Abstract
Herbicides are the most commonly used means of controlling the growth of Ambrosia artemisiifolia L. Their constant use has led to the development of resistant populations. They can be evaluated by studying seed germination and the corresponding grown plants, but A. artemisiifolia exhibits seed dormancy, preventing germination and delaying research. Here, we developed a simple and rapid method to stimulate germination of freshly collected or stored A. artemisiifolia seeds. The germination of A. artemisiifolia freshly collected/stored seeds was evaluated after storage, stratification, and chemical treatments (ethephon, gibberellic acid (GA3), thiourea, KNO3). Ethephon or ethephon + GA3 improved freshly collected seed germination by 88 and 95%, respectively, and germination of stored seeds by 78 and 80%, respectively. In addition, placing the seeds of A. artemisiifolia in ethephon, GA3, ethephon + GA3, or thiourea solutions caused the freshly collected seeds to germinate faster than stored seeds or nontreated seeds. In contrast, the conditioning of seeds in these solutions favored germination of stored seeds, especially when ethephon + GA3 or GA3 was used. Imbibition of the freshly collected A. artemisiifolia seeds in a mixture of ethephon and GA3 can effectively overcome primary dormancy when rapid experimental results are needed. For seeds requiring prolonged storage, conditioning in ethephon, GA3, or thiourea solutions may be applied to promote germination.
1. Introduction
Ambrosia artemisiifolia L. (common ragweed) is a highly competitive summer annual weed that originated from North America and occurs in all arable crops that are sown or planted in mid-spring [1]. Its wide range of tolerated conditions and high germination rates across a broad spectrum of temperatures [2], photoperiods, pH, and soil salinity levels, as well as its high variability, must be considered crucial conditions for its rapid spread [3], which poses a serious threat to public health [4]. Extensive research in mainland Croatia has shown that A. artemisiifolia is the most common broadleaf weed among arable crops [5]. Herbicides are usually used to control its growth, but their constant use has led to resistant biotypes [6].
The efficacy of herbicides and the development of herbicide-resistant populations can be evaluated by studying germinated seeds and the corresponding grown plants. However, freshly collected A. artemisiifolia seeds exhibit primary dormancy, also known as endogenous (physiological) dormancy [7], which can inhibit germination for more than six months, thus prolonging the time that is required for plant growth in greenhouse experiments [8]. Given that seed dormancy release occurs naturally at low temperatures and in humid conditions during the winter, freshly collected seeds can be stratified at 4 °C in moist sand. However, this method is time-consuming, requiring up to 12 weeks to promote germination [9].
Stratified A. artemisiifolia seeds germinate when the balance changes between the growth-promoting hormone gibberellic acid (GA3) and the growth-inhibiting hormone abscisic acid [7,9,10,11]. Ethylene also promotes seed germination of many species by reducing the sensitivity of the seeds to endogenous abscisic acid [12]. Therefore, plant growth hormones or their combination have been used to shorten primary dormancy as well as to initiate germination and plant growth. In particular, ethephon (synthetic form of ethylene in liquid state), GA3, and kinetin (cytokinin) have been used to release primary and secondary dormancy in A. artemisiifolia seeds, but a single hormone application does not significantly stimulate germination. In contrast, the combination of ethylene and gibberellin significantly promotes the germination of some weed species seeds [13,14,15]. The germination of A. trifida seeds can also be significantly promoted using chemical agents instead of water, but seedlings with abnormal growth occasionally develop [16].
Other methods have been used to release the dormancy of various Ambrosia species, including nitrate fertilizers, mechanical scarification, and embryo extraction [9,16,17]. Among them, embryo excision from A. trifida seeds may be the most effective method, but it is laborious and, therefore, unsuitable for large numbers of seeds. To our knowledge, embryo excision has not yet been tested on A. artemisiifolia seeds.
