
Does Abiotic Host Stress Favour Dothideomycete-Induced Disease Development?


Abstract
:1. Introduction
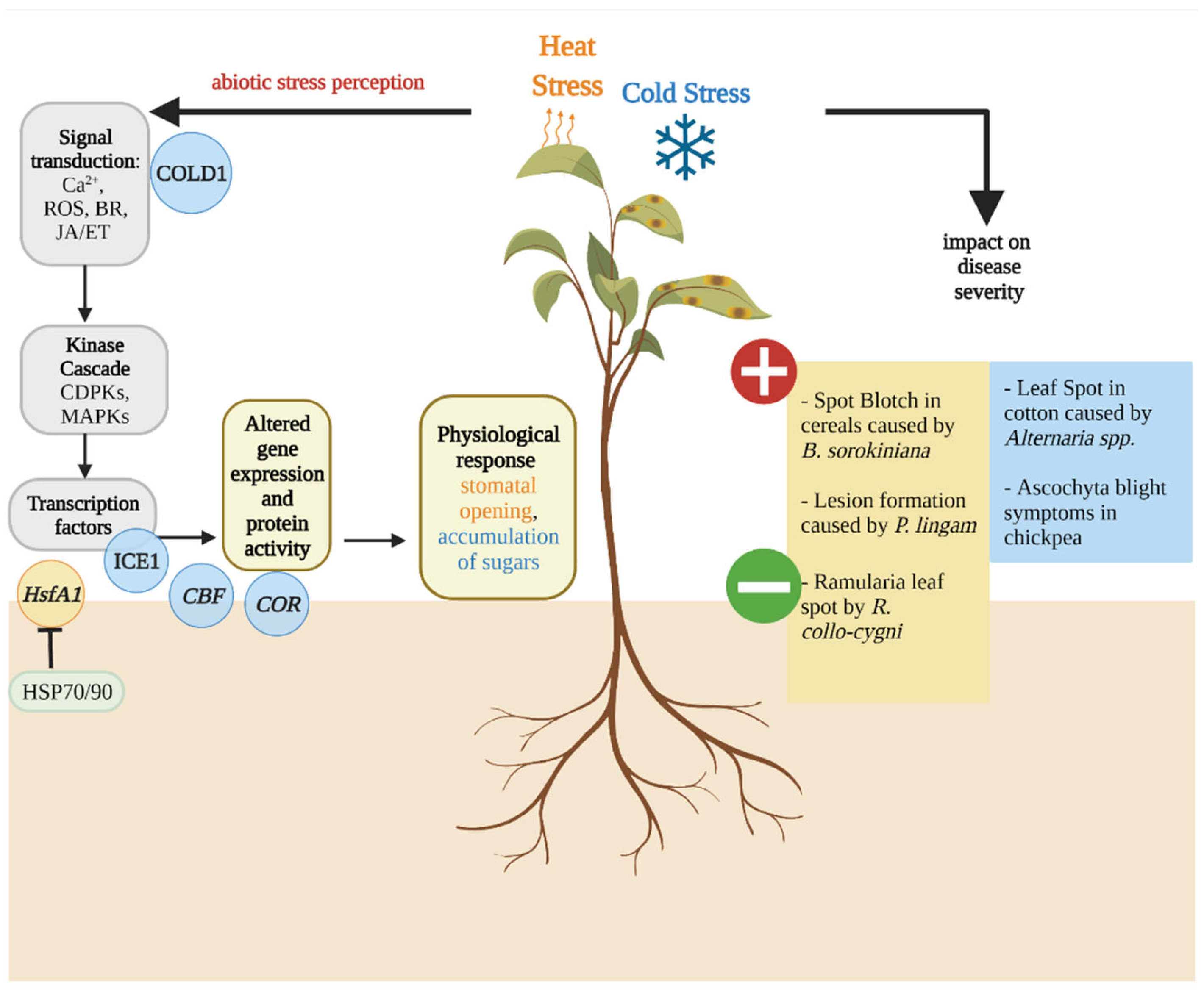
2. Overview of the Abiotic Stress Response in Plants
3. Temperature Stress Affects Dothideomycete-Induced Disease Development
3.1. Effect of Host Heat Stress on Dothideomycete-Induced Diseases
3.2. Effect of Host Cold Stress on Dothideomycete-Induced Diseases
4. Water Stress in the Host Affects Pathogenic Dothideomycete-Plant Interactions
4.1. Effect of Waterlogging and Flooding Stress in the Host on Dothideomycete-Induced Disease Development
4.2. Drought Stress Affects the Development of Dothideomycete-Induced Diseases
5. Salt Stress Influences Dothideomycete-Induced Diseae
6. Light Stress Affects Dothideomycete-Induced Disease Development
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Plant pathogen infection risk tracks global crop yields under climate change. Nat. Clim. Chang. 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Pereira, A. Plant abiotic stress challenges from the changing environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic stress in crop species: Improving tolerance by applying plant metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Atafar, Z.; Mesdaghinia, A.; Nouri, J.; Homaee, M.; Yunesian, M.; Ahmadimoghaddam, M.; Mahvi, A.H. Effect of fertilizer application on soil heavy metal concentration. Environ. Monit. Assess. 2010, 160, 83–89. [Google Scholar] [CrossRef]
- Manzoor, J.; Sharma, M.; Wani, K.A. Heavy metals in vegetables and their impact on the nutrient quality of vegetables: A review. J. Plant Nutr. 2018, 41, 1744–1763. [Google Scholar] [CrossRef]
- Snowdon, R.J.; Wittkop, B.; Chen, T.W.; Stahl, A. Crop adaptation to climate change as a consequence of long-term breeding. Theor. Appl. Genet. 2021, 134, 1613–1623. [Google Scholar] [CrossRef]
- Redman, R.S.; Kim, Y.O.; Woodward, C.J.D.A.; Greer, C.; Espino, L.; Doty, S.L.; Rodriguez, R.J. Increased fitness of rice plants to abiotic stress via habitat adapted symbiosis: A strategy for mitigating impacts of climate change. PLoS ONE 2011, 6, e14823. [Google Scholar] [CrossRef]
- Lohani, N.; Jain, D.; Singh, M.B.; Bhalla, P.L. Engineering Multiple Abiotic Stress Tolerance in Canola, Brassica napus. Front. Plant Sci. 2020, 11, 3. [Google Scholar] [CrossRef]
- Chojak-Koźniewska, J.; Kuźniak, E.; Zimny, J. The effects of combined abiotic and pathogen stress in plants: Insights from salinity and Pseudomonas syringae pv lachrymans interaction in cucumber. Front. Plant Sci. 2018, 9, 1691. [Google Scholar] [CrossRef]
- Vaughan, M.M.; Huffaker, A.; Schmelz, E.A.; Dafoe, N.J.; Christensen, S.A.; McAuslane, H.J.; Alborn, H.T.; Allen, L.H.; Teal, P.E.A. Interactive effects of elevated [CO2] and drought on the maize phytochemical defense response against mycotoxigenic Fusarium verticillioides. PLoS ONE 2016, 11, e0159270. [Google Scholar] [CrossRef] [Green Version]
- Sewelam, N.; El-Shetehy, M.; Mauch, F.; Maurino, V.G. Combined abiotic stresses repress defense and cell wall metabolic genes and render plants more susceptible to pathogen infection. Plants 2021, 10, 1946. [Google Scholar] [CrossRef]
- Creelman, R.A.; Mullet, J.E. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. USA 1995, 92, 4114–4119. [Google Scholar] [CrossRef] [Green Version]
- Horváth, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Huot, B.; Castroverde, C.D.M.; Velásquez, A.C.; Hubbard, E.; Pulman, J.A.; Yao, J.; Childs, K.L.; Tsuda, K.; Montgomery, B.L.; He, S.Y. Dual impact of elevated temperature on plant defence and bacterial virulence in Arabidopsis. Nat. Commun. 2017, 8, 1808. [Google Scholar] [CrossRef]
- Vaughan, M.M.; Huffaker, A.; Schmelz, E.A.; Dafoe, N.J.; Christensen, S.; Sims, J.; Martins, V.F.; Swerbilow, J.; Romero, M.; Alborn, H.T.; et al. Effects of elevated [CO2] on maize defence against mycotoxigenic Fusarium verticillioides. Plant Cell Environ. 2014, 37, 2691–2706. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594. [Google Scholar] [CrossRef] [Green Version]
- Tiley, A.M.; Karki, S.J.; Feechan, A. Zymoseptoria Tritici. In eLS; John Wiley & Sons, Ltd: Chichester, UK, 2018; pp. 1–8. [Google Scholar] [CrossRef]
- See, P.T.; Schultz, N.; Moffat, C.S. Evaluation of Pyrenophora tritici-repentis infection of wheat heads. Agriculture 2020, 10, 417. [Google Scholar] [CrossRef]
