Abstract
The taxonomy of the subfamily Linoideae at the intergeneric and section levels has been questioned throughout the years, and the evolution of floral characters remains poorly understood. In particular, the evolution of flower color is still uncertain, despite its ecological importance and being one of the most variable and striking traits in Angiospermae. We evaluated the phylogenetic relationships of the genera and sections and used the phylogeny to reconstruct the ancestral state of flower color. The results suggest reevaluating the taxonomic status of segregated genera and re-incorporating them into Linum. Four of the five sections currently accepted were recovered as monophyletic (Cathartolinum, Dasylinum, Linum, and Syllinum). We propose accepting the section Stellerolinon and reevaluating Linopsis, whose representatives were recovered in three separate clades. The ancestral flower color for Linoideae was yellow-white. The flower colors purple and yellow-white were recovered at the deepest nodes of the two main clades. Pink, blue, and red colors were the most recent to evolve. These results appear to be related to diversification events, biogeographical history, and ecological aspects of the subfamily. Our reconstruction constitutes the first plausible scenario that explores the evolution of flower color, leading to new testable hypotheses for future research on the flax group.
1. Introduction
Linoideae Arnott is one of the two known subfamilies in Linaceae and the larger of the two. The subfamily is distributed mainly in temperate regions, with the greatest diversity concentrated in the Mediterranean Basin and Southwestern Asia, with some representatives extending to tropical and subtropical latitudes [1,2]. It comprises approximately 210 species in 8 genera [1,3], with Linum L. being the most diverse and commercially important [2,4,5]. Recognition of the genus is attributed to the cultivation of Linum usitatissimum L., commonly known as flax or linseed [6]. However, some wild species have recently been used for other purposes, such as Linum perenne L. and L. grandiflorum Desf, which have been used as ornamentals [7,8], and L. rupestre (A. Gray) Engelm. ex A. Gray and L. scabrellum Planch., which have been used for their medicinal properties [9,10,11].
The species of the genus have great morphological diversity, due in part to the wide range of environments where they live, so species can be difficult to characterize [12,13,14]. Therefore, Linum has been subject to several taxonomic changes. For example, although Linoideae was initially organized in the genera Anisadenia Wall., Linum, Radiola Hill, and Reinwardtia Dumort, for Planchon [15,16], Tirpitzia Hallier f. was segregated from Reinwardtia due to the presence of species with tubular corollas characteristic and winged seeds [17]. Furthermore, the monotypic genera Cliococca Bab. and Sclerolinon C. M. Rogers, as well as the genus Hesperolinon (A. Gray) Small with 13 species, originally confined in Linum, were segregated from this genus based on morphological characteristics [18,19,20,21].
Cliococca selaginoides (Lam.) C. M. Rogers & Mildner was initially described as Linum selaginoides Lam. [22], and later, Babington [23], based on the description of cultivated plants at the Cambridge Botanical Garden, considered that there were sufficient characteristics to establish the existence of a new genus: Cliococca. Rogers and Mildner [21] reassessed the genera and validated their segregation. Similarly, Hesperolinon was also described as a section of Linum by Gray [24]. Small [18], based on morphological characters, such as the number of carpels and styles, circumscribed the section at the genus level with H. californicum (Benth.) Small. as the type. This status was supported by Sharsmith [19], who detailed the description of this genus. However, its separation from the rest of the flaxes is controversial. Although the genus shows clear morphological differences [18,25], there are no clear differences at the molecular level, and its return to Linum has been suggested [1,26]. For its part, Sclerolinon digynum (A. Gray) C. M. Rogers was described under the basonym Linum digynum (A. Gray). Brewer and Watson [27] and Trelease [28,29] noted the resemblance of this species to Hesperolinon. It was the first time that the species was separated from flaxes. A century later, Sharsmith [19] pointed out the need to reassess this species to accurately determine what genus it belongs to and validated Sclerolinon as a genus.
The studies of Planchon [15,16], Winkler [30], and Ockendon and Walters [31] constituted the basis for the establishment of the five sections of the Linum genus currently recognized in most taxonomic works: Cathartolinum (Rchb.) Grised, Dasylinum (Planch.) Juz., Linum (Planch.) Juz, Linopsis (Rchb.), and Syllinum Griseb. However, phylogenetic analyses show discrepancies in the validity of some sections. McDill et al. [2], from molecular characters, recovered only two of the five sections, Dasylinum and Cathartolinum, as natural groups. On the other hand, McDill and Simpson [1], from plastid DNA data, recovered Linum, Dasylinum, Syllinum, and Cathartolinum as natural groups. Meanwhile, intending to track the evolution of style polymorphism, Ruiz-Martín et al. [13] and Maguilla et al. [32] recovered only three sections as monophyletic (Dasylinum, Syllinum, and Cathartolinum). Furthermore, other classifications at the section level have recently been evaluated in the group [33]. Therefore, shedding light on this debate is necessary.
The evolution of heterostyly in Linum has been extensively studied. Nonetheless, the evolution of other floral features, such as the widely variable color of the corolla among species, has not been studied. Flower color diversity is one of the most striking features of angiosperm radiation. Moreover, it appears to be one of the most evolutive changed traits, often differing between sister species [34,35].
Corolla color spots, patterns, and lines are common across angiosperms and, in ecological terms, are important for mediating plant–animal interactions. For example, color patterns on petals can enhance the pollinator’s ability to detect flowers [36,37], orient it to floral rewards [38], and increase the likelihood of effective pollination [39].
The genetic changes that lead to flower color transitions have been studied in detail [40,41,42]. They are valuable not only for pollination studies [43] but also in evolution. The evolution of character states can be revealed when such states are traced in phylogenetic trees [44]. To clarify the intra- and infrageneric evolutionary relationships of the subfamily Linoideae and contribute for the first time to the knowledge of the evolution of flower color in the group, the aims of this study were: (a) to elucidate the phylogenetic relationships between the genera of Linoideae using molecular characters, (b) to evaluate the phylogenetic position of the five sections of Linum, and (c) to reconstruct the ancestral flower color of the lineages of the subfamily.
2. Results
2.1. Phylogenetic Analysis
The concatenated dataset consisted of 2989 characters, 1631 of which were conserved sites, 1316 were variable sites, and 951 were parsimony-informative. The parsimony analysis (PA) resulted in 240 most-parsimonious trees with a length of 3347, a consistency index (Ci), excluding uninformative characters, of 0.44, and a retention index (Ri) of 0.86. Although significant congruence was observed between topologies derived from Bayesian Inference (BI, Figure 1) and PA analyses (Supplementary Figure S3), with only minor differences in the placement of some taxa, in the topology obtained with parsimony, some poorly resolved clades were recovered. Consequently, hereafter, only the BI tree is described and discussed with the corresponding support values (Figure 1).
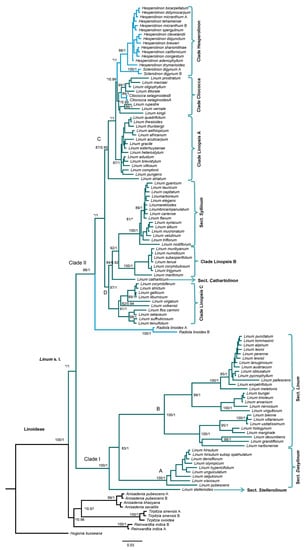
Figure 1.
Bayesian Inference tree of Linoideae based on combined plastid (ndhF, matK, and trnL-F) and ITS dataset. The bootstrap values (BS; left) and posterior probabilities (PP; right) are labeled above the branches. Only support values of BS ≥ 80% and PP ≥ 0.9 are shown. * No support values. A representative of Hugonia (Hugonideae) was used as an outgroup.
The subfamily Linoideae was recovered as a well-supported monophyletic group (BS = 100%, PP = 1), with their members resolved into two major clades. The first one comprised the Anisadenia and Tirpitzia genera, forming a monophyletic group with moderate support (BS ≤ 80%, PP = 0.97), with Reinwardtia as its sister group (BS ≤ 80%, PP = 0.96). The second clade included all Linum species and segregated genera (BS ≤ 80%, PP = 1) resolved in two main clades (I and II).
