
Effect of the Application of Hydrolysate of Chlorella vulgaris Extracted by Different Techniques on the Growth of Pelargonium × hortorum


 ,
,  and
and
Abstract
1. Introduction
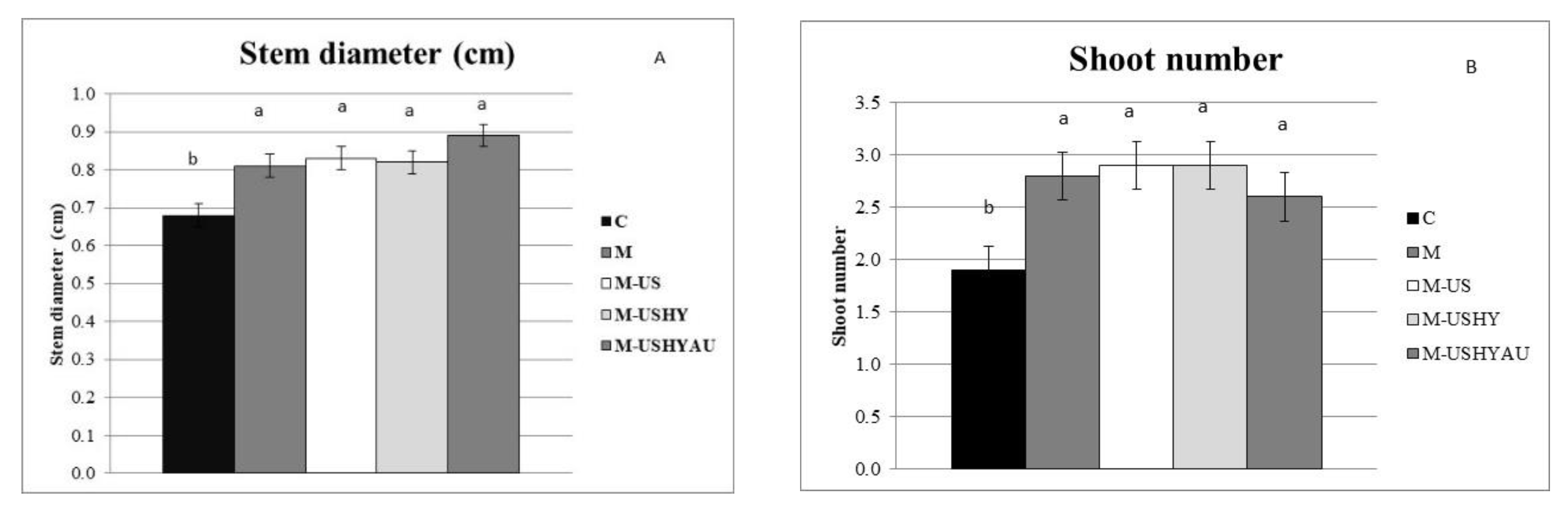
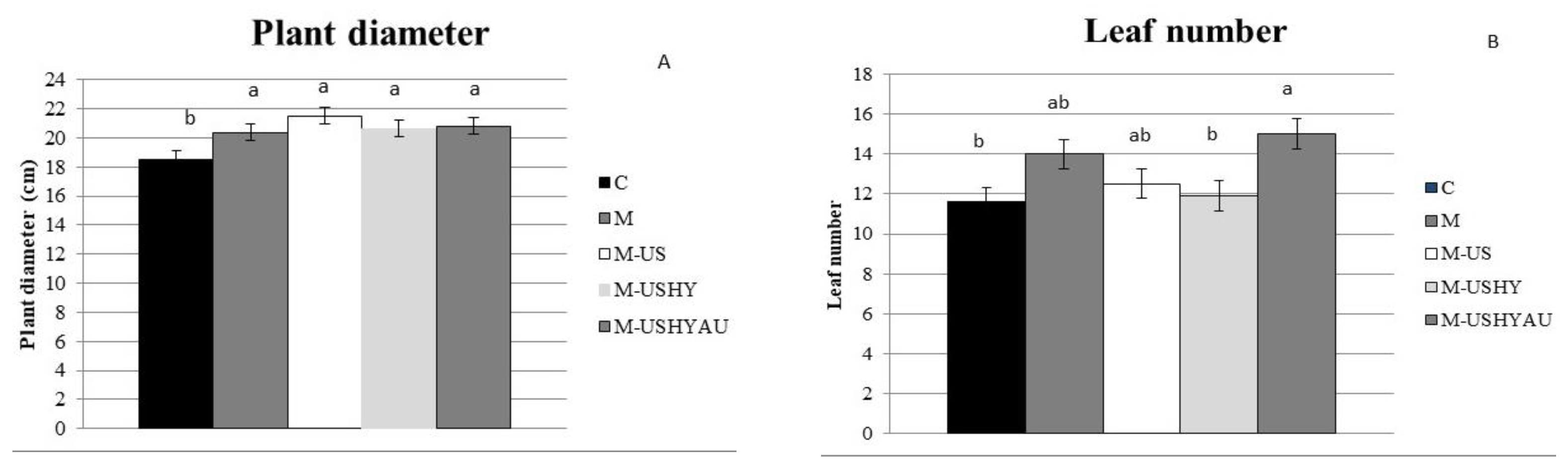
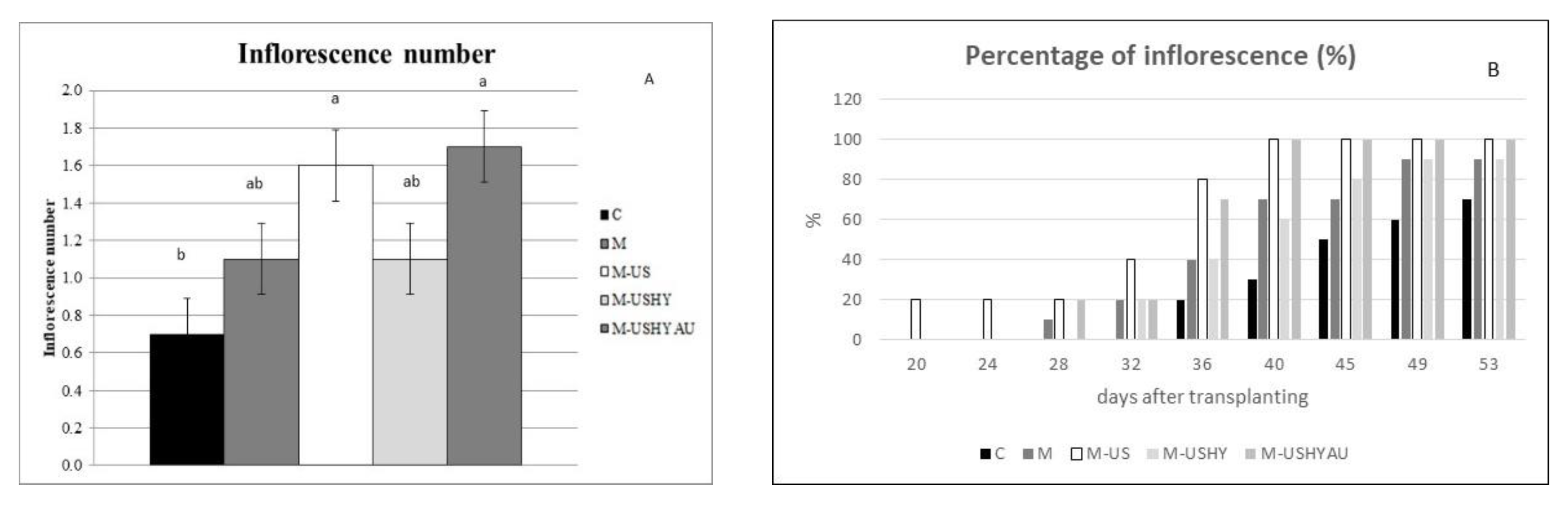
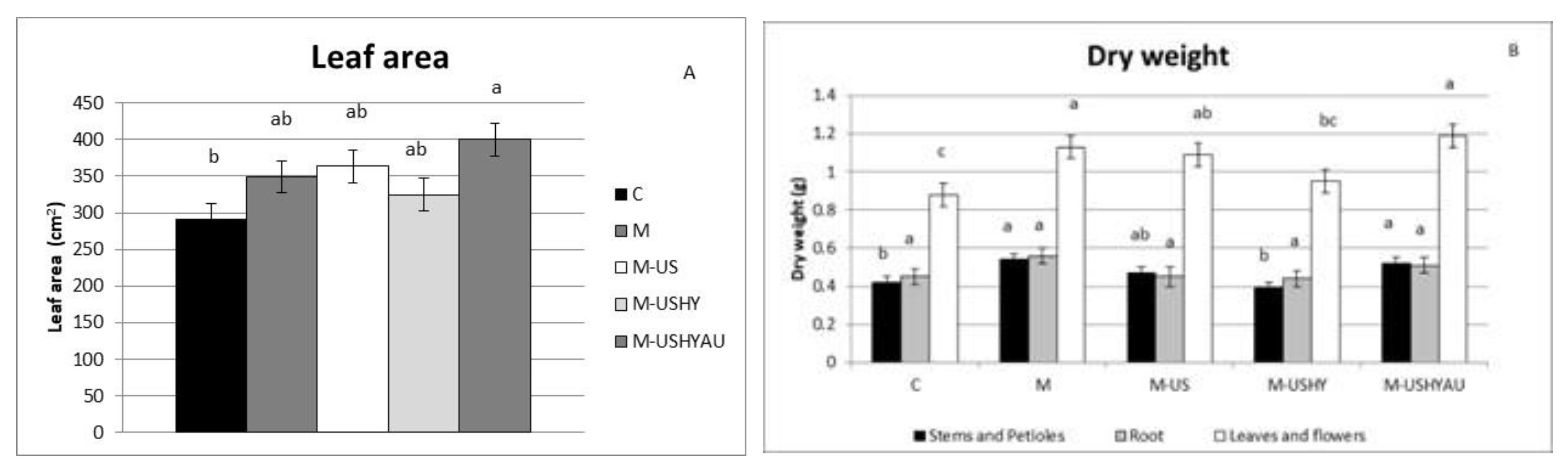
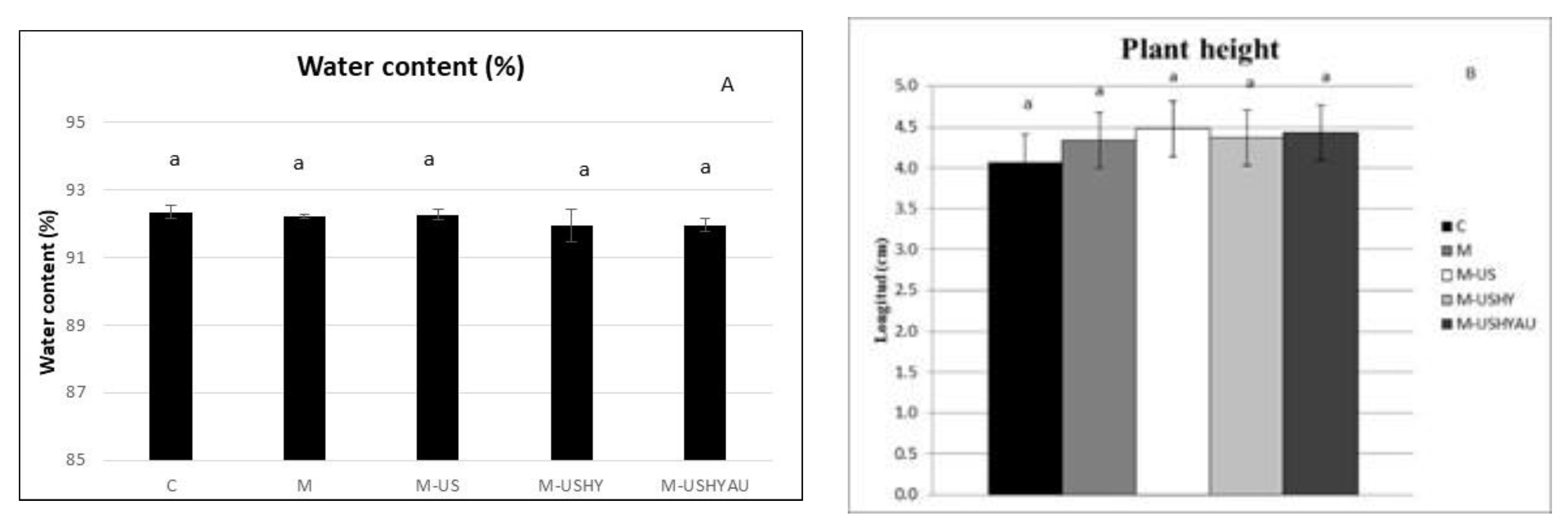
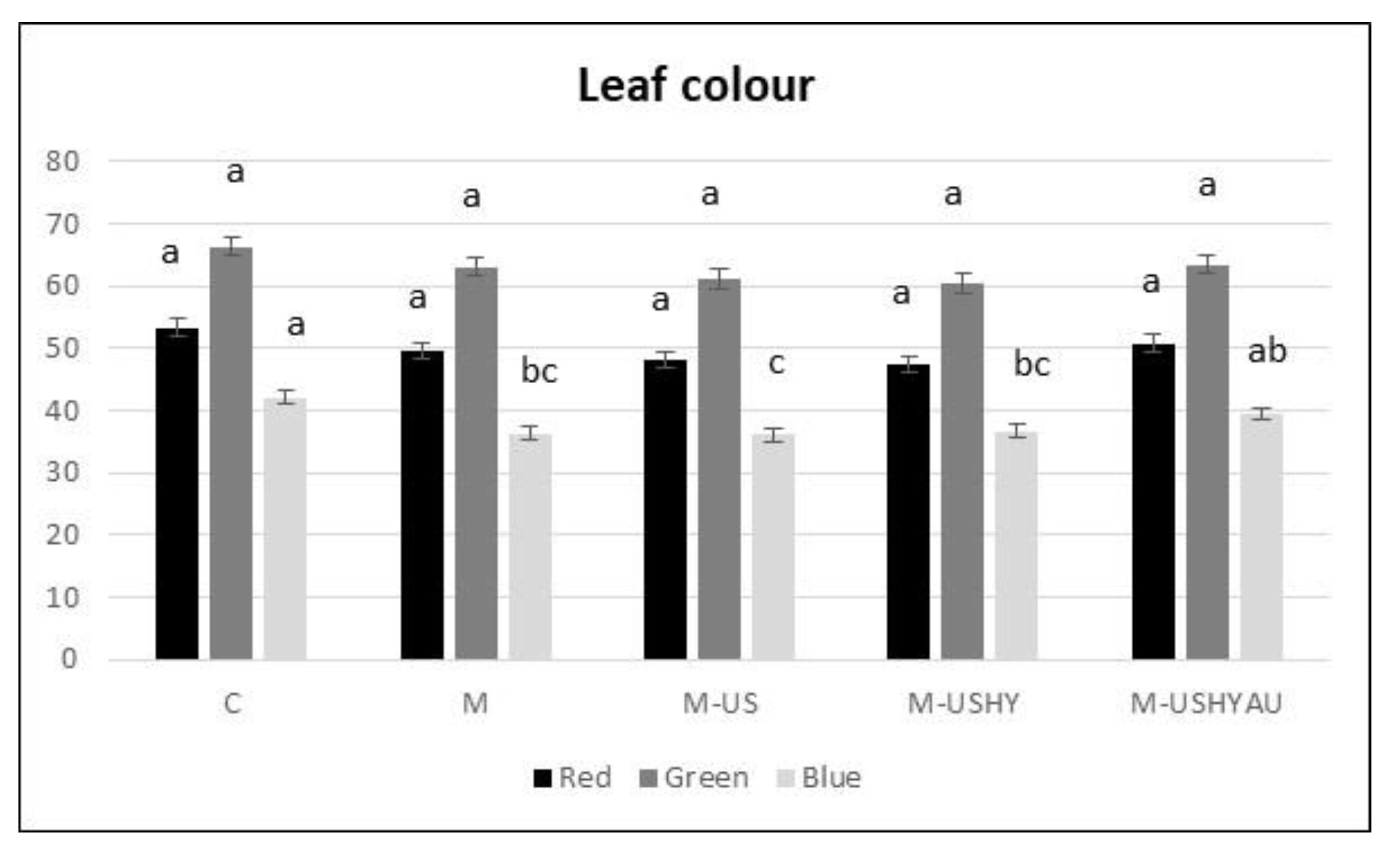
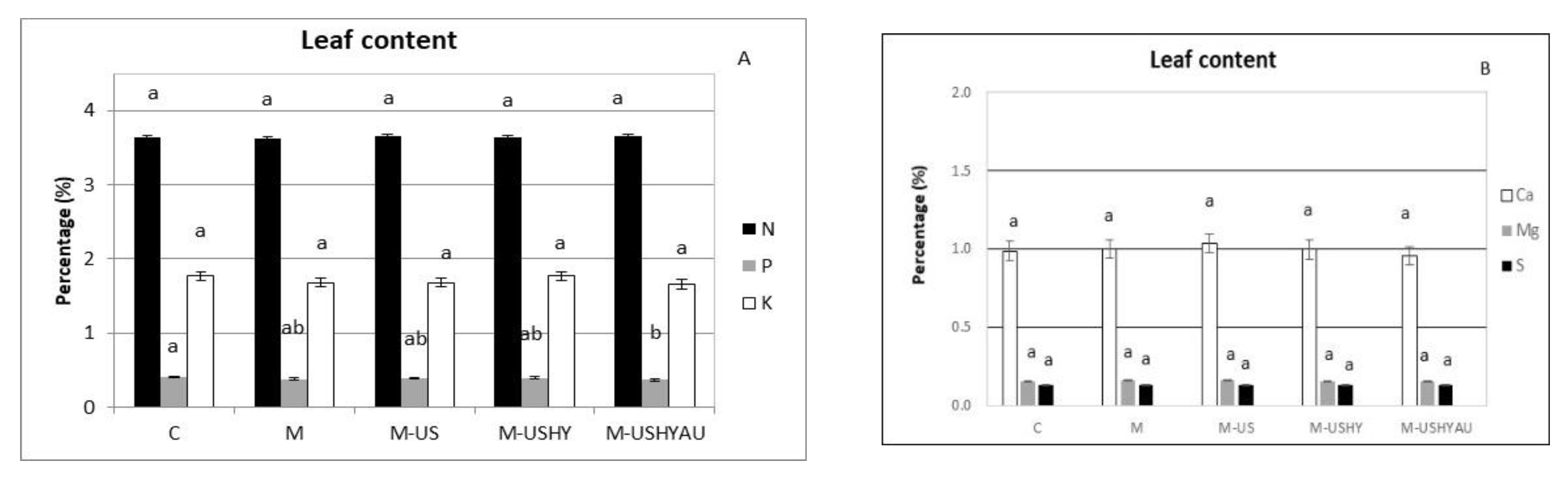
2. Results
3. Material and Methods
3.1. Microalgae Biomass Production
3.2. Microalgae Application and Treatments
3.3. Greenhouse Trial
3.4. Biometric Parameters
3.5. Plant Analysis
3.6. Experimental Design and Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Puglisi, I.; Barone, V.; Sidella, S.; Coppa, M.; Broccanello, C.; Gennari, M.; Baglieri, A. Biostimulant activity of humic-like substances from agro-industrial waste on Chlorella vulgaris and Scenedesmus quadricauda. Eur. J. Phycol. 2018, 53, 433–442. [Google Scholar] [CrossRef]
- Ventura, S.; Nobre, B.; Ertekin, F.; Hayes, M.; Garciá-Vaquero, M.; Vieira, F.; Koc, M.; Gouveia, L.; Aires-Barros, M.; Palavra, A. Extraction of value-added compounds from microalgae. In Energy, Microalgae-Based Biofuels and Bioproducts (Woodhead Publishing Series); Gonzalez-Fernandez, C., Muñoz, R., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 461–483. [Google Scholar] [CrossRef]