- Churchill, A.C.L. Mycosphaerella fijiensis, the black leaf streak pathogen of banana: Progress towards understanding pathogen biology and detection, disease development, and the challenges of control. Mol. Plant Pathol. 2011, 12, 307–328. [Google Scholar] [CrossRef]
- Noar, R.D.; Thomas, E.; Daub, M.E. A novel polyketide synthase gene cluster in the plant pathogenic fungus Pseudocercospora fijiensis. PLoS ONE 2019, 14, e0212229. [Google Scholar] [CrossRef] [Green Version]
- Westman, S.M.; Kloth, K.J.; Hanson, J.; Ohlsson, A.B.; Albrectsen, B.R. Defence priming in Arabidopsis—A Meta-Analysis. Sci. Rep. 2019, 9, 13309. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3544. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Sonnewald, U. Differences and commonalities of plant responses to single and combined stresses. Plant J. 2017, 90, 839–855. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Luan, S. The CBL-CIPK network in plant calcium signaling. Trends Plant Sci. 2009, 14, 37–42. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef]
- Fichman, Y.; Mittler, R. Rapid systemic signaling during abiotic and biotic stresses: Is the ROS wave master of all trades? Plant J. 2020, 102, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.H.; Willmann, M.R.; Chen, H.C.; Sheen, J. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiol. 2002, 129, 469–485. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zhou, J.M. Receptor-Like Cytoplasmic Kinases: Central Players in Plant Receptor Kinase-Mediated Signaling. Annu. Rev. Plant Biol. 2018, 69, 267–299. [Google Scholar] [CrossRef] [Green Version]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for Plant Growth and Mitigation of Abiotic Stresses: A Metabolomics Perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef]
- Sandrini, M.; Nerva, L.; Sillo, F.; Balestrini, R.; Chitarra, W.; Zampieri, E. Abiotic Stress and Belowground Microbiome: The Potential of Omics Approaches. Int. J. Mol. Sci. 2022, 23, 1091. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Santner, A.; Calderon-Villalobos, L.I.A.; Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Arbona, V.; Manzi, M.; Zandalinas, S.I.; Vives-Peris, V.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Physiological, metabolic, and molecular responses of plants to abiotic stress. In Stress Signaling in Plants: Genomics and Proteomics Perspective; Springer: Berlin/Heidelberg, Germany, 2016; Volume 2, pp. 1–35. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.A.D.; Suzuki, N.; Ciftci-Yilmaz, S.U.L.T.A.N.; Mittler, R.O.N. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant. Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, S.; Nagarajan, S. Abiotic tolerance and crop improvement. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic Foundation; Springer: Dordrecht, The Netherlands, 2010; pp. 1–11. ISBN 9789048131112. [Google Scholar]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Han, X.; Wu, J.; Zheng, S.; Shang, Z.; Sun, D.; Zhou, R.; Li, B. A heat-activated calcium-permeable channel–Arabidopsis cyclic nucleotide-gated ion channel 6–is involved in heat shock responses. Wiley Online Libr. 2012, 70, 1056–1069. [Google Scholar] [CrossRef]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Network of Plant Heat Stress Response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 Overexpression Induces COR Genes and Enhances Freezing Tolerance. Science 1998, 280, 104–106. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Harding, S.A.; Guikema, J.A.; Paulsen, G.M. Photosynthetic decline from high temperature stress during maturation of wheat: I. Interaction with senescence processes. Plant Physiol. 1990, 92, 648–653. [Google Scholar] [CrossRef] [Green Version]
- Bheemanahalli, R.; Sunoj, V.S.J.; Saripalli, G.; Prasad, P.V.V.; Balyan, H.S.; Gupta, P.K.; Grant, N.; Gill, K.S.; Jagadish, S.V.K. Quantifying the impact of heat stress on pollen germination, seed set, and grain filling in spring wheat. Crop Sci. 2019, 59, 684–696. [Google Scholar] [CrossRef]
- Aiqing, S.; Somayanda, I.; Sebastian, S.V.; Singh, K.; Gill, K.; Prasad, P.V.V.; Jagadish, S.V.K. Heat Stress during Flowering Affects Time of Day of Flowering, Seed Set, and Grain Quality in Spring Wheat. Crop Sci. 2018, 58, 380–392. [Google Scholar] [CrossRef]