Clade I (BS = 100%, PP = 1) recovered the section Dasylinum as monophyletic (clade A: BS = 100%, PP = 1) and almost all members of the section Linum, also as a natural group (clade B: BS = 100%, PP = 1), except for L. stelleroides Planch, which was positioned as the basal clade. Within clade B, the position of L. usitatissimum showed discrepancies between PA and BI analyses. In the topology obtained with PA, L. bienne Mill. was positioned as the sister species of L. usitatissimum, although with a low support value (BS < 80%), while in the BI analysis, the cultivated species was positioned as the sister of the clade formed by L. bienne and L. villarianum Pau., with strong support (PP = 1). Notwithstanding this inconsistency, this clade showed robust support in both analyses (BS = 100%, PP = 1). Additionally, this clade was consistently recovered as a sister of the clade composed of L. hologynum Rchb. and L. marginale A.Cunn. ex. Planch. (BS = 95%, PP = 1). Interestingly, in the sect. Linum (clade B), several species of socioeconomic interest were recovered. For example, in addition to agronomic species, those of ornamental interest, L. narbonense L., L. grandiflorum Desf., and L. decumbens Desf., formed a clade together but with no support (BS < 80%, PP < 0.9). For its part, L. perenne, also of ornamental importance, was recovered forming a natural group with Linum leonii F. W. Schultz, L. alpinum Jacq., L. tommasinii (Rchb.), and L. punctatum C. Presl., albeit without enough support (BS < 80%, PP < 0.9).
Clade II integrated most of the Linum species (31%), as well as the segregated genera. Here, two large well-resolved clades were recovered (C and D), with Radiola linoides positioned as their sister group with strong support values (BS = 98%, PP = 1). Clade C (BS = 87%, PP = 0.95) included three minor clades: the clade Hesperolinon (BS < 80%, PP = 1), which integrated the genera Hesperolinon and Sclerolinon; the clade Cliococca (BS ≤ 80%, PP = 0.99), which was recovered as the sister of clade Hesperolinon and included seven species of the so-called section Linopsis [(Linum oligophyllum Willd. Ex Schult., L. littorale A. St. Hil, L. macraei Benth., L. prostratum Dombey ex Lam., L. rupestre (A. Gray) Engelm. ex A. Gray, L. vernale Wooton, and L. kingii S. Watson) together with the monospecific genus Cliococca; and the clade Linopsis A (BS = 97%, PP = 1), which integrated more representatives of the sect. Linopsis.
Clade D (BS = 87%, PP = 1) recovered four minor clades: the section Syllinum with strong support values (BS = 100%, PP = 1) and sister to clade Linopsis B, integrating some species from the sect. Linopsis (BS = 100%, PP = 1). The clade composed of the monotypic section Cathartolinum, which was positioned as basal to the clades Linopsis B and Syllinum (BS = 84%, PP = 0.92), and the fourth clade integrating the remaining species of the sect. Linopsis (clade Linopsis C) had high support values (BS = 91%, PP = 1).
2.2. Reconstruction of Ancestral Flower Color
The results of the analysis in RASP based on the S-DIVA model (Figure 2) indicated that the flower color of the most recent common ancestor of all living linoids (41.44 Ma: 95% HPD 36.95–47.45 Ma; Supplementary Figure S4) was most likely yellow-white (AC) (node I, Table 1). The same state was recovered for node II (P = 0.30), composed of the genera Anisadenia, Tirpitzia, and Reinwardtia, and node III, which corresponds to the species of Linum plus segregated genera (=clade Linum s.l., P = 0.16), which diversified during the Eocene (Priabonian) 34.77 Ma (95% HPD: 20.96–46.61 Ma).
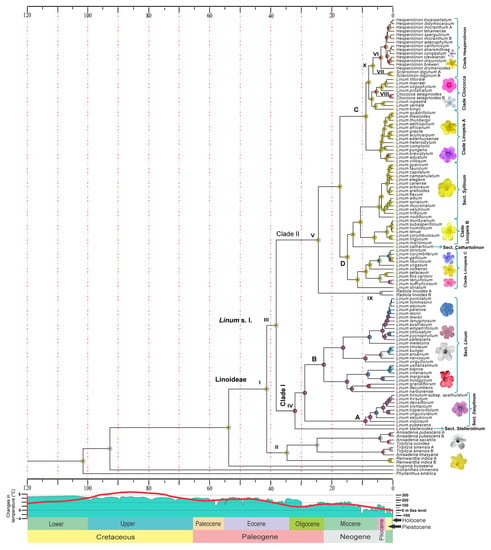
Figure 2.
Reconstruction of ancestral states of flower color in the subfamily Linoideae based on S-DIVA analysis. Colored circles at nodes indicate the most likely color state, as estimated by RASP. Roman numerals represent key nodes. The colors at the tips represent the current color of the flower. Global temperature scales and timing of sea-level fluctuations were modified from Zachos et al. [45] and Haq et al. [46], respectively. The geological time scale is in Ma (million years).

Table 1.
Divergence time estimates from BEAST analysis and probabilities of the ancestral color of the flower estimated with the Statistical Dispersal-Vicariance Analysis (S-DIVA) in RASP for key nodes of Linoideae in analysis of concatenated data. 95% HPD = 95% highest posterior density; Ma = million years; AC = area code; P = probability value.
Within the clade Linum s.l., node IV (=clade I) corresponds to sects. Dasylinum and Linum, which recovered purple as the most likely color ancestral state (P = 0.32), emerging in the Oligocene at approximately 32.16 Ma (95% HPD 26.33–37.7 Ma). In this same epoch, node V (=clade II, P = 0.62, Table 1), which includes sects. Linopsis, Cathartolinum, and Syllinum, as well as segregated genera, showed yellow-white flower color as the most likely ancestral state (Table 1, Figure 2).
Regarding the segregated genera, the clade Hesperolinon (node X) recovered yellow as the ancestral color of the flower during the Miocene (Tortonian, 7.95 Ma), which was also recovered for the genus Sclerolinon (node VII, P = 1). For the same period, the red color was detected as emerging in the clade of purple flowers in the mid-Miocene (13.5 Ma). The pink color emerged between the end-Miocene and Pleistocene (0.22–5.78 Ma) in the genus Hesperolinon (node VI), with the color yellow-pink as the ancestral state (P = 0.86), and in Cliococca (node VIII) with the color white-pink (P = 1). The above, unlike Radiola (node IX), showed white as the most likely state (P = 1).
The parsimony ancestral state reconstruction analysis (Supplementary Figure S5) resulted in flower color reconstructions like those obtained with the Bayesian approach (S-DIVA). Minor differences were detected at deep nodes. With parsimony analysis, nodes I, II, III, and V recovered the yellow color as the ancestral state. With RASP, these same nodes retrieved yellow-white as the most likely state. The rest of the nodes similarly recovered the ancestral state in both analyses. According to the parsimony analysis, the pink and white colors of the flowers showed multiple independent origins in subclade II (node V). In subclade I (node IV), the ancestral color was pink-purple.
It is important to note that each of the two large clades into Linum s.l. shows a color affinity. In clade I (node IV), corresponding to sections Dasylinum and Linum, the ancestors show mainly a purple flower color, followed by pink. Clade II (node V) shows a greater distribution of the yellow color among the ancestors and the current species, which is therefore a plesiomorphic state. The pink and blue colors indicate more recent states (~15 and 10 Ma, respectively). For its part, the red color, according to our analysis, is an apomorphic state also of recent origin, since it is present only in two current species, L. decumbens and L. grandiflorum, both forming a monophyletic group.
3. Discussion
3.1. Phylogenetic Analysis
Our results support the close relationship between Anisadenia and Tirpitzia, with Reinwatdtia as the sister group, in contrast with McDill and Simpson [1], who recovered Anisadenia and Reinwardtia as the most closely related genera. Similarly, Ruiz-Martín et al. [13] obtained different results. They recovered Tirpitzia and Reinwardtia as monophyletic, with Anisadenia as the sister genus. The inconsistency in the phylogenetic position of these three genera was pointed out by McDill et al. [2], who concluded that, although the relationship between these genera is uncertain, they are consistently sister to the flax clade.
Within the clade of Linum s.l., two large subclades were recovered, consistent with previous phylogenetic work [1,2,13,26,33]. The largest of these subclades integrated Hesperolinon, Sclerolinon, Clioccoca, and Radiola. These genera were originally circumscribed within Linum [18,19,23] and, later, due to differences in morphological characters, segregated and categorized into the taxonomic rank of the genus. Considering the current circumscription, Linum is paraphyletic, which has already been widely highlighted in various phylogenetic studies [1,2,13,26]. However, our results add to those cited above that do not support the current circumscription of the genera, since all of them are nested within the different clades of flax with high support values. Therefore, it has been proposed to reconsider the return of these genera to Linum. If so, the genus will be recovered as a natural group.