Growth-promoting hormones have been used to break dormancy in R. crispus seeds [15]. However, the effects of dormancy-breaking growth-promoting hormones or chemicals on their germination and normal plant development have not yet been reported. Therefore, in this study we examined the response of freshly collected and stored A. artemisiifolia seeds to dormancy-breaking methods, with the aim of developing a simple and time-efficient method for stimulating the germination of A. artemisiifolia seeds. The effective stimulation of germination in dormant A. artemisiifolia seeds would help support future investigations into the effect of herbicides on A. artemisiifolia and consequently into overcoming herbicide resistance.
Freshly collected seeds exhibit primary dormancy [7], while prolonged seed storage can lead to seed senescence (loss of vigor) and plant death [18]. Laboratory experiments were first performed to investigate the effects of seed storage, cold stratification, chemical treatments, and their combination on the germination rate and germination dynamics of both seed types. The most effective dormancy-breaking methods that were identified in the laboratory experiments were then applied under greenhouse conditions to detect whether they give rise to growth abnormalities during the emergence and early growth of A. artemisiifolia seeds.
2. Results
2.1. Laboratory Experiments
The laboratory experiments indicated significant differences in the germination of freshly collected and cold-stored A. artemisiifolia seeds depending on the duration of cold storage, stratification time, or type of chemical treatments (Table 1). A significant difference was found for the adjusted means of the two factors that were studied, freshly collected/cold-stored seeds and treatments, and for the interaction between these factors.

Table 1.
Analysis of variance for the effect of dormancy-breaking methods on Ambrosia artemisiifolia seeds.
2.1.1. Seed Storage
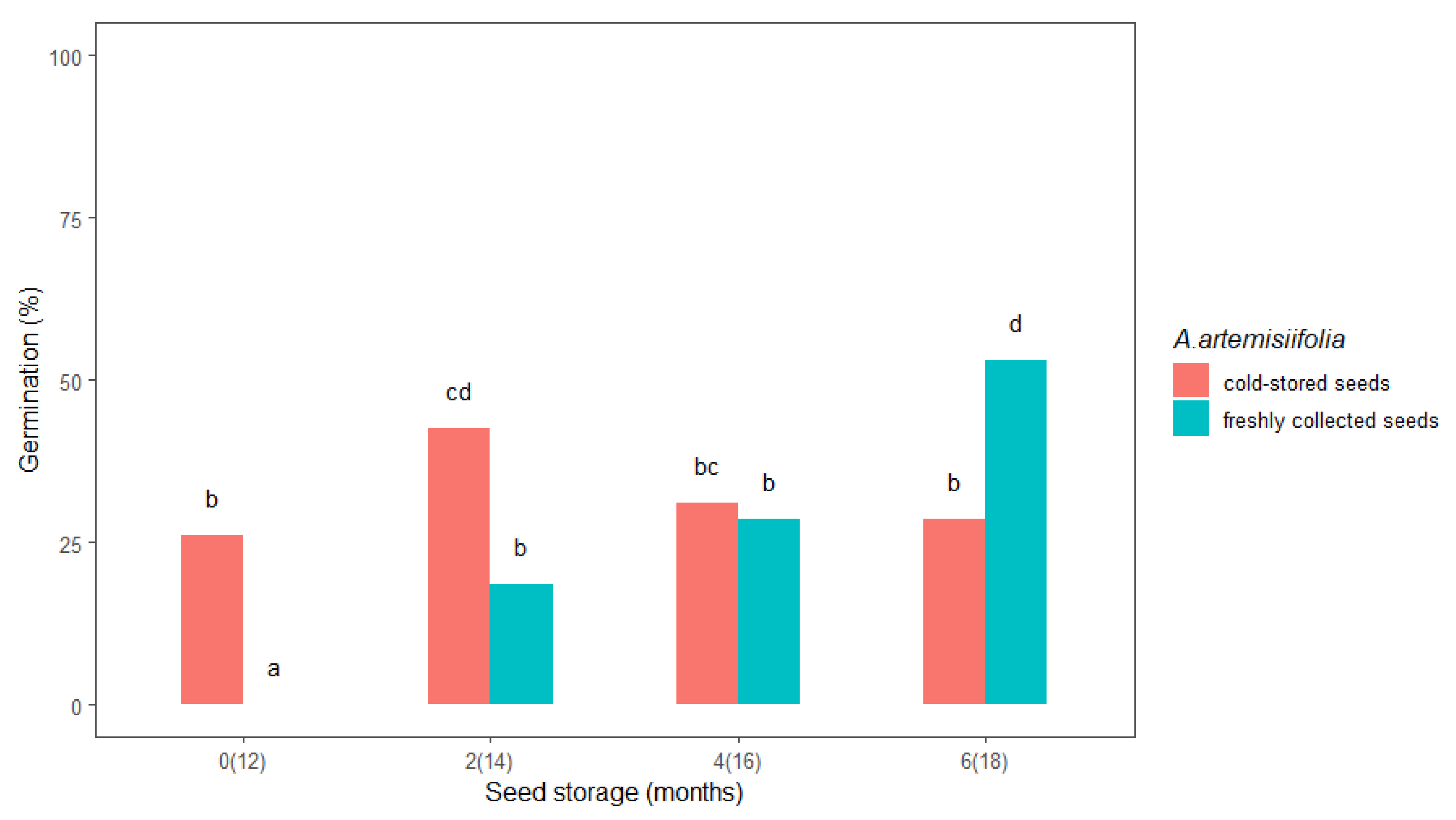
Prolonged cold storage seemed to favor the A. artemisiifolia seeds that were already in cold storage compared to the freshly collected seeds. A significant difference in the germination percentage was observed between freshly collected and cold-stored seeds in the first test (0, i.e., 12 months) and after two (14) and six (18) months of storage (Figure 1). The freshly collected seeds did not germinate in the first test, while the cold-stored seeds had a germination rate of 26%, and after two months of storage, the freshly collected and cold-stored seeds germinated at 18.5% and 42.5%, respectively. In general, the germination rate after zero, two, and four months was higher for the cold-stored seeds. Interestingly, germination of the freshly collected seeds increased from 0% after zero months of storage to 53% after six months, while germination of the cold-stored seeds decreased after four and six months. In addition, germination of the cold-stored seeds after six months of storage was almost half (28.5%) that of the freshly collected seeds after six months of storage.
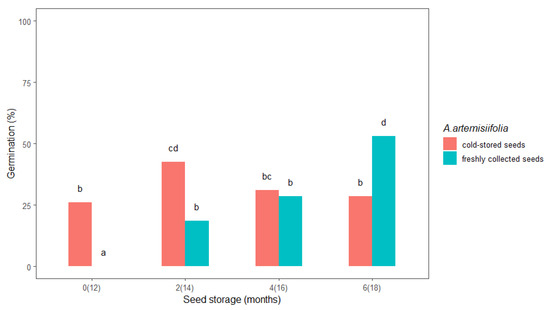
Figure 1.
The effect of storage duration on freshly collected and cold-stored Ambrosia artemisiifolia seeds. The estimated marginal means of eight replicates are shown (standard error = 3.1). The means that are labelled with the same letter are not significantly different (p < 0.05) according to Tukey’s test. Numbers on the x-axis indicate the months of seed storage for the freshly collected seeds and the total months of storage in parentheses for the cold-stored seeds.
2.1.2. Cold Stratification
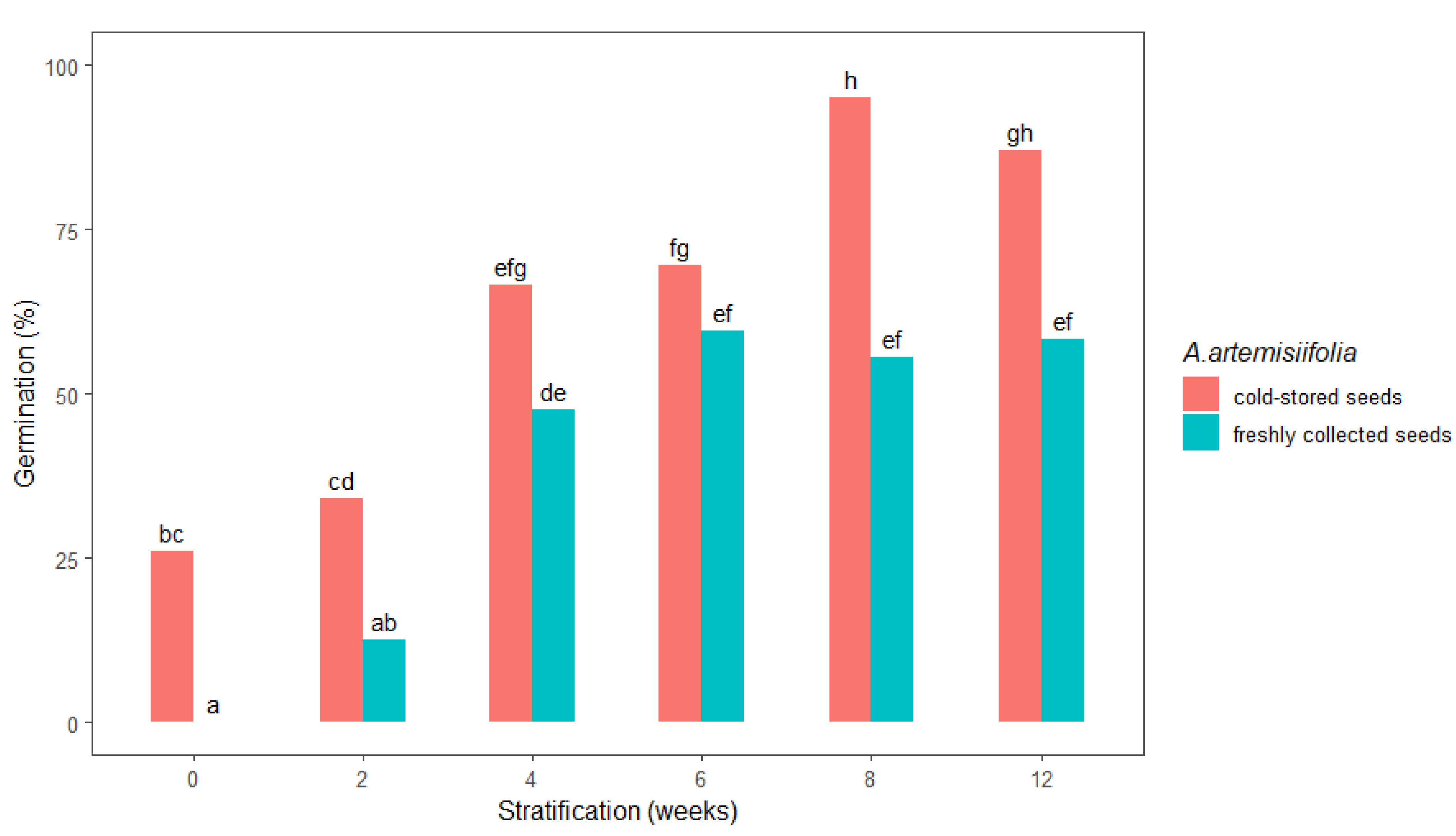
The germination of stratified A. artemisiifolia seeds was highly dependent on the duration of the stratification period and the studied freshly collected or cold-stored seeds (Figure 2). In particular, no germination occurred in the unstratified freshly collected seeds, whereas their germination was promoted as the stratification period increased. Stratification for shorter than four weeks did not satisfactorily break the seed dormancy (Figure 2), but germination of about 55% was achieved after 6, 8, and 12 weeks of stratification, respectively. In contrast, the germination percentage of unstratified cold-stored A. artemisiifolia seeds (26%) was significantly higher than that of the unstratified freshly collected seeds (0%). Increasing the duration of stratification promoted germination of the cold-stored seeds, with significantly higher germination rates after 8 and 12 weeks of stratification of 95% and 87% recorded, respectively. Statistical analysis further confirmed that there was no significant difference in the germination of the freshly collected seeds that were stratified for 4, 6, 8, or 12 weeks (Figure 2).