- Cheng, C.; Gao, X.; Feng, B.; Sheen, J.; Shan, L.; He, P. Plant immune response to pathogens differs with changing temperatures. Nat. Commun. 2013, 4, 2530. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Chand, R.; Vasistha, N.K.; Pandey, S.P.; Kumar, U.; Mishra, V.K.; Joshi, A.K. Spot blotch disease of wheat: The current status of research on genetics and breeding. Wiley Online Libr. 2018, 67, 508–531. [Google Scholar] [CrossRef]
- Dubin, H.J.; Van Ginkel, M. The status of wheat diseases and disease research in warmer areas. In Wheat for the Nontraditional Warm Areas: In Proceedings of the International Conference, Foz do Iguacu, Brazil, 29 July–3 August 1990; CIMMYT United Nations Development Program: Mexico City, Mexico, 1991; pp. 125–145. [Google Scholar]
- Duveiller, E.; Dubin, H.J.; Reeves, J.; McNab, A. Helminthosporium Blights of Wheat: Spot Blotch and Tan Spot; International Maize and Wheat Improvement Center: Mexico City, Mexico, 1997. [Google Scholar]
- Joshi, A.K.; Mishra, B.; Chatrath, R.; Ortiz Ferrara, G.; Singh, R.P. Wheat improvement in India: Present status, emerging challenges and future prospects. Euphytica 2007, 157, 431–446. [Google Scholar] [CrossRef]
- Joshi, A.K.; Ortiz-Ferrara, G.; Crossa, J.; Singh, G.; Sharma, R.C.; Chand, R.; Parsad, R. Combining superior agronomic performance and terminal heat tolerance with resistance to spot blotch (Bipolaris sorokiniana) of wheat in the warm humid Gangetic Plains of South Asia. Field Crops Res. 2007, 103, 53–61. [Google Scholar] [CrossRef]
- Chaurasia, S.; Chand, R.; Joshi, A.K. Relative dominance of Alternaria triticina Pras. et Prab. and Bipolaris sorokiniana (Sacc.) Shoemaker in different growth stages of wheat (T. aestivum L.). Z. Fur Pflanzenkrankh. Und Pflanzenschutz 2000, 107, 176–181. [Google Scholar]
- Sharma, R.C.; Duveiller, E.; Ortiz-Ferrara, G. Progress and challenge towards reducing wheat spot blotch threat in the Eastern Gangetic Plains of South Asia: Is climate change already taking its toll? Field Crops Res. 2007, 103, 109–118. [Google Scholar] [CrossRef]
- Mercado Vergnes, D.; Renard, M.E.; Duveiller, E.; Maraite, H. Effect of growth stage on host sensitivity to helminthosporol toxin and susceptibility to Cochliobolus sativus causing spot blotch on wheat. Physiol. Mol. Plant Pathol. 2006, 68, 14–21. [Google Scholar] [CrossRef]
- Schützendübel, A.; Stadler, M.; Wallner, D.; Von Tiedemann, A. A hypothesis on physiological alterations during plant ontogenesis governing susceptibility of winter barley to ramularia leaf spot. Plant Pathol. 2008, 57, 518–526. [Google Scholar] [CrossRef]
- Havis, N.D.; Brown, J.K.M.; Clemente, G.; Frei, P.; Jedryczka, M.; Kaczmarek, J.; Kaczmarek, M.; Matusinsky, P.; McGrann, G.R.D.; Piotrowska, M.; et al. Ramularia collo-cygni—An emerging pathogen of barley crops. Phytopathology 2015, 105, 895–904. [Google Scholar] [CrossRef] [Green Version]
- McGrann, G.R.D.; Steed, A.; Burt, C.; Goddard, R.; Lachaux, C.; Bansal, A.; Corbitt, M.; Gorniak, K.; Nicholson, P.; Brown, J.K.M. Contribution of the drought tolerance-related Stress-responsive NAC1 transcription factor to resistance of barley to Ramularia leaf spot. Mol. Plant Pathol. 2015, 16, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Mařík, P.; Šnejdar, Z.; Matušinsky, P. Expression of resistance to ramularia leaf spot in winter barley cultivars grown in conditions of the Czech Republic. Czech J. Genet. Plant Breed. 2011, 47, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Hoheneder, F.; Hofer, K.; Groth, J.; Herz, M.; Heß, M.; Hückelhoven, R. Ramularia leaf spot disease of barley is highly host genotype-dependent and suppressed by continuous drought stress in the field. J. Plant Dis. Prot. 2021, 128, 749–767. [Google Scholar] [CrossRef]
- Syme, R.A.; Tan, K.C.; Hane, J.K.; Dodhia, K.; Stoll, T.; Hastie, M.; Furuki, E.; Ellwood, S.R.; Williams, A.H.; Tan, Y.F.; et al. Comprehensive Annotation of the Parastagonospora nodorum Reference Genome Using Next-Generation Genomics, Transcriptomics and Proteogenomics. PLoS ONE 2016, 11, e0147221. [Google Scholar] [CrossRef]
- Shaw, M.W.; Bearchell, S.J.; Fitt, B.D.L.; Fraaije, B.A. Long-term relationships between environment and abundance in wheat of Phaeosphaeria nodorum and Mycosphaerella graminicola. New Phytol. 2008, 177, 229–238. [Google Scholar] [CrossRef]
- Downie, R.C.; Lin, M.; Corsi, B.; Ficke, A.; Lillemo, M.; Oliver, R.P.; Phan, H.T.T.; Tan, K.C.; Cockram, J. Septoria nodorum blotch of wheat: Disease management and resistance breeding in the face of shifting disease dynamics and a changing environment. Phytopathology 2021, 111, 906–920. [Google Scholar] [CrossRef]
