
Ecological Niche Modeling of Water Lily (Nymphaea L.) Species in Australia under Climate Change to Ascertain Habitat Suitability for Conservation Measures

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Variable Selection and Performance of the Models
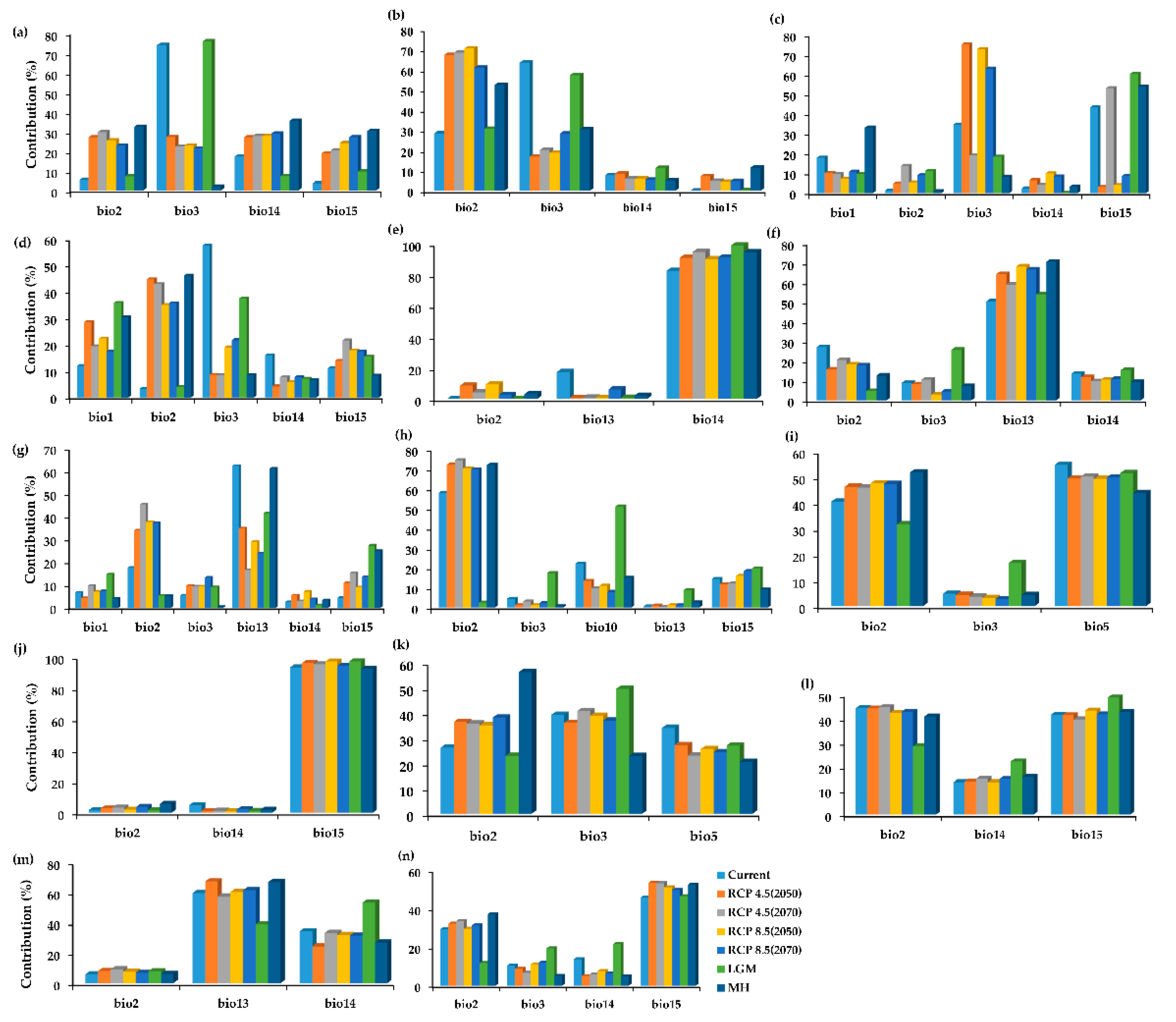
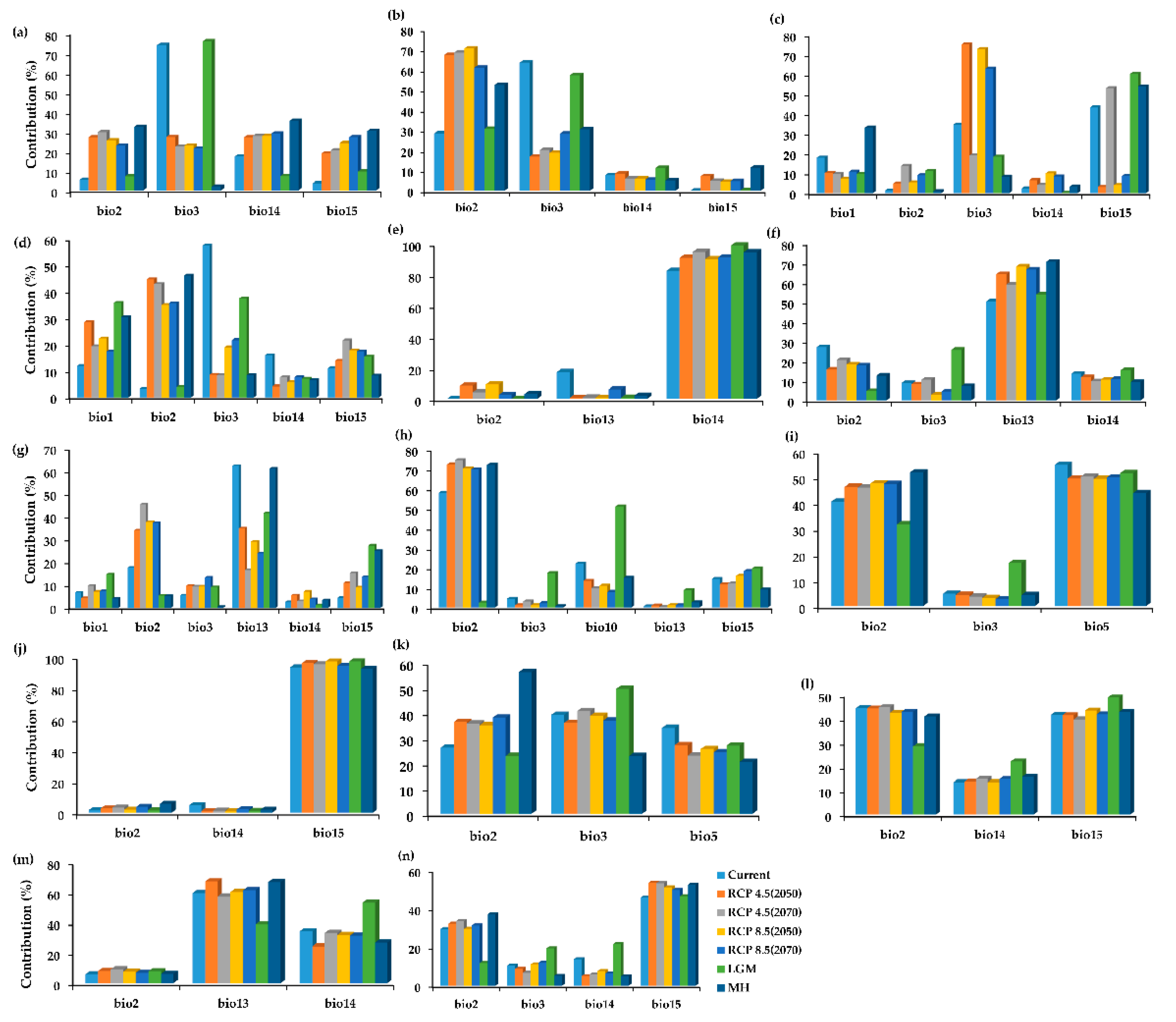