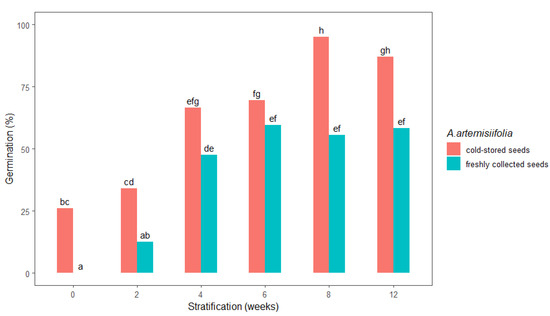
Figure 2.
Total germination 14 days after sowing of freshly collected or cold-stored Ambrosia artemisiifolia seeds at different stratification intervals (0, 2, 4, 6, 8, and 12 weeks). The estimated marginal means of eight replicates are shown (standard error = 4.49). Means following the same letter are not significantly different (p < 0.05) according to Tukey’s test.
2.1.3. Chemical Treatments
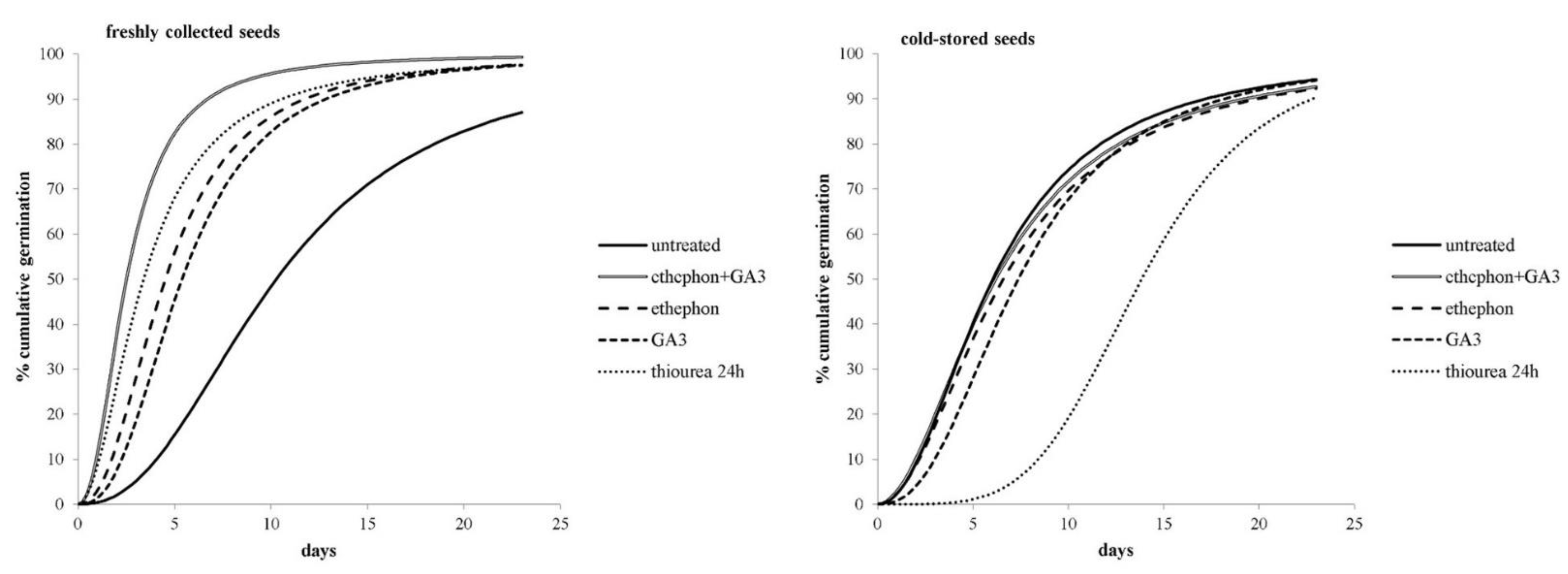
Freshly collected or cold-stored A. artemisiifolia seeds were also treated with various growth-promoting chemicals, and the effects on germination were determined. Although cumulative seed germination was significantly affected by the type of chemical treatment that was used (Figure 3), it showed a stronger dependence on the time of seed collection and storage (Figure 4).
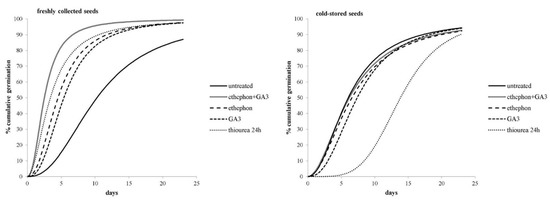
Figure 3.
Cumulative germination over 14 days of the freshly collected or cold-stored seeds of A. artemisiifolia that were treated or not treated with chemical treatments. The lines represent logistic models that were generated with the statistical program Bioassay (97).
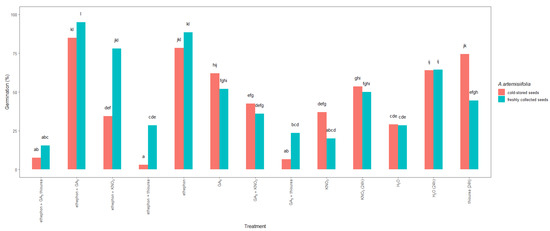
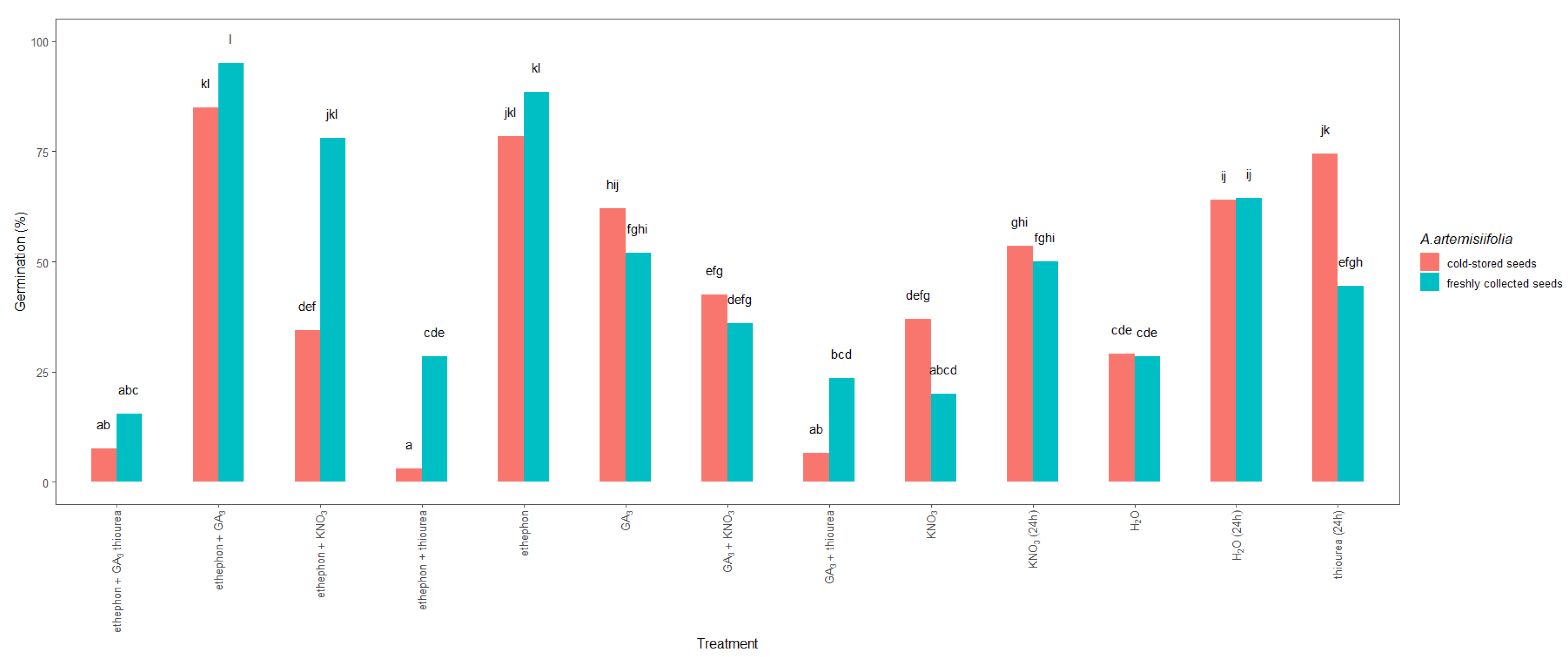
Figure 4.