- Sudhakar, M.P.; Ramesh Kumar, B.; Mathimani, T.; Arunkumar, K. A review on bioenergy and bioactive compounds from microalgae and macroalgae-sustainable energy perspective. J. Clean. Prod. 2019, 228, 1320–1333. [Google Scholar] [CrossRef]
- Tan, J.S.; Lee, S.Y.; Chew, K.W.; Lam, M.K.; Lim, J.W.; Ho, S.-H.; Show, P.L. A review on microalgae cultivation and harvesting, and their biomass extraction processing using ionic liquids. Bioengineered 2020, 11, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Acién, F.G.; Gómez-Serrano, C.; Morales-Amaral, M.M.; Fernandez-Sevilla, F.G.A.; Molina-Grima, E. Wastewater treatment using microalgae: How realistic a contribution might it be to significant urban wastewater treatment? Appl. Microbiol. Biotechnol. 2016, 100, 9013–9022. [Google Scholar] [CrossRef] [PubMed]
- Mathimani, T.; Pugazhendhi, A. Utilization of algae for biofuel, bio-products and bio-remediation. Biocatal. Agric. Biotechnol. 2019, 17, 326–330. [Google Scholar] [CrossRef]
- Bayona-Morcillo, P.J.; Plaza, B.M.; Gómez-Serrano, C.; Rojas, E.; Jiménez-Becker, S. Effect of the foliar application of cyanobacterial hydrolysate (Arthrospira platensis) on the growth of Petunia x hybrida under salinity conditions. J. Appl. Phycol. 2020, 32, 4003–4011. [Google Scholar] [CrossRef]
