Abstract
Plant cell wall remodeling is an important process during plant responses to heat stress. Pectins, a group of cell wall polysaccharides with a great diversity of complex chemical structures, are also involved in heat stress responses. Enzymatic activity of the pectin methyl esterases, which remove methyl groups from pectins in the cell wall, is regulated by DUF642 proteins, as described in different plants, including Arabidopsis thaliana and Oryza sativa. Our results demonstrated that heat stress altered the expression of the DUF642 gene, BIIDXI. There was an important decrease in BIIDXI expression during the first hour of HS, followed by an increase at 24 h. bdx-1 seedlings had less tolerance to heat stress but presented a normal heat stress response; HSFA2 and HSP22 expressions were highly increased, as they were in WT seedlings. Thermopriming triggered changes in pectin methyl esterase activity in WT seedlings, while no increases in PME activity were detected in bdx-1 seedlings at the same conditions. Taken together, our results suggest that BIIDXI is involved in thermotolerance via PME activation.
1. Introduction
The cell wall has a complex structure, formed mainly by the polysaccharide cellulose, hemicelluloses and pectins, and by proteins from different families. The cell wall actively participates during the different stages of plant development and in response to abiotic and biotic stresses [1]. High temperatures induce heat stress by promoting important alterations in plant growth and development [2]. Plants exposed to sublethal temperatures acquire thermotolerance, a mechanism that allows survival at higher temperatures that are normally lethal. Under laboratory conditions, thermotolerance acquisition can be induced using thermopriming treatment. In addition to the accumulation of heat shock proteins (HSPs), plant cell wall remodeling has also been considered as an integral part of the heat response network, as it was recently demonstrated for salt-induced responses [3,4]. In the mold Aspergillus fumigatus, cell wall stress generated by heat stress actively participates in the perception and signaling of stress, triggering the coordinated expression of heat shock transcription factors (HSFs) and HSPs [5].
Heat stress may also promote mechanisms to resist recurrent heat stress events, i.e., thermomemory. The participation of HSFs, HSPs, primary metabolism enzymes, growth regulators, and epigenetic processes in thermomemory has been widely described [2]. The involvement of plant cell wall remodeling processes in thermomemory has not yet been described but is suggested by the upregulation of plant cell wall protein-encoding genes by the transcription factor HSFA2 [6].
The plant cell wall polysaccharides known as pectins are the most sensitive to the mechanical deformation that occurs in plant cells during the different stages of development and during the responses to biotic and abiotic stresses [3,7]. Pectins have a great diversity of complex chemical structures. Among the five different groups of pectins, homogalacturonans (HGs) are the most abundant. These polysaccharides are linear chains of α-1,4-linked D-galacturonic acid (DGalA) residues whose chemical properties, determined by methyl esterification and acetyl esterification, may be influenced by development and environmental conditions [8].
Highly methyl-esterified HGs are secreted into the cell wall, where their degree of esterification is determined by the activity of pectin methyl esterases (PMEs). The removal of the methyl group by PMEs determines the degree of methylation of pectins, promoting both relaxation and increased rigidity of the cell wall: the de-methyl-esterified HGs chains can be the substrate for enzymes such as polygalacturonases (PGs) and pectate lyases (PLs) that promote cell wall relaxation; likewise, the acid groups present in the de-methyl-esterified HG chains can interact with calcium ions found in the cell wall, forming structures called “egg boxes” that confer greater rigidity to the wall [9]. The regulation of the degree of pectin-methyl esterification present in the cell wall is determined by the combined activity of PMEs and the proteins that regulate their activity: the pectin methyl esterase inhibitors (PMEIs) and the DUF642 family proteins [10,11].
