Comparative Study of Temporal Changes in Pigments and Optical Properties in Sepals of Helleborus odorus and H. niger from Prebloom to Seed Production


Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling Locations
2.2. Sepal Morphological and Chemical Properties, and SLA
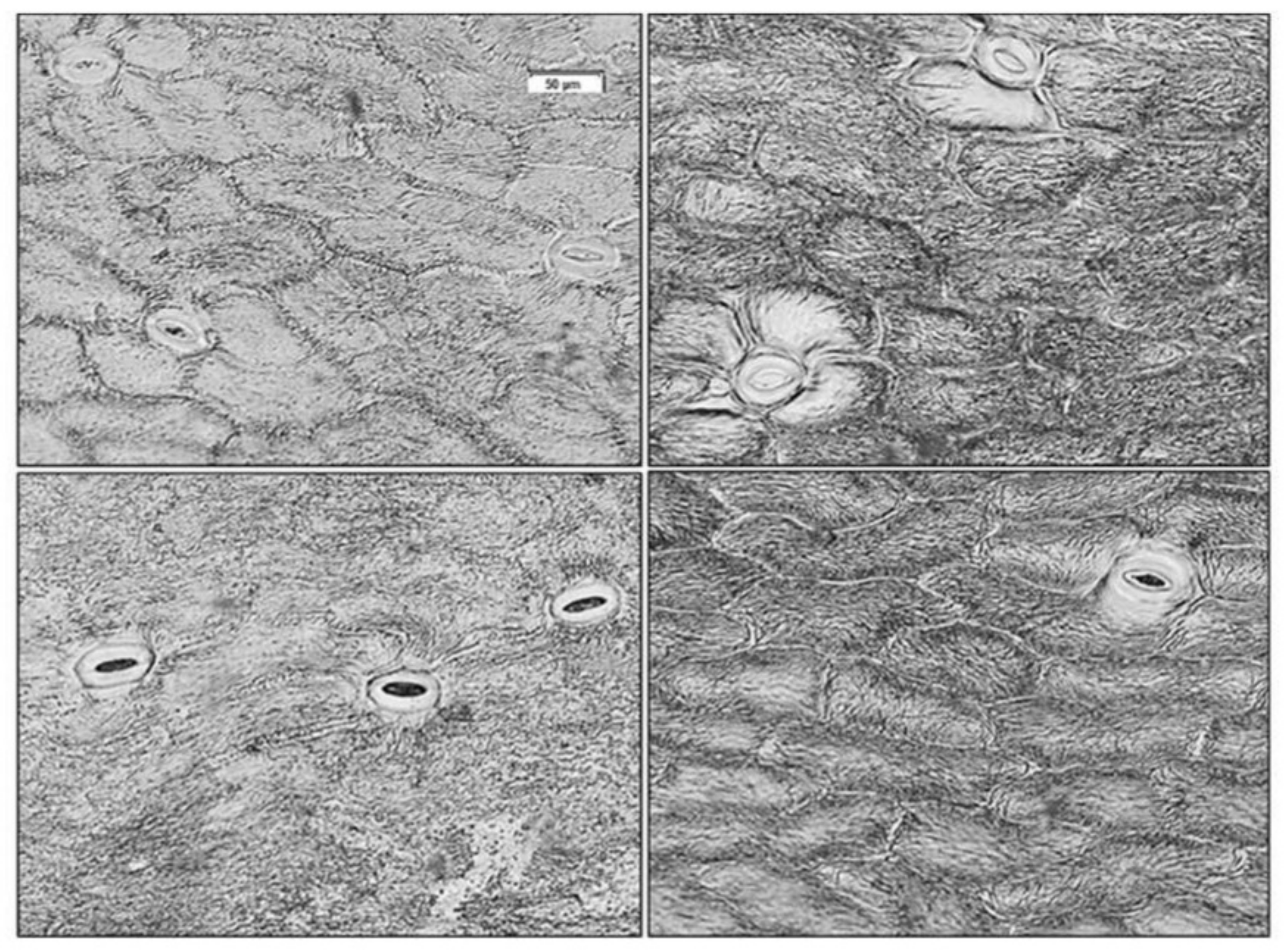
2.3. Sepal Optical Properties
2.4. Statistical Analyses
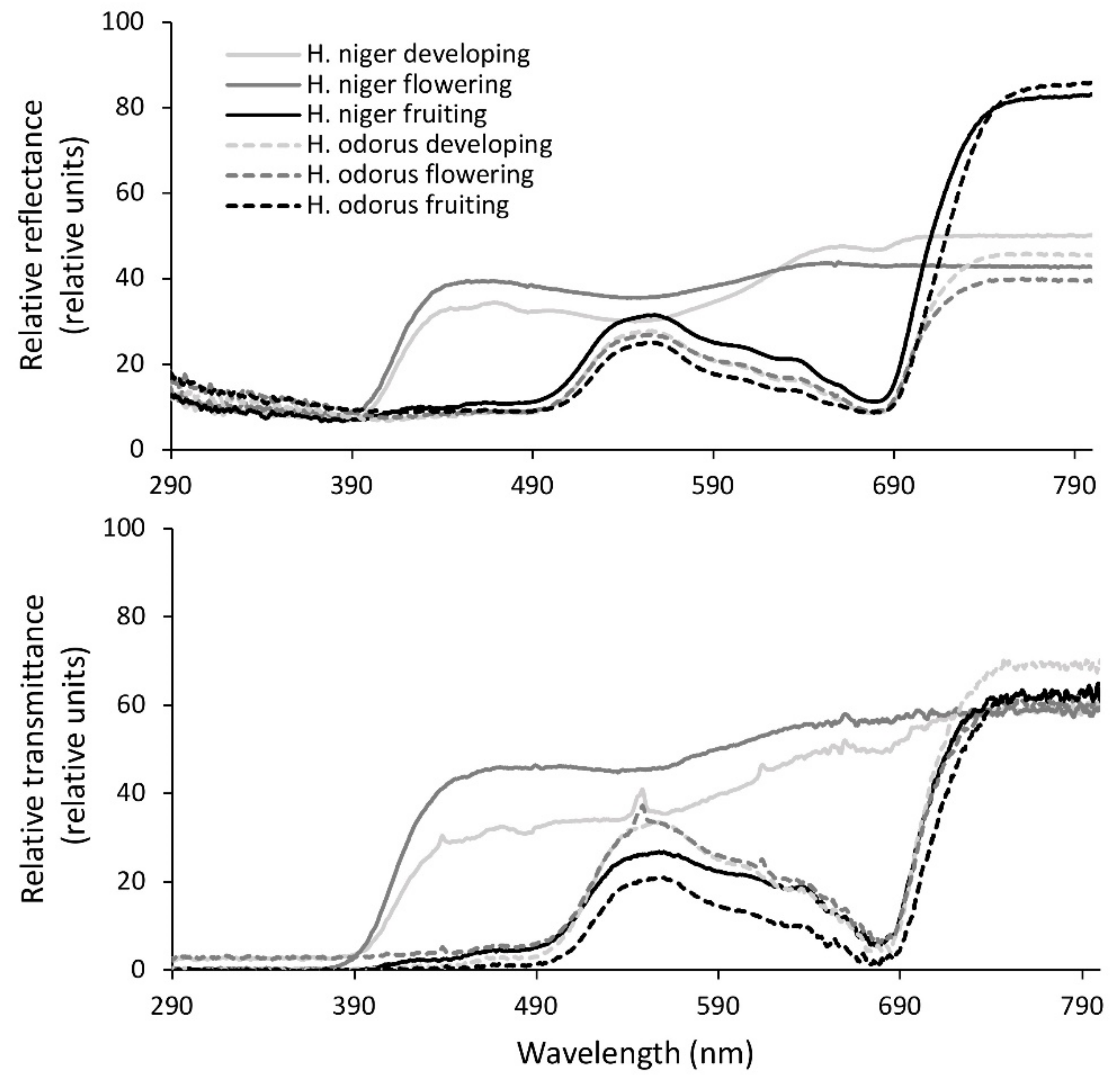
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Larcher, W. Physiological Plant Ecology, 4th ed.; Springer: Berlin, Germany, 2003; p. 513. [Google Scholar]
- Klančnik, K.; Levpušček, M.; Gaberščik, A. Variegation and red abaxial epidermis define the leaf optical properties of Cyclamen purpurascens. Flora 2016, 224, 87–95. [Google Scholar] [CrossRef]
- Gilliam, F. (Ed.) The Herbaceous Layer in Forests of Eastern North America; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Kim, H.J.; Jung, J.B.; Jang, Y.L.; Sung, J.H.; Park, P.S. Effects of experimental early canopy closure on the growth and reproduction of spring ephemeral Erythronium japonicum in a montane deciduous forest. J. Plant Biol. 2015, 58, 164–174. [Google Scholar] [CrossRef]
- Grašič, M.; Sovdat, T.; Gaberščik, A. Frond Optical Properties of the Fern Phyllitis scolopendrium Depend on Light Conditions in the Habitat. Plants 2020, 9, 1254. [Google Scholar] [CrossRef] [PubMed]
- Grašič, M.; Planinc, G.; Gaberščik, A. Bracts and basal leaves in Hacquetia epipactis differ in their spectral signatures. Biologia 2021, 76, 831–840. [Google Scholar] [CrossRef]
- Williams, K.; Koch, G.W.; Mooney, H.A. The carbon balance of flowers of Diplacus aurantiacus (Scrophulariaceae). Oecologia 1985, 66, 530–535. [Google Scholar] [CrossRef]
- Aschan, G.; Pfanz, H. Non-foliar photosynthesis—A strategy of additional carbon acquisition. Flora Morphol. Distrib. Funct. Ecol. Plants 2003, 198, 81–97. [Google Scholar] [CrossRef]
- Esteban, R.; Fernández-Marín, B.; Becerril, J.M.; García-Plazaola, J.I. Photoprotective implications of leaf variegation in E. denscanis L. and P. officinalis L. J. Plant Physiol. 2008, 165, 1255–1263. [Google Scholar] [CrossRef]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The Evolution of Plant Functional Variation: Traits, Spectra, and Strategies. Int. J. Plant Sci. 2003, 164, S143–S164. [Google Scholar] [CrossRef]
- Yoshimura, H.; Zhu, H.; Wu, Y.; Ma, R. Spectral properties of plant leaves pertaining to urban landscape design of broad-spectrum solar ultraviolet radiation reduction. Int. J. Biometeorol. 2010, 54, 179–191. [Google Scholar] [CrossRef]
- Klančnik, K.; Mlinar, M.; Gaberščik, A. Heterophylly results in a variety of “spectral signatures” in aquatic plant species. Aquat. Bot. 2012, 98, 20–26. [Google Scholar] [CrossRef]
- Liew, O.; Chong, P.; Li, B.; Asundi, A. Signature Optical Cues: Emerging Technologies for Monitoring Plant Health. Sensors 2008, 8, 3205–3239. [Google Scholar] [CrossRef] [Green Version]
- Bavcon, J.; Vreš, B.; Mlinar, C.; Smolej, H.; Batič, F.; Praprotnik, N.; šiftar, A. Telohi (Helleborus L.) v Sloveniji = Helleborus (Helleborus L.) in Slovenia; Založba Primus: Brežice, Slovenia, 2012. [Google Scholar]
- Fassou, G.; Kougioumoutzis, K.; Iatrou, G.; Trigas, P.; Papasotiropoulos, V. Genetic Diversity and Range Dynamics of Helleborus odorus subsp. cyclophyllus under Different Climate Change Scenarios. Forests 2020, 11, 620. [Google Scholar] [CrossRef]
- Guitián, J.; Larrinaga, A.R. The role of post-floral persistent perianth in Helleborus viridis subsp. occidentalis (Ranunculaceae). Nord. J. Bot. 2014, 32, 852–857. [Google Scholar] [CrossRef] [Green Version]
- Šušek, A.; Ivančič, A. Pollinators of Helleborus niger in Slovenian naturally occuring populations. Acta Agric. Slov. 2006, 87, 205–211. [Google Scholar]
- Vesprini, J.L.; Pacini, E. Pollination ecology in sympatric winter flowering Helleborus (Ranunculaceae). Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 627–632. [Google Scholar] [CrossRef]
- Shahri, W.; Tahir, I.; Islam, S.T.; Bhat, M.A. Physiological and biochemical changes associated with flower development and senescence in so far unexplored Helleborus orientalis Lam. cv. Olympicus. Physiol. Mol. Biol. Plants 2011, 17, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Schmitzer, V.; Mikulic-Petkovsek, M.; Stampar, F. Sepal phenolic profile during Helleborus niger flower development. J. Plant Physiol. 2013, 170, 1407–1415. [Google Scholar] [CrossRef]
- Wraber, T.; Jogan, N.; Podobnik, A.; Ravnik, V.; Turk, B.; Vreš, B. Mala Flora Slovenije: Ključ za Določanje Praprotnic in Semenk; Tehniška Založba Slovenije: Ljubljana, Slovenia, 2007. [Google Scholar]
- Salopek-Sondi, B.; Magnus, V. Developmental Studies in the Christmas Rose (Helleborus niger L.). Int. J. Plant Dev. Biol. 2007, 1, 151–159. [Google Scholar]
- Salopek-Sondi, B. Reproductive Development of the Christmas Rose (Helleborus niger L.): The Role of Plant Hormones. Croat. Chem. Acta 2011, 84, 277–285. [Google Scholar] [CrossRef]
- Bavcon, J. Helleborus Diversity in Slovenia. In European Botanic Gardens in a Changing World: Insights into Eurogard VI; Krigas, N., Tsoktouridis, G., Cook, C.-M., Mylona, P., Maloupa, E., Eds.; Balkan Botanic Garden of Kroussia (Hellenic Agriculture Organisation-Demeter) and Botanic Gardens Conservation International: Thessaloniki, Greece, 2014; pp. 53–62. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, 171–178. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Extraction of Phtosynthetic Tissues: Chlorophylls and Carotenoids. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.2.1–F4.2.6. [Google Scholar] [CrossRef]
- Drumm, H.; Mohr, H. The mode of interaction between blue (UV) light photoreceptor and phytochrome in anthocyanin formation of the Sorghum seedling. Photochem. Photobiol. 1978, 27, 241–248. [Google Scholar] [CrossRef]
- Caldwell, M.M. Solar Ultraviolet Radiation as an Ecological Factor for Alpine Plants. Ecol. Monogr. 1968, 38, 243–268. [Google Scholar] [CrossRef]
- Klančnik, K.; Vogel-Mikuš, K.; Gaberščik, A. Silicified structures affect leaf optical properties in grasses and sedge. J. Photochem. Photobiol. B Biol. 2014, 130, 1–10. [Google Scholar] [CrossRef]
- Salopek-Sondi, B.; Kovač, M.; Ljubešić, N.; Magnus, V. Fruit initiation in Helleborus niger L. triggers chloroplast formation and photosynthesis in the perianth. J. Plant Physiol. 2000, 157, 357–364. [Google Scholar] [CrossRef]
- Salopek-Sondi, B. Developing fruit direct post-floral morphogenesis in Helleborus niger L. J. Exp. Bot. 2002, 53, 1949–1957. [Google Scholar] [CrossRef]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin, Germany, 2003. [Google Scholar]
- Aschan, G.; Pfanz, H.; Vodnik, D.; Batič, F. Photosynthetic performance of vegetative and reproductive structures of green hellebore (Helleborus viridis L. agg.). Photosynthetica 2005, 43, 55–64. [Google Scholar] [CrossRef]