Thus, Hesperolinon is consistently recovered as a monophyletic group nested within one of the large subclades of Linum, coinciding with previous works [1,2,13,26,33]. This contrasts significantly with the morphological evidence shown in a recent taxonomic study [25]. In addition, Hesperolinon has been highlighted as the only ecological and evolutionarily notable group showing extreme diversification in serpentine soils [1,19,47,48,49]. Geographically, the species of the genus are distributed in a narrow region within the Californian biogeographic province, which has demonstrated not only geographic but also edaphic endemism [4,19,25,26]. Considering these remarkable group differences, the results reported here, and Gray’s [24] proposal, we hypothesize that the Hesperolinon species should be treated at the section rank, probably together with Sclerolinon, which form a well-supported natural group. This latter genus, originally described as Linum digynum, shows important morphological similarities to Hesperolinon, which caused it to be transferred to that genus by Sharsmith [19]. However, it was finally elevated to generic rank by Rogers [20] based on the presence of a bicarpellate fruit 4-locular due to the presence of false septa, nuclei with one seed each. Since then, there has been no revision of Sclerolinon to give certainty that it is a monotypic genus.
Similarly, we suggest that Cliococca, which was initially described as Linum selaginoides Lam. until Babington [23] and Rogers and Mildner [21] validated its segregation based on a few morphological characters (presence of decumbent, leafy stems arising from an extensive subterranean pattern, and the presence of indehiscent capsules), should not be considered in the rank of genus. Based on our results, Cliococca selaginoides is more closely related to L. oligophyllum, L. littorale, L. macraei, and L. prostratum, with high support values. This is also consistent with the different phylogenies of the group [2,13,33]. Returning to the notes made by Rogers and Mildner [21] in their reevaluation of the genus Clioccoca, they mentioned that there are sufficient similarities to Linum; however, some characters, such as the indehiscent 10-segmented fruit, imbricate corolla, and the unique pollen morphology, suggest that C. selaginoides be maintained as a distinct genus from the flaxes. Under these considerations, future work should focus on evaluating the species at the population level and consider the possible hypothesis that it may be a hybrid, even more so when its original description was based on cultivated material from the Cambridge Botanic Garden. In addition, our results also do not support the current Radiola circumscription, despite its segregation from Linum based on morphological characters [3]. Most likely, it is a section.
Our results support the sectional division of four of the five sections proposed by Ockendon and Walters [31]. These are Dasylinum, Cathartolinum, and Syllinum, and without considering L. stelleroides, the sect. Linum was also recovered as monophyletic with high support values. Considering our results, we support Yuzepchuk’s [50] proposal to recognize L. stelleroides as a monotypic section called Stellerolinon Juz. ex Prob. The same has already been considered in recent works [4,33,51]. This is based not only on the fact that the species has a geographical distribution range that is different from the rest (Eastern Asia) but also on the presence of stipulate glands provided with a small stipe, a character that does not appear in any other section. Furthermore, the chromosome number of L. stelleroides is 2n = 20, different from sects. Linum and Dasylinum, which have the chromosomal base n = 8, 9, or 15 [31,50,52,53]. Meanwhile, all species of the sect. Linum lack stipulate glands [2]. For its part, Linopsis, the largest of all of the sections, was not recovered as a natural group, and its species were distributed in three clades. According to Planchon [15,16], Winkler [30], Rogers [54], and Ruiz-Martín et al. [13], the characters that describe Linopsis are very broad and variable, so a more detailed taxonomic treatment for this section, as well as the inclusion of ecological and biogeographical features, could support the proposal to divide Linopsis into independent sections.
Several recent works have shown the importance of identifying and studying the wild relatives of cultivated plants [55,56,57,58]. As sources of new genetic diversity, crop wild relatives have been used for many decades for plant breeding, contributing to a wide range of beneficial agronomic and nutritional traits [55,59]. Linum ussitatisimum is not an exception, as it is the species of the greatest importance and was used in ancient times for agronomical purposes in the subfamily, and, in recent years, its production demand has increased [60,61]. Hence, knowing the phylogenetic relationships of this species is essential to explore and estimate the potential use of available resources from its sister species.
Although there is no clarity about the sister group of the cultivated species, our results show a close relationship with L. villarianun and L. bienne. The relationship of cultivated flax with L. bienne was mentioned by McDill et al. [2], McDill and Simpson [1], Schneider et al. [26], Ruiz-Martín et al. [13], Sheidai et al. [62], and, recently, Bolsheva et al. [33], and it has been widely studied since it is considered the old flax wild form cultivated and the wild ancestor of the modern cultivated flax [2,12,63,64]. Something important is that, except for the work by Ruiz-Martín et al. [13], no studies have included L. villarianum in their analyses. The phylogenetic closeness of this last species with L. ussitatisimum represents an opportunity to focus efforts on studying it under the premise that it represents an important potential resource. However, this analysis only includes 54% of the species of the subfamily and 73% of the sect. Linum. Therefore, it is likely that by including the remaining species, the phylogenetic relationships of L. usitatissimum will change. As shown in this analysis, just by including one more species, the relationships with cultivated flax are uncertain and weak. It is urgent and essential to represent the remaining 46% of Linoideae. Including the rest of the taxa will surely shed light on this controversy, and the hypotheses proposed here will be tested.
Although they attract less attention, the rest of the species that have gained interest for their uses should not be left out. Among those, L. grandiflorum and L. narbonense stand out, reported as ornamentals [8], and our results recovered them forming a clade together with L. decumbens. These three species are the sister group of the clade that integrates cultivated flax. Linum perenne and L. lewisii, also reported as ornamentals [65,66,67], were recovered with a close relationship and as sister species to L. punctatum, L. tommasinii, L. alpinum, and L. leonii. All of them belong to the sect. Linum. The present work is the first to incorporate L. narbonense into a phylogenetic analysis, whose position had not been evaluated before and which has also been reported for medicinal use [68]. Other species of the genus have been recognized for their traditional uses. Linum rupestre, for example, is used in some localities in the state of Chiapas, Mexico, as a medicinal plant [11]. This species was recovered in the present analysis in a monophyletic group with L. vernale and L. kingii. Despite the importance of these and other flax species in medicine, phytochemistry, and ornamentals, several of them have not been a focus of interest, and little or nothing has been explored, not only at the molecular level but also in terms of their potential for use.
3.2. Reconstruction of Ancestral Flower Color
The diversity of the colors of flowers is one of the most striking characteristics of the radiation of angiosperms since this character has allowed us to identify and measure the interaction with animals [34,35,36,37]. The subfamily Linoideae has been characterized by the inclusion of a great diversity of colors in the corolla, which has translated into the ornamental interest in several of its species [5,6,14,25,54,69]. Our study provides the first tentative evidence that the color of the ancestral flowers of Linoideae was most likely yellow-white. Flower color transitions in Linoideae have not received much attention from an evolutionary perspective either. McDill et al. [2] were the first to formally recognize a clade of blue flowers and another clade of yellow flowers in Linum s.l., data that were perpetuated in the subsequent publications of the group. However, this identification was only based on the flower colors of the current species, without delving further into its evolutionary significance.
Based on our analyses of the reconstruction of ancestral states, yellow-white was the plesiomorphic state (41.44 Ma: 95% HPD 36.95–47.45 Ma), from which the purple flower evolved in the Oligocene at 32.16 Ma (clade IV: sects. Linum, Dasyllinum, and Stellerolinon), followed by blue and red colors in the late Miocene (~13 Ma), with pink as the most recent color to evolve (end-Miocene–Pleistocene), coinciding with the period of the greatest radiation in the subfamily. Previous studies have implicated flower color shifts in speciation [70,71]. To the extent that flower color plays a role in speciation events, it is important to determine what evolutionary forces underlie its divergence.
The emergence of the purple color (node IV) in one of the major clades in Linum s.l. in the late Eocene and Oligocene coincided with a period of low temperatures, which, in the Northern Hemisphere, became too cold [48,72]. According to the ancestral areas reconstructed by Maguilla et al. [32], in this period, the most recent common ancestor to Linum s.l. inhabited the Western Palearctic (i.e., Europe, North Africa, northern and central Arabian Peninsula, and part of temperate Asia). This cold event in the north probably caused the expansion of the lineages, since this climatic change promoted the establishment of communities dominated by temperate vegetation, mainly herbaceous species [73]. In addition, this could have favored the establishment of flax species, since they have shown a preference for habitats dominated by grasses and small herbaceous plants [1,2,13].
