

Metabolic Integration of Spectral and Chemical Cues Mediating Plant Responses to Competitors and Herbivores


Abstract
1. Introduction
2. Results
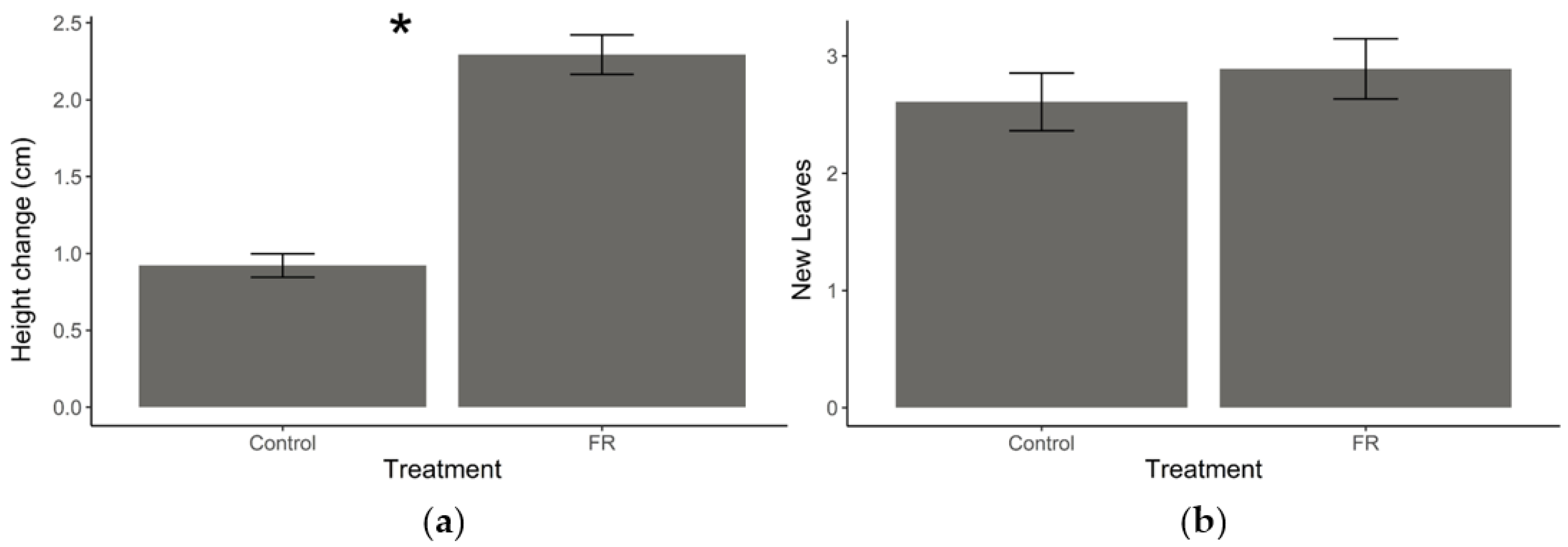
2.1. Effect of FR Light on Plant Growth
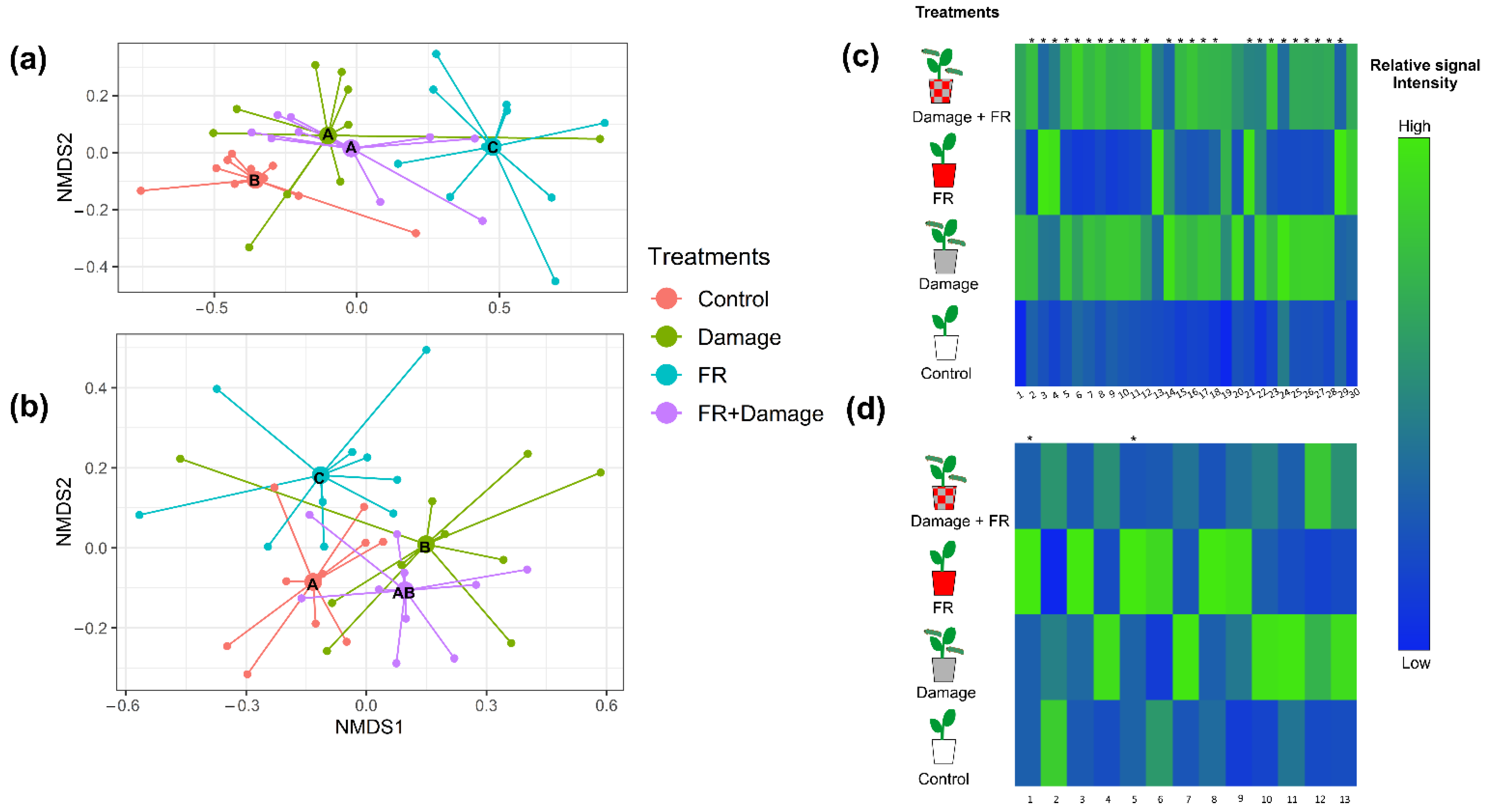
2.2. Effect of Herbivory and FR Radiation on Plant Chemistry
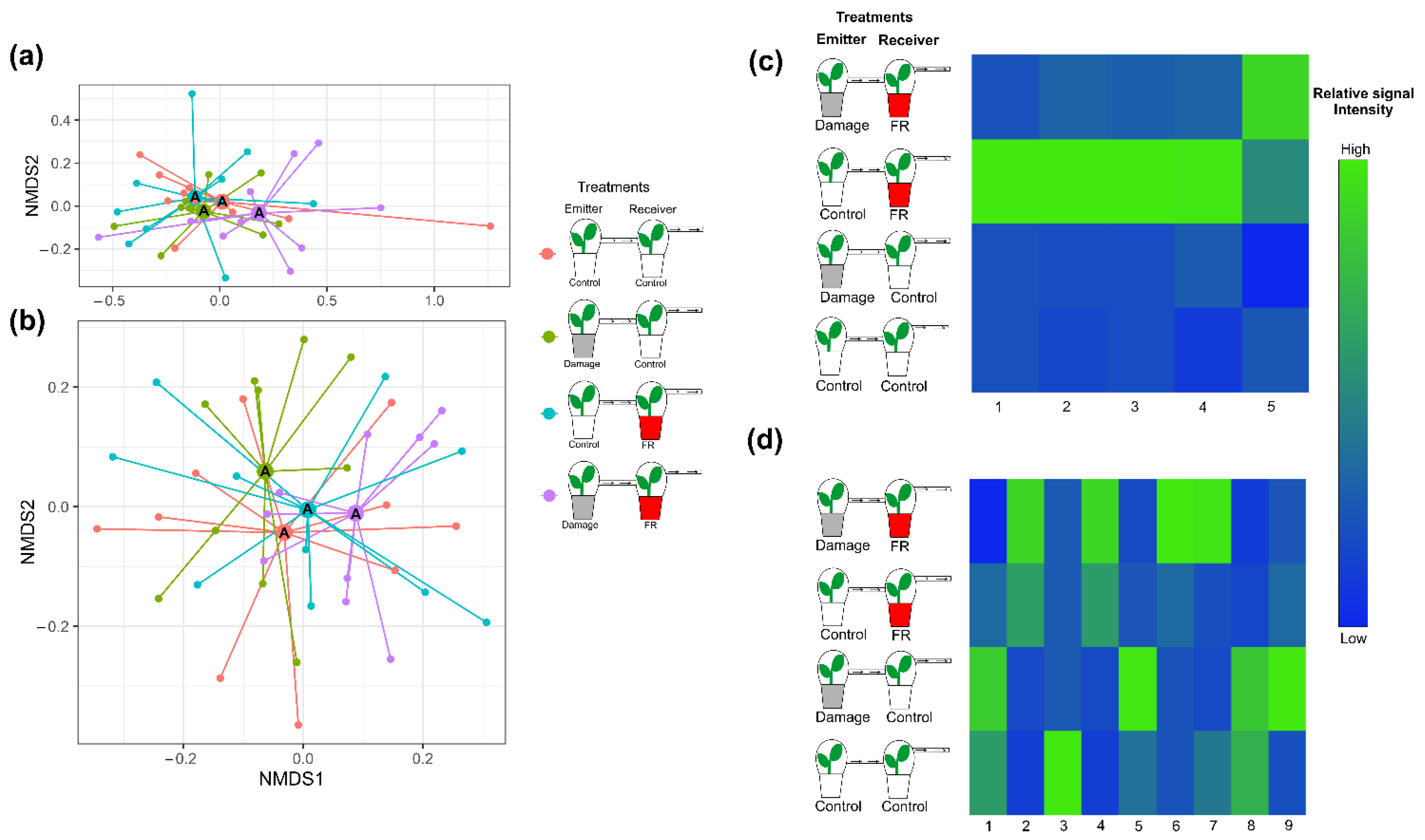
2.3. Effect of FR Light on the Perception of VOCs
3. Discussion
3.1. FR Light and Plant Growth
3.2. Effect of Increased FR Light on Constitutive and Herbivory-Induced Secondary Metabolism
3.3. Effect of Ingreased FR Light on the Perception of HIPVs from Neighbors
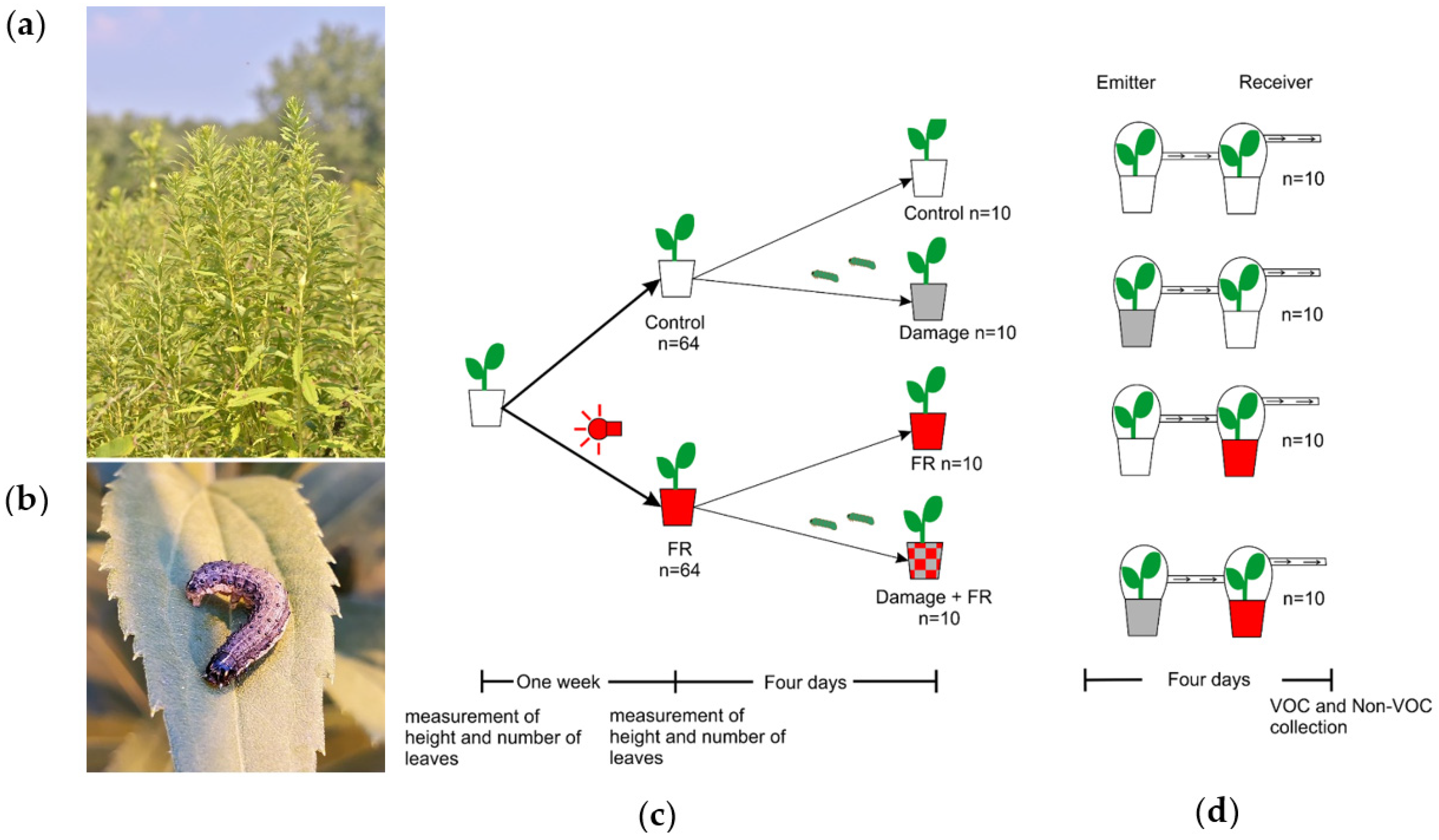
4. Materials and Methods
4.1. Plant Material
4.2. Secondary Metabolite Analysis
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, M.; Chory, J.; Fankhauser, C. Light Signal Transduction in Higher Plants. Annu. Rev. Genet. 2004, 38, 87–117. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R.F.; Takaki, M.; Azevedo, R.A. Plant Pigments: The Many Faces of Light Perception. Acta Physiol. Plant. 2011, 33, 241–248. [Google Scholar] [CrossRef]
- Khait, I.; Obolski, U.; Yovel, Y.; Hadany, L. Sound Perception in Plants. Semin. Cell Dev. Biol. 2019, 92, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Ton, J. Long-Distance Signalling in Plant Defence. Trends Plant Sci. 2008, 13, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A. The Information Landscape of Plant Constitutive and Induced Secondary Metabolite Production. Curr. Opin. Insect Sci. 2015, 8, 47–53. [Google Scholar] [CrossRef]
- Bae, H.; Mishra, R.C. Plant Cognition: Ability to Perceive ‘Touch’ and ‘Sound’. In Sensory Biology of Plants; Sopory, S., Ed.; Springer Nature: Singapore, 2019; pp. 137–162. ISBN 978-981-13-8921-4. [Google Scholar]
- Karban, R. Plant Behaviour and Communication. Ecol. Lett. 2008, 11, 727–739. [Google Scholar] [CrossRef]