- Brcko, A.; Pěnčík, A.; Magnus, V.; Prebeg, T.; Mlinarić, S.; Antunović, J.; Lepeduš, H.; Cesar, V.; Strnad, M.; Rolčík, J.; et al. Endogenous Auxin Profile in the Christmas Rose (Helleborus niger L.) Flower and Fruit: Free and Amide Conjugated IAA. J. Plant Growth Regul. 2012, 31, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.M. Post-floral perianth functionality: Contribution of persistent sepals to seed development in Helleborus foetidus (Ranunculaceae). Am. J. Bot. 2005, 92, 1486–1491. [Google Scholar] [CrossRef]
- Klančnik, K.; Pančić, M.; Gaberščik, A. Leaf optical properties in amphibious plant species are affected by multiple leaf traits. Hydrobiologia 2014, 737, 121–130. [Google Scholar] [CrossRef]
- Van der Kooi, C.J.; Dyer, A.G.; Kevan, P.G.; Lunau, K. Functional significance of the optical properties of flowers for visual signalling. Ann. Bot. 2019, 123, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kay, Q.O.N.; Daoud, H.S.; Stirton, C.H. Pigment distribution, light reflection and cell structure in petals. Bot. J. Linn. Soc. 1981, 83, 57–83. [Google Scholar] [CrossRef]
- Peitsch, D.; Fietz, A.; Hertel, H.; de Souza, J.; Ventura, D.F.; Menzel, R. The spectral input systems of hymenopteran insects and their receptor-based colour vision. J. Comp. Physiol. A 1992, 170, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Kelber, A.; Vorobyev, M.; Osorio, D. Animal colour vision—Behavioural tests and physiological concepts. Biol. Rev. Camb. Philos. Soc. 2003, 78, 81–118. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Species and Developmental Phase | |||||
|---|---|---|---|---|---|---|
| Developing Phase | Flowering Phase | Fruiting Phase | ||||
| H. odorus | H. niger | H. odorus | H. niger | H. odorus | H. niger | |
| Chlorophyll a (mg cm−2) | 1.65 ± 0.36 b | 0.58 ± 0.29 a | 1.14 ± 0.50 ab | 1.53 ± 0.53 b | 4.51 ± 1.36 d | 3.14 ± 0.91 c |
| Chlorophyll b (mg cm−2) | 2.37 ± 0.84 b | 0.65 ± 0.68 a | 1.26 ± 0.39 a | 1.18 ± 0.44 a | 4.35 ± 2.00 c | 2.59 ± 0.78 b |
| Carotenoids (mg cm−2) | 0.37 ± 0.13 a | 0.34 ± 0.13 a | 0.66 ± 0.16 b | 0.71 ± 0.18 b | 1.01 ± 0.28 c | 1.08 ± 0.26 c |
| Anthocyanins (au cm−2) | 0.03 ± 0.01 b | 0.01 ± 0.01 a | 0.05 ± 0.02 c | 0.07 ± 0.02 cd | 0.06 ± 0.02 cd | 0.08 ± 0.02 d |
| UV-B-AS (au cm−2) | 0.58 ± 0.09 a | 0.58 ± 0.17 a | 0.65 ± 0.14 ab | 0.52 ± 0.09 a | 0.78 ± 0.20 bc | 0.88 ± 0.22 c |
| UV-A-AS (au cm−2) | 1.72 ± 0.14 b | 1.61 ± 0.07 a | 1.65 ± 0.10 ab | 1.59 ± 0.07 a | 1.65 ± 0.04 ab | 1.59 ± 0.11 a |
| Sepal thickness (µm) | 183± 34 a | 226 ± 61 bc | 194 ± 11 a | 205 ± 21 ab | 240 ± 17 cd | 266 ± 13 d |
| SLA (cm2 mg−1) | 1.05 ± 0.17 c | 1.18 ± 0.35 c | 1.04 ± 0.23 bc | 1.01 ± 0.23 abc | 0.84 ± 0.12 ab | 0.83 ± 0.09 a |
| Fv/Fm (au) | 0.77 ± 0.03 c | 0.31 ± 0.20 a | 0.76 ± 0.02 c | 0.45 ± 0.16 b | 0.79 ± 0.01 c | 0.79 ± 0.03 c |
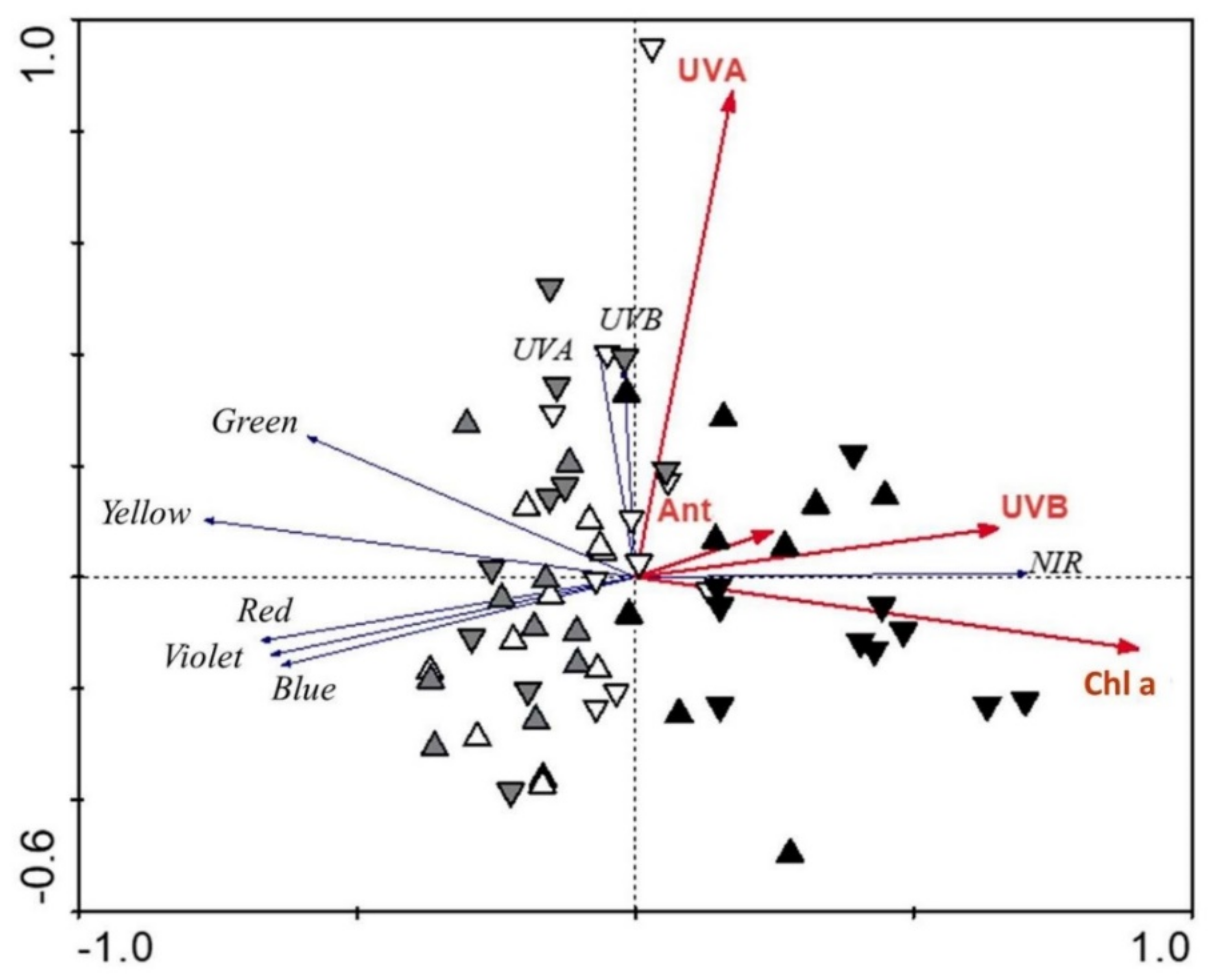
| Sepal Traits | Fv/Fm | Anthocyanins | Chl. a | Chl. b | UV-B-AS | UV-A-AS | |
|---|---|---|---|---|---|---|---|
| UV−B | D | 0.58 | 0.35 | 0.54 | 0.63 | 0.14 | 0.17 |
| FL | 0.63 | −0.23 | −0.25 | 0.11 | 0.47 | 0.35 | |
| FR | −0.06 | 0.36 | 0.25 | −0.28 | 0.29 | −0.56 | |
| UV−A | D | 0.43 | 0.17 | 0.34 | 0.43 | 0.27 | 0.23 |
| FL | 0.57 | −0.13 | −0.17 | 0.05 | 0.43 | 0.37 | |
| FR | −0.24 | −0.36 | −0.45 | −0.31 | −0.01 | 0.25 | |
| Violet | D | −0.70 | −0.61 | −0.73 | −0.57 | −0.06 | −0.50 |
| FL | −0.82 | 0.37 | 0.33 | −0.18 | −0.52 | −0.33 | |
| FR | −0.26 | −0.48 | −0.51 | −0.16 | −0.09 | 0.44 | |
| Blue | D | −0.71 | −0.61 | −0.72 | −0.55 | −0.08 | −0.48 |
| FL | −0.84 | 0.33 | 0.33 | −0.19 | −0.51 | −0.35 | |
| FR | −0.32 | −0.50 | −0.54 | −0.22 | −0.31 | 0.48 | |