PMEs constitute a multigene family in plants that are involved in different physiological processes during plant development. In Arabidopsis thaliana, there are 66 PME-related genes with differential expressions during plant development [12]. In A. thaliana seedlings, decreases in gene expression of some PMEs resulted from exposure to heat shock, and the PME7, PME28, PME34, and PME53 were observed to participate in the thermotolerance process. The seedlings of the pme7, pme28, and pme34 mutant lines have a higher sensitivity to HS [13], while the seedlings of the pme53 mutant line have a higher thermotolerance [14]. The increase in temperature promotes an increase in PME activity that participates in cell wall remodeling in response to heat stress, a crucial process for the acquisition of thermotolerance [3,13,15]. The increase in PME activity could be a result of their regulation through other interacting proteins, as has been suggested by Bosch and Hepler [16]; PME enzymatic activity regulation is likely more crucial than the regulation at the translation or protein accumulation level.
PMEI proteins were first identified in kiwi [17], and ever since, they have been reported to modulate PME activity in a wide variety of biological contexts, participating in cell adhesion, cell wall stress signaling, and defense, among other processes [18]. The participation of these proteins has been described in response to cold and salinity stress [19], but they are poorly studied during heat stress response, despite previous suggestions by Wu et al. [3] that specific PMEIs might be involved in the regulation of PME34 activity during this process.
The DUF642 family is a family of cell wall proteins specific to spermatophyte plants. In Arabidopsis, 10 genes have been described for this family that are grouped into four different clades (A-D) and are differentially expressed during plant development [11,20]. DUF642 proteins have been described in cell wall proteomes of different tissues in Arabidopsis and other plants [21]. The DUF642 proteins encoded by the genes At4g32460 (BIIDXI) and At5g11420 interact in vitro with the AtPME3 (pectin methyl esterase 3, At3g14310) protein [22]. The DUF642–PME protein interaction has been verified for the rice DUF642 protein, named DFOT1, and the homologue of PME3 [23]. Different tissues in the BIIDXI and At2g41800 (TEEBE) loss-of-function and overexpression lines show alterations in PME activity. In seedlings, the bdx-1 and teb-1 mutants show a decrease in PME activity, while OEBDX and GCTEB overexpression lines show an increase [24]. For bdx-1 and teb-1, a decrease in the detection of demethyl-esterified pectins in the seed endosperm and in the epidermal cells of the hypocotyl, respectively, has also been described [25,26]. In rice, the dfot 1 mutant showed a lower degree of methyl esterification in the cell wall of the lodicule cells, a structure involved in the opening of the flower [23].
The increase in PME activity in response to high temperature determines a remodeling of the cell wall that could participate in the perception and signaling of heat stress [3,27]. To address the role of the DUF642 proteins in the processes of thermotolerance and thermomemory through the regulation of PME activity, we first investigated the effect of different temperatures on the expression levels of three DUF642 genes: At3g08030, BIIDXI, and DGR2 (DUF642 L-GALL RESPONSIVE GENE 2 (At5g25460)). We found that bdx-1 seedlings are more sensitive to heat, and the dynamics of total PME activity during treatment is different from that of WT plants. Our results provide evidence that the DUF642 family proteins could be involved in the regulation of PME activity under heat stress.
2. Results
2.1. BIIDXI Expression Decreases during Thermopriming
Heat stress decreased the expression levels of DUF642 genes from peach fruits, and a decrease in the accumulation of DUF642 proteins in peach fruits and Brachipodium leaves in response to heat was also reported [28]. To investigate if DUF642 genes from A. thaliana could be involved in thermotolerance acquisition and/or in thermomemory (Figure 1A), we first determined the expression levels of At3g08030, BIIDXI, and At5g25460, the DUF642 genes that are highly expressed in seedlings (Figure S1 and Figure 1B). No differences in expression levels of At3g08030 and At5g25460 were recorded during thermotolerance and thermomemory, described in Figure 1A. The BIIDXI gene expression level decreased in response to heat stress at 37 °C, with no further increase throughout the thermopriming phase (Figure 1B). At 24 h after priming, an increase in BIIDXI expression was observed (Figure 1C).