Total germination 14 days after sowing of freshly collected or cold-stored Ambrosia artemisiifolia seeds that were imbedded or treated with growth-promoting hormones or fertilizers. The estimated marginal means of eight replicates (standard error = 3.35) are shown. According to Tukey’s test, means following the same letter are not significantly different (p < 0.05). No germination was observed in seeds that were treated with thiourea + KNO3 or GA3 + thiourea + KNO3; data not shown.
In general, the germination of nontreated freshly collected seeds started an average of two days earlier, and reached 50% at 4.2 days earlier, than the cold-stored seeds (Figure 3). Further treatment of freshly collected seeds with ethephon, GA3, ethephon + GA3, or thiourea significantly accelerated their germination compared to the nontreated seeds. For example, nontreated seeds reached 50% germination after 10.4 days, while the chemically-treated seeds required only 2.5–5.4 days (Figure 3). Conversely, the cold-stored seeds that were imbedded in thiourea took more than twice as long as the nontreated seeds to achieve 10% and 50% germination. Similar results were also found for GA3-treated cold-stored seeds, whereas the germination dynamic of cold-stored seeds that were treated with ethephon or ethephon + GA3 was similar to that of the untreated seeds.
Significant differences were also observed between the studied seeds (Figure 4). In particular, the germination of the freshly collected seeds that were treated with ethephon + KNO3 reached nearly a maximum, 78%, whereas a germination of only 34% was achieved in the similarly treated cold-stored seeds. In contrast, imbibition in thiourea resulted in 74% germination of the cold-stored seeds but only 44% germination of the freshly collected seeds. Moreover, when ethephon or ethephon + GA3 was used, the germination of the cold-stored and freshly collected A. artemisiifolia seeds improved to 78–95%, with the freshly collected seeds showing higher germination than the cold-stored seeds. In contrast, the application of nitrogen fertilizers or their combination with hormones resulted in significantly lower germination compared to other treatments, and a significant interaction was observed between the type of treatment and the seed collection time and storage (Table 1). Hormones and fertilizers were also used as a tank mix (ethephon + thiourea, ethephon + GA3 + thiourea, or GA3 + thiourea), but an antagonistic effect was observed, leading to low germination in both the freshly collected and cold-stored seeds.
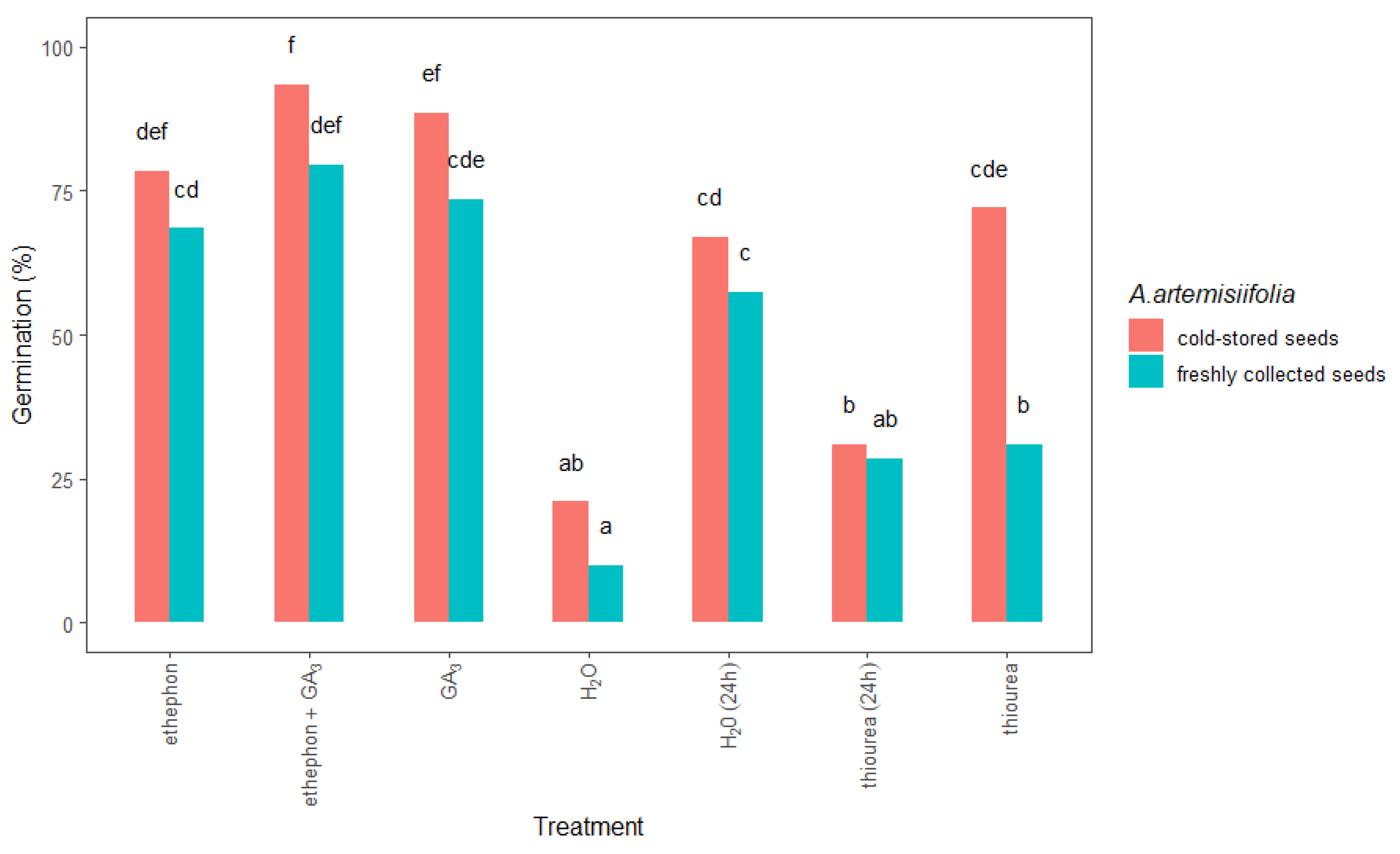
Furthermore, conditioning in chemical solutions significantly improved the germination of the freshly collected and cold-stored A. artemisiifolia seeds compared to both types of nontreated seeds (Figure 5). In all cases, this process was more effective on the cold-stored than on the freshly collected seeds, especially when the seeds were conditioned with ethephon + GA3 (93% cold-stored vs. 79% freshly collected) or GA3 (88% vs. 73%). The application of ethephon alone also promoted the germination of cold-stored and freshly collected seeds by 78% and 68%, respectively. Interestingly, the germination of the cold-stored seeds was significantly higher (72%) than that of the freshly collected seeds (34%) after conditioning with thiourea.