The Miocene was characterized by greater aridity that allowed the further expansion of sclerophyll shrublands and woodlands [48,74,75]. In this geological time, the red color evolved from a purple ancestor between the mid-Miocene (~13.5 Ma) and late Miocene (~7 Ma). In that same epoch, the blue color also emerged from different ancestors throughout clade I. At this point, the most likely ancestral area of the lineages was the Western Palearctic, and currently, this clade is essentially Eurasian [32]. In the middle of the Miocene, the starting phase of global cooling and rapid aridification led to the expansion of grassland and xeric vegetation [75,76]. The impact of the dry climate had important effects not only on the Western Palearctic but also across the entire Northern Hemisphere [77]. These events led to important diversification events, supported by growing evidence for species in the western Mediterranean [78,79,80]. These changes were likely what caused the greatest radiation of flax species in the Mediterranean Basin, producing its current status as a Linoideae hotspot.
The effects were also significant in Africa, where the summer monsoon was drastically reduced by the narrowing of the Tethys Sea during the Tortonian age (11.6–7.2 Ma) [81]. These changes altered the composition and distribution of Northern Hemisphere flora [76,82] and coincided with the emergence of the two important lineages in clade C (American and African lineages). This cladogenetic event was congruent with several geological events. The rise of the African lineage (clade Linopsis A) [13,32] was concordant with the connection formed between Africa and south-western Asia due to the collision of the Afro-Arabian plate with the Iranian and Anatolian plates [83]. Furthermore, the main collision with Eurasia resulted in the closure of the Tethys Sea with the formation of the Gomphoterium land bridge during the mid-Miocene, causing African and Eurasian biota to interact [84]. On the other hand, the colonization of the American continent (American lineage: North American = clade Hesperolinon + South American = clade Clioccoca) was congruent with an existing connection between North America and Eurasia. It had been assumed that lineages with divergence times between Eurasia and America younger than the Eocene would not have passed across a North Atlantic land bridge (NALB) but rather across the Bering Strait [85]. However, a review of Neogene sedimentary rocks with plants from Iceland [86,87,88,89,90] revealed rich warm-temperate to temperate flora that lasted at least until 9–8 Ma. This was supported by various studies on temperate flora that corroborated a migration between America and Eurasia via the NALB during the Miocene and up to the Pliocene [86,88,90,91,92,93]. At the end of the Miocene and the middle Pliocene (7.95–3.75 Ma), the pink color arose in the North America and South America lineages independently, both evolving from ancestors with yellow flowers. It is important to highlight that the yellow and pink colors dominate the flowers (Figure 2) of the American lineages, and yellow dominate the African lineages [32]. Results can be influenced by the time of separation of both continents with their respective biogeographical events. These conditions probably favored the diversification and fixation of the pink color through the current species.
The pink color also emerged in the Eurasian species L. viscosum and L. pubescens in clade A in the late Miocene–Pliocene and more recently in the Asian genera Anisadenia and Tirpitzia during the Pliocene–Pleistocene (~2.5 Ma). This last epoch was characterized by glacial and interglacial cycles that resulted in the evolution and migration of many plant lineages, favoring their diversification to alternate environments [79]. It is currently known that the Mediterranean Basin served as a refuge for many species during the Tertiary and Quaternary glaciations and as a source for the subsequent colonization of adjacent areas as Asia regions [94]. However, although the geological and paleoclimatic events mentioned could suggest a close relationship between the biogeographical history of the group and the color of the flowers, they do not explain why the pleisiomorphic yellow state is mainly maintained throughout clade II and the purple state is maintained in clade I. Here, it is likely that pollinators played a more important role.
Flower color transitions usually accompany a shift in pollination mode [95]. Observations on pollination in Linum or sister genera are scarce, scattered in the literature, and/or have not been updated [96,97,98,99]. The little that is known about the group has been generalized from those studies, and it is mentioned that Linum flowers are typically pollinated by insects such as honeybees, bumblebees, flies, and butterflies [13,99]. It is known that, due to differences in preferences, different functional groups of pollinators may select different flower colors [95,100,101]. However, many floral radiations exhibit a remarkable variety of colors despite members sharing the same functional group of pollinators [102,103,104,105]. The above suggests that the pollinator shift model does not fully account for the diversity of colors across angiosperms [106]. In addition, it is important to mention that we examined the color evolution of the corolla based on human perception and not the ultraviolet light (UV) spectrum that pollinators can detect, mainly insects [107]. It has been suggested that the evolution of human-visible patterns is associated with the evolution of larger flowers, but the evolution of UV patterns is correlated with the evolution of smaller flowers [108]. Therefore, we suggest complementing this work from an ecological perspective, since little is known about the types of pollinators and details about pollination in Linoideae. Moreover, color vision can vary among insect species, so this information cannot be generalized [109].
A few studies have examined the tempo of discrete changes in flower color, such as gains or losses of pigmentation [110,111]. According to our results, the most likely scenario is that pigmentation was gained in a speciation event from an ancestral lineage of white-yellow flowers. According to Ng and Smith [112], the appearance of a gain of pigmentation mutant in an ancestral population could also lead to the emergence of a new lineage if this trait allows or even promotes dispersal to a new region. This is consistent with the geological period with the greatest diversification of Linoideae, which coincides with dispersal events to new geographical areas of several lineages, and provides an initial assessment of the possible role of flower color in dispersal to new habitats. Nonetheless, we cannot rule out a pollinator-mediated scenario where a sub-population disperses to a new region with a different pollinator fauna that selects for colored flowers [111,113]. Studies about pollinators are needed to test this hypothesis. Once this knowledge is acquired, it will be possible to know and understand how Linoideae’s pollinators, especially insects, perceive colors and thus understand plant-pollinator interactions more precisely in this group.
Our study provides a novel picture of the flower color of the most recent ancestor of all living Linoideae and the earliest steps of color polymorphisms. The ancestral flower color for Linoideae was yellow-white, with the purple color dominating clade I ancestors and the yellow color dominant throughout clade II, suggesting a scenario closely related to the biogeographical history of the group and its pollinators. However, new progress in reconstructing the evolutionary steps and integrating breakthroughs in evo-devo and ecological research is still necessary. Likewise, the taxonomic status of the segregated genera was explored, and as a result, we propose here that they be reconsidered so that they are returned to Linum and that the current sectional status is reevaluated. These results are a contribution toward an understanding of floral color pattern diversity and evolution, as well as the systematics of the subfamily Linoideae.
4. Material and Methods
4.1. Taxon Sampling
The taxonomic diversity of Linoideae was represented by 451 accessions of 113 species covering the eight genera of the subfamily (Supplementary Table S1). Hugonia busseana Engl. (subfamily Hugonioideae) was included as an outgroup along with Phyllanthus emblica L. and Ixonanthes chinensis (Hook. & Arn.) Champ., representing the two sister families of Linaceae: Phyllanthaceae and Ixonanthaceae [114]. Sequences from these taxa were retrieved from GenBank (Supplementary Table S1) and correspond to four DNA regions: the nuclear ribosomal DNA ITS1-5.8S-ITS2 (ITS) region, Maturase K (matK), NADH-dehydrogenase subunit F (ndhF), and the intergenic spacer between tRNALeu and tRNAPhe (trnL-trnF). Sequences were aligned using PhyDe software [115] with the Muscle algorithm [116], followed by a final adjustment by visual inspection.
4.2. Phylogenetic Analysis
Phylogenetic analyses were performed separately for ITS and cpDNA (matK, ndhF, and trnL-trnF) and in combination (ET = ITS + cpDNA). Analyses were performed with H. busseana (subfamily Hugonioideae) as the outgroup. The congruence of the phylogenetic signals from ITS and cpDNA was evaluated by visual comparison of their respective topologies. Furthermore, an incongruence length difference (ILD) test [117], implemented in PAUP v4.0a168 [118] as the partition-homogeneity test between the ITS and cpDNA datasets, was conducted. The partition homogeneity test revealed that partitions were homogeneous (P > 0.05). There were no strongly conflictive topologies found among molecular data partitions (Supplementary Figures S1 and S2). Therefore, we concatenated both datasets for further analyses and discussion.
Phylogenetic reconstruction was carried out using Parsimony Analysis (PA) and Bayesian Inference (IB) approaches. Of the regions recovered from GenBank, 5.98% were missing in some taxa; hence, they were coded as “missing data (?)”. PA was performed with the heuristic search implemented in TNT v1.5 [119] with 100 iterations with the TBR (Tree Bisection Reconnection) algorithm, retaining 100 trees per iteration. Gaps were recorded as missing. The shortest trees obtained were saved for calculating the strict consensus tree. Statistical branch support was determined by bootstrap (BS) analysis running 1000 sampling replicates with replacement and collapsing those clades with a value lower than 50% through the “Cutoff” option.