- Smith, H. Physiological and Ecological Function within the Phytochrome Family. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 289–315. [Google Scholar] [CrossRef]
- De Wit, M.; Spoel, S.H.; Sanchez-Perez, G.F.; Gommers, C.M.M.; Pieterse, C.M.J.; Voesenek, L.A.C.J.; Pierik, R. Perception of Low Red: Far-Red Ratio Compromises Both Salicylic Acid- and Jasmonic Acid-Dependent Pathogen Defences in Arabidopsis. Plant J. 2013, 75, 90–103. [Google Scholar] [CrossRef]
- Rockwell, N.C.; Su, Y.S.; Lagarias, J.C. Phytochrome Structure and Signaling Mechanisms. Annu. Rev. Plant Biol. 2006, 57, 837–858. [Google Scholar] [CrossRef]
- Casal, J.J.; Sánchez, R.A.; Deregibus, V.A. The Effect of Light Quality on Shoot Extension Growth in Three Species of Grasses. Ann. Bot. 1987, 59, 1–7. [Google Scholar] [CrossRef]
- Cortés, L.E.; Weldegergis, B.T.; Boccalandro, H.E.; Dicke, M.; Ballaré, C.L. Trading Direct for Indirect Defense? Phytochrome B Inactivation in Tomato Attenuates Direct Anti-Herbivore Defenses Whilst Enhancing Volatile-Mediated Attraction of Predators. New Phytol. 2016, 212, 1057–1071. [Google Scholar] [CrossRef] [PubMed]
- Izaguirre, M.M.; Mazza, C.A.; Biondini, M.; Baldwin, I.T.; Ballaré, C.L. Remote Sensing of Future Competitors: Impacts on Plants Defenses. Proc. Natl. Acad. Sci. USA 2006, 103, 7170–7174. [Google Scholar] [CrossRef]
- Fernández-Milmanda, G.L.; Crocco, C.D.; Reichelt, M.; Mazza, C.A.; Köllner, T.G.; Zhang, T.; Cargnel, M.D.; Lichy, M.Z.; Fiorucci, A.S.; Fankhauser, C.; et al. A Light-Dependent Molecular Link between Competition Cues and Defence Responses in Plants. Nat. Plants 2020, 6, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Kegge, W.; Ninkovic, V.; Glinwood, R.; Welschen, R.A.M.; Voesenek, L.A.C.J.; Pierik, R. Red:Far-Red Light Conditions Affect the Emission of Volatile Organic Compounds from Barley (Hordeum Vulgare), Leading to Altered Biomass Allocation in Neighbouring Plants. Ann. Bot. 2015, 115, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, T.A.; Schwieterman, M.L.; Gilbert, J.L.; Jaworski, E.A.; Langer, K.M.; Jones, C.R.; Rushing, G.V.; Hunter, T.M.; Olmstead, J.; Clark, D.G.; et al. Light Modulation of Volatile Organic Compounds from Petunia Flowers and Select Fruits. Postharvest Biol. Technol. 2013, 86, 37–44. [Google Scholar] [CrossRef]
- Carvalho, S.D.; Schwieterman, M.L.; Abrahan, C.E.; Colquhoun, T.A.; Folta, K.M. Light Quality Dependent Changes in Morphology, Antioxidant Capacity, and Volatile Production in Sweet Basil (Ocimum Basilicum). Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Tegelberg, R.; Julkunen-Tiitto, R.; Aphalo, P.J. Red:Far-Red Light Ratio and UV-B Radiation: Their Effects on Leaf Phenolics and Growth of Silver Birch Seedlings. Plant Cell Environ. 2004, 27, 1005–1013. [Google Scholar] [CrossRef]
- Kuo, T.C.; Chen, C.; Chen, S.; Lu, I.; Chu, M.; Huang, L.; Lin, C.; Chen, C.; Lo, H.; Jeng, S.; et al. The Effect of Red Light and Far-Red Light Conditions on Secondary Metabolism in Agarwood. BMC Plant Biol. 2015, 15, 1–10. [Google Scholar] [CrossRef]
- Leone, M.; Keller, M.M.; Cerrudo, I.; Ballaré, C.L. To Grow or Defend? Low Red: Far-Red Ratios Reduce Jasmonate Sensitivity in Arabidopsis Seedlings by Promoting DELLA Degradation and Increasing JAZ10 Stability. New Phytol. 2014, 204, 355–367. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, Function and Metabolic Engineering of Plant Volatile Organic Compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Becker, C.; Desneux, N.; Monticelli, L.; Fernandez, X.; Michel, T.; Lavoir, A.V. Effects of Abiotic Factors on HIPV-Mediated Interactions between Plants and Parasitoids. Biomed Res. Int. 2015, 2015, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M.; Baldwin, I.T. The Evolutionary Context for Herbivore-Induced Plant Volatiles: Beyond the “Cry for Help”. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Karban, R.; Shiojiri, K.; Ishizaki, S. Plant Communication—Why Should Plants Emit Volatile Cues? J. Plant Interact. 2011, 6, 81–84. [Google Scholar] [CrossRef]
- Karban, R. Plant Communication System. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 1–24. [Google Scholar] [CrossRef]
- Okada, K.; Abe, H.; Arimura, G.I. Jasmonates Induce Both Defense Responses and Communication in Monocotyledonous and Dicotyledonous Plants. Plant Cell Physiol. 2015, 56, 16–27. [Google Scholar] [CrossRef]
- Morrell, K.; Kessler, A. Plant Communication in a Widespread Goldenrod: Keeping Herbivores on the Move. Funct. Ecol. 2017, 31, 1049–1061. [Google Scholar] [CrossRef]
- Kalske, A.; Shiojiri, K.; Uesugi, A.; Sakata, Y.; Morrell, K.; Kessler, A. Insect Herbivory Selects for Volatile-Mediated Plant-Plant Communication. Curr. Biol. 2019, 29, 3128–3133.e3. [Google Scholar] [CrossRef] [PubMed]
- Erb, M. Volatiles as Inducers and Suppressors of Plant Defense and Immunity—Origins, Specificity, Perception and Signaling. Curr. Opin. Plant Biol. 2018, 44, 117–121. [Google Scholar] [CrossRef]
- Heil, M. Herbivore-Induced Plant Volatiles: Targets, Perception and Unanswered Questions. New Phytol. 2014, 204, 297–306. [Google Scholar] [CrossRef]
- Gallie, D.R. Ethylene Receptors in Plants—Why so Much Complexity? F1000Prime Rep. 2015, 7, 1–12. [Google Scholar] [CrossRef]
- Sugimoto, K.; Matsui, K.; Iijima, Y.; Akakabe, Y.; Muramoto, S.; Ozawa, R.; Uefune, M.; Sasaki, R.; Alamgir, K.M.; Akitake, S.; et al. Intake and Transformation to a Glycoside of (Z)-3-Hexenol from Infested Neighbors Reveals a Mode of Plant Odor Reception and Defense. Proc. Natl. Acad. Sci. USA 2014, 111, 7144–7149. [Google Scholar] [CrossRef] [PubMed]