- Bhathal, J.S.; Loughman, R.; Speijers, J. Yield reduction in wheat in relation to leaf disease from yellow (tan) spot and septoria nodorum blotch. Eur. J. Plant Pathol. 2003, 109, 435–443. [Google Scholar] [CrossRef]
- Bishaw, Z.; Struik, P.C.; Van Gastel, A.J.G. Farmer’s seed sources and seed quality: 2. seed health. Int. J. Plant Prod. 2013, 7, 637–658. [Google Scholar]
- Yang, C.; Zou, Z.; Fernando, W.G.D. The Effect of Temperature on the Hypersensitive Response (HR) in the Brassica napus–Leptosphaeria maculans Pathosystem. Plants 2021, 10, 843. [Google Scholar] [CrossRef]
- Huang, Y.J.; Evans, N.; Li, Z.Q.; Eckert, M.; Chèvre, A.M.; Renard, M.; Fitt, B.D. Temperature and leaf wetness duration affect phenotypic expression of Rlm6-mediated resistance to Leptosphaeria maculans in Brassica napus. New Phytol. 2006, 170, 129–141. [Google Scholar] [CrossRef]
- West, J.S.; Kharbanda, P.D.; Barbetti, M.J.; Fitt, B.D.L. Epidemiology and management of Leptosphaeria maculans (phoma stem canker) on oilseed rape in Australia, Canada and Europe. Plant Pathol. 2001, 50, 10–27. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Nurhasanah Ritonga, F.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stresses. Volume II. Water, Radiation, Salt, and Other Stresses; Academic Press: Cambridge, MA, USA, 1980. [Google Scholar]
- John, R.; Anjum, N.A.; Sopory, S.K.; Akram, N.A.; Ashraf, M. Some key physiological and molecular processes of cold acclimation. Biol. Plant. 2016, 60, 603–618. [Google Scholar] [CrossRef]
- Chen, L.J.; Xiang, H.Z.; Miao, Y.; Zhang, L.; Guo, Z.F.; Zhao, X.H.; Lin, J.W.; Li, T.L. An Overview of Cold Resistance in Plants. J. Agron. Crop Sci. 2014, 200, 237–245. [Google Scholar] [CrossRef]
- Lucau-Danila, A.; Toitot, C.; Goulas, E.; Blervacq, A.S.; Hot, D.; Bahrman, N.; Sellier, H.; Lejeune-Hénaut, I.; Delbreil, B. Transcriptome analysis in pea allows to distinguish chilling and acclimation mechanisms. Plant Physiol. Biochem. 2012, 58, 236–244. [Google Scholar] [CrossRef]
- Zhao, J.; Li, S.; Jiang, T.; Liu, Z.; Zhang, W.; Jian, G.; Qi, F. Chilling stress-the key predisposing factor for causing alternaria alternata infection and leading to cotton (Gossypium hirsutum L.) leaf senescence. PLoS ONE 2012, 7, e36126. [Google Scholar] [CrossRef] [Green Version]
- Kemal, S.A.; Bencheqroun, S.K.; Hamwieh, A.; Imtiaz, M. Effects of temperature stresses on the resistance of chickpea genotypes and aggressiveness of Didymella rabiei isolates. Front. Plant Sci. 2017, 8, 1607. [Google Scholar] [CrossRef] [Green Version]
- Bashan, Y.; Hernandez-Saavedra, N.Y. Alternaria-blight of cotton: Epidemiology and transmission. In Alternaria Biology, Plant Diseases and Metabolites; Chelkowski, J., Visconti, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1992; pp. 233–266. [Google Scholar]
- Bashi, E.; Sachs, Y.; Rotem, J. Relationships between disease and yield in cotton fields affected by Alternaria macrospora. Phytoparasitica 1983, 11, 89–98. [Google Scholar] [CrossRef]
- Muehlbauer, F.J.; Chen, W. Resistance to ascochyta blights of cool season food legumes. Ascochyta Blights Grain Legum. 2007, 119, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Nizam, S.; Sinha, M.; Verma, P.K. Comparative Transcriptome Analysis of the Necrotrophic Fungus Ascochyta rabiei during Oxidative Stress: Insight for Fungal Survival in the Host Plant. PLoS ONE 2012, 7, e33128. [Google Scholar] [CrossRef]
- Nene, Y.L. A review of Ascochyta blight of chickpea. Trop. Pest Manag. 1982, 28, 61–70. [Google Scholar] [CrossRef]
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.K.; Kwon, W.-T.; Laprise, R. Regional Climate Projections—Supplementary Material; Cambridge University Press: Cambridge, UK, 2007; Volume 23. [Google Scholar]
- Fukao, T.; Bailey-Serres, J. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 16814–16819. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann-Benning, S.; Kende, H. On the Role of Abscisic Acid and Gibberellin in the Regulation of Growth in Rice. Plant Physiol. 1992, 99, 1156–1161. [Google Scholar] [CrossRef]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef]