The IB analyses were carried out in MrBayes v3.2.7a [120] with the Markov Chain Monte Carlo (MCMC) technique. Model parameters were fixed according to the values obtained with jModeltest v2.1.10 [121] for each of the matrices and selected with Akaike’s criterion (AIC) [122]. The models used were TVM + G for cpDNA and GTR + I + G for the concatenated matrix and ITS dataset. Each MCMC analysis was run for 10 million generations with four MCMC chains—one cold and three heated—starting from different random points in the parameter space with a discarded burn-in of 25% and sampled every 1000th generation. The outgroup was never forced to be monophyletic during searches. Nodes with posterior probabilities (PP) > 60% were retained in the majority-rule consensus tree. Finally, the trees obtained were visualized and edited in FigTree v1.4 [123].
4.3. Reconstruction of Ancestral Flower Color
Reconstruction of ancestral states using model-based methods requires a phylogenetic tree with branch lengths proportional to time, i.e., a timeline. The precedent is to avoid bias by assuming a strict correlation between molecular and morphological evolutionary rates. Therefore, molecular dating analyses were conducted using BEAST v1.10.1 [124] with the individual datasets (Supplementary Material Table S2) and concatenated matrix under an uncorrelated lognormal relaxed-clock model. Based on the results of the study by Xi et al. [125], two secondary calibration points were selected. The first of them was used to calibrate the root node of Phyllanthaceae + [(Ixonanthaceae + Linaceae) under a normal distribution (mean = 102.5; SD = 4.03). The second was used to calibrate the stem node of Linaceae + Ixonanthaceae (mean = 90; SD = 8.65). A third calibration point from fossil pollen grain data unequivocally attributed to Linum from the late Eocene from the Ebro basin in northeastern Spain [126,127] was used to calibrate the minimum stem node divergence of this genus under a lognormal distribution (mean = 1; SD = 1; offset = 35.55).
The model parameter implemented for the molecular clock implemented was GTR + I + G for all datasets. This was the closest model to those calculated according to the AIC in jModelTest. Tree priors were modeled with a birth–death process, which models speciation and extinction patterns. Three independent MCMC analyses were run for ITS and ET, each with 50,000,000 generations. For the cpDNA, 120,000,000 generations were carried out in four independent analyses. In all cases, sampling was performed every 1000th generation. The convergence and stationarity of the estimated parameter values were assessed according to effective sample size (ESS > 200), traces, and Bayesian density plots using Tracer v1.7. (Germany) [128]. The log files were combined using LogCombiner. A maximum-clade-credibility (MCC) tree representing the maximum a posteriori topology, with mean divergence times and a posterior probability limit of 0.9, was calculated after the removal of 10% of trees as burn-in using TreeAnnotator v1.10.4. (New Zeland) [129]. Finally, the trees were visualized with FigTree v1.4. (UK) [123].
We recorded the flower colors of 112 species of Linoideae using herbarium data, systematic and taxonomic studies, regional floras, and a database [5,6,13,54,130,131,132,133]. We did not use any general family descriptions or make any assumptions that all species of a genus share the same character state. The Bayesian method of ancestral state reconstruction (GTR + I + G model) Statistical Dispersal-Vicariance Analysis (S-DIVA) [134], implemented in RASP v3.2.1 (China) (Reconstruct Ancestral State in Phylogenies) [135], was performed to reconstruct the ancestral flower color. Each terminal in the tree was coded for six color states divided into the following categories: yellow, blue, white, purple, red, and pink. Flower colors with the highest probability value are indicated by the colored circle at each node of the tree, and the probability values are given in Table 1. For comparison, the ancestral color of flowers was also reconstructed using parsimony, as implemented in Mesquite v2.75 (Canada) [136]. All characters were treated as unordered.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11121579/s1. Table S1: GenBank accessions numbers for ndhF, matK, trnL-F, and ITS; Table S2: Divergence time estimates from BEAST analysis for key nodes of the Linoideae subfamily based on partitioned data; Figure S1: Phylogenetic relationships of the Linoideae subfamily based on combined plastid DNA: (A) Parsimony tree. (B) Bayesian Inference tree; Figure S2: Phylogenetic relationships of the Linoideae subfamily based on ITS: (A) Parsimony tree. (B) Bayesian Inference tree; Figure S3: Parsimony analysis tree of Linoideae based on combined plastid and ITS dataset; Figure S4: Beast maximum clade credibility tree of Linoideae inferred from combined ITS, ndhF, matK, and trnL-F; Figure S5: Reconstruction of ancestral states of flower color in the subfamily Linoideae based on Most Parsimonious Reconstruction.
Author Contributions
Conceptualization, M.B.-H.; methodology, M.B.-H. and A.V.-H.; formal analysis, A.V.-H. and M.B.-H.; investigation, A.V.-H. and M.B.-H.; resources, A.V.-H., M.B.-H. and D.G.; data curation, A.V.-H.; writing—original draft preparation, A.V.-H., M.B-H. and D.G.; writing—review and editing, M.B.-H., A.V.-H. and D.G.; visualization, M.B.-H.; supervision, M.B.-H.; project administration, M.B.-H.; funding acquisition, A.V.-H. and M.B.-H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Council of Science and Technology, student grant A.V.-H.-CONACYT- 765451, and Colegio de Postgraduados M.B.-H.-CP-E01-55-02.
Institutional Review Board Statement
The study did not require ethical approval.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Monserrat Vázquez for her valuable comments on the manuscript and Sarahí Barrientos for support with editing the images. The authors express their gratitude to two anonymous reviewers for their valuable comments to improve this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McDill, J.; Simpson, B.B. Molecular phylogenetics of Linaceae with complete generic sampling and data from two plastid genes. Bot. J. Linn. Soc. 2011, 165, 64–83. [Google Scholar] [CrossRef]
- McDill, J.; Repplinger, M.; Simpson, B.B.; Kadereit, W.J. The phylogeny of Linum and Linaceae subfamily Linoideae, with implications for their systematics, biogeography, and evolution of heterostyly. Syst. Bot. 2009, 34, 386–405. [Google Scholar] [CrossRef]
- Dressler, S.M.; Repplinger, M.; Bayer, C. Linaceae. In Flowering Plants. Eudicots, 1st ed.; Kubitzki, K., Ed.; Springer: Heidelberg, Germany, 2014; Volume 10, pp. 237–246. [Google Scholar]
- Melnikova, N.V.; Kudryavtseva, A.V.; Zelenin, A.V.; Lakunina, V.A.; Yurkevich, O.Y.; Speranskaya, A.S.; Snezhkina, A.V. Retrotransposon-based molecular markers for analysis of genetic diversity within the genus. Linum. BioMed Res. Int. 2014, 2014, 231589. [Google Scholar] [CrossRef] [PubMed]
- Rzedowski, J.; Calderón de Rzedowski, G. Linaceae. In Flora del Bajío y de Regiones Adyacentes; Instituto de Ecología A.C.: Pátzcuaro, Mexico, 1992; Volume 6, pp. 1–22. ISSN 0188-5170. [Google Scholar]
- Rzedowski, J.; Calderón de Rzedowski, G. Fascículo 5. Linaceae. In Flora del Valle de Tehuacán-Cuicatlán, 1st ed.; Universidad Nacional Autónoma de México: Mexico City, México, 1994; Volume 5, pp. 5–19. [Google Scholar]