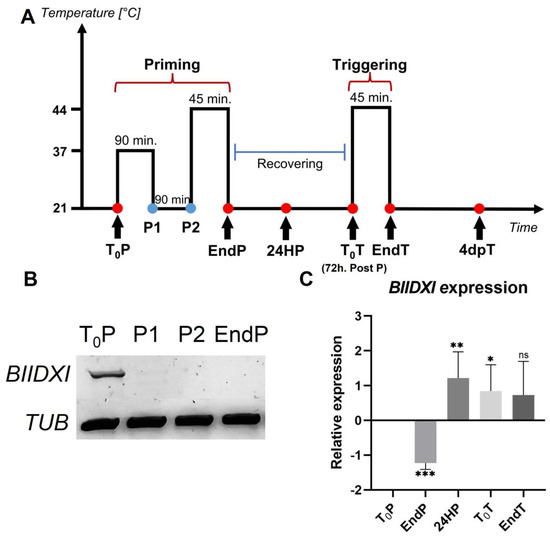
Figure 1.
BIIDXI expression during thermotolerance and thermomemory. (A) Schematic representation of the protocol (based on Olas et al. [29]). Seedlings were collected and frozen immediately at the beginning and at the end of priming (T0P and EndP), 24 h after the end of priming (24HP), 72 h after priming/start of triggering (T0T), at the end of triggering (EndT), and 4 days after triggering (4dpT). (B) BIIDXI expression within the priming, including sublethal heat treatment (C) on the BIIDXI expression level during triggering. The expression values are relative to T0P expression. Error bars indicate S.D. (n = 4). Statistical analyses were performed using Student’s t-test. Asterisks indicate statistically significant differences to T0P (* p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant).
2.2. The bdx-1 Mutant Is Affected by Thermopriming Treatment
The differential effects of priming and triggering on BIIDXI gene expression suggest that this gene could be involved in thermotolerance and/or thermomemory processes, so the effect of both treatments on the development of WT, overexpression (OEBDX) transgenic lines, and loss-of-function transgenic line seedlings (bdx-1) were performed. The detrimental effects of priming and triggering on seedling development were quantified as the percentage of cotyledon bleaching in seedlings (Figure 2). After three days of priming treatment, 80% of the bdx-1 seedlings showed cotyledon bleaching, a significantly higher percentage than WT and OEBDX seedlings, which did not show significant differences between them (Figure 2A,B). Leaf number in bdx-1 seedlings was also significantly lower after priming, in contrast to untreated seedlings (7d post priming). Triggering did not affect the percentage of cotyledon bleaching in any of the three lines, but a lower growth of rosette leaves was observed.
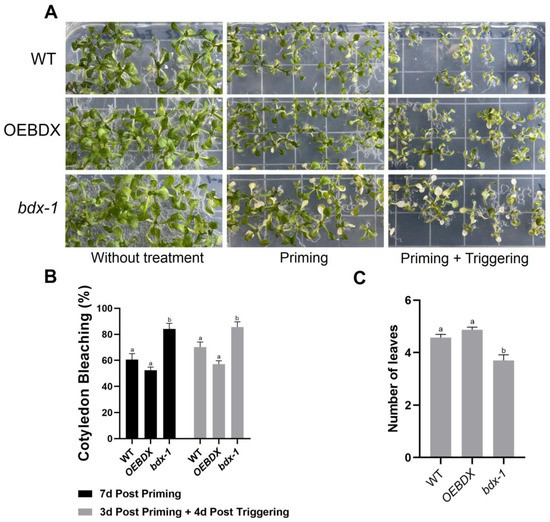
Figure 2.
Phenotypic changes of WT, bdx-1, and OEBDX seedlings during thermotolerance and thermomemory. (A) WT Col-0, bdx-1, and OEBDX seedlings were grown on MS medium. Seedