
Viability of Embryo Sacs and Fruit Set in Different Plum (Prunus domestica L.) Cultivars Grown under Norwegian Climatic Conditions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Time of Flowering and Climatic Conditions
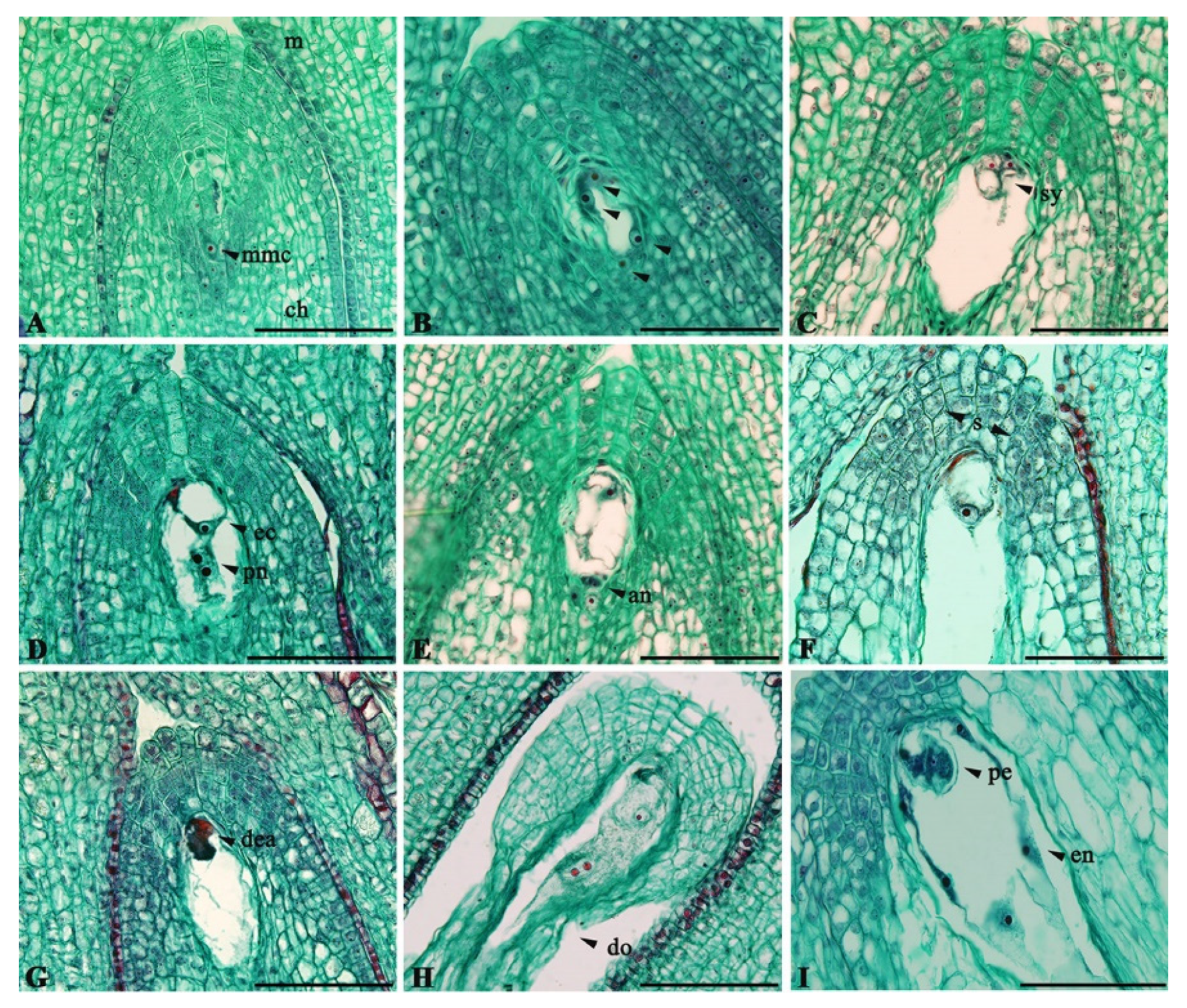