- Touré, A.; Xu, X.M. Flaxseed lignans: Source, biosynthesis, metabolism, antioxidant activity, bio-active components, and health benefits. Compr. Rev. Food Sci. Food Saf. 2010, 9, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Mridula, D.; Rehal, J.; Barnwal, P. Flaxseed: A potential source of food, feed and fiber. Crit. Rev. Food Sci. Nutr. 2011, 51, 210–222. [Google Scholar] [CrossRef]
- Lautié, E.; Quintero, R.; Fliniaux, M.A.; Villarreal, M.L. Selection methodology with scoring system: Application to Mexican plants producing podophyllotoxin related lignans. J. Ethnopharmacol. 2008, 120, 402–412. [Google Scholar] [CrossRef]
- Alonso-Castro, J.A.; Villarreal, M.L.; Sálazar-Olivo, A.L.; Gomez-Sanchez, M.; Dominguez, F.; Garcia-Carranca, A. Mexican medicinal plants used for cancer treatment: Pharmacological phytochemical and ethnobotanical studies. J. Ethnopharmacol. 2011, 133, 945–972. [Google Scholar] [CrossRef]
- Barrera-Robles, P.J.; Burgos-Hernández, M.; Ruíz-Acevedo, A.D.; Castillo-Campos, G. La familia Linaceae en México: Estado actual y perspectivas. Bot. Sci. 2020, 98, 560–572. [Google Scholar] [CrossRef]
- Fu, Y.-B.; Peterson, G.; Diederichsen, A.; Richards, K.W. RAPD analysis of genetic relationships of seven flax species in the genus Linum L. Genet. Resour. Crop Evol. 2002, 49, 253–259. [Google Scholar] [CrossRef]
- Ruiz-Martín, J.; Santos-Gally, R.; Escudero, M.; Midgley, J.J.; Pérez-Barrales, R.; Arroyo, J. Style polymorphism in Linum (Linaceae): A case of Mediterranean parallel evolution? J. Plant Biol. 2018, 20, 100–111. [Google Scholar] [CrossRef]
- Burgos-Hernández, M.; Castillo-Campos, G. Contribución al conocimiento del género Linum (Linaceae) en Veracruz, México. Acta Bot. Mex. 2019, 3, 126. [Google Scholar] [CrossRef]
- Planchon, J.E. Sur la famille des Linées. Lond. J. Bot. 1847, 6, 588–603. [Google Scholar]
- Planchon, J.E. Sur la famille des Linées. Lond. J. Bot. 1848, 7, 165–186, 473–501, 507–528. [Google Scholar]
- Hallier, H. Beitrëge zur Kenntnis der Linaceae (DC. 1819) Dumort. Beih. Bot. Cent. Abt. 1 1921, 39, 1–178. [Google Scholar]
- Small, J.K. Linaceae. N. Am. Flora 1907, 25, 67–87. [Google Scholar]
- Sharsmith, H.K. The genus Hesperolinon (Linaceae). Univ. Calif. Publ. Bot. 1961, 32, 235–314. [Google Scholar]
- Rogers, C.M. Sclerolinon, a new genus in the Linaceae. Madroño 1966, 18, 181–184. [Google Scholar]
- Rogers, C.M.; Mildner, R. The reevaluation of the genus Cliococca (Linaceae) of South America. Rhodora 1971, 73, 560–565. [Google Scholar]
- Lamarck, J.B. Encyclopédie Méthodique, Botanique 1, 1st ed.; Panckoucke: Paris, France, 1791; p. 496. [Google Scholar]
- Babington, C.C. Description of a new genus of Lineae. Trans. Linn. Soc. Lond. 1842, 19, 33–34. [Google Scholar] [CrossRef][Green Version]
- Gray, A. Linum sect. Hesperolinon. Proc. Am. Acad. Arts 1865, 6, 521. [Google Scholar]
- González-Velasco, J.; Burgos-Hernández, M.; Galván-Escobedo, I.G.; Castillo-Campos, G. Taxonomic update of the flax family in Mexico. Phytotaxa 2022, 549, 141–184. [Google Scholar] [CrossRef]
- Schneider, A.C.; Freyman, W.A.; Guilliams, C.M.; Springer, Y.P.; Baldwin, B.G. Pleistocene radiation of the serpentine—Adapted genus Hesperolinon and other divergence times in Linaceae (Malpighiales). Am. J. Bot. 2016, 103, 221–232. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brewer, W.H.; Watson, S.; Gray, A. Botany [of California], 1st ed.; Cambridge University Press: Cambridge, MA, USA, 1876. [Google Scholar]
- Trelease, W. A revision of North American Linaceae. Trans. Acad. Sci. St. Louis 1887, 5, 7–20. [Google Scholar]
- Trelease, W. Linaceae. In Synoptical Flora of North America, 1st ed.; American Book Company: New York, NY, USA, 1897; Volume 2, pp. 344–349. [Google Scholar]
- Winkler, H. Linaceae. In Die Natürlichen Pflanzenfamilien, 19th ed.; Engelmann: Leipzig, Germany, 1931; Volume 10, pp. 82–130. [Google Scholar]
- Ockendon, D.J.; Walters, S.M. Linum. In Flora Europea, 1st ed.; Cambridge University Press: Cambridge, MA, USA, 1968; pp. 206–211. [Google Scholar]
- Maguilla, E.; Escudero, M.; Ruiz-Martín, J.; Arroyo, J. Origin and diversification of flax and their relationship with heterostyly across the range. J. Biogeogr. 2021, 48, 1994–2007. [Google Scholar] [CrossRef]
- Bolsheva, N.L.; Melnikova, N.V.; Dvorianinova, E.M.; Mironova, L.N.; Yurkevich, O.Y.; Amosova, A.V.; Krasnov, G.S.; Dimitriev, A.A.; Muravenko, O.V. Clarification of the position of Linum stelleroides Planch. within the phylogeny of the genus Linum L. Plants. 2022, 11, 652. [Google Scholar] [CrossRef]
- Wesselingh, R.A.; Arnold, M.L. Pollinator behaviour and the evolution of Louisiana iris hybrid zones. J. Evol. Biol. 2000, 13, 171–180. [Google Scholar] [CrossRef]
- Martén-Rodríguez, S.; Fenster, C.B.; Agnarsson, I.; Skog, L.E.; Zimmer, E.A. Evolutionary breakdown of pollination specialization in a Caribbean plant radiation. New Phytol. 2010, 188, 403–441. [Google Scholar] [CrossRef]
- Lehrer, M.; Horridge, G.A.; Zhang, S.W.; Gadagkar, R. Shape vision in bees: Innate preference for flower-like patterns. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1995, 347, 123–137. [Google Scholar] [CrossRef]
- Koski, M.H.; Ashman, T.L. Dissecting pollinator responses to a ubiquitous ultraviolet floral pattern in the wild. Funct. Ecol. 2014, 28, 868–877. [Google Scholar] [CrossRef]
- Manning, A. The Effect of Honey-Guides. Behaviour 1956, 9, 114–139. [Google Scholar] [CrossRef]
- Hansen, D.M.; van der Niet, T.; Johnson, S.D. Floral signposts: Testing the significance of visual ‘nectar guides’ for pollinator behaviour and plant fitness. Proc. R. Soc. B 2012, 279, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Streisfeld, M.A.; Rausher, M.D. Altered trans-regulatory control of gene expression in multiple anthocyanin genes contributes to adaptive flower color evolution in Mimulus aurantiacus. Mol. Biol. Evol. 2009, 26, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D.; Rausher, M.D. Gene loss and parallel evolution contribute to specie difference in flower color. Mol. Biol. Evol. 2011, 28, 2799–2810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, C.; Yao, X.; Wang, F.; Wu, J.; King, G.J.; Liu, F. Disruption of a carotenoid cleavage dioxygenase 4 gene converts flower colour from white to yellow in Brassica species. New Phytol. 2015, 206, 1513–1526. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.A.R.; Fenster, C.B.; Hereford, J.; Huang, S.; Ree, R.H. Floral diversity a community structure in Pedicularis (Orobanchaceae). Ecology 2012, 93, 182–193. [Google Scholar] [CrossRef]
- Lawson, D.A.; Rands, S.A. The evolution of floral guides: Using a genetic algorithm to investigate the evolution of floral cue arrangements. Biol. J. Linn. Soc. Lond. 2018, 123, 739–753. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef]
- Haq, B.U.; Hardenbol, J.A.N.; Vail, P.R. Chronology of fluctuating sea levels since the Triassic. Science 1987, 235, 1156–1167. [Google Scholar] [CrossRef]
- Raven, P.H.; Axelrod, D.I. Origin and relationships of the California flora. Univ. Calif. Publ. Bot. 1978, 72, 1–134. [Google Scholar]
- Anacker, B.L.; Whittall, J.B.; Goldberg, E.E.; Harrison, S.P. Origins and consequences of serpentine endemism in the California flora. Evolution 2011, 65, 365–376. [Google Scholar] [CrossRef]
- Springer, Y.P. Do extreme environments provide a refuge from pathogens? A phylogenetic test using serpentine flax. Am. J. Bot. 2009, 96, 2010–2021. [Google Scholar] [CrossRef] [PubMed]
- Yuzepchuk, S.V. Genus Linum-Linaceae Dumort. In Flora SSSR (Flora of the Soviet Union); Shishkin, B.K., Bobrov, E.G., Eds.; Izdatel’stvo Akademii Nauk SSSR: Moscow, Russia, 1949; Volume 14, pp. 84–146. [Google Scholar]