| Green | D | −0.34 | −0.38 | −0.37 | −0.16 | −0.15 | −0.34 |
| FL | −0.73 | 0.33 | 0.29 | −0.30 | −0.45 | −0.35 | |
| FR | −0.29 | −0.50 | −0.52 | −0.08 | −0.34 | 0.55 | |
| Yellow | D | −0.64 | −0.59 | −0.66 | −0.45 | −0.12 | −0.43 |
| FL | −0.80 | 0.32 | 0.32 | −0.24 | −0.48 | −0.41 | |
| FR | −0.27 | −0.50 | −0.50 | −0.03 | −0.28 | 0.58 | |
| Red | D | −0.85 | −0.70 | −0.85 | −0.70 | −0.08 | −0.45 |
| FL | −0.85 | 0.29 | 0.34 | −0.15 | −0.50 | −0.41 | |
| FR | 0.10 | 0.26 | 0.23 | 0.22 | −0.23 | 0.01 | |
| NIR | D | −0.74 | −0.62 | −0.66 | −0.34 | −0.04 | −0.41 |
| FL | −0.70 | 0.02 | 0.13 | −0.19 | −0.34 | −0.37 | |
| FR | 0.10 | 0.26 | 0.23 | 0.22 | −0.23 | 0.01 |
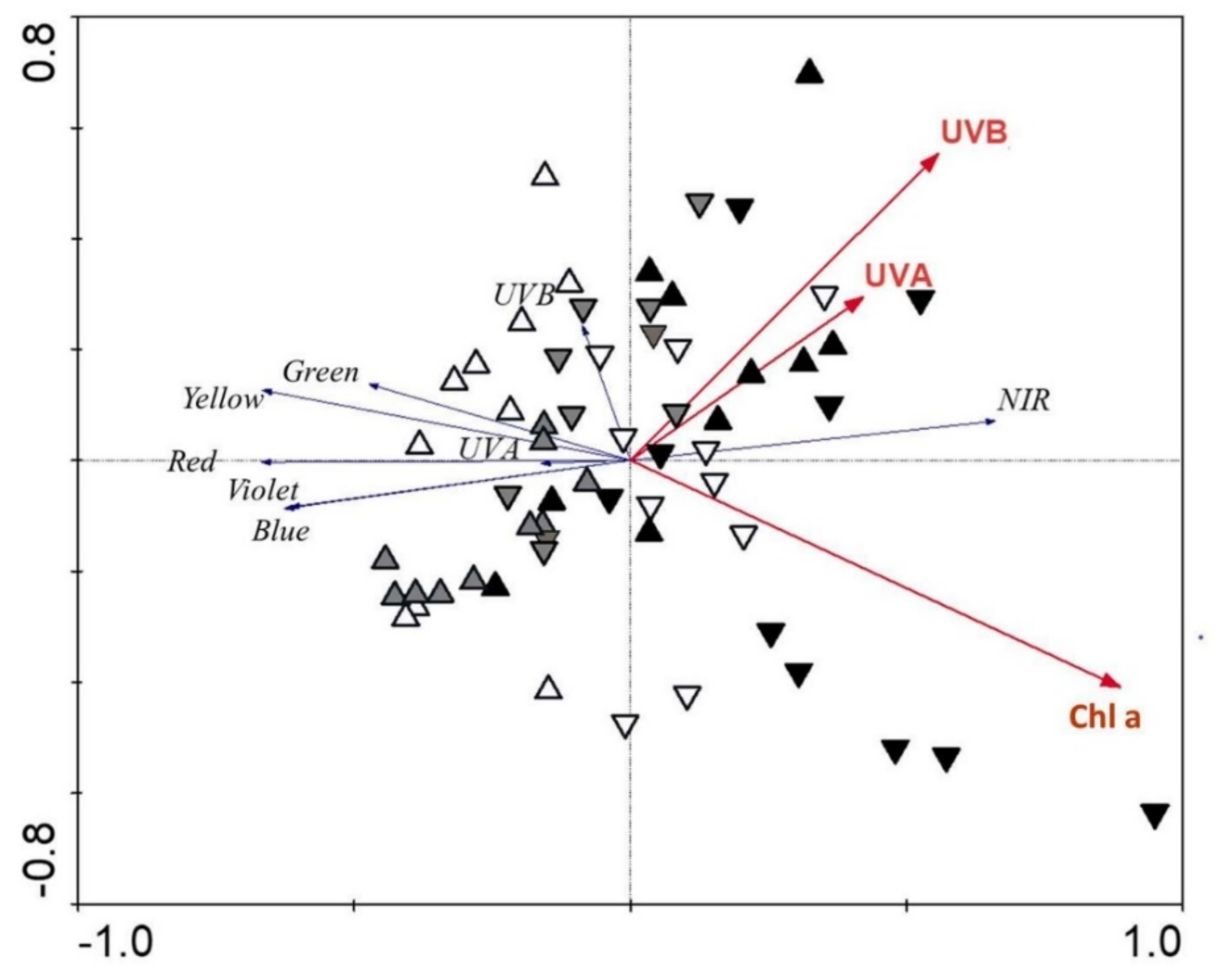
| Sepal Traits | Fv/Fm | Anthocyanins | Chl. a | Chl. b | Carotenoids | UV-B-AS | UV-A-AS | |
|---|---|---|---|---|---|---|---|---|
| UV-B | D | −0.30 | 0.10 | −0.26 | −0.25 | 0.23 | 0.52 | −0.08 |
| FL | −0.39 | 0.28 | 0.41 | 0.10 | 0.37 | −0.00 | −0.36 | |
| FR | −0.11 | −0.02 | −0.13 | 0.01 | −0.26 | −0.17 | 0.24 | |
| UV-A | D | −0.58 | −0.60 | −0.65 | −0.48 | −0.42 | −0.32 | −0.32 |
| FL | −0.07 | 0.21 | 0.16 | 0.03 | −0.04 | 0.05 | −0.18 | |
| FR | −0.37 | −0.35 | −0.43 | 0.11 | 0.44 | −0.33 | 0.67 | |
| Violet | D | −0.75 | −0.62 | −0.78 | −0.63 | −0.18 | −0.04 | −0.43 |
| FL | −0.80 | 0.36 | 0.30 | −0.21 | 0.20 | −0.55 | −0.40 | |
| FR | −0.34 | −0.29 | −0.38 | 0.20 | 0.49 | −0.43 | 0.73 | |
| Blue | D | −0.74 | −0.65 | −0.77 | −0.61 | −0.20 | −0.12 | −0.42 |
| FL | −0.83 | 0.34 | 0.30 | −0.22 | 0.21 | −0.56 | −0.39 | |
| FR | −0.28 | −0.32 | −0.38 | −0.06 | 0.40 | −0.28 | 0.55 | |
| Green | D | −0.39 | −0.31 | −0.46 | −0.23 | −0.08 | −0.07 | −0.12 |
| FL | −0.78 | 0.37 | 0.26 | −0.28 | 0.19 | −0.55 | −0.37 | |