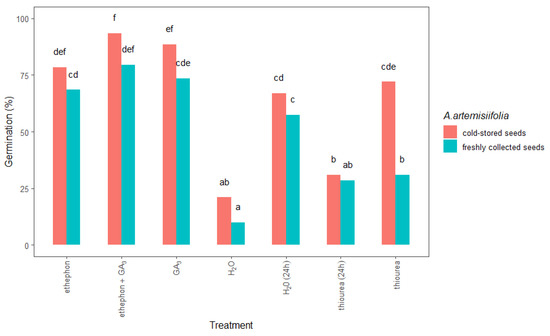
Figure 5.
Total germination at 14 days after sowing freshly collected or cold-stored Ambrosia artemisiifolia seeds that were conditioned in water or chemical solution. The estimated marginal means of eight replicates (standard error = 3.35) are shown. According to Tukey’s test, the means following the same letter are not significantly different (p < 0.05). No germination was observed in seeds that were treated with thiourea + KNO3 or GA3 + thiourea + KNO3; data not shown.
2.2. Greenhouse Experiment
The laboratory experiments indicated that imbibition or conditioning in ethephon, GA3, ethephon + GA3, or thiourea were the most effective dormancy-breaking methods for A. artemisiifolia seeds. Therefore, these treatments were further applied in a greenhouse experiment to examine their effect on the emergence and development of A. artemisiifolia plants.
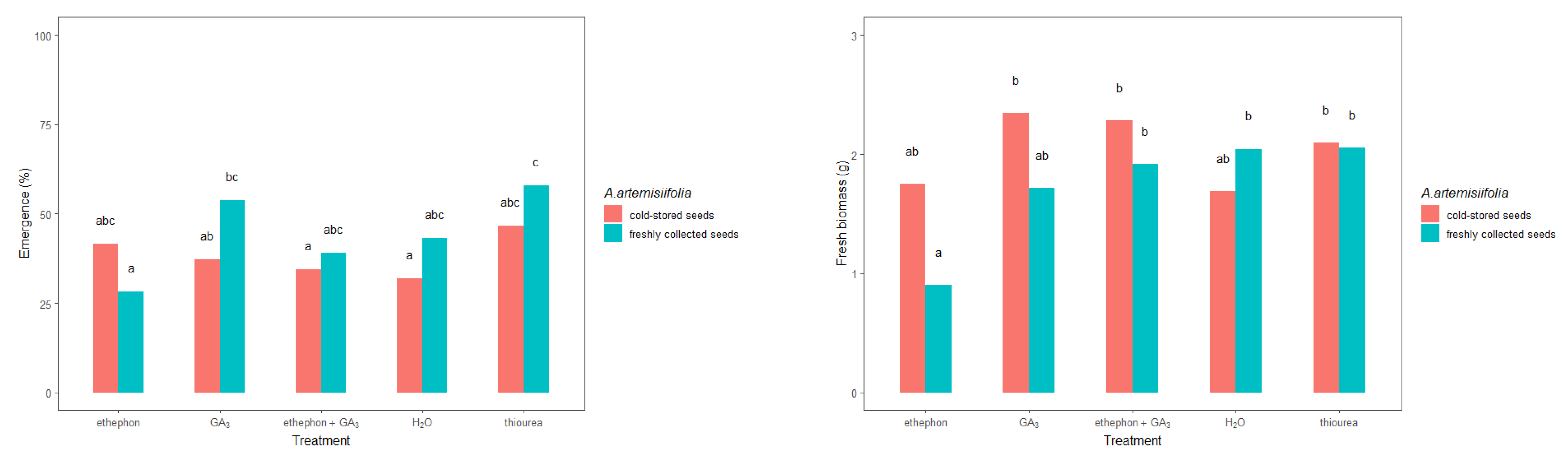
The freshly collected and cold-stored seeds responded differently to the applied treatments, consistent with a treatment–seed interaction (Table 2). In general, the freshly collected A. artemisiifolia seeds emerged in greater numbers than the cold-stored seeds in most cases (Figure 6). However, a significant difference was observed only in the freshly collected seeds that were treated with ethephon + GA3 and thiourea. Moreover, only these two treatments resulted in emergence that was greater than 50%: 53.7% for ethephon + GA3 and 57.8% for thiourea.
Table 2.
Analysis of variance of hormones and nitrogen fertilizers treatments on the emergence, hypocotyl, and fresh biomass of A. artemisiifolia seedlings.
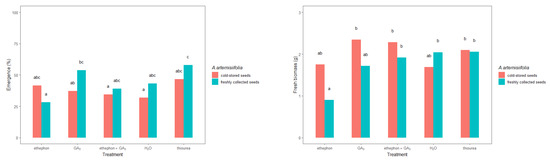
Figure 6.
Initial growth parameters of young Ambrosia artemisiifolia plants that were grown from freshly collected or cold-stored seeds under various chemical treatments or conditioning in the greenhouse. The estimated marginal means of 16 replicates are shown for the freshly collected seeds (standard error = 4.19) and cold-stored seeds (standard error = 0.201). According to Tukey’s test, means following the same letter are not significantly different (p < 0.05).
The fresh biomass of A. artemisiifolia differed depending on the treatment or seed. In particular, freshly collected seeds that were treated with GA3 or conditioned in thiourea developed heavier seedlings (2.38 g and 2.52 g, respectively) than the nontreated seeds (1.89 g). Similar differences were also observed in the cold-stored seeds that were treated with ethephon (2.23 g), ethephon + GA3 (2.89 g) or GA3 (2.98), or conditioned in thiourea (2.63 g). In contrast, no significant differences were observed in the heights of plants that developed from the freshly collected and cold-stored seeds, except for freshly collected seeds that were conditioned in ethephon (5.07 cm) and cold-stored seeds that were conditioned in water (7.80 cm), which developed significantly lower plants than the respective nontreated seeds (9.03 and 8.60 cm, respectively). None of the other treatments led to significant differences in plant height.
Growth-promoting chemicals also led to abnormalities in the growth of A. artemisiifolia. For example, internodes formed between the seedling and the first pair of true leaves on seven plants that were grown from ethephon + GA3-treated cold-stored seeds. In addition, small deformations were observed on the first leaves of four plants that were grown from seeds that were imbedded or conditioned with GA3 or ethephon + GA3. In contrast, no deformations were detected in plants that derived from seeds that were imbedded or conditioned in thiourea (Figure 7).

Figure 7.
Ambrosia artemisiifolia young plants showing slight deformations upon chemical treatments.
3. Discussion
In this study, we investigated the ability of growth-promoting chemicals to break the dormancy of freshly collected or cold-stored A. artemisiifolia seeds. Although numerous studies have reported different germination patterns between different populations, probably due to their different dormancy levels [8] or genetic variability [19,20], the present study may be the first report on the susceptibility of A. artemisiifolia seeds to dormancy-breaking methods.