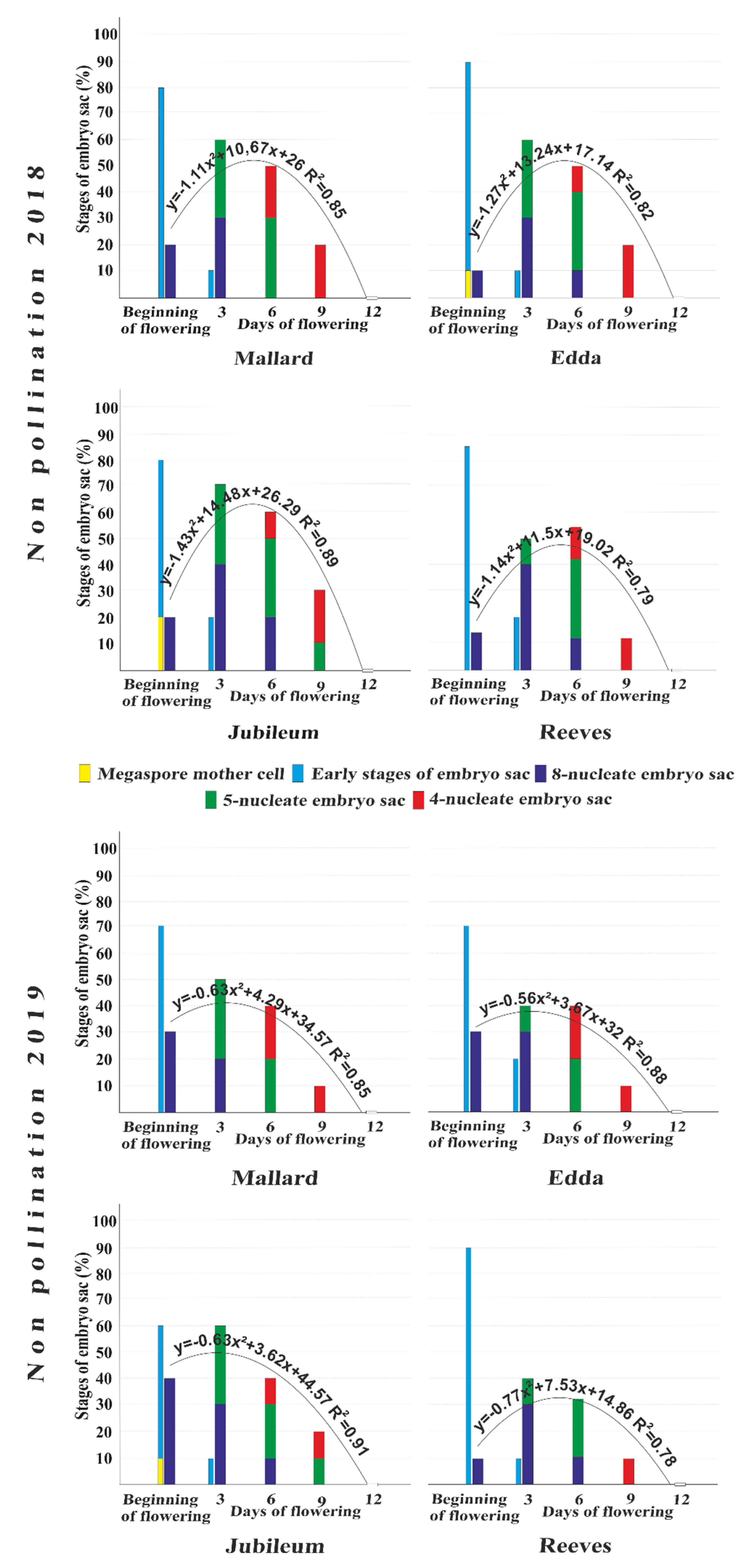
2.2. Development and Viability of Embryo Sacs
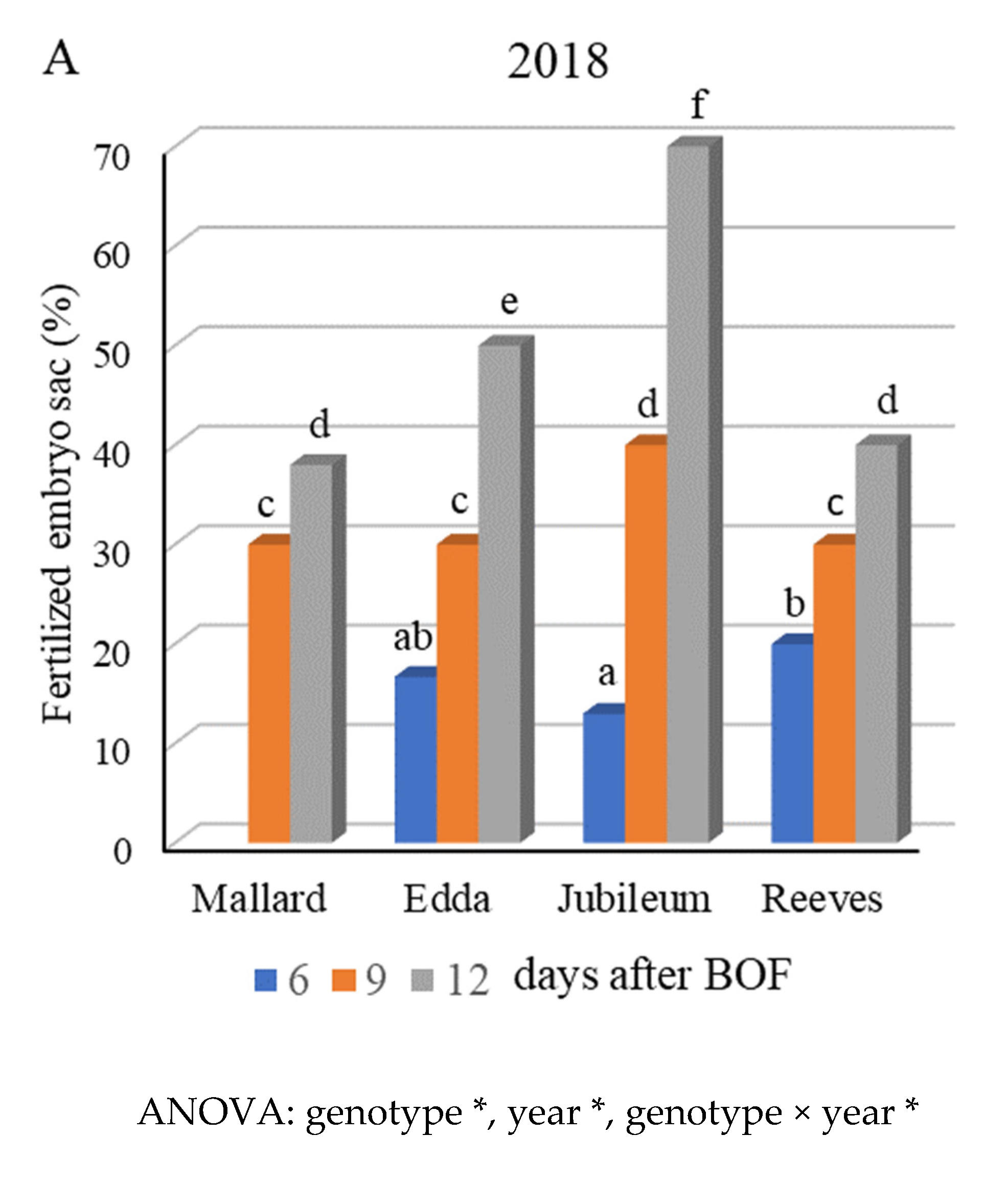
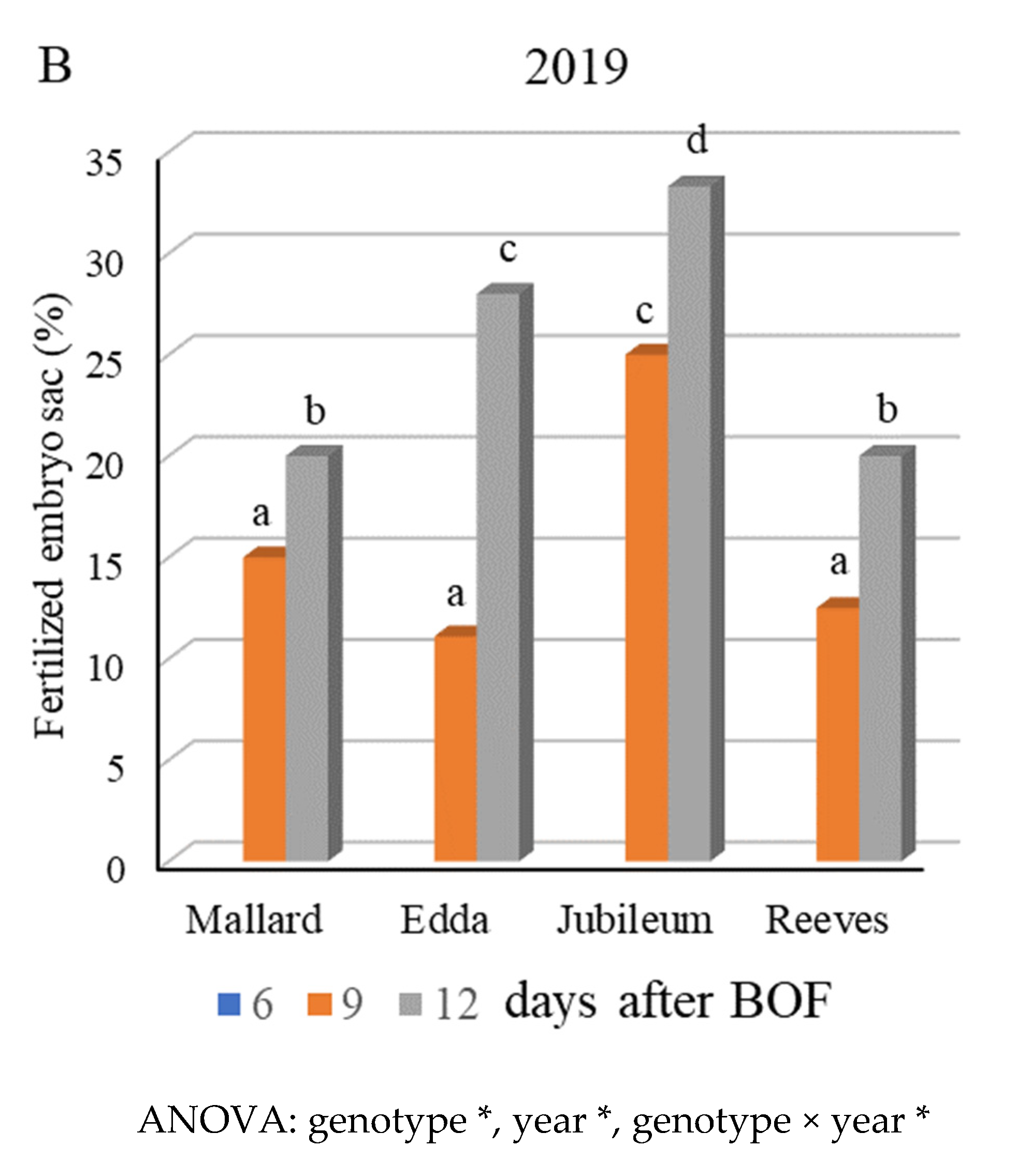
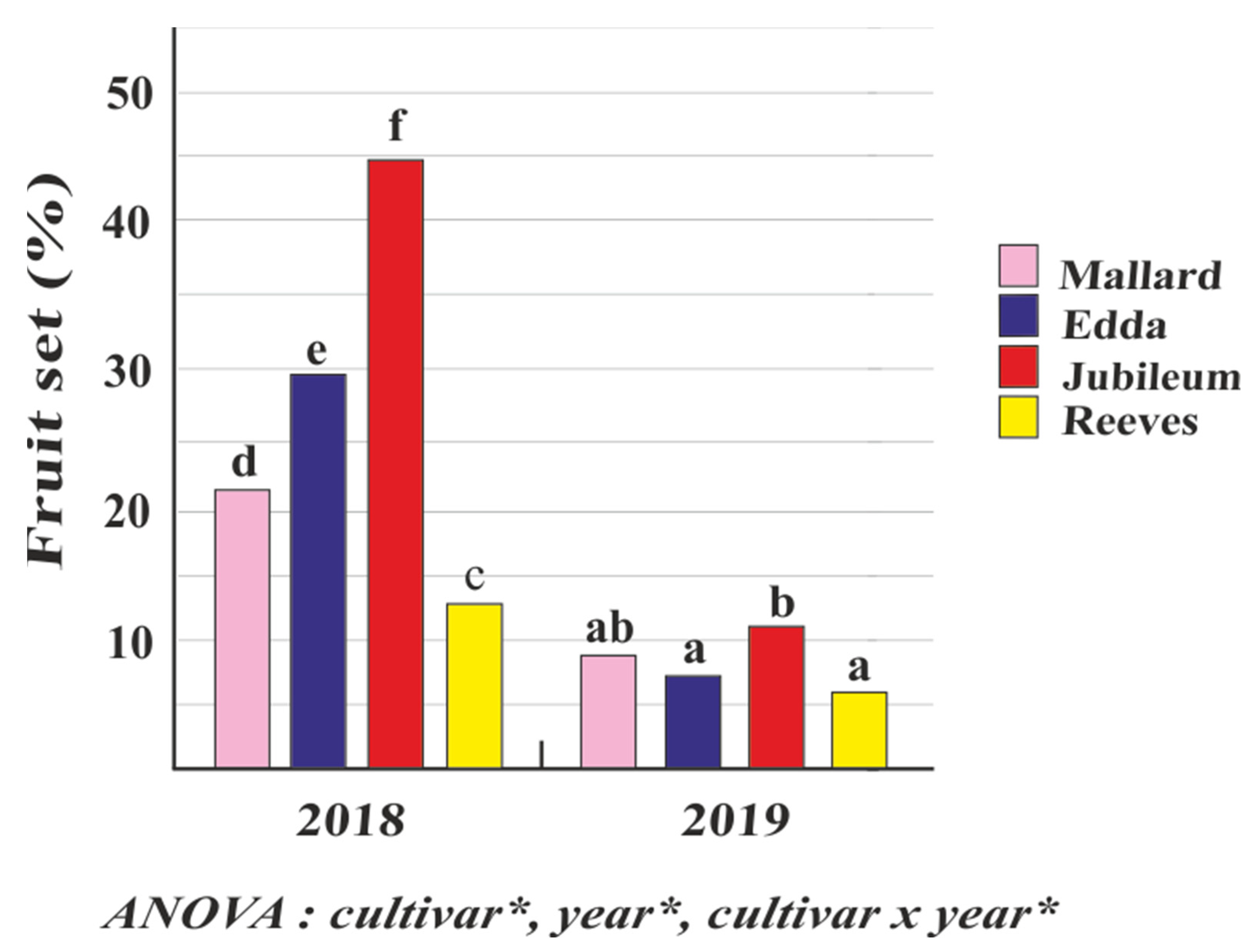
2.3. Fertilization Success and Fruit Set
3. Discussion
3.1. Development and Viability of Embryo Sacs
3.2. Fertilization Success and Fruit Set
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Flowering Time and Climate Conditions
4.3. Microscopic Preparations
4.4. Fruit Set
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meland, M.; Frøynes, O.; Fotirić Akšić, M.; Pojskić, N.; Kalamujić Stroil, B.; Lasi, L.; Gaši, F. Identifying pollen donors and success rate of individual pollinizers in European plum (Prunus domestica L.) using microsatellite markers. Agronomy 2020, 10, 264. [Google Scholar] [CrossRef] [Green Version]
- Sekse, L. Plum production in Norway. Acta Hortic. 2007, 734, 23–26. [Google Scholar] [CrossRef]
- FAOStat. 2020. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 22 November 2021).
- Redalen, G. Plum growing in Norway, at 60° N. Acta Hortic. 2002, 577, 385–389. [Google Scholar] [CrossRef]
- Hjeltnes, S.H.; Røen, K.B. Pollen germination of some plum (Prunus domestica L.) cultivars under different temperatures. Acta Hortic. 2019, 1260, 113–116. [Google Scholar] [CrossRef]
- Hjeltnes, S.H. Plum breeding in Norway—Past, present and future. Acta Hortic. 1994, 359, 63–68. [Google Scholar] [CrossRef]
- Vangdal, E.; Flatland, S.; Nordbø, R. Fruit quality changes during marketing of new plum cultivars (Prunus domestica L.). Hortic. Sci. 2007, 34, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Hartman, W.; Neumuller, M. Plum Breeding. In Breeding Plantation Tree Crops: Temperate Species; Mohan Jain, S., Priyadarshan, P.M., Eds.; Springer: New York, NY, USA, 2009; pp. 161–189. [Google Scholar]
- Cerović, R.; Mićić, N. Oprašivanje i oplodnja jabučastih i koštičavih voćaka. Jugosl. Voćarstvo 1996, 30, 73–98. [Google Scholar]
- Cerović, R.; Mićić, N. Functionality of embryo sacs as related to their viability and fertilization success in sour cherry. Sci. Hortic. 1999, 79, 227–235. [Google Scholar] [CrossRef]
- Dyś, B. Cyto-embryological studies in self-incompatible and self-fertile cultivars of sour-cherries (Cerasus vulgaris Mill.). II Development of embryo sacs and ovules at some stages of florescence. Gen. Pol. 1984, 25, 171–180. [Google Scholar]
- Furukawa, Y.; Bukovac, M.J. Embryo sac development in sour cherry during the pollination period as related to fruit set. HortScience 1989, 24, 1005–1008. [Google Scholar]
- Alburquerque, N.; Burgos, L.; Egea, J. Variability in the developmental stage of apricot ovules at anthesis and its relationship with fruit set. Ann. Appl. Biol. 2002, 141, 147–152. [Google Scholar] [CrossRef]
- Cerović, R.; Fotirić Akšić, M.; Đorđević, M.; Meland, M. The effects of pollinizers on pollen tube growth and fruit set of European plum (Prunus domestica L.) in a Nordic climate. Sci. Hortic. 2021, 288, 110390. [Google Scholar] [CrossRef]
- Hedhly, A. Sensitivity of flowering plant gametophytes to temperature fluctuations. Environ. Exp. Bot. 2011, 74, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Koskela, E.; Kemp, H.; van Dieren, M.C.A. Flowering and pollination studies with European plum (Prunus domestica L.) cultivars. Acta Hortic. 2010, 874, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. The effect of temperature on stigmatic receptivity in sweet cherry (Prunus avium L.). Plant Cell Environ. 2003, 26, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Stösser, R. Zur Befruchtungsbiologie der Zwetschensorte ‘Valjevka’. Erwerbsobstbau 2002, 44, 71–75. [Google Scholar]
- Đorđević, M.; Cerović, R.; Radičević, S.; Nikolić, D.; Milošević, N.; Glišić, I.; Marić, S.; Lukić, M. Pollen tube growth and embryo sac development in ‘Pozna Plava’ plum cultivar related to fruit set. Erwerbsobstbau 2019, 61, 313–322. [Google Scholar] [CrossRef]
- Cerović, R.; Ružić, Đ.; Mićić, N. Viability of plum ovules at different temperatures. Ann. Appl. Biol. 2000, 137, 53–59. [Google Scholar] [CrossRef]
- Ruiz, D.; Egea, J. Ovule development at anthesis in apricot (Prunus armeniaca L.) varieties in a Mediterranean climate. Ann. Appl. Biol. 2007, 151, 43–51. [Google Scholar] [CrossRef]
- Radičević, S.; Cerović, R.; Ðorđević, M. Ovule senescence and unusual pollen tube growth in the ovary of sweet cherry as affected by pistilar genotype and temperature. Span. J. Agric. Res. 2018, 16, e0704. [Google Scholar] [CrossRef]
- Williams, R.R. Factors affecting pollination in fruit trees. In Physiology of Tree Crops; Luckwill, L.C., Cutting, C.V., Eds.; Academic Press: London, UK; New York, NY, USA, 1970; pp. 193–207. [Google Scholar]
- Roeder, S.; Serra, S.; Musacchi, S. Effective pollination period and parentage effect on pollen tube growth in apple. Plants 2021, 10, 1618. [Google Scholar] [CrossRef]
- Egea, J.; Burgos, L. Ovule differences between single-kernelled and double-kernelled fruits in almond (Prunus dulcis). Ann. Appl. Biol. 2000, 136, 291–295. [Google Scholar] [CrossRef]
- Ruiz, D.; Campoy, J.A.; Egea, J. Ovule development at anthesis in Japanese plum (Prunus salicina Lindl.). Span. J. Agric. Res. 2010, 8, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, S.; Viti, R.; Guerriero, R. Observations on the fertilization process in self-pollinated flowers of cultivar “Leccino”. Acta Hortic. 2002, 586, 521–524. [Google Scholar] [CrossRef]
- Fotirić Akšić, M.; Cerović, R.; Hjeltnes, S.H.; Meland, M. The effective pollination period in European plum (Prunus domestica L.) cultivars in Western Norway. Horticulturae 2022, 8, 55. [Google Scholar] [CrossRef]
- Pimienta, E.; Polito, V.S. Ovule abortion in ‘Nonpareil’ almond (Prunus dulcis [MILL.] D. A. WEBB). Am. J. Bot. 1983, 69, 913–920. [Google Scholar] [CrossRef]
- Burgos, L.; Berenguer, T.; Egea, J. Embryo-sac development in pollinated and non-pollinated flowers of two apricot cultivars. J. Hortic. Sci. 1995, 70, 35–39. [Google Scholar] [CrossRef]
- Burgos, L.; Alburquerque, N.; Egea, J. Review. Flower biology in apricot and its implications for breeding. Span. J. Agric. Res. 2004, 2, 227–241. [Google Scholar] [CrossRef]
- Arbeloa, A.; Herrero, M. Development of ovular structures in peach [Prunus persica (L.) Batsch]. New Phytol. 1991, 118, 527–534. [Google Scholar] [CrossRef]
- Stösser, R.; Anvari, S.F. On the senescence of ovules in cherries. Sci. Hortic. 1982, 16, 29–38. [Google Scholar] [CrossRef]
- Guerra, M.; Wünsch, A.; López-Corrales, M.; Rodrigo, J. Lack of fruit set caused by ovule degeneration in Japanese plum. J. Am. Soc. Hortic. Sci. 2011, 136, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.M.; Liu, L.J. Temperature, fruit set, and embryo sac development in ‘Italian’ prune. J. Am. Soc. Hortic. Sci. 1973, 98, 193–197. [Google Scholar]
- Dittmann, K.; Stösser, R. Development of ovules in relation to premature fruit drop in Prunus-species. J. Appl. Bot. 1999, 73, 86–98. [Google Scholar]
- Rodrigo, J.; Herrero, M. Influence of intraovular reserves on ovule fate in apricot (Prunus armeniaca L.). Sex. Plant Reprod. 1998, 11, 86–93. [Google Scholar] [CrossRef]
- Cerović, R.; Vujičić, R.; Mićić, N. Localization of polysaccharides in the ovary of sour cherry. Gartenbauwissenschaft 1999, 64, 40–46. [Google Scholar]
- Herrero, M. Changes in the ovary related to pollen tube growth. Ann. Bot. 2000, 85, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Willemse, M.T.M.; van Went, J.L. The female gametophyte. In Embryology of Angiosperms; Johri, B.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 159–196. [Google Scholar]
- Yadegari, R.; Drews, N.G. Female gametophyte development. Plant Cell 2004, 16, 133–141. [Google Scholar] [CrossRef]
- Dresselhaus, T.; Márton, M.L. Micropylar pollen tube guidance and burst: Adapted from defense mechanisms? Curr. Opin. Plant Biol. 2009, 12, 773–780. [Google Scholar] [CrossRef]
- Huang, B.-Q.; Russell, S.D. Female germ unit: Organization, isolation, and function. Int. Rev. Cytol. 1992, 140, 233–293. [Google Scholar] [CrossRef]
- Reiser, L.; Fischer, L.R. The ovule and the embryo sac. Plant Cell 1993, 5, 1291–1301. [Google Scholar] [CrossRef]
- Skinner, D.J.; Sundaresan, V. Recent advances in understanding female gametophyte development. F1000Research 2018, 7, 804. [Google Scholar] [CrossRef]
- Song, X.; Yuan, L.; Sundaresan, V. Antipodal cells persist through fertilization in the female gametophyte of Arabidopsis. Plant Reprod. 2014, 27, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Costa Tura, J.; Mackenzie, K.A.D. Ovule and embryo sac development in Malus pumila L. cv. Coxʹs Orange Pippin, from dormancy to blossom. Ann. Bot. 1990, 66, 443–450. [Google Scholar] [CrossRef]
- Rodrigo, J.; Herrero, M. Effects of pre-blossom temperatures on flower development fruit set in apricots. Sci. Hortic. 2002, 92, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Đorđević, M.; Cerović, R.; Radičević, S.; Glišić, I.; Milošević, N.; Marić, S.; Lukić, M. Abnormalities in the ovule development of the European plum cultivar ‘Pozna Plava’ in the days following anthesis. Sci. Hortic. 2019, 252, 222–228. [Google Scholar] [CrossRef]
- Herrero, M. Male and female synchrony and the regulation of mating in flowering plants. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Postweiler, K.; Stösser, R.; Anvary, S.F. The effect of different temperatures on the viability of ovules in cherries. Sci. Hortic. 1985, 25, 235–239. [Google Scholar] [CrossRef]
- Cerović, R.; Ružić, Đ. Senescence of ovules at different temperatures and their effect on the behavior of pollen tubes in sour cherry. Sci. Hortic. 1992, 51, 321–327. [Google Scholar] [CrossRef]
- Stösser, R.; Anvary, S.F. Über die Lebensdauer von Samenanlagen in Bezichung zum Fruchtansatz beim Steinobst. Erwerbsobstbau 1990, 32, 134–137. [Google Scholar]
- Keulemans, J.; Van Laer, H. Effective pollination period of plums: The influence of temperature on pollen germination and pollen tube growth. In Manipulation of Fruiting; Wright, C.J., Ed.; Butterworths: London, UK, 1989; pp. 159–171. [Google Scholar]
- Burgos, L.; Egea, J. Apricot embryo-sac development in relation to fruit set. J. Hortic. Sci. 1993, 68, 203–208. [Google Scholar] [CrossRef]
- Cerović, R.; Fotirić Akšić, M.; Milena Ðorđević, M.; Meland, M. Functionality of embryo sacs in pear cultivars ‘Ingeborg’ and ‘Celina’ as related to fruit set under Nordic climate. Plants 2020, 9, 1716. [Google Scholar] [CrossRef] [PubMed]
- Sanzol, J.; Herrero, M. The “effective pollination period” in fruit trees. Sci. Hortic. 2001, 90, 1–17. [Google Scholar] [CrossRef]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. Global warming and sexual plant reproduction. Trends Plant Sci. 2009, 14, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Meier, U. Growth stages of mono- and dicotyledonous plants. In Federal Biological Research Centre for Agriculture and Forestry, 2nd ed.; BBCH Monograph; Blackwell Wissenschafts: Berlin, Germany, 2001; pp. 1–158. [Google Scholar]
- Gerlach, D. A rapid safranin-crystal violet-light green staining sequence for paraffin sections of plant materials. Stain Technol. 1969, 44, 210–211. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerović, R.; Akšić, M.F.; Đorđević, M.; Meland, M. Viability of Embryo Sacs and Fruit Set in Different Plum (Prunus domestica L.) Cultivars Grown under Norwegian Climatic Conditions. Plants 2022, 11, 219. https://doi.org/10.3390/plants11020219
Cerović R, Akšić MF, Đorđević M, Meland M. Viability of Embryo Sacs and Fruit Set in Different Plum (Prunus domestica L.) Cultivars Grown under Norwegian Climatic Conditions. Plants. 2022; 11(2):219. https://doi.org/10.3390/plants11020219
Chicago/Turabian StyleCerović, Radosav, Milica Fotirić Akšić, Milena Đorđević, and Mekjell Meland. 2022. "Viability of Embryo Sacs and Fruit Set in Different Plum (Prunus domestica L.) Cultivars Grown under Norwegian Climatic Conditions" Plants 11, no. 2: 219. https://doi.org/10.3390/plants11020219
APA StyleCerović, R., Akšić, M. F., Đorđević, M., & Meland, M. (2022). Viability of Embryo Sacs and Fruit Set in Different Plum (Prunus domestica L.) Cultivars Grown under Norwegian Climatic Conditions. Plants, 11(2), 219. https://doi.org/10.3390/plants11020219