- Etterson, J.R.; Delf, D.E.; Craig, T.P.; Ando, Y. NOTE/NOTE Parallel Patterns of Clinal Variation in Solidago altissima in Its Native Range in Central USA and Its Invasive Range in Japan. Botany 2008, 97, 91–97. [Google Scholar] [CrossRef]
- Howard, M.M.; Kalske, A.; Kessler, A. Eco-Evolutionary Processes Affecting Plant–Herbivore Interactions during Early Community Succession. Oecologia 2018, 187, 547–559. [Google Scholar] [CrossRef]
- Maddox, G.D.; Root, R.B. Resistance to 16 Diverse Species of Herbivorous Insects within a Population of Goldenrod, Solidago altissima: Genetic Variation and Heritability. Oecologia 1987, 72, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Maddox, G.D.; Root, R.B. Structure of the Encounter between Goldenrod (Solidago altissima) and Its Diverse Insecr Fauna. Ecology 1990, 71, 2115–2124. [Google Scholar] [CrossRef]
- Carson, W.P.; Root, R.B. Herbivory and Plant Species Coexistence: Community Regulation by an Outbreaking Phytophagous Insect. Ecol. Monogr. 2000, 70, 73–99. [Google Scholar] [CrossRef]
- Bode, R.F.; Kessler, A. Herbivore Pressure on Goldenrod (Solidago altissima L., Asteraceae): Its Effects on Herbivore Resistance and Vegetative Reproduction. J. Ecol. 2012, 100, 795–801. [Google Scholar] [CrossRef]
- Uesugi, A.; Kessler, A. Herbivore Exclusion Drives the Evolution of Plant Competitiveness via Increased Allelopathy. New Phytol. 2013, 198, 916–924. [Google Scholar] [CrossRef]
- Rubin, I.N.; Ellner, S.P.; Kessler, A.; Morrell, K.A. Informed Herbivore Movement and Interplant Communication Determine the Effects of Induced Resistance in an Individual-Based Model. J. Anim. Ecol. 2015, 84, 1273–1285. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant Responses to Red and Far-Red Lights, Applications in Horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Fankhauser, C.; Batschauer, A. Shadow on the Plant: A Strategy to Exit. Cell 2016, 164, 15–17. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M.; Kurepin, L.V.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M. Uncoupling Light Quality from Light Irradiance Effects in Helianthus Annuus Shoots: Putative Roles for Plant Hormones in Leaf and Internode Growth. J. Exp. Bot. 2007, 58, 2145–2157. [Google Scholar] [CrossRef] [PubMed]
- Pierik, R.; Ballaré, C.L.; Dicke, M. Ecology of Plant Volatiles: Taking a Plant Community Perspective. Plant Cell Environ. 2014, 37, 1845–1853. [Google Scholar] [CrossRef]
- Cipollini, D.; Lieurance, D.M. Expression and Costs of Induced Defense Traits in Alliaria Petiolata, a Widespread Invasive Plant. Basic Appl. Ecol. 2012, 13, 432–440. [Google Scholar] [CrossRef]
- Mason, H.S.; Mullet, J.E. Expression of Two Soybean Vegetative Storage Protein Genes during Development and in Response to Water Deficit, Wounding, and Jasmonic Acid. Plant Cell 1990, 2, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Mason, H.S.; DeWald, D.B.; Creelman, R.A.; Mullet, J.E. Coregulation of Soybean Vegetative Storage Protein Gene Expression by Methyl Jasmonate and Soluble Sugars. Plant Physiol. 1992, 98, 859–867. [Google Scholar] [CrossRef]
- Dewald, D.B.; Sadka, A.; Mullet, J.E. Sucrose Modulation of Soybean Vsp Gene Expression Is Inhibited by Auxin. Plant Physiol. 1994, 104, 439–444. [Google Scholar] [CrossRef][Green Version]
- Thornburg, R.W.; Li, X. Wounding Nicotiana Tabacum Leaves Causes a Decline in Endogenous Indole-3-Acetic Acid. Plant Physiol. 1991, 96, 802–805. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Baldwin, I.T.; Zhang, Z.P.; Diab, N.; Ohnmeiss, T.E.; McCloud, E.S.; Lynds, G.Y.; Schmelz, E.A. Quantification, Correlations and Manipulations of Wound-Induced Changes in Jasmonic Acid and Nicotine in Nicotiana Sylvestris. Planta 1997, 201, 397–404. [Google Scholar] [CrossRef]
- Cooper-Driver, G.A.; Le Quesne, P.W. Diterpenoids as Insect Antifeedants and Growth Inhibitors: Role in Solidago Species. In Allelochemicals: Role in Agriculture and Forestry; Waller, G.R., Ed.; American Chemical Society: Washington, DC, USA, 1987; pp. 534–550. ISBN 9780841209923. [Google Scholar]
- Uesugi, A.; Kessler, A. Herbivore Release Drives Parallel Patterns of Evolutionary Divergence in Invasive Plant Phenotypes. J. Ecol. 2016, 104, 876–886. [Google Scholar] [CrossRef]
- Kessler, A.; Halitschke, R.; Diezel, C.; Baldwin, I.T. Priming of Plant Defense Responses in Nature by Airborne Signaling between Artemisia Tridentata and Nicotiana Attenuata. Oecologia 2006, 148, 280–292. [Google Scholar] [CrossRef]
- Crepy, M.A.; Casal, J.J. Photoreceptor-Mediated Kin Recognition in Plants. New Phytol. 2015, 205, 329–338. [Google Scholar] [CrossRef]
- Semchenko, M.; Saar, S.; Lepik, A. Plant Root Exudates Mediate Neighbour Recognition and Trigger Complex Behavioural Changes. New Phytol. 2014, 204, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Kalske, A. Plant Secondary Metabolite Diversity and Species Interactions. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 115–138. [Google Scholar] [CrossRef]
- Mertens, D.; Boege, K.; Kessler, A.; Koricheva, J.; Thaler, J.S.; Whiteman, N.K.; Poelman, E.H. Predictability of Biotic Stress Structures Plant Defence Evolution. Trends Ecol. Evol. 2021, 36, 444–456. [Google Scholar] [CrossRef]
- Barbosa, P.; Hines, J.; Kaplan, I.; Martinson, H.; Szczepaniec, A.; Szendrei, Z. Associational Resistance and Associational Susceptibility: Having Right or Wrong Neighbors. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 1–20. [Google Scholar] [CrossRef]
- Lawson, S.K.; Sharp, L.G.; Powers, C.N.; McFeeters, R.L.; Satyal, P.; Setzer, W.N. Volatile Compositions and Antifungal Activities of Native American Medicinal Plants: Focus on the Aster0aceae. Plants 2020, 9, 126. [Google Scholar] [CrossRef]