2.2. The Variable Contribution
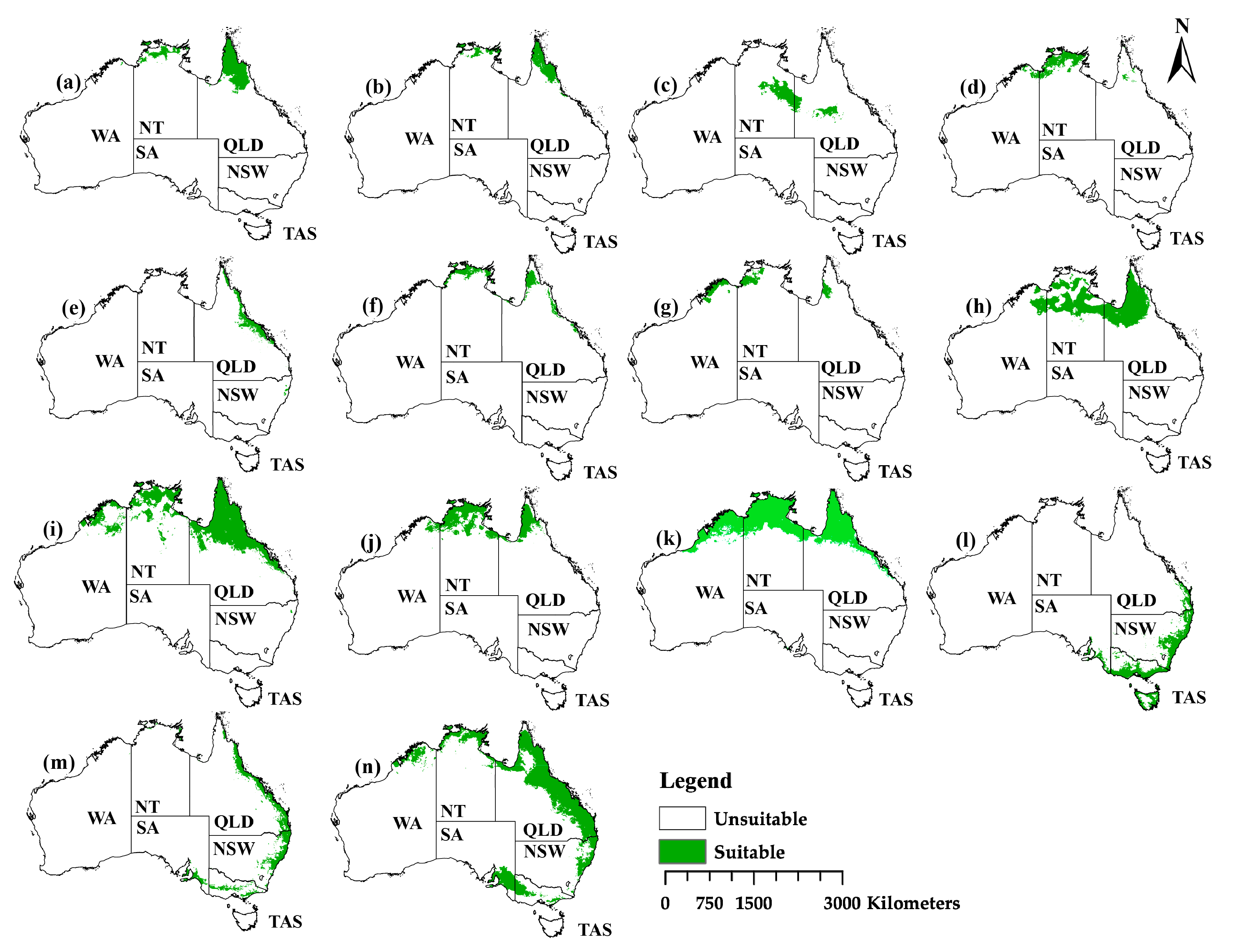
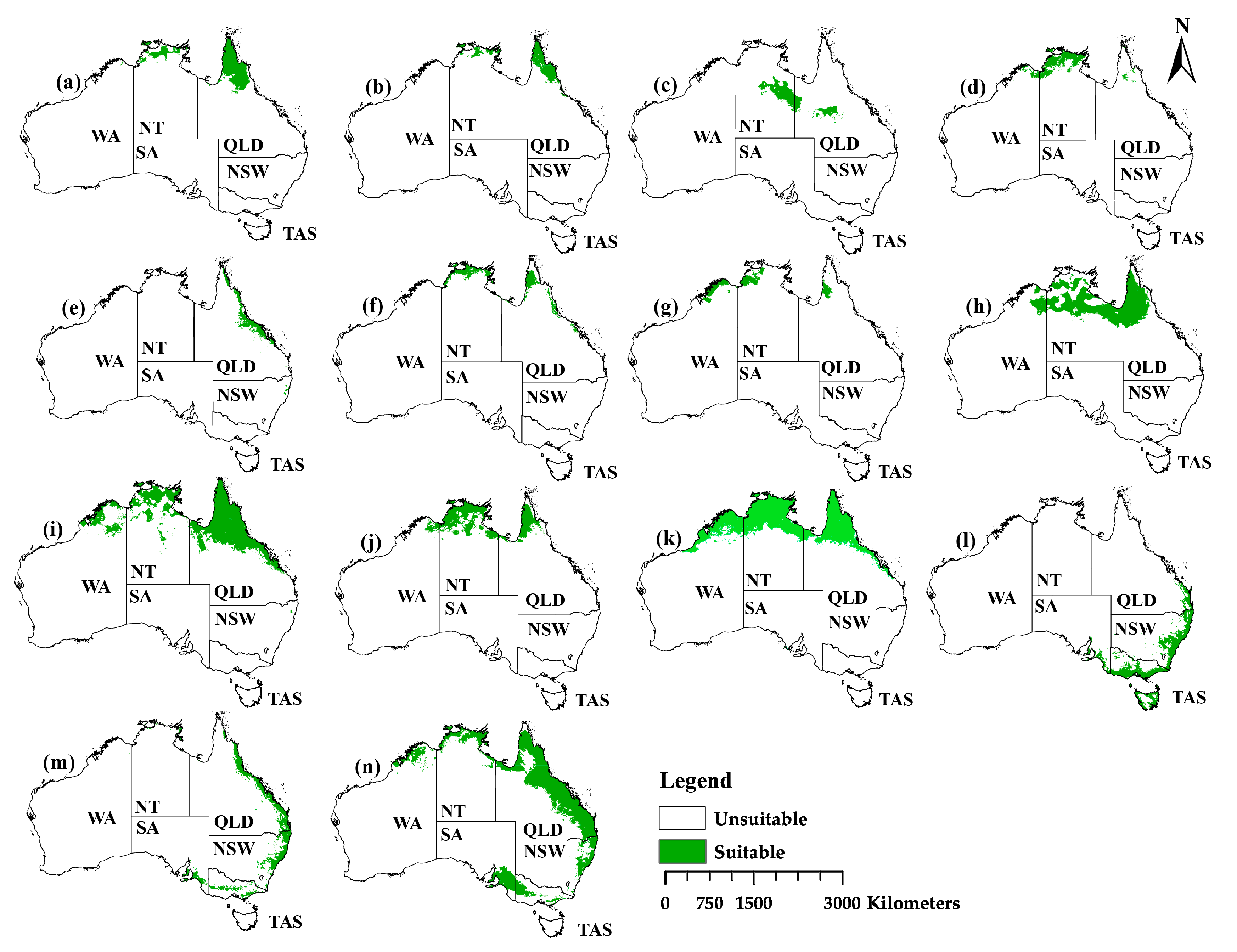
2.3. The Current Distribution
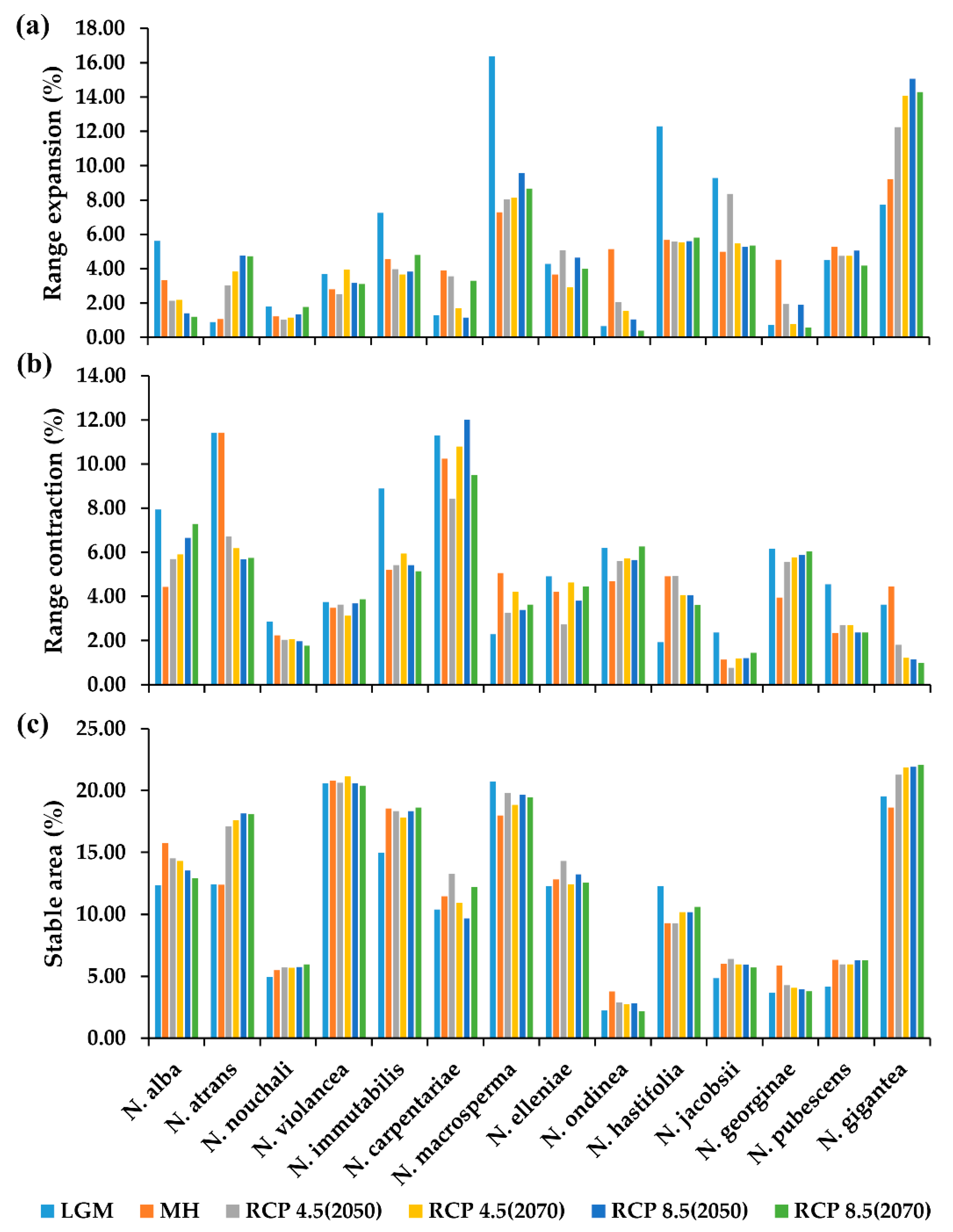
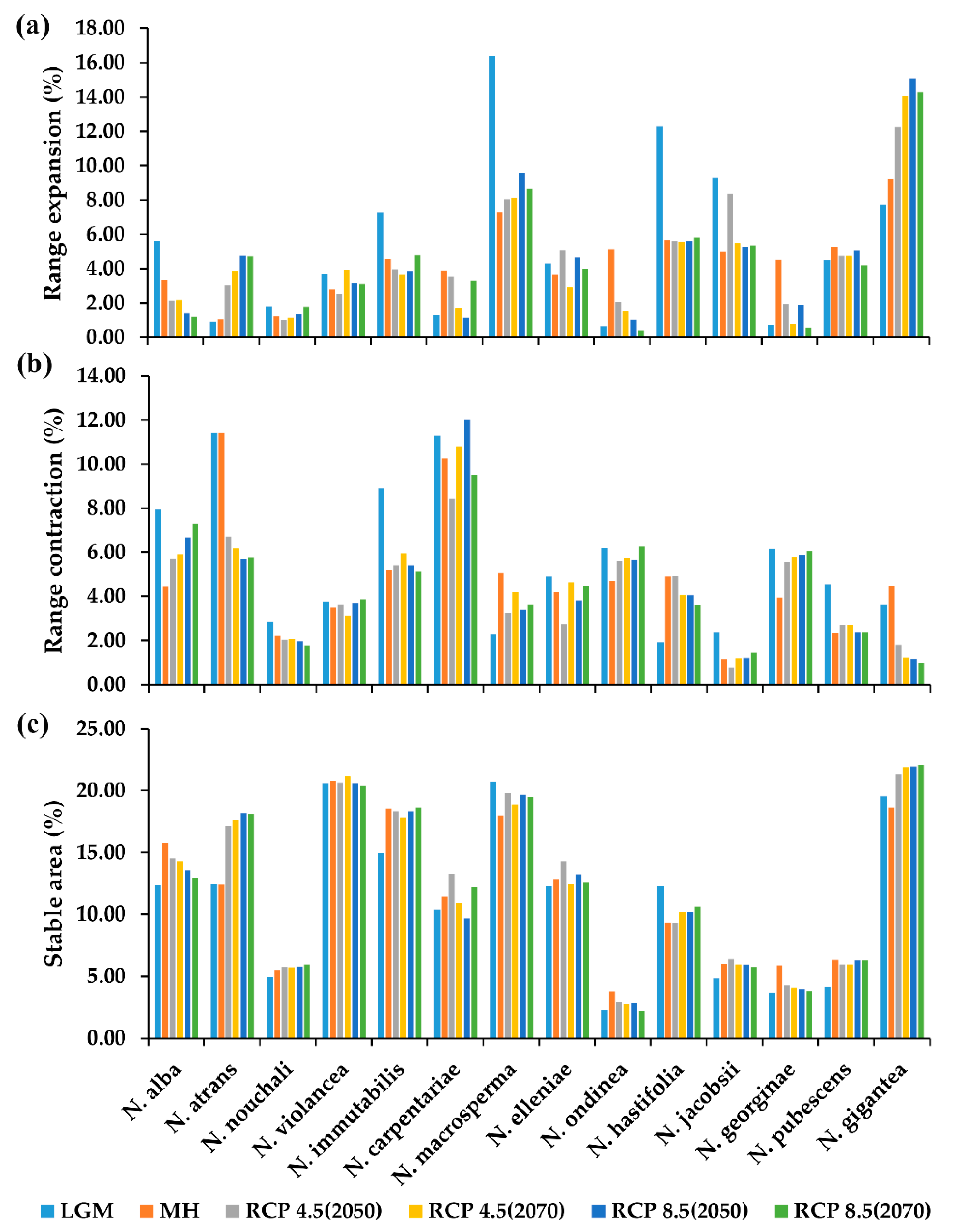
2.4. The Projection Changes for the Past and Future Distribution
3. Discussion
4. Materials and Methods