- Melcher, K.; Ng, L.M.; Zhou, X.E.; Soon, F.F.; Xu, Y.; Suino-Powell, K.M.; Park, S.Y.; Weiner, J.J.; Fujii, H.; Chinnusamy, V.; et al. A gate–latch–lock mechanism for hormone signalling by abscisic acid receptors. Nature 2009, 462, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Palmer, T.N.; Räisänen, J. Quantifying the risk of extreme seasonal precipitation events in a changing climate. Nature 2002, 415, 512–514. [Google Scholar] [CrossRef]
- Christensen, J.H.; Christensen, O.B. Severe summertime flooding in Europe. Nature 2003, 421, 805–806. [Google Scholar] [CrossRef]
- Hemantaranjan, A. Flooding: Abiotic Constraint Limiting Vegetable Productivity. Adv. Plants Agric. Res. 2014, 1, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Tewari, S.; Mishra, A. Flooding Stress in Plants and Approaches to Overcome. In Plant Metab. Regul. Under Environ. Stress; Ahmad, P., Ahanger, M.A., Singh, V.P., Tripathi, D.K., Alam, P., Alyemeni, M.N., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 355–366. [Google Scholar] [CrossRef]
- Parent, C.; Capelli, N.; Berger, A.; Crèvecoeur, M.; Dat, J.F. An overview of plant responses to soil waterlogging. Plant Stress 2008, 2, 20–27. [Google Scholar]
- Lu, Y.; Watanabe, A.; Kimura, M. Contribution of plant photosynthates to dissolved organic carbon in a flooded rice soil. Biogeochemistry 2004, 71, 1–15. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Rennenberg, H. Molecular and physiological responses of trees to waterlogging stress. Plant Cell Environ. 2014, 37, 2245–2259. [Google Scholar] [CrossRef]
- Irfan, M.; Hayat, S.; Hayat, Q.; Afroz, S.; Ahmad, A. Physiological and biochemical changes in plants under waterlogging. Protoplasma 2010, 241, 3–17. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Havis, N.D.; Nyman, M.; Oxley, S.J.P. Evidence for seed transmission and symptomless growth of Ramularia collo-cygni in barley (Hordeum vulgare). Plant Pathol. 2014, 63, 929–936. [Google Scholar] [CrossRef]
- McGrann, G.R.D.; Havis, N.D. Ramularia leaf spot: A newly important threat to barley production. Outlooks Pest Manag. 2017, 28, 65–69. [Google Scholar] [CrossRef]
- McGrann, G.R.D.; Brown, J.K.M. The role of reactive oxygen in the development of Ramularia leaf spot disease in barley seedlings. Ann. Bot. 2018, 121, 415–430. [Google Scholar] [CrossRef] [Green Version]
- Thornton, P.K.; Ericksen, P.J.; Herrero, M.; Challinor, A.J. Climate variability and vulnerability to climate change: A review. Glob. Chang. Biol. 2014, 20, 3313–3328. [Google Scholar] [CrossRef]
- Alamri, S.; Hu, Y.; Mukherjee, S.; Aftab, T.; Fahad, S.; Raza, A.; Ahmad, M.; Siddiqui, M.H. Silicon-induced postponement of leaf senescence is accompanied by modulation of antioxidative defense and ion homeostasis in mustard (Brassica juncea) seedlings exposed to salinity and drought stress. Plant Physiol. Biochem. 2020, 157, 47–59. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Guan, X.K.; Song, L.; Wang, T.C.; Turner, N.C.; Li, F.M. Effect of Drought on the Gas Exchange, Chlorophyll Fluorescence and Yield of Six Different-Era Spring Wheat Cultivars. J. Agron. Crop Sci. 2015, 201, 253–266. [Google Scholar] [CrossRef]
- Loggini, B.; Scartazza, A.; Brugnoli, E.; Navari-Izzo, F. Antioxidative Defense System, Pigment Composition, and Photosynthetic Efficiency in Two Wheat Cultivars Subjected to Drought. Plant Physiol. 1999, 119, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Allu, A.D.; Soja, A.M.; Wu, A.; Szymanski, J.; Balazadeh, S. Salt stress and senescence: Identification of cross-talk regulatory components. J. Exp. Bot. 2014, 65, 3993–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Zong, W.; Li, X.; Ning, J.; Hu, H.; Li, X.; Xiao, J.; Xiong, L. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 2013, 64, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Irulappan, V.; Kandpal, M.; Saini, K.; Rai, A.; Ranjan, A.; Sinharoy, S.; Senthil-Kumar, M. Drought stress exacerbates fungal colonization and endodermal invasion and dampens defense responses to increase dry root rot in chickpea. Mol. Plant Microbe Interact. 2022. [Google Scholar] [CrossRef]
- Marquez, N.; Giachero, M.L.; Declerck, S.; Ducasse, D.A. Macrophomina phaseolina: General Characteristics of Pathogenicity and Methods of Control. Front. Plant Sci. 2021, 12, 666. [Google Scholar] [CrossRef]
- Schroeder, M.M.; Lai, Y.; Shirai, M.; Alsalek, N.; Tsuchiya, T.; Roberts, P.; Eulgem, T. A novel Arabidopsis pathosystem reveals cooperation of multiple hormonal response-pathways in host resistance against the global crop destroyer Macrophomina phaseolina. Sci. Rep. 2019, 9, 20083. [Google Scholar] [CrossRef]