- Ramesh-Kumar, B.; Deviram, G.; Mathimani, T.; Anh Duc, P.; Pugazhendhi, A. Microalgae as rich source of polyunsaturated fatty acids. Biocatal. Agric. Biotechnol. 2019, 17, 583–588. [Google Scholar] [CrossRef]
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable Sources of Plant Biostimulation: Microalgae as a Sustainable Means to Improve Crop Performance. Front. Plant Sci. 2018, 9, 1782. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae—A review. J. Algal. Biomass. Util. 2012, 3, 89–100. [Google Scholar]
- Panahi, Y.; Khosroushahi, A.Y.; Sahebkar, A.; Heidari, H.R. Impact of Cultivation Condition and Media Content on Chlorella vulgaris Composition. Adv. Pharm. Bull. 2019, 9, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, G.; Moovendhan, M.; Arumugam, A.; Matharasi, A.; Dineshkumar, R.; Sampathkumar, P. Evaluation of Chemical Composition and In Vitro Antiinflammatory Effect of Marine Microalgae Chlorella vulgaris. Waste Biomass Valor. 2018, 10, 3263–3270. [Google Scholar] [CrossRef]
- Stirk, W.A.; Ördög, V.; Novák, O.; Rolčík, J.; Strnad, M.; Bálint, P.; van Staden, J. Auxin and cytokinin relationships in 24 microalgal strains(1). J. Phycol. 2013, 49, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Wild, K.J.; Trautmann, A.; Katzenmeyer, M.; Steingaß, H.; Posten, C.; Rodehutscord, M. Chemical composition and nutritional characteristics for ruminants of the microalgae Chlorella vulgaris obtained using different cultivation conditions. Algal Res. 2018, 38, 101385. [Google Scholar] [CrossRef]
- Giuffrida, F.; Rouphael, Y.; Toscano, S.; Scuderi, D.; Romano, D.; Rivera, C.M.; Colla, G.; Leonardi, C. A simple model for nondestructive leaf area estimation in bedding plants. Photosynthetica 2011, 49, 380–388. [Google Scholar] [CrossRef]
- Hogue, E.; Wilcox, G.E.; Cantliffe, D.J. Effect of soil P levels on phosphate fraction in tomato leaves. J. Am. Soc. Hortic. Sci. 1970, 95, 174–176. [Google Scholar] [CrossRef]
- Krom, M.D. Spectrophotometric determination of NH4+: A study of a modified berthelot reaction using salicylate and dichloroisocyanurate. Analyst 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Gitau, M.M.; Farkas, A.; Balla, B.; Ördög, V.; Futó, Z.; Maróti, G. Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago truncatula. Plants 2021, 10, 1060. [Google Scholar] [CrossRef]
- Tejada-Ruiz, S.; Gonzalez-Lopez, C.; Rojas, E.; Jiménez-Becker, S. Effect of the Foliar Application of Microalgae Hydrolysate (Arthrospira platensis) and Silicon on the Growth of Pelargonium hortorum L.H. Bailey under Salinity Conditions. Agronomy 2020, 10, 1713. [Google Scholar] [CrossRef]
- Rahimi, S.; Naderi, R.; Ghaemaghami, S.; Kalatejari, S.; Farham, B. Study on Effects of Different Plant Growth Regulators Types in Shoot Regeneration and Node Formation of Sutsuki Azalea (Rhododendron indicum): A Commercially Important Bonsai. Procedia Eng. 2013, 59, 240–246. [Google Scholar] [CrossRef][Green Version]
- Refaay, D.A.; El-Marzoki, E.M.; Abdel-Hamid, M.I.; Haroun, S.A. Effect of foliar application with Chlorella vulgaris, Tetradesmus dimorphus, and Arthrospira platensis as biostimulants for common bean. J. Appl. Phycol. 2021, 33, 3807–3815. [Google Scholar] [CrossRef]
- Ciereszko, I. Regulatory roles of sugars in plant growth and development. Acta Soc. Bot. Pol. 2018, 87. [Google Scholar] [CrossRef]
- Rachidi, F.; Benhima, R.; Sbabou, L.; El Arroussi, H. Microalgae polysaccharides bio-stimulating effect on tomato plants: Growth and metabolic distribution. Biotechnol. Rep. 2020, 25, e00426. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef]
- Francesca, S.; Cirillo, V.; Raimondi, G.; Maggio, A.; Barone, A.; Rigano, M.M. A Novel Protein Hydrolysate-Based Biostimulant Improves Tomato Performances under Drought Stress. Plants 2021, 10, 783. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, X.; Cui, Y.; Yuan, W. Ultrasound for microalgal cell disruption and product extraction: A review. Ultrason. Sonochem. 2022, 87, 06054. [Google Scholar] [CrossRef]