- Martinez Arbizu, P. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis, Version 0.4; 2020. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis#readme (accessed on 11 September 2022).
- R Team Core. R: A Language and Environment for Statistical Computing, Version 2.6.2; R Foundation for Statistical Computing: Vienna, Austria, 2021.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Control | Damage | FR |
|---|---|---|---|
| Damage | F1,18 = 3.872, p = 0.001 * | ||
| FR | F1,18 = 19.12, p = 0.001 * | F1,18 = 7.85, p = 0.001 * | |
| FR + Damage | F1,18 = 5.70 p = 0.001 * | F1,18 = 0.86, p = 0.283 | F1,18 = 7.7928, p = 0.001 * |
| Treatment | Control | Damage | FR |
|---|---|---|---|
| Damage | F1,18 = 2.5411, p = 0.03 * | ||
| FR | F1,18 = 7.851, p = 0.003 * | F1,18 = 3.75, p = 0.001 * | |
| FR + Damage | F1,18 = 1.33, p = 0.153 | F1,18 = 0.887, p = 0.461 | F1,18 = 4.7507, p = 0.002 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chautá, A.; Kessler, A. Metabolic Integration of Spectral and Chemical Cues Mediating Plant Responses to Competitors and Herbivores. Plants 2022, 11, 2768. https://doi.org/10.3390/plants11202768
Chautá A, Kessler A. Metabolic Integration of Spectral and Chemical Cues Mediating Plant Responses to Competitors and Herbivores. Plants. 2022; 11(20):2768. https://doi.org/10.3390/plants11202768
Chicago/Turabian StyleChautá, Alexander, and André Kessler. 2022. "Metabolic Integration of Spectral and Chemical Cues Mediating Plant Responses to Competitors and Herbivores" Plants 11, no. 20: 2768. https://doi.org/10.3390/plants11202768
APA StyleChautá, A., & Kessler, A. (2022). Metabolic Integration of Spectral and Chemical Cues Mediating Plant Responses to Competitors and Herbivores. Plants, 11(20), 2768. https://doi.org/10.3390/plants11202768