Our results indicate that the germination of A. artemisiifolia seeds depends significantly on the seed age. Specifically, only 26% of seeds that were stored at 4 °C for 12 months germinated (Figure 1), while further extension of the cold storage for six months did not improve their germination. This result is inconsistent with earlier studies, where the storage of A. artemisiifolia seeds at 4 or 5 °C for extended periods was found to break dormancy [21]. In contrast, 53% of the freshly collected seeds in our study germinated after six months of cold storage (Figure 1), implying that the seed age may limit germination due to the loss of viability [16,22,23]. Our findings were confirmed by the tetrazolium (2,3,5-tryphenyl tetrazolium chloride) test, where the cold-stored seeds showed lower viability than the freshly collected seeds (data not shown). Moreover, 50% and 90% germination occurred more than 4 and 8.6 days earlier, respectively, with the freshly collected seeds than with the cold-stored seeds (Figure 3), suggesting that germinating seeds should be obtained quite soon after harvest and that an efficient method should be developed to overcome dormancy in freshly collected seeds.
The stimulating effect of ethephon, GA3, thiourea, or KNO3 on the germination of Ambrosia trifida seeds has already been reported [17]. The present study extends the literature by showing that direct application or conditioning of A. artemisiifolia seeds with these chemicals has different effects on seed germination depending on the type of growth-promoting agents and the seed storage conditions. In particular, the application of ethephon or ethephon + GA3 either directly to the Petri dishes or as seed conditioning promoted the germination of A. artemisiifolia seeds more than the other treatments, and the effect was greater on the freshly collected seeds than on the cold-stored ones. Conversely, conditioning in a chemical solution (GA3 or thiourea) improved germination of the cold-stored seeds more than that of the freshly collected seeds (Figure 4), as already reported for A. trifida that were conditioned in water or chemical solution [16]. We also found that hormones (ethephon and GA3) were more effective than nitrogen fertilizers (KNO3 and thiourea) on the germination of freshly collected A. artemisiifolia seeds (Figure 4). Although nitrogen fertilizers can break the inhibition of seed germination, they cannot directly break dormancy, which makes them more effective at lower dormancy levels [24]. This difference between the chemical agents could explain the higher germination of the cold-stored seeds compared to the freshly collected seeds after imbibition in KNO3 or thiourea (Figure 4). In contrast, growth-promoting hormones had a better effect on the freshly collected A. artemisiifolia seeds, as the exogenous application of ethephon + GA3 or ethephon stimulated seed germination by 95% and 88%, respectively. These results are consistent with previous studies, where the imbibition or conditioning of cold-stored A. trifida seeds with GA3 did not improve germination compared to treatment with water [16]. Using higher GA3 concentrations is unlikely to help, since it may exacerbate prolonged internode growth [25]. As an alternative approach, GA3 has been used in a tank mixture with ethephon or kinetin, which moderately promoted germination [15]. In the present study, freshly collected or cold-stored A. artemisiifolia seeds were conditioned in a GA3 solution, which led to 88% and 73% germination, respectively, probably due to the leaching of these compounds during conditioning [16]. Although using sandpaper to release embryos from seeds requires time and skill, we found that it can significantly promote the germination of A. artemisiifolia seeds when GA3 is used for conditioning (Figure 5). However, adding other chemicals (ethephon + GA3 or thiourea) to the solution did not lead to greater germination than when seeds were exposed to the same chemicals by imbibition or exogenous application. Similar results have been reported for A. trifida, where seeds that were conditioned in a chemical solution developed almost four times fewer germinated seeds than seeds that were conditioned in water [16]. Based on these results, we conclude that the seed conditioning process is favorable only for cold-stored seeds that are treated with GA3.
The use of growth-promoting chemicals accelerated the germination dynamics of only the freshly collected seeds (Figure 3). The lag phase from the beginning of incubation to the onset of germination was reduced to 2.1–3.3 days in seeds that ere imbedded with growth-promoting hormones and to 3.3 days in seeds that were imbedded with thiourea, while 50% germination was achieved about twice as fast as in the nontreated seeds. These results make growth-promoting hormones an attractive method to stimulate A. artemisiifolia germination without the need for stratification or a post-ripening period, as suggested earlier [16,17]. Seed stratification is a standard method of reducing physiological dormancy in A. artemisiifolia. However, we found that satisfactory germination can be achieved only if the seeds are exposed for at least 8 weeks to the stratification period, as reported in numerous previous studies [9,26,27,28,29,30]. Although the mechanism of stratification-induced release from dormancy is not known, germination may occur due to depletion of three silver-containing polypeptides during the stratification period [31] or possibly due to physical contact of the abrasive sand mixture with the seeds, weakening the embryo and thus stimulating germination [16]. However, in the present study, increasing the stratification period from 4 to 12 weeks did not promote the germination of freshly collected seeds that were maintained in a moist sand mixture (Figure 2).
The seedling stage of plants is the most vulnerable period in the life cycle of plants, as the growth of the seedlings directly affects the development and fitness of adult plants [32]. Therefore, here we investigated the emergence patterns and growth behavior of cold-stored and freshly collected A. artemisiifolia seeds that were imbedded or conditioned with growth-promoting hormones or thiourea. In most cases, the measured parameters (hypocotyl length, seedling weight, and height) showed negligible or only slightly significant differences between the cold-stored or freshly collected seeds. However, seeds that were conditioned in ethephon solution developed slightly longer hypocotyls or seedlings with lower heights and weights than the other treatments. Moreover, negligible deformations, i.e., development of internodes or deformation of first leaves, were observed in 3% of the plants that developed from ethephon or ethephon + GA3-treated seeds. Therefore, we conclude that the growth of A. artemisiifolia plants is not significantly affected when the seeds are treated with hormones or nitrogen fertilizers. However, since certain deformations were observed in the initial growth of A. artemisiifolia upon treatment with ethephon, the effect of these disturbances on further plant growth should be investigated in future studies, especially in resistance trials. Specifically, plants that are grown from ethephon-treated seeds should be treated with herbicides, and their response should be compared with plants that are emerging from nontreated seeds.
Our study clearly supports the theory that treatment with ethephon or ethephon + GA3 is the most effective method to overcome dormancy in A. artemisiifolia seeds when rapid experimental results are needed. Abnormal seedling growth was detected in A. trifida under the same conditions [16], but we did not observe such effects here. Prolonged cold storage should be avoided when the seed supply is limited because cold-stored seeds show lower viability than freshly collected ones. In the event that extensive cold storage is inevitable, A. artemisiifolia seeds can be conditioned in ethephon, GA3, or thiourea solution to promote germination. Nevertheless, the effects of these dormancy-breaking methods on plant growth should be further studied, especially after the use of herbicides.
4. Materials and Methods
4.1. Seed Collection and Storage
Mature seeds of A. artemisiifolia L. were collected in October 2018 and 2019 from plants that were grown in soybean fields in Požeška, Slavonia County, Badljevina (45°30′42″ N; 17°11′34″ E) and in Zagreb County, Topolje (45°42′36″ N; 16°20′24″ E). All of the seeds were collected from more than 20 grown A. artemisiifolia plants to adequately represent the genetic variability of a strictly allogamous species [33]. After each collection, the seeds were manually sorted by size and color, considering the relationship between size and germination [34] as well as their morphological and functional variability [35]. Only uniform seeds and those that were visually free of predator damage were used in the experiments. Seeds that were collected in 2018 were stored in a refrigerator at 4 °C for 12 months before treatment and are hereinafter referred to as “cold-stored” seeds. Seeds that were collected in 2019 were immediately used and are hereinafter referred to as “freshly collected” seeds. All of the seeds were subjected to continuous germination testing after 0, 2, 4, and 6 months of cold storage (i.e., 12, 14, 16, and 18 months of cold storage for the seeds that were designated as cold-stored seeds).
4.2. Laboratory Experiments
4.2.1. Cold Stratification
The freshly collected and cold-stored seeds were first sterilized with 3% hydrogen peroxide for 6 min to inactivate pathogens, and then placed in glass Petri dishes (150 mm diameter, 25 mm height) containing 100 g of sterilized quartz sand and 50 mL distilled water. The Petri dishes were subsequently sealed with Parafilm to prevent evaporation and stored at 4 °C for 0 (non-stratified seeds), 2, 4, 6, 8, and 12 weeks. At each stratification time point, the A. artemisiifolia seeds were extracted from the sand and subjected to a 14-day germination test.
4.2.2. Chemical Treatments
To assess the ability of chemical treatments to promote germination of freshly collected or cold-stored A. artemisiifolia seeds, nitrogen fertilizers (potassium nitrate (KNO3) and thiourea) and synthetic hormones (GA3 and ethephon) were applied as single treatments or in combination [16,17]. The seeds were either imbedded (24 h) or conditioned (48 h) in aqueous solutions of the growth promoters, or the aqueous solutions were added directly to the seed-containing Petri dishes (Table 3). For the treatment with nitrogen fertilizers, the seeds were imbedded in their aqueous solution for 24 h, or five of the aqueous solutions was added directly to the Petri dishes. In contrast, the hormone solutions were added directly to the Petri dishes.
Table 3.
Treatments that were applied to freshly collected or cold-stored Ambrosia artemisiifolia seeds in the laboratory and the greenhouse.
Seed conditioning was performed based on a previous study on A. trifida seeds [16], where the seed embryo was released by incising the seed crown. However, embryo incision could not be applied to the A. artemisiifolia seeds because the embryo is fused with the seed coat. Therefore, the seed coat was manually removed with sandpaper (P 40) and the embryo was released during the conditioning process. The scarified seeds were placed in an Erlenmeyer flask that was filled with 200 mL of deionized water or a chemical solution. A flexible hose that was attached to an air supply (Flamingo, Crawfish 1800, Limburg, The Netherlands), which touched the bottom of the surface, was then placed in an Erlenmeyer flask. The opening at the top of the flask was sealed with Parafilm to prevent evaporation and to stabilize the flask tube. In addition, air was distributed evenly through the tube to prevent water or aqueous solution from leaking and to ensure the continuous movement of the seeds in the flask. After 48 h, the A. artemisiifolia seeds were removed from the flask, and a germination test was performed.
4.2.3. Germination Tests
The germination tests were performed using a randomized complete block design with four replicates to determine the germination percentages of cold-stratified or chemically-treated seeds. The germination dynamics were assessed by counting the number of germinated seeds at 24 h intervals over a 14-day period [36].
The nontreated seeds were also tested after storage at 4 °C for 0, 2, 4, and 6 months (seed lot 2019) and 12, 14, 16, and 20 months (seed lot 2018) to evaluate germination over time without the application of dormancy-breaking methods.
For each germination test, 25 seeds were placed in sterile plastic Petri dishes (90 mm diameter) that were lined with filter paper (80 g m−2, 21/N, Munktell, Paul Marienfeld GmbH & Co. KG, Lauda-Königshofen, Germany) and 5 mL of distilled water or chemical solution was added. The Petri dishes were then sealed with Parafilm and placed in a climatic chamber (Memmert, UF 260, Schwabach, Germany) at a constant temperature of 25/15 °C with 70% humidity under standard photoperiod conditions (12 h:12 h) and a light intensity of 40–50 µmol m−2 s−1. The germinated seeds were counted daily for two weeks. Seed germination was defined as the development of 1 mm long radicles.
4.3. Greenhouse Experiment
Plant pots (12 cm diameter) were filled with a mixture of humus substrate (Potground H, Klasman, Geeste, Germany) and sterile soil in equal weight ratio (1:1). The soil, which was sampled from the Šašinovec Experimental Station at University of Zagreb (45°51′05.2″ N 68 16°10′34.1″ E), was classified as sandy clay with pH 7.74 (H2O), 7.04 (KCl), 4.22% humus, and 2.9% calcium carbonate and was sterilized before use at 100 °C for 30 min to eliminate all viable weed seeds [37].
Ambrosia artemisiifolia seeds were imbedded or conditioned with ethephon, GA3, ethephon + GA3, or thiourea solutions at the same concentrations as those that were used in the laboratory experiments (Table 3). Then, the seeds were sown at a depth of 1.5 cm [21,38] by placing the narrowest part from which the radicle develops towards the bottom of the pot. Afterward, the pots were placed in a greenhouse with an average ambient temperature of 15.8 °C and natural light conditions. An irrigation system (Water Control Master, GARDENA, Ulm, Germany) was used to maintain the humidity of the medium throughout the experiment. All of the plants were cultivated for 21 days and the number of emerged seedlings, as well as the hypocotyl length and the total plant height were measured every seven days. Growth deformities were also assessed qualitatively. The experiment was conducted using a complete block design in eight replicates with 20 seeds per pot.
4.4. Statistical Analysis
The laboratory and greenhouse experiment were performed twice with four and eight replicates each time, respectively. No differences were identified between the experiment using the t-test (p = 0.6638), and the data were combined for analysis. All the replicates were treated as a random effect, while the seed storage, stratification period, and seed treatments were treated as fixed effects. The normality and homogeneity of variance of all the data were evaluated, and an ANOVA was performed. The marginal means were estimated based on the best fitted linear model. The significance of differences between the estimated marginal means of variables was assessed using Tukey’s test. Differences with p < 0.05 were considered statistically significant. The statistical analysis was performed in R programming language and environment. The germination dynamics were quantified using a logistic function in the statistical program Bioassay 97, and the resulting germination time course was used to determine the time that was required for the germination of 10% (t10) or 50% (t50) of the sown seeds.
Author Contributions
Conceptualization, M.Š. methodology, M.Š., L.K. and V.Š.; software, L.P. and V.Š.; validation, M.Š.; formal analysis, V.Š. and L.P.; investigation, L.K. and V.Š.; resources, M.Š.; data curation, V.Š. and L.K.; writing—original draft preparation, M.Š.; writing— review and editing, L.K. and V.Š.; visualization, M.Š., V.Š. and L.P.; supervision, M.Š.; project administration, M.Š.; funding acquisition—M.Š. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The experiment was supported by the Croatian Ministry of Agriculture within the project “Monitoring of pest resistance to plant protection products (2018–2020)” led by Tanja Gotlin-Čuljak from the Department of Agricultural Zoology of the Faculty of Agriculture University of Zagreb.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Di Tommaso, A. Germination behavior of common ragweed (Ambrosia artemisiifolia) populations across a range of salinities. Weed Sci. 2004, 52, 1002–1009. [Google Scholar] [CrossRef]
- Pickett, S.T.; Baskin, J.M. The role of temperature and light in the germination behavior of Ambrosia artemisiifolia. Bull. Torrey Bot. Club 1973, 100, 165–170. [Google Scholar] [CrossRef]
- Sang, W.; Liu, X.; Axmacher, J.C. Germination and emergence of Ambrosia artemisiifolia L. under changing environmental conditions in China. Plant Species Biol. 2011, 26, 125–133. [Google Scholar] [CrossRef]
- Brandes, D.; Nitzsche, J. Biology, introduction, dispersal, and distribution of common ragweed (Ambrosia artemisiifolia L.) with special regard to Germany. Nachr. Deut Pflanzenschutzd 2006, 58, 286–291. [Google Scholar]
- Šarić, T.; Ostojić, Z.; Stefanović, L.; Deneva Milanova, S.; Kazinczi, G.; Tyšer, L. The changes of the composition of weed flora in south eastern and central Europe as affected by cropping practices. Herbologia 2011, 12, 8–12. [Google Scholar]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2021. Available online: www.weedsicence.org (accessed on 21 January 2021).