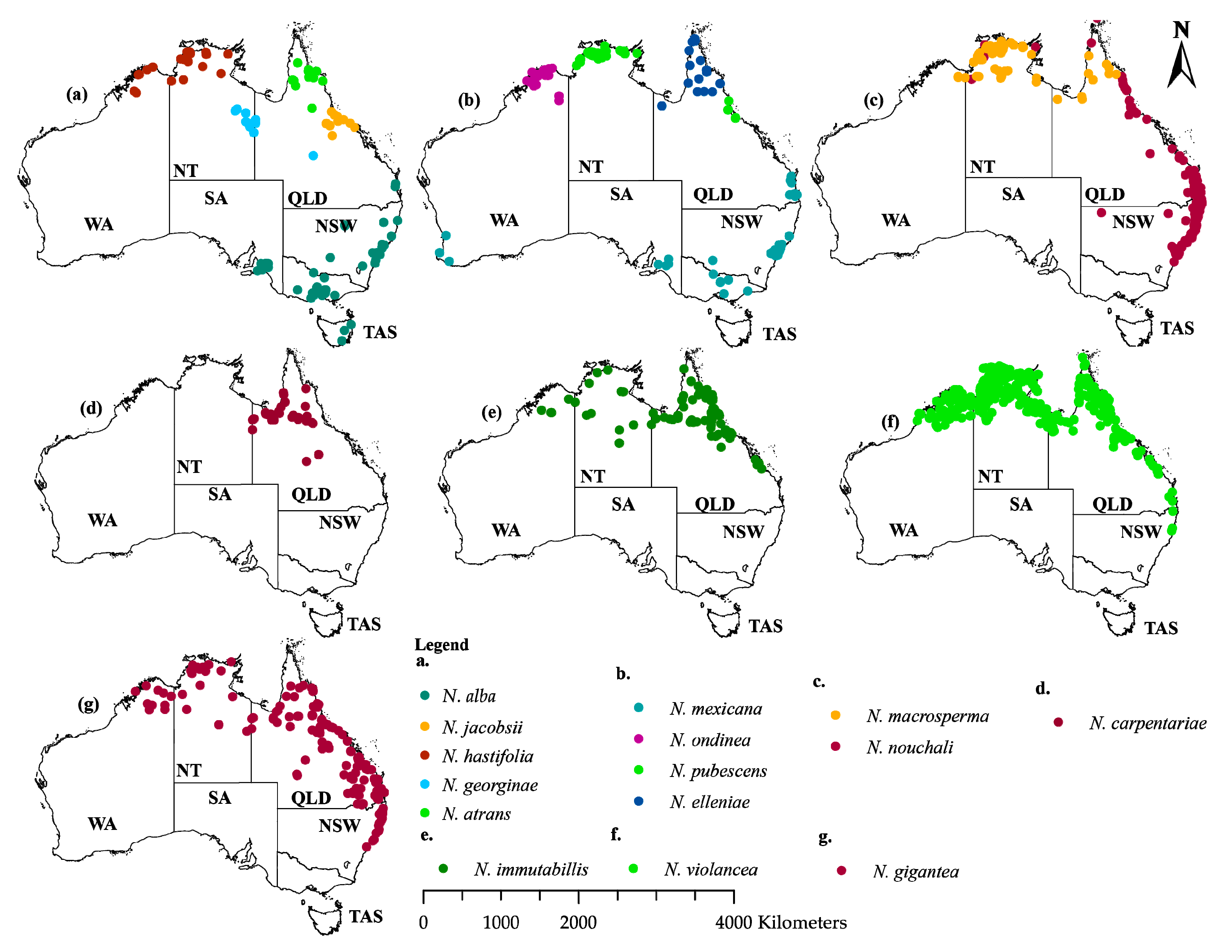
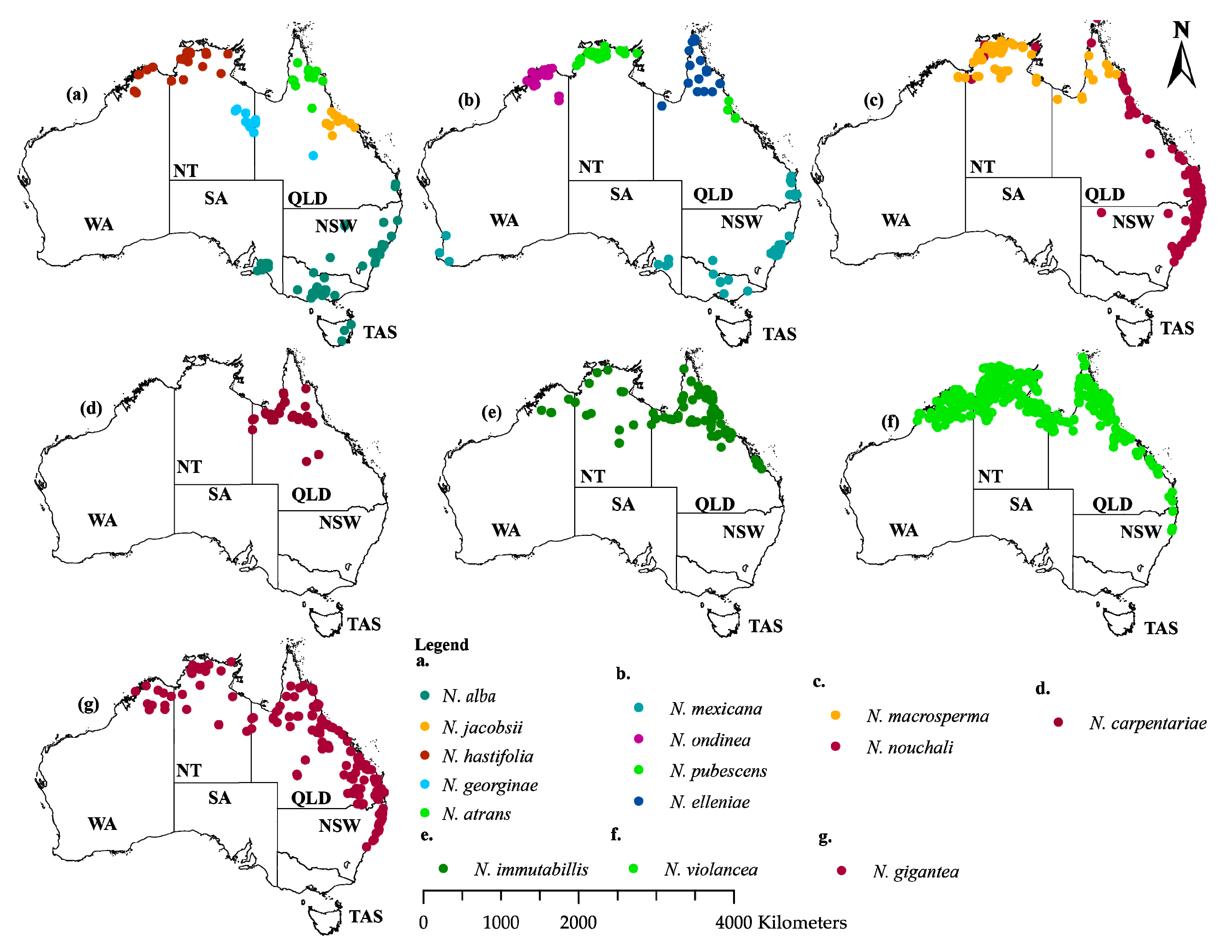
4.1. Distribution Data
4.2. Climatic Data
4.3. Model Building and Evaluation
4.4. Distribution of Habitat Suitability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Houghton, J.T.; Ding, Y.; Griggs, D.J.; Noguer, M.; van der Linden, P.J.; Dai, X.; Maskell, K.; Johnson, C. Climate Change 2001: The Scientific Basis: Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Alexander, L.V.; Allen, S.K.; Bindoff, N.L.; Bréon, F.M.; Church, J.A.; Cubasch, U.; Emori, S. Technical summary. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; pp. 33–115. [Google Scholar]
- WWF. Living Planet: Report 2016: Risk and Resilience in a New Era; World Wide Fund for Nature. 2016. Available online: http://assets.wwf.org.uk/custom/lpr2016/ (accessed on 28 March 2022).
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CSIRO, Burealal of Meteorology. State of the Climate 2012. Available online: www.climatechangeinaustralia.gov.au (accessed on 12 January 2022).