- Bolsheva, N.L.; Melnikova, N.V.; Kirov, I.V.; Speranskaya, A.S.; Krinitsina, A.A.; Dmitriev, A.A.; Muravenko, O.V. Evolution of blue-flowered species of genus Linum based on high-throughput sequencing of ribosomal RNA genes. BMC Evol. Biol. 2017, 17, 253. [Google Scholar] [CrossRef] [PubMed]
- Martzenitzina, K.K. The chromosomes of some species of the genus Linum L. Bull. Appl. Bot. Genet. Plant Breed. 1927, 17, 25–264. [Google Scholar]
- Petrova, A.V. IOPB Chromosome number reports XXXV. Taxon 1927, 21, 161–166. [Google Scholar]
- Rogers, C.M. The systematics of Linum sect. Linopsis. Plant Syst. Evol. 1982, 140, 225–234. [Google Scholar] [CrossRef]
- Castañeda-Alvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Toll, J. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef] [PubMed]
- Von Wettberg, E.J.; Chang, P.L.; Başdemir, F.; Carrasquila-Garcia, N.; Korbu, L.B.; Moenga, S.M.; Bedada, G.; Greenlon, A.; Moriuchi, K.S.; Singh, V.; et al. Ecology and genomics of an important crop wild relative as a prelude to agricultural innovation. Nat. Commun. 2018, 9, 649. [Google Scholar] [CrossRef]
- Nair, K.P. Utilizing crop wild relatives to combat global warming. Adv. Agron. 2019, 153, 175–258. [Google Scholar] [CrossRef]
- Vincent, H.; Hole, D.; Maxted, N. Congruence between global crop wild relative hotspots and biodiversity hotspots. Biol. Conserv. 2022, 265, 109432. [Google Scholar] [CrossRef]
- Brozynska, M.; Furtado, A.; Henry, R.J. Genomics of crop wild relatives: Expanding the gene pool for crop improvement. Plant Biotechnol. J. 2016, 14, 1070–1085. [Google Scholar] [CrossRef]
- Eyres, L. Flaxseed fibre—A functional superfood? Food N. Z. 2015, 15, 24. [Google Scholar]
- Kaur, P.; Waghmare, R.; Kumar, V.; Rasane, P.; Kaur, S.; Gat, Y. Recent advances in utilization of flaxseed as potential source for value addition. OCL 2018, 25, A304. [Google Scholar] [CrossRef]
- Sheidai, M.; Darini, S.; Talebi, S.M.; Koohdar, F.; Ghasemzadeh-Baraki, S. Molecular systematic study in the genus Linum (Linaceae) in Iran. Acta Bot. Hung. 2019, 61, 421–434. [Google Scholar] [CrossRef]
- Tammes, T. The genetics of the genus Linum. Bibliogr Genet. 1928, 4, 1–36. [Google Scholar]
- Allaby, R.G.; Peterson, G.W.; Merriwether, A.; Fu, Y.B. Evidence of the domestication history of flax (Linum usitatissimum) from genetic diversity of the sad2 locus. Theor. Appl. Genet. 2005, 112, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Maxted, N.; Scholten, M.; Codd, R.; Ford-Lloyd, B. Creation and use of a national inventory of crop wild relatives. Biol. Conserv. 2007, 140, 142–159. [Google Scholar] [CrossRef]
- Cullis, J.O. Diagnosis and management of anaemia of chronic disease: Current status. Br. J. Haematol. 2011, 154, 289–300. [Google Scholar] [CrossRef]
- Tork, D.G.; Anderson, N.O.; Wyse, D.L.; Betts, K.J. Domestication of perennial flax using an ideotype approach for oilseed, cut flower, and garden performance. Agronomy 2019, 9, 707. [Google Scholar] [CrossRef]
- Mohammed, M.M.; Christensen, L.P.; Ibrahim, N.A.; Awad, N.E.; Zeid, I.F.; Pedersen, E.B.; Jensen, K.B.; Colla, P.L. Anti-HIV-1 activities of the extracts from the medicinal plant Linum grandiflorum Desf. In Proceedings of 4th Conference on Research and Development of Pharmaceutical Industries (Current Challenges). Med. Aromat. Plant Sci. Biotechnol. 2009, 3, 37–41. [Google Scholar]
- Rogers, C.M. Relationships of Hesperolinon and Linum (Linaceae). Madroño 1975, 23, 153–159. [Google Scholar]
- Bradshaw, H.D.; Wilbert, S.M.; Otto, K.G.; Schemske, D.W. Genetic mapping of floral traits associated with reproductive isolation in monkey flowers (Mimulus). Nature 1995, 376, 762–765. [Google Scholar] [CrossRef]
- Van der Niet, T.; Johnson, S.D. Phylogenetic evidence for pollinator driven diversification of angiosperms. Trends Ecol. Evol. 2012, 27, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.G.; Fairbanks, R.G.; Mountain, G.S. Tertiary oxygen isotope synthesis, sea level history and continental margin erosion. Paleoceanogr. Paleoclimatol. 1987, 2, 1–19. [Google Scholar] [CrossRef]
- Tiffney, B.H.; Manchester, S.R. The use of geological and paleontological evidence in evaluating plant phylogeographic hypotheses in the Northern Hemisphere Tertiary. Int. J. Plant Sci. 2001, 162, S3–S17. [Google Scholar] [CrossRef]
- Bush, M.; Flenley, J.; Gosling, W. Tropical Rainforest Responses to Climatic Change, 2nd ed.; Springer: Berlin, Germany, 2011; pp. 1–34, 85–123. [Google Scholar] [CrossRef]
- Herbert, T.D.; Lawrence, K.T.; Tzanova, A.; Peterson, L.C.; Caballero-Gill, R.; Kelly, C.S. Late Miocene global cooling and the rise of modern ecosystems. Nat. Geosci. 2016, 11, 843–847. [Google Scholar] [CrossRef]
- Milne, R.I.; Abbott, R.J. The origin and evolution of tertiary relict floras. Adv. Bot. Res. 2002, 38, 281–314. [Google Scholar] [CrossRef]
- Weijermars, R. Neogene tectonics in the Western Mediterranean may have caused the Messinian Salinity Crisis and an associated glacial event. Tectonophysics 1988, 148, 211–219. [Google Scholar] [CrossRef]
- Casimiro-Soriguer, R.; Talavera, M.; Balao, F.; Terrab, A.; Herrera, J.; Talavera, S. Phylogeny and genetic structure of Erophaca (Leguminosae), a East-West Mediterranean disjunct genus from the Tertiary. Mol. Phylogenet. Evol. 2010, 56, 441–450. [Google Scholar] [CrossRef]
- Fernández-Mazuecos, M.; Jiménez-Mejías, P.; Rotllan-Puig, X.; Vargas, P. Perspectives in plant ecology, evolution and systematics narrow endemics to Mediterranean islands: Moderate genetic diversity but narrow climatic niche of the ancient, critically endangered Naufraga (Apiaceae). Perspect. Plant Ecol. Evol. Syst. 2014, 16, 190–202. [Google Scholar] [CrossRef]
- García-Castaño, J.L.; Terrab, A.; Ortiz, M.A.; Stuessy, T.F.; Talavera, S. Patterns of phylogeography and vicariance of Chamaerops humilis L. (Palmae). Turk. J. Bot. 2014, 38, 1132–1146. [Google Scholar] [CrossRef]
- Zhang, Z.; Ramstein, G.; Schuster, M.; Li, C.; Contoux, C.; Yan, Q. Aridification of the Sahara Desert caused by Tethys Sea shrinkage during the Late Miocene. Nature 2014, 513, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Medail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Popov, S.; Rögl, F.; Rozanov, A.; Steininger, F.F.; Shcherba, I.; Kovac, M. Lithological-paleogeographic maps of Paratethys. Late Eocene to Pliocene. Cour. Forschungsinst. Senckenberg 2004, 250, 1–46. [Google Scholar]
- Rögl, F. Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geol. Carpath. 1999, 50, 339–349. [Google Scholar]
- Donoghue, M.J.; Smith, S.A. Patterns in the assembly of temperate forests around the Northern Hemisphere. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 1633–1644. [Google Scholar] [CrossRef]
- Grímsson, F.; Denk, T. Fagus from the Miocene of Iceland: Systematics and biogeographical considerations. Rev. Palaeobot. Palynol. 2005, 134, 27–54. [Google Scholar] [CrossRef]
- Grímsson, F.; Denk, T. Floristic turnover in Iceland from 15 to 6 Ma extracting biogeographical signals from fossil floral assemblages. J. Biogeogr. 2007, 34, 1490–1504. [Google Scholar] [CrossRef]
- Grímsson, F.; Denk, T.; Símonarson, L.A. Middle Miocene floras of Iceland—The early colonization of an island? Rev. Palaeobot. Palynol. 2007, 144, 181–219. [Google Scholar] [CrossRef]
- Grímsson, F.; Denk, T.; Zetter, R. Pollen, fruits, and leaves of Tetracentron (Trochodendraceae) from the Cainozoic of Iceland and western North America and their palaeobiogeographic implications. Grana 2008, 47, 1–14. [Google Scholar] [CrossRef]