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Fogliatto, S.; Milan, M.; De Palo, F.; Vidotto, F. The effect of various after-ripening temperature regimens on the germination behaviour of Ambrosia artemisiifolia. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2019, 154, 165–172. [Google Scholar] [CrossRef]
- Willemsen, R.W.; Rice, E.L. Mechanism of seed dormancy in Ambrosia artemisiifolia. Am. J. Bot. 1972, 59, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.J. Abscisic acid and gibberellic acid regulation of seed germination and dormancy. Biologist 1969, 51, 91–99. [Google Scholar]
- Ame, R.D. A model of seed dormancy. Bot. Rev. 1968, 34, 1–31. [Google Scholar]
- Corbineau, F.; Xia, Q.W.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a key factor in the regulation of seed dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matilla, A.J. Ethylene in seed formation and germination. Seed Sci. Res. 2000, 10, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Ghassemian, M.; Nambara, E.; Cutler, S.; Kawaide, H.; Kamiya, Y.; McCourt, P. Regulation of abscisic acid signaling by the ethylene response pathway in arabidopsis. Plant Cell 2000, 12, 1117–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samimy, C.; Khan, A.A. Secondary dormancy, growth-regulator effects, and embryo growth potential in curly dock (Rumex crispus) seeds. Weed Sci. 1983, 31, 153–158. [Google Scholar] [CrossRef]
- Harre, N.T.; Weller, S.C.; Young, B.G. An improved method to shorten physiological dormancy of giant ragweed (Ambrosia trifida) seed. Weed Sci. 2019, 67, 205–213. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Breaking seed dormancy during dry storage: A useful tool of major problem for successful restoration via direct seeding? Plants 2020, 9, 636. [Google Scholar] [CrossRef]
- Page, E.R.; Nurse, R.E. Comparing physical, chemical, and cold stratification methods for alleviating dormancy of giant ragweed (Ambrosia trifida) seeds. Weed Technol. 2015, 29, 311–317. [Google Scholar] [CrossRef]
- Kazinczi, G.; Biro, K.; Beres, I.; Ferger, B. Intraspecific differences in the germination of common ragweed (Ambrosia artemisiifolia L.). (Fajon belüli), (intraspecifikus) különbségek az ürömlevelű parlagfű (Ambrosia artemisiifolia L.) csírázásában). Növényvédelem 2006, 42, 477–481. [Google Scholar]
- Ciappetta, S.; Ghiani, A.; Gilardelli, F.; Bonini, M.; Citterio, S.; Gentili, R. Invasion of Ambrosia artemisiifolia in Italy: Assessment via analysis of genetic variability and herbarium data. Flora Morphol. Distrib. Funct. Ecol. Plants 2016, 223, 106–113. [Google Scholar] [CrossRef]
- Dinelli, G.; Marotti, I.; Catizone, P.; Bosi, S.; Tanveer, A.; Abbas, R.N.; Pavlovic, D. Germination ecology of Ambrosia artemisiifolia L. and Ambrosia trifida L. biotypes suspected of glyphosate resistance. Cent. Eur. J. Biol. 2013, 8, 286–296. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Secondary dormancy in the seeds of the common ragweed Ambrosia artemisiifolia. Bull. Torrey Bot. Club 1970, 97, 302–305. [Google Scholar] [CrossRef]
- Šoštarčić, V.; Košćak, L.; Culjak, A.; Jerić, I.; Šćepanović, M. Utjecaj hladnog skladištenja na klijanje korovne vrste Ambrosia artemisiifolia L. Fragm. Phytomedica 2020, 34, 16–26. [Google Scholar]
- Boddy, L.G.; Bradford, K.J.; Fischer, A.J. Stratification requirements for seed dormancy alleviation in a wetland weed. PLoS ONE 2013, 8, e71457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tieng, M.T. Germination Inhibition and Promotion of Ragweed (Ambrosia trifida L.) and Smartweed (Polygonum pensylvanicum L.). Ph.D. Thesis, Purdue University, West Lafayette, IN, USA, 1962. [Google Scholar]
- Šoštarčić, V.; Masin, R.; Turčinov, M.; Carin, N.; Šćepanović, M. Intrapopulation variability in morphological and functional properties of Ambrosia artemisiifolia L. seeds. J. Cent. Eur. Agric. 2020, 21, 366–378. [Google Scholar] [CrossRef]
- Vink, J.P.; Soltani, N.; Robinson, D.E.; Tardif, F.J.; Lawton, M.B.; Sikkema, P.H. Occurrence and distribution of glyphosate-resistant giant ragweed (Ambrosia trifida L.) in southwestern Ontario. Can. J. Plant Sci. 2012, 92, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Westhoven, A.M.; Davis, V.M.; Gibson, K.D.; Weller, S.C.; Johnson, W.G. Field presence of glyphosate-resistant horseweed (Conyza canadensis), common lambsquarters (Chenopodium album), and giant ragweed (Ambrosia trifida) biotypes with elevated tolerance to glyphosate. Weed Technol. 2008, 22, 544–548. [Google Scholar] [CrossRef]
- Moretti, M.L.; Van Horn, C.R.; Robertson, R.R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.G.; Sammons, R.D.; Wang, D.; Ge, X.; et al. Glyphosate resistance in Ambrosia trifida. Part 2. Rapid response physiology and non-target site resistance. Pest Manag. Sci. 2018, 74, 1079–1088. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Jha, P.; Steckel, L.E.; Scott, R.C. Confirmation and control of glyphosate-resistant giant ragweed (Ambrosia trifida) in Tennessee. Weed Technol. 2010, 24, 64–70. [Google Scholar] [CrossRef]
- Ballard, T.O.; Foley, M.E.; Bauman, T.T. Germination, viability, and protein changes during cold stratification of giant ragweed (Ambrosia trifida L.) seed. J. Plant Physiol. 1996, 149, 229–232. [Google Scholar] [CrossRef]
- Vange, V.; Heuch, I.; Vandvik, V. Do seed mass and family affect germination and juvenile performance in Knautia arvensis? A study using failure-time methods. Acta Oecol. 2004, 25, 169–178. [Google Scholar] [CrossRef]
- Burgos, N.R.; Tranel, P.J.; Streibig, J.C.; Davis, V.M.; Shaner, D.; Norsworthy, J.K.; Ritz, C. Review: Confirmation of resistance to herbicides and evaluation of resistance levels. Weed Sci. 2013, 61, 4–20. [Google Scholar] [CrossRef]
- Schutte, B.J.; Regnier, E.E.; Harrison, S.K. Seed dormancy and adaptive seedling emergence timing in giant ragweed (Ambrosia trifida). Weed Sci. 2012, 60, 19–26. [Google Scholar] [CrossRef]
- Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Šoštarčić, V.; Masin, R.; Loddo, D.; Brijačak, E.; Šćepanović, M. Germination Parameters of Selected Summer Weeds: Transferring of the AlertInf Model to Other Geographical Regions. Agronomy 2021, 11, 292. [Google Scholar] [CrossRef]
- Dahlquist, R.; Prather, T.; Stapleton, J. Time and temperature requirements for weed seed thermal death. Weed Sci. 2007, 55, 619–625. [Google Scholar] [CrossRef]
- Guillemin, J.P.; Chauvel, B. Effects of the seed weight and burial depth on the seed behavior of common ragweed (Ambrosia artemisiifolia). Weed Biol. Manag. 2011, 11, 217–223. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).