BC |
| R5 | G12 | |||
|---|---|---|---|---|
| ppm Gliadin% of the Control | ppm Gliadin% of the Control | |||
| Mean | 100.57 | 127.17 | ||
| F for VARIETIES | 173.39 | *** | 29.79 | *** |
| F for TREATMENTS | 26.90 | *** | 4.32 | *** |
| F for VAR*TRE | 31.69 | *** | 3.84 | *** |
| LSD (cal by t) | 2.70 | 2.97 | ||
| Bánkúti 1201 (DS) | 101.50 | C | 101.77 | B |
| Mv Mambó (DT) | 97.27 | B | 111.62 | C |
| Mv Palotás (HS) | 110.86 | D | 90.97 | A |
| Mv Hombár (HT) | 92.79 | A | 200.69 | D |
| LSD (cal by t) | 2.11 | 1.41 | ||
| Control | 100.00 | B | 100.00 | B |
| Drought A (DS_anth) | 98.35 | B | 144.24 | E |
| Heat A (HT_anth) | 92.92 | A | 97.16 | A |
| Comb A (CMB_anth) | 107.54 | D | 191.60 | E |
| Drought 6–10 DPA (DS_postanth) | 103.43 | D | 121.61 | D |
| Heat 6–10 DPA (HT_postanth) | 101.99 | C | 120.35 | C |
| Comb 6–10 DPA (CMB_postanth) | 102.05 | C | 120.39 | C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagy-Réder, D.; Birinyi, Z.; Rakszegi, M.; Békés, F.; Gell, G. The Effect of Abiotic Stresses on the Protein Composition of Four Hungarian Wheat Varieties. Plants 2022, 11, 1. https://doi.org/10.3390/plants11010001
Nagy-Réder D, Birinyi Z, Rakszegi M, Békés F, Gell G. The Effect of Abiotic Stresses on the Protein Composition of Four Hungarian Wheat Varieties. Plants. 2022; 11(1):1. https://doi.org/10.3390/plants11010001
Chicago/Turabian StyleNagy-Réder, Dalma, Zsófia Birinyi, Marianna Rakszegi, Ferenc Békés, and Gyöngyvér Gell. 2022. "The Effect of Abiotic Stresses on the Protein Composition of Four Hungarian Wheat Varieties" Plants 11, no. 1: 1. https://doi.org/10.3390/plants11010001
APA StyleNagy-Réder, D., Birinyi, Z., Rakszegi, M., Békés, F., & Gell, G. (2022). The Effect of Abiotic Stresses on the Protein Composition of Four Hungarian Wheat Varieties. Plants, 11(1), 1. https://doi.org/10.3390/plants11010001