- Milić, D.; Radenković, S.; Radišić, D.; Andrić, A.; Nikolić, T.; Vujić, A. Stability and changes in the distribution of Pipiza hoverflies (Diptera, Syrphidae) in Europe under projected future climate conditions. PLoS ONE 2019, 14, e0221934. [Google Scholar] [CrossRef] [PubMed]
- Gaurnaut, R. The Garnaut Climate Change Review: Final Report; Cambridge University Press: Cambridge, UK, 2008; ISBN 9780521744447. [Google Scholar]
- Cait, W.; World Resources Institute. Climate Analysis Indicators Tool (WRI, CAIT); WRI CAIT: Washington, DC, USA, 2012; Available online: http://cait2.wri.org (accessed on 24 March 2022).
- Change, I.C. Synthesis Report. Contribution of Working Groups I. II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change 2014, 151. Available online: https://epic.awi.de/ (accessed on 24 March 2022).
- Dalziell, E.L.; Lewandrowski, W.; Merritt, D.J. Increased salinity reduces seed germination and impacts upon seedling development in Nymphaea L. (Nymphaeaceae) from northern Australia’s freshwater wetlands. Aquat. Bot. 2020, 165, 103235. [Google Scholar] [CrossRef]
- Oliver, E.C.; Benthuysen, J.A.; Bindoff, N.L.; Hobday, A.J.; Holbrook, N.J.; Mundy, C.N.; Perkins-Kirkpatrick, S.E. The unprecedented 2015/16 Tasman Sea marine heatwave. Nat. Commun. 2017, 8, 16101. [Google Scholar] [CrossRef] [Green Version]
- Saintilan, N.; Rogers, K.; Kelleway, J.J.; Ens, E.; Sloane, D.R. Climate change impacts on the coastal wetlands of Australia. Wetlands 2019, 39, 1145–1154. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Murphy, K.; Efremov, A.; Davidson, T.A.; Molina-Navarro, E.; Fidanza, K.; Betiol, T.C.C.; Chambers, P.; Grimaldo, J.T.; Martins, S.V.; Springuel, I. World distribution, diversity and endemism of aquatic macrophytes. Aquat. Bot. 2019, 158, 103127. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Araújo, M.B.; Anderson, R.P.; Márcia Barbosa, A.; Beale, C.M.; Dormann, C.F.; Early, R.; Garcia, R.A.; Guisan, A.; Maiorano, L.; Naimi, B. Standards for distribution models in biodiversity assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wolter, C.; Xian, W.; Jeschke, J.M. Species distribution models have limited spatial transferability for invasive species. Ecol. Lett. 2020, 23, 1682–1692. [Google Scholar] [CrossRef] [PubMed]
- Strubbe, D.; Beauchard, O.; Matthysen, E. Niche conservatism among non-native vertebrates in Europe and North America. Ecography 2015, 38, 321–329. [Google Scholar] [CrossRef]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [Green Version]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, S.; Porter, C. Nymphaeaceae. Flora Aust. 2007, 2, 259–275. [Google Scholar]
- Beaumont, L.J.; Hughes, L. Potential changes in the distributions of latitudinally restricted Australian butterfly species in response to climate change. Glob. Chang. Biol. 2002, 8, 954–971. [Google Scholar] [CrossRef]
- Argent, R. Australia State of the Environment 2016: Inland Water, Independent Report to the Australian Government Minister for the Environment and Energy. Commonwealth of Australia 2017 Australia State of the Environment 2016: Inland Water Is Licensed by the Commonwealth of Australia for Use under a Creative Commons Attribution 4.0 International Licence with the Exception of the Coat of Arms of the Commonwealth of Australia, the Logo of the Agency Responsible for Publishing the Report and Some Content Supplied by Third Parties. For License Conditions See Creative Commons. Org/Licenses/By/4.0. The Commonwealth of Australia Has Made All Reasonable Efforts to Identify and Attribute Content Supplied by Third Parties That Is Not Licensed for Use under Creative Commons Attribution 4.0 International. 2017, 10, 94. Available online: http://www.epochfuture.com (accessed on 18 March 2022).
- Steffen, W.; Burbidge, A.A.; Hughes, L.; Kitching, R.; Lindenmeyer, D.; Musgrave, W.; Smith, M.S.; Werner, P.A. Australia’s Biodiversity and Climate Change; CSIRO Publishing: Clayton, Australia, 2009. [Google Scholar]
- Cabrelli, A.; Beaumont, L.; Hughes, L. The impacts of climate change on Australian and New Zealand flora and fauna. In Austral Ark: The State of Wildlife in Australia and New Zealand; Cambridge University Press: Cambridge, UK, 2015; pp. 65–82. [Google Scholar] [CrossRef]
- Last, P.R.; White, W.T.; Gledhill, D.C.; Hobday, A.J.; Brown, R.; Edgar, G.J.; Pecl, G. Long-term shifts in abundance and distribution of a temperate fish fauna: A response to climate change and fishing practices. Glob. Ecol. Biogeogr. 2011, 20, 58–72. [Google Scholar] [CrossRef]
- Nzei, J.M.; Ngarega, B.K.; Mwanzia, V.M.; Musili, P.M.; Wang, Q.F.; Chen, J.M. The past, current, and future distribution modeling of four water lilies (Nymphaea) in Africa indicates varying suitable habitats and distribution in climate change. Aquat. Bot. 2021, 173, 103416. [Google Scholar] [CrossRef]
- Ngarega, B.K.; Nzei, J.M.; Saina, J.K.; Halmy, M.W.A.; Chen, J.M.; Li, Z.Z. Mapping the habitat suitability of Ottelia species in African. Plan Divers. 2021. [Google Scholar] [CrossRef]