- Denk, T.; Grimsson, F.; Zetter, R.; Símonarson, L. Late Cainozoic Floras of Iceland: 15 Million Years of Vegetation and Climate History in the Northern North Atlantic; Springer Science & Business Media: New York, NY, USA, 2011; Volume 35. [Google Scholar] [CrossRef]
- Akhmetiev, M.A.; Bratzeva, G.M.; Giterman, R.E.; Golubeva, L.V.; Moiseyeva, A.I. Late Cainozoic Stratigraphy and Flora of Iceland; Academy of Sciences of the USSR: Moscow, Russia, 1978. [Google Scholar]
- Denk, T.; Grímsson, F.; KvaČek, Z. The Miocene floras of Iceland and their significance for late Cainozoic North Atlantic biogeography. Bot. J. Linn. Soc. 2005, 149, 369–417. [Google Scholar] [CrossRef]
- Milne, R.I. Phylogeny and biogeography of Rhododendron subsection Pontica, a group with a tertiary relict distribution. Mol. Phylogenet. Evol. 2004, 33, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D. Plant Evolution in the Mediterranean: Insights for Conservation, 2nd ed.; Oxford University Press: New York, NY, USA, 2020; pp. 10–20. [Google Scholar]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Williams, I.H.; Martin, A.P.; Clark, S.J. Pollination requirements of linseed (Linum usitatissimum). J. Agric. Sci. 1990, 115, 347–352. [Google Scholar] [CrossRef]
- Kearnsaf1, C.A.; Inouye, D.W. Fly pollination of Linum lewish (Linaceae). Am. J. Bot. 1994, 81, 1091–1095. [Google Scholar] [CrossRef]
- Gürbüz, B. Determination of cross-pollination in flax (Linum usitatissimum) using different experimental designs. J. Agric. Sci. 1999, 133, 31–35. [Google Scholar] [CrossRef]
- Lebel, M.; Obolski, U.; Hadany, L.; Sapir, Y. Pollinator-mediated selection on floral size and tube color in Linum pubescens: Can differential behavior and preference in different times of the day maintain dimorphism? Ecol. Evol. 2018, 8, 1096–1106. [Google Scholar] [CrossRef]
- Whittall, J.B.; Hodges, S.A. Pollinator shifts drive increasingly long nectar spurs in columbine flowers. Nature 2007, 447, 706–709. [Google Scholar] [CrossRef]
- Campbell, D.R.; Bischoff, M.; Lord, J.M.; Robertson, A.W. Flower color influences insect visitation in alpine New Zealand. Ecology 2010, 91, 2638–2649. [Google Scholar] [CrossRef]
- Armbruster, W.S. Can indirect selection and genetic context contribute to trait diversification? A transition-probability study of blossom-colour evolution in two genera. J. Evol. Biol. 2002, 15, 468–486. [Google Scholar] [CrossRef]
- Cooley, A.M.; Carvallo, G.; Willis, J.H. Is floral diversification associated with pollinator divergence? Flower shape, flower colour and pollinator preference in Chilean Mimulus. Ann. Bot. 2008, 101, 641–650. [Google Scholar] [CrossRef]
- Smith, C.I.; Godsoe, W.K.; Tank, S.; Yoder, J.B.; Pellmyr, O. Distinguishing coevolution from covicariance in an obligate pollination mutualism: Asynchronous divergence in Joshua tree and its pollinators. Evolution 2008, 62, 2676–2687. [Google Scholar] [CrossRef] [PubMed]
- Paget-Seekins, J. Ribes (Grossulariaceae) Pollination in Northern California: Strong Overlap in Visitor Assemblages Despite Floral Diversity. Master’s Thesis, The Faculty of Humboldt State University, Arcata, CA, USA, 2012. [Google Scholar]
- Muchhala, N.; Johnsen, S.; Smith, S.D. Competition for hummingbird pollination shapes flower color variation in Andean Solanaceae. Evolution 2014, 68, 2275–2286. [Google Scholar] [CrossRef] [PubMed]
- Song, B.M.; Lee, C.H. Toward a mechanistic understanding of color vision in insects. Front. Neural Circuits 2018, 12, 16. [Google Scholar] [CrossRef]
- Koski, M.H. Macroevolution of flower color patterning: Biased transition rates and correlated evolution with flower size. Front. Plant Sci. 2020, 945. [Google Scholar] [CrossRef]
- Van der Kooi, C.J.; Stavenga, D.G.; Arikawa, K.; Belušič, G.; Kelber, A. Evolution of insect color vision: From spectral sensitivity to visual ecology. Annu. Rev. Entomol. 2021, 66, 435–461. [Google Scholar] [CrossRef]
- Smith, S.D.; Miller, R.E.; Otto, S.P.; FitzJohn, R.G.; Rausher, M.D. The effects of flower color transitions on diversification rates in morning glories (Ipomoea subg. Quamoclit, Convolvulaceae). In Darwin’s Heritage Today, Proceedings of the Darwin 200 Beijing International Conference, Beijing, China, 24–26 October 2009; Higher Education Press: Beijing, China, 2010; pp. 202–226. [Google Scholar]
- Sobel, J.M.; Streisfeld, M.A. Flower color as a model system for studies of plant evo-devo. Front. Plant Sci. 2013, 4, 321. [Google Scholar] [CrossRef]
- Ng, J.; Smith, S.D. How traits shape trees: New approaches for detecting character state-dependent lineage diversification. J. Evol. Biol. 2014, 27, 2035–2045. [Google Scholar] [CrossRef]
- Waser, N.M.; Campbell, D.R. Ecological speciation in flowering plants. In Adaptive Speciation, 1st ed.; Cambridge University Press: Cambridge, UK, 2004; pp. 264–277. [Google Scholar]
- The Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Müller, J.; Müller, K.; Neinhuis, C.; Quandt, D. PhyDE-Phylogenetic Data Editor. 2005. Available online: http://www.phyde.de (accessed on 22 January 2021).
- Edgar, R.C. MUSCLE: Multiple sequence aligment with high accurancy and high troughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Farris, J.S.; Kallersjo, M.; Kluge, A.G.; Bult, C. Constructing a significance test for incongruence. Syst. Biol. 1995, 44, 570–572. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP* Phylogenetic Analysis Using Parsimony Version 4.0a168. Available online: http://paup.sc.fsu.edu (accessed on 25 June 2021).
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, A.S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MRBAYES 3.2: Efficient Bayesian phylogenetic inference and model selection across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 3 May 2021).
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef]
- Xi, Z.; Ruhfel, B.R.; Schaefer, H.; Amorim, A.M.; Sugumaran, M.; Wurdack, K.J.; Endress, P.K.; Matthews, M.L.; Stevens, P.F.; Mathews, S.; et al. Phylogenomics and a posteriori data partitioning resolve the Cretaceous angiosperm radiation Malpighiales. Proc. Natl. Acad. Sci. USA 2012, 109, 17519–17524. [Google Scholar] [CrossRef]
- Punt, W.; Den Breejen, P. Linaceae. Rev. Palaeobot. Palynol. 1981, 33, 75–115. [Google Scholar] [CrossRef]
- Cavagnetto, C.; Anadón, P. Preliminary palynological data on floristic and climatic changes during the Middle Eocene-Early Oligocene of the eastern Ebro Basin, northeast Spain. Rev. Palaeobot. Palynol. 1996, 92, 281–305. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Virus Evol. 2018, 67, 901. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A. TreeAnnotator v.2.4.3. 2016. Available online: http://beast.community/treeannotator (accessed on 12 May 2021).
- Rogers, C.M. Yellow flowered species of Linum in Eastern North America. Brittonia 1963, 15, 97–122. [Google Scholar] [CrossRef]
- Rogers, C.M. Yellow-flowered Linum (Linaceae) in Texas. Sida 1964, 1, 328–336. [Google Scholar]
- Rogers, C.M. Yellow-flowered species of Linum in Central America and western North America. Brittonia 1968, 20, 107–135. [Google Scholar] [CrossRef]
- POWO, Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 1 May 2022).
- Yan, Y.; Harris, A.J.; He, X. S-DIVA (Statistical Dispersal-Vicariance Analysis): A tool for inferring biogeographic histories. Mol. Phylogenet. Evol. 2010, 56, 848–850. [Google Scholar] [CrossRef]
- Yu, Y.; Blair, C.; He, X.J. RASP 4: Ancestral State Reconstruction Tool for Multiple Genes and Characters. Mol. Biol. Evol. 2020, 37, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 2.75. 2011. Available online: http://mesquiteproject.org (accessed on 22 January 2022).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).