- Singh, S.K. Influence of Cropping Systems on Macrophomina phaseolina Populations in Soil. Plant Dis. 1990, 74, 812. [Google Scholar] [CrossRef]
- Kaur, S.; Dhillon, G.S.; Brar, S.K.; Vallad, G.E.; Chand, R.; Chauhan, V.B. Emerging phytopathogen Macrophomina phaseolina: Biology, economic importance and current diagnostic trends. Crit. Rev. Microbiol. 2012, 38, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Mengistu, A.; Ray, J.D.; Smith, J.R.; Arelli, P.R.; Bellaloui, N.; Chen, P.; Shannon, G.; Boykin, D. Effect of charcoal rot on selected putative drought tolerant soybean genotypes and yield. Crop Prot. 2018, 105, 90–101. [Google Scholar] [CrossRef]
- Mihail, J.D. Macrophomina phaseolina: Spatio-temporal dynamics of inoculum and of disease in a highly susceptible crop. Phytopathology 1989, 79, 848–855. [Google Scholar] [CrossRef]
- Goudarzi, A.; Banihashemi, Z.; Maftoun, M. Effect of water potential on sclerotial germination and mycelial growth of Macrophomina phaseolina. Phytopathol. Mediterr. 2008, 47, 107–114. [Google Scholar]
- Rengasamy, P. Soil processes affecting crop production in salt-affected soils. Funct. Plant Biol. 2010, 37, 613–620. [Google Scholar] [CrossRef]
- Paul, K.; Pauk, J.; Kondic-Spika, A.; Grausgruber, H.; Allahverdiyev, T.; Sass, L.; Vass, I. Co-occurrence of mild salinity and drought synergistically enhances biomass and grain retardation in wheat. Front. Plant Sci. 2019, 10, 501. [Google Scholar] [CrossRef]
- Nedjimi, B. Salinity tolerance: Growth, mineral nutrients, and roles of organic osmolytes, case of Lygeum spartum L., A review. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: Delhi, India, 2015; pp. 27–35. [Google Scholar] [CrossRef]
- Arzani, A.; Ashraf, M. Smart Engineering of Genetic Resources for Enhanced Salinity Tolerance in Crop Plants. CRC Crit. Rev. Plant Sci. 2016, 35, 146–189. [Google Scholar] [CrossRef]
- Mushtaq, Z.; Faizan, S.; Gulzar, B. Salt stress, its impacts on plants and the strategies plants are employing against it: A review. J. Appl. Biol. Biotechnol. 2020, 8, 81–91. [Google Scholar] [CrossRef]
- Zhu, J.K. Cell signaling under salt, water and cold stresses. Curr. Opin. Plant Biol. 2001, 4, 401–406. [Google Scholar] [CrossRef]
- Liu, J.; Ishitani, M.; Halfter, U.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000, 97, 3730–3734. [Google Scholar] [CrossRef]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B.; Sussman, M.R. A peptide hormone and its receptor protein kinase regulate plant cell expansion. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Kita, D.; Peaucelle, A.; Cartwright, H.N.; Doan, V.; Duan, Q.; Liu, M.C.; Maman, J.; Steinhorst, L.; Schmitz-Thom, I.; et al. The FERONIA Receptor Kinase Maintains Cell-Wall Integrity during Salt Stress through Ca2+ Signaling. Curr. Biol. 2018, 28, 666–e5. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ellwood, S.R.; Oliver, R.P.; Friesen, T.L. Pyrenophora teres: Profile of an increasingly damaging barley pathogen. Mol. Plant Pathol. 2011, 12, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Murray, G.M.; Brennan, J.P. Estimating disease losses to the Australian barley industry. Australas. Plant Pathol. 2010, 39, 85–96. [Google Scholar] [CrossRef]
- Grewal, T.S.; Rossnagel, B.G.; Pozniak, C.J.; Scoles, G.J. Mapping quantitative trait loci associated with barley net blotch resistance. Theor. Appl. Genet. 2008, 116, 529–539. [Google Scholar] [CrossRef]
- Ben Alaya, A.; Rabhi, F.; Hessini, K.; Djébali, N. Pyrenophora teres growth and severity of net blotch on barley under salt stress. Eur. J. Plant Pathol. 2021, 161, 709–722. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant-pathogen interactions. Curr. Opin. Plant Biol. 2005, 8, 409–414. [Google Scholar] [CrossRef]
- Ben Rejeb, I.; Pastor, V.; Mauch-Mani, B. Plant Responses to Simultaneous Biotic and Abiotic Stress: Molecular Mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef]