- Finlayson, C.M.; Lowry, J.; Bellio, M.G.; Nou, S.; Pidgeon, R.; Walden, D.; Humphrey, C.; Fox, G. Biodiversity of the wetlands of the Kakadu Region, northern Australia. Aquat. Sci. 2006, 68, 374–399. [Google Scholar] [CrossRef]
- Karadada, J.; Karadada, L.; Goonack, W.; Mangolamara, G.; Bunjuck, W.; Karadada, L.; Djanghara, B.; Mangolamara, S.; Oobagooma, J.; Charles, A.; et al. Uunguu Plants and Animals, Aboriginal Biological Knowledge from Wunambal Gaambera Country in the North-West Kimberley, Australia; Northern Territory Botanical Bulletin No. 35; Department of Natural Resources Aboriginal Knowledge: Plants and Animals:: Kimberley, Australia, 2011. [Google Scholar]
- Jackson, S.T.; Webb, R.S.; Anderson, K.H.; Overpeck, J.T.; Webb, T., III; Williams, J.W.; Hansen, B.C. Vegetation and environment in eastern North America during the last glacial maximum. Quat. Sci. Rev. 2000, 19, 489–508. [Google Scholar] [CrossRef]
- Hobday, A.J.; Lough, J.M. Projected climate change in Australian marine and freshwater environments. Mar. Freshw. Res. 2011, 62, 1000–1014. [Google Scholar] [CrossRef] [Green Version]
- Morrongiello, J.R.; Beatty, S.J.; Bennett, J.C.; Crook, D.A.; Ikedife, D.N.; Kennard, M.J.; Kerezsy, A.; Lintermans, M.; McNeil, D.G.; Pusey, B.J.; et al. Climate change and its implications for Australia’s freshwater fish. Mar. Freshw. Res. 2011, 62, 1082–1098. [Google Scholar] [CrossRef] [Green Version]
- Eliot, I.; Finlayson, C.; Waterman, P. Predicted climate change, sea-level rise and wetland management in the Australian wet-dry tropics. Wetl. Ecol. Manag. 1999, 7, 63–81. [Google Scholar] [CrossRef]
- Pusey, B.; Kennard, M.J.; Arthington, A.H. Freshwater Fishes of North-Eastern Australia; CSIRO Publishing: Clayton, Australia, 2004. [Google Scholar]
- Hamilton, S.K. Biogeochemical implications of climate change for tropical rivers and floodplains. Hydrobiologia 2010, 657, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.F.; Timbal, B. A review of recent climate variability and climate change in southeastern Australia. Int. J. Climatol. A J. R. Meteorol. Soc. 2008, 28, 859–879. [Google Scholar] [CrossRef]
- Suppiah, R.; Hennessy, K.; Whetton, P.; McInnes, K.; Macadam, I.; Bathols, J.; Ricketts, J.; Page, C. Australian climate change projections derived from simulations performed for the IPCC 4th Assessment Report. Aust. Meteorol. Mag. 2007, 56, 131–152. [Google Scholar]
- Löhne, C.; Wiersema, J.H.; Borsch, T. The unusual Ondinea, actually just another Australian water-lily of Nymphaea subg. Anecphya (Nymphaeaceae). Willdenowia 2009, 39, 55–58. [Google Scholar] [CrossRef]
- Bennett, J.; Ling, F.; Graham, B.; Grose, M.; Corney, S.; White, C.; Holz, G.; Post, D.; Gaynor, S.; Bindoff, N. Climate Futures for Tasmania: Water and Catchments Technical Report. 2010. Available online: http://www.climatechange.tas.gov.au/government_action/climate_futures (accessed on 17 March 2022).
- Araújo, M.B.; Rahbek, C. How does climate change affect biodiversity? Science 2006, 313, 1396–1397. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.r-project.org (accessed on 17 March 2022).
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. A J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The community climate system model version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E. The Global 2002: Priority ecoregions for global conservation. Ann. Mo. Bot. Gard. 2002, 89, 199–224. [Google Scholar] [CrossRef]
- O’brien, R.M. A caution regarding rules of thumb for variance inflation factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Bioclimatic Variables | |||||||
|---|---|---|---|---|---|---|---|---|
| bio1 | bio2 | bio3 | bio5 | bio10 | bio13 | bio14 | bio15 | |
| N. atrans | 1.650 | 1.426 | 1.442 | - | - | - | 1.975 | 1.873 |
| N. carpentariae | - | 1.687 | - | - | - | - | 2.555 | 2.048 |
| N. elleniae | 1.616 | 1.405 | 1.442 | - | - | - | 1.973 | 1.948 |
| N. georginae | 1.706 | 1.505 | 1.440 | - | - | - | 2.165 | 1.831 |
| N. gigantea | - | 2.029 | 1.186 | 2.032 | - | - | - | - |
| N. hastifolia | 1.839 | 1.331 | 1.348 | - | - | - | 1.818 | 1.961 |
| N. immutabilis | - | 1.556 | - | - | - | - | 2.371 | 1.947 |
| N. jacobsii | - | 2.647 | - | - | - | 2.584 | 2.272 | - |
| N. macrosperma | - | 2.140 | - | - | - | 1.910 | 1.809 | - |
| N. ondinea | 2.326 | 2.297 | 1.620 | - | - | 6.540 | 4.597 | 6.692 |
| N. violancea | - | 1.642 | 1.596 | - | - | - | 2.564 | 2.555 |
| N. nouchali | - | 1.902 | 1.128 | 1.859 | - | - | - | - |
| N. alba | - | 3.861 | 2.244 | - | 3.908 | 3.428 | - | 4.659 |
| N. pubescens | - | 2.515 | 3.739 | - | 3.514 | 2.843 | - | |
| Species | Features Class | rm Value | Current Habitat Suitability (Km2) |
|---|---|---|---|
| N. alba | LQ | 0.5 | 564,703.2154 |
| N. atrans | LQH | 1.5 | 288,466.6287 |
| N. carpentariae | LQHP | 1.5 | 814,252.6658 |
| N. elleniae | LQHP | 3 | 206,789.3887 |