- Saha, S.; Shukla, S.K.; Singh, H.R.; Singh, B.; Jha, S.K. Chapter 16—Bioactive compounds from microalgae. In An Integration of Phycoremediation Processes in Wastewater Treatment; Shah, M., Rodriguez-Couto, S., Vargas De La Cruz, C.B., Biswas, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 337–358. [Google Scholar] [CrossRef]
- Kim, H.U. Lipid Metabolism in Plants. Plants 2020, 9, 871. [Google Scholar] [CrossRef]
- Keris-Sen, U.D.; Sen, U.; Soydemir, G.; Gurol, M.D. An investigation of ultrasound effect on microalgal cell integrity and lipid extraction efficiency. Bioresour. Technol. 2014, 152, 407–413. [Google Scholar] [CrossRef]
- Deenu, A.; Naruenartwongsakul, S.; Kim, S.M. Optimization and economic evaluation of ultrasound extraction of lutein from Chlorella vulgaris. Biotechnol. Bioprocess Eng. 2013, 18, 1151–1162. [Google Scholar] [CrossRef]
- Parniakov, O.; Apicella, E.; Koubaa, M.; Barba, F.; Grimi, N.; Lebovka, N.; Pataro, G.; Ferrari, G.; Vorobiev, E. Ultrasound-assisted green solvent extraction of high-added value compounds from microalgae Nannochloropsis spp. Bioresour. Technol. 2015, 198, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid Metabolism in Plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Hajnal-Jafari, T.; Seman, V.; Stamenov, D.; Duric, S. Effect of Chlorella vulgaris on Growth and Photosynthetic Pigment Content in Swiss Chard (Beta vulgaris L. subsp. cicla). Pol. J. Microbiol. 