- Gorshkov, V.; Tsers, I. Plant susceptible responses: The underestimated side of plant–pathogen interactions. Biol. Rev. 2022, 97, 45–66. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, K.A.; El-shawy, E.A.; Mohamed Hafez, Y.; Abdel-dayem, S.M.A.; Chidya, R.C.G.; Saneoka, H.; Sabagh, A. EL Nano-Silver and non-traditional compounds mitigate the adverse effects of net blotch disease of barley in correlation with up-regulation of antioxidant enzymes. Pakistan J. Bot. 2020, 52, 1065–1072. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. J. Integr. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kang, M.H.; Kim, J.Y.; Lim, P.O. The Role of Light and Circadian Clock in Regulation of Leaf Senescence. Front. Plant Sci. 2021, 12, 669170. [Google Scholar] [CrossRef] [PubMed]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N. ROS-dependent signalling pathways in plants and algae exposed to high light: Comparisons with other eukaryotes. Free Radic. Biol. Med. 2018, 122, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2020, 44, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Maai, E.; Nishimura, K.; Takisawa, R.; Nakazaki, T. Light stress-induced chloroplast movement and midday depression of photosynthesis in sorghum leaves. Plant Prod. Sci. 2020, 23, 172–181. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fritschi, F.B.; Mittler, R. Global Warming, Climate Change, and Environmental Pollution: Recipe for a Multifactorial Stress Combination Disaster. Trends Plant Sci. 2021, 26, 588–599. [Google Scholar] [CrossRef]
- Griebel, T.; Zeier, J. Light regulation and daytime dependency of inducible plant defenses in arabidopsis: Phytochrome signaling controls systemic acquired resistance rather than local defense. Plant Physiol. 2008, 147, 790–801. [Google Scholar] [CrossRef] [Green Version]
- Vannier, N.; Agler, M.; Hacquard, S. Microbiota-mediated disease resistance in plants. PLoS Pathog. 2019, 15, e1007740. [Google Scholar] [CrossRef] [Green Version]
- Trotta, A.; Rahikainen, M.; Konert, G.; Finazzi, G.; Kangasjärvi, S. Signalling crosstalk in light stress and immune reactions in plants. Philos. Trans. R. Soc. B 2014, 369, 20130235. [Google Scholar] [CrossRef] [Green Version]
- De Vallavieille-Pope, C.; Huber, L.; Leconte, M.; Bethenod, O. Preinoculation effects of light quantity on infection efficiency of Puccinia striiformis and P. triticina on wheat seedlings. Phytopathology 2002, 92, 1308–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makepeace, J.C.; Havis, N.D.; Burke, J.I.; Oxley, S.J.P.; Brown, J.K.M. A method of inoculating barley seedlings with Ramularia collo-cygni. Plant Pathol. 2008, 57, 991–999. [Google Scholar] [CrossRef]
- Li, Z.; Wakao, S.; Fischer, B.; Niyogi, K. Sensing and responding to excess light. Annu. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Heiser, I.; Heß, M.; Schmidtke, K.-U.; Vogler, U.; Miethbauer, S.; Liebermann, B. Fatty acid peroxidation by rubellin B, C and D, phytotoxins produced by Ramularia collo-cygni (Sutton et Waller). Physiol. Mol. Plant Pathol. 2004, 64, 135–143. [Google Scholar] [CrossRef]
- Dussart, F.; Douglas, R.; Sjökvist, E.; Hoebe, P.N.; Spoel, S.H.; McGrann, G.R.D. Genome-Based Discovery of Polyketide-Derived Secondary Metabolism Pathways in the Barley Pathogen Ramularia collo-cygni. Mol. Plant-Microbe Interact. 2018, 31, 962–975. [Google Scholar] [CrossRef] [Green Version]
- Dussart, F.; Jakubczyk, D. Biosynthesis of Rubellins in Ramularia collo-cygni—Genetic Basis and Pathway Proposition. Int. J. Mol. Sci. 2022, 23, 3475. [Google Scholar] [CrossRef]
- Daamen, R.; Stol, W. Surveys of cereal diseases and pests in the Netherlands. 5. Occurrence of Septoria spp. in winter wheat. Neth. J. Plant Pathol. 1992, 98, 369–376. [Google Scholar] [CrossRef]
- Arraiano, L.S.; Brading, P.A.; Brown, J.K.M. A detached seedling leaf technique to study resistance to Mycosphaerella graminicola (anamorph Septoria tritici) in wheat. Plant Pathol. 2001, 50, 339–346. [Google Scholar] [CrossRef]
- Mycosphaerella pini. EPPO Bull. 2005, 35, 303–306. [CrossRef]
- Bradshaw, R.E. Dothistroma (red-band) needle blight of pines and the dothistromin toxin: A review. For. Pathol. 2004, 34, 163–185. [Google Scholar] [CrossRef]
- Kabir, M.S.; Ganley, R.J.; Bradshaw, R.E. Dothistromin toxin is a virulence factor in dothistroma needle blight of pines. Plant Pathol. 2015, 64, 225–234. [Google Scholar] [CrossRef]
- Busogoro, J.P.; Etame, J.; Lognay, G.; Messiaen, J.; Van Cutsem, P.; Lepoivre, P. Analysis of the mechanisms of action of Mycosphaerella fijiensis toxins during the development of black leaf streak disease. In Banana Improvement: Cellular, Molecular Biology, and Induced Mutations. Proceedings of A Meeting Held in Leuven, Belgium, 24–28 September 2001; Science Publishers, Inc.: New York, NY, USA, 2004; pp. 171–181. [Google Scholar]
- Noar, R.D.; Thomas, E.; Daub, M.E. Genetic Characteristics and Metabolic Interactions between Pseudocercospora fijiensis and Banana: Progress toward Controlling Black Sigatoka. Plants 2022, 11, 948. [Google Scholar] [CrossRef] [PubMed]
- Hartman, P.E.; Suzuki, C.K.; Stack, M.E. Photodynamic production of superoxide in vitro by altertoxins in the presence of reducing agents. Appl. Environ. Microbiol. 1989, 55, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daub, M.E.; Ehrenshaft, M. The photoactivated Cercospora toxin cercosporin: Contributions to plant disease and fundamental biology. Annu. Rev. Phytopathol. 2000, 38, 461–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, T.; Shimanuki, T.; Araki, T.; Sakamura, S. Phleichrome; a new phytotoxic compound produced by Cladosporium phlei. Agric. Biol. Chem. 1975, 39, 1683–1684. [Google Scholar] [CrossRef]
- Daub, M.E.; Herrero, S.; Chung, K.R. Photoactivated perylenequinone toxins in fungal pathogenesis of plants. FEMS Microbiol. Lett. 2005, 252, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Upchurch, R.G.; Walker, D.C.; Rollins, J.A.; Ehrenshaft, M.; Daub, M.E. Mutants of Cercospora kikuchii altered in cercosporin synthesis and pathogenicity. Appl. Environ. Microbiol. 1991, 57, 2940–2945. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, S.; Okubo, A.; Akiyama, Y.; Fuwa, K. Cercosporin: A novel photodynamic pigment isolated from Cercospora kikuchii. Agric. Biol. Chem. 1975, 39, 287–288. [Google Scholar] [CrossRef] [Green Version]
- Daub, M.; Chung, K.-R. Cercosporin: A photoactivated toxin in plant disease. APSnet Featur. 2007, 1–12. [Google Scholar] [CrossRef]
- Sloat, L.L.; Davis, S.J.; Gerber, J.S.; Moore, F.C.; Ray, D.K.; West, P.C.; Mueller, N.D. Climate adaptation by crop migration. Nat. Commun. 2020, 11, 1243. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stress | Pathogen | Disease/Crop | Effect on Disease Development | References |
|---|---|---|---|---|
| Heat | Bipolaris sorokiniana (Cochliobolus sativus) | Spot blotch in cereals | Increase in spot blotch | [57,58,59,62,63,64,65] |
| Ramularia collo-cygni | Ramularia leaf spot (RLS) in barley | Decrease in RLS symptom development | [81,82] | |
| Parastagnorospora nodorum (Phaeosphaeria nodorum) | Septoria nodrum blotch and glume blotch in wheat | Reduced P. nodrum DNA in winter wheat grains | [71,72,75] | |
| Phoma lingam (Leptosphaeria maculans) | Blackleg and Phoma stem canker in oilseed rape | Increased lesion formation | [91] | |
| Cold | Alternaria spp. (A. alternata and A. macrospora) | Leaf spot in cotton | Increase in leaf spot | [85] |
| Ascochyta rabiei (Didymella rabiei) | Ascochyta blight in chickpea | Increase in Ascochyta blight symptoms | [86] | |
| Waterlogging & Flooding | Ramularia collo-cygni | RLS in barley | Increase in RLS symptom development | [119,120] |
| Drought | Ramularia collo-cygni | RLS in barley | Drought led to increased resistance to RLS | [80,82] |
| Macrophomina phaseolina | Dry root rot (DRR) in chickpea | Drought led to enhanced DRR infection and colonisation | [117] | |
| Salinity | Drechslera teres (Pyrenophora teres) | Net blotch in barley | High salt concentration led to increase in net blotch | [137,142] |
| Light | Ramularia collo-cygni | RLS in barley | Increased RLS disease symptoms | [120,162] |
| Zymoseptoria tritici (Mycosphaerella graminicola) | Septoria tritici blotch in wheat | Positive correlation between hours of sunshine and disease prevalence | [158,159] | |
| Dothistroma septosporum (Mycosphaerella pini) | Needle blight in various pine trees | Light intensity increased formation of symptoms | [160,161,162] | |
| Pseudocercospora fijiensis (Mycosphaerella fijiensis) | Black Sigatoka disease in banana | Shade decreases black Sigatoka lesion formation | [22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Röhrig, L.; Dussart, F. Does Abiotic Host Stress Favour Dothideomycete-Induced Disease Development? Plants 2022, 11, 1615. https://doi.org/10.3390/plants11121615
Röhrig L, Dussart F. Does Abiotic Host Stress Favour Dothideomycete-Induced Disease Development? Plants. 2022; 11(12):1615. https://doi.org/10.3390/plants11121615
Chicago/Turabian StyleRöhrig, Laura, and Francois Dussart. 2022. "Does Abiotic Host Stress Favour Dothideomycete-Induced Disease Development?" Plants 11, no. 12: 1615. https://doi.org/10.3390/plants11121615