| N. georginae | LQH | 1 | 192,787.1066 |
| N. gigantea | LQHP | 0.5 | 1,303,516.112 |
| N. hastifolia | LQHP | 1.5 | 199,145.3796 |
| N. immutabilis | LQH | 1 | 948,548.1007 |
| N. jacobsii | LQH | 2.5 | 142,273.8545 |
| N. macrosperma | LQH | 0.5 | 427,824.8378 |
| N. nouchali | LQHP | 0.5 | 453,647.2079 |
| N. ondinea | LQH | 1 | 120,521.2861 |
| N. pubescens | LQHPT | 1 | 162,148.2411 |
| N. violancea | LQHP | 0.5 | 1,137,450.324 |
| Species | Current | RCP 4.5 (2050) | RCP 4.5 (2070) | RCP 8.5 (2050) | RCP 8.5 (2070) | LGM | MH |
|---|---|---|---|---|---|---|---|
| N. alba | 0.876 (0.030) | 0.850 (0.026) | 0.875 (0.030) | 0.887 (0.043) | 0.844 (0.049) | 0.849 (0.032) | 0.897 (0.021) |
| N. atrans | 0.852 (0.133) | 0.859 (0.063) | 0.903 (0.061) | 0.870 (0.081) | 0.906 (0.069) | 0.912 (0.041) | 0.876 (0.075) |
| N. carpentariae | 0.897 (0.060) | 0.907 (0.033) | 0.888 (0.054) | 0.880 (0.047) | 0.918 (0.045) | 0.888 (0.043) | 0.897 (0.020) |
| N. elleniae | 0.858 (0.053) | 0.887 (0.053) | 0.878 (0.056) | 0.899 (0.057) | 0.894 (0.051) | 0.892 (0.056) | 0.873 (0.062) |
| N. georginae | 0.933 (0.060) | 0.929 (0.071) | 0.919 (0.065) | 0.952 (0.063) | 0.923 (0.094) | 0.949 (0.051) | 0.929 (0.066) |
| N. gigantea | 0.829 (0.017) | 0.843 (0.020) | 0.849 (0.030) | 0.845 (0.027) | 0.857 (0.035) | 0.864 (0.016) | 0.838 (0.016) |
| N. hastifolia | 0.850 (0.058) | 0.863 (0.069) | 0.890 (0.045) | 0.871 (0.047) | 0.853 (0.071) | 0.824 (0.189) | 0.876 (0.069) |
| N. immutabilis | 0.883 (0.022) | 0.906 (0.011) | 0.882 (0.023) | 0.898 (0.016) | 0.893 (0.033) | 0.875 (0.019) | 0.903 (0.025) |
| N. jacobsii | 0.930 (0.049) | 0.936 (0.032) | 0.938 (0.055) | 0.947 (0.031) | 0.947 (0.048) | 0.912 (0.035) | 0.919 (0.035) |
| N. macrosperma | 0.899 (0.022) | 0.890 (0.021) | 0.902 (0.023) | 0.907 (0.031) | 0.902 (0.020) | 0.869 (0.023) | 0.908 (0.020) |
| N. nouchali | 0.959 (0.005) | 0.956 (0.013) | 0.956 (0.012) | 0.959 (0.011) | 0.963 (0.010) | 0.945 (0.014) | 0.955 (0.012) |
| N. ondinea | 0.901 (0.065) | 0.913 (0.053) | 0.946 (0.047) | 0.934 (0.058) | 0.950 (0.045) | 0.948 (0.040) | 0.912 (0.043) |
| N. pubescens | 0.962 (0.020) | 0.942 (0.019) | 0.960 (0.012) | 0.948 (0.015) | 0.959 (0.006) | 0.941 (0.030) | 0.929 (0.018) |
| N. violancea | 0.896 (0.009) | 0.903 (0.009) | 0.908 (0.006) | 0.901 (0.008) | 0.907 (0.006) | 0.884 (0.007) | 0.900 (0.008) |
| Species | Current | LGM | MH | RCP 4.5 (2050) | RCP 8.5 (2050) | RCP 4.5 (2070) | RCP 8.5 (2070) |
|---|---|---|---|---|---|---|---|
| N. alba | 0.3045 | 0.4618 | 0.2835 | 0.3045 | 0.3209 | 0.2957 | 0.2864 |
| N. atrans | 0.3649 | 0.5166 | 0.4495 | 0.3649 | 0.3931 | 0.3851 | 0.3282 |
| N. carpentariae | 0.2657 | 0.2762 | 0.3093 | 0.2657 | 0.2427 | 0.2323 | 0.267 |
| N. elleniae | 0.427 | 0.4036 | 0.3695 | 0.427 | 0.3686 | 0.4071 | 0.3776 |
| N. georginae | 0.4203 | 0.4576 | 0.3858 | 0.4203 | 0.3524 | 0.3354 | 0.4671 |
| N. gigantea | 0.3244 | 0.2812 | 0.3083 | 0.3244 | 0.2374 | 0.2307 | 0.2477 |
| N. hastifolia | 0.5105 | 0.3385 | 0.476 | 0.5105 | 0.4554 | 0.4571 | 0.41 |
| N. immutabilis | 0.263 | 0.2779 | 0.2242 | 0.263 | 0.2589 | 0.1886 | 0.2051 |
| N. jacobsii | 0.5673 | 0.597 | 0.5929 | 0.5673 | 0.5944 | 0.6119 | 0.5631 |
| N. macrosperma | 0.1733 | 0.1669 | 0.1807 | 0.1733 | 0.1618 | 0.1857 | 0.1686 |
| N. nouchali | 0.145 | 0.1523 | 0.1305 | 0.145 | 0.129 | 0.1268 | 0.1283 |
| N. ondinea | 0.4065 | 0.4328 | 0.3101 | 0.4065 | 0.3521 | 0.3685 | 0.4189 |
| N. pubescens | 0.1814 | 0.349 | 0.2098 | 0.1814 | 0.2221 | 0.2264 | 0.229 |
| N. violancea | 0.2178 | 0.2379 | 0.1992 | 0.2178 | 0.1725 | 0.1666 | 0.1933 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nzei, J.M.; Mwanzia, V.M.; Ngarega, B.K.; Musili, P.M.; Wang, Q.-F.; Chen, J.-M.; Li, Z.-Z. Ecological Niche Modeling of Water Lily (Nymphaea L.) Species in Australia under Climate Change to Ascertain Habitat Suitability for Conservation Measures. Plants 2022, 11, 1874. https://doi.org/10.3390/plants11141874
Nzei JM, Mwanzia VM, Ngarega BK, Musili PM, Wang Q-F, Chen J-M, Li Z-Z. Ecological Niche Modeling of Water Lily (Nymphaea L.) Species in Australia under Climate Change to Ascertain Habitat Suitability for Conservation Measures. Plants. 2022; 11(14):1874. https://doi.org/10.3390/plants11141874
Chicago/Turabian StyleNzei, John M., Virginia M. Mwanzia, Boniface K. Ngarega, Paul M. Musili, Qing-Feng Wang, Jin-Ming Chen, and Zhi-Zhong Li. 2022. "Ecological Niche Modeling of Water Lily (Nymphaea L.) Species in Australia under Climate Change to Ascertain Habitat Suitability for Conservation Measures" Plants 11, no. 14: 1874. https://doi.org/10.3390/plants11141874
APA StyleNzei, J. M., Mwanzia, V. M., Ngarega, B. K., Musili, P. M., Wang, Q.-F., Chen, J.-M., & Li, Z.-Z. (2022). Ecological Niche Modeling of Water Lily (Nymphaea L.) Species in Australia under Climate Change to Ascertain Habitat Suitability for Conservation Measures. Plants, 11(14), 1874. https://doi.org/10.3390/plants11141874