2020, 69, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, T.R. Physical and chemical cell disruption for the recovery of intracellular proteins. Bioprocess Technol. 1991, 12, 57–83. [Google Scholar] [PubMed]
- Middelberg, A.P.J. Process-scale disruption of microorganisms. Biotechnol. Adv. 1995, 13, 491–551. [Google Scholar] [CrossRef]
- Salinas-Salazar, C.; Garcia-Perez, J.S.; Chandra, R.; Castillo-Zacarias, C.; Iqbal, H.M.N.; Parra-Saldívar, R. Methods for Extraction of Valuable Products from Microalgae Biomass. In Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Springer: Singapore, 2019; pp. 245–263. [Google Scholar] [CrossRef]
- Stirk, W.A.; Bálint, P.; Vambe, M.c.M.; Lovász, C.; Molnár, Z.; van Staden, J.; Ördög, V. Effect of cell disruption methods on the extraction of bioactive metabolites from microalgal biomass. J. Biotechnol. 2020, 307, 35–43. [Google Scholar] [CrossRef]
- Kusvuran, S. Microalgae (Chlorella vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Hortic. Plant J. 2021, 7, 221–231. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. vulgaris | ||
|---|---|---|
| Ethylene | ACC | 11.19 |
| Cytokinins | Trans-Zeatin | 813.65 |
| Isopentenyl-adenine | 3816.28 | |
| Gibberellins | GA1 | 7.28 |
| GA3 | 0.01 | |
| GA4 | 26.62 | |
| Auxins | Indoleacetic acid | 2.53 |