| FR | −0.25 | −0.38 | −0.43 | 0.02 | 0.42 | −0.34 | 0.63 | |
| Yellow | D | −0.61 | −0.55 | −0.67 | −0.45 | −0.17 | −0.17 | −0.21 |
| FL | −0.81 | 0.39 | 0.30 | −0.24 | 0.20 | −0.54 | −0.35 | |
| FR | −0.26 | −0.39 | −0.44 | 0.11 | 0.46 | −0.41 | 0.71 | |
| Red | D | −0.77 | −0.73 | −0.80 | −0.64 | −0.26 | −0.26 | −0.33 |
| FL | −0.83 | 0.37 | 0.34 | −0.18 | 0.21 | −0.54 | −0.35 | |
| FR | −0.09 | −0.22 | −0.16 | −0.10 | 0.30 | 0.07 | 0.29 | |
| NIR | D | 0.07 | −0.03 | 0.06 | 0.12 | −0.49 | −0.27 | 0.43 |
| FL | −0.30 | 0.22 | 0.02 | −0.10 | 0.44 | −0.35 | 0.04 | |
| FR | −0.03 | 0.34 | 0.26 | −0.25 | −0.29 | 0.29 | −0.57 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grašič, M.; Dacar, M.; Gaberščik, A. Comparative Study of Temporal Changes in Pigments and Optical Properties in Sepals of Helleborus odorus and H. niger from Prebloom to Seed Production. Plants 2022, 11, 119. https://doi.org/10.3390/plants11010119
Grašič M, Dacar M, Gaberščik A. Comparative Study of Temporal Changes in Pigments and Optical Properties in Sepals of Helleborus odorus and H. niger from Prebloom to Seed Production. Plants. 2022; 11(1):119. https://doi.org/10.3390/plants11010119
Chicago/Turabian StyleGrašič, Mateja, Maja Dacar, and Alenka Gaberščik. 2022. "Comparative Study of Temporal Changes in Pigments and Optical Properties in Sepals of Helleborus odorus and H. niger from Prebloom to Seed Production" Plants 11, no. 1: 119. https://doi.org/10.3390/plants11010119
APA StyleGrašič, M., Dacar, M., & Gaberščik, A. (2022). Comparative Study of Temporal Changes in Pigments and Optical Properties in Sepals of Helleborus odorus and H. niger from Prebloom to Seed Production. Plants, 11(1), 119. https://doi.org/10.3390/plants11010119