| Other hormones | ABA | 2.17 |
| Salicylic acid | 408.86 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayona-Morcillo, P.J.; Gómez-Serrano, C.; González-López, C.V.; Massa, D.; Jiménez-Becker, S. Effect of the Application of Hydrolysate of Chlorella vulgaris Extracted by Different Techniques on the Growth of Pelargonium × hortorum. Plants 2022, 11, 2308. https://doi.org/10.3390/plants11172308
Bayona-Morcillo PJ, Gómez-Serrano C, González-López CV, Massa D, Jiménez-Becker S. Effect of the Application of Hydrolysate of Chlorella vulgaris Extracted by Different Techniques on the Growth of Pelargonium × hortorum. Plants. 2022; 11(17):2308. https://doi.org/10.3390/plants11172308
Chicago/Turabian StyleBayona-Morcillo, Pedro José, Cintia Gómez-Serrano, Cynthia Victoria González-López, Daniele Massa, and Silvia Jiménez-Becker. 2022. "Effect of the Application of Hydrolysate of Chlorella vulgaris Extracted by Different Techniques on the Growth of Pelargonium × hortorum" Plants 11, no. 17: 2308. https://doi.org/10.3390/plants11172308
APA StyleBayona-Morcillo, P. J., Gómez-Serrano, C., González-López, C. V., Massa, D., & Jiménez-Becker, S. (2022). Effect of the Application of Hydrolysate of Chlorella vulgaris Extracted by Different Techniques on the Growth of Pelargonium × hortorum. Plants, 11(17), 2308. https://doi.org/10.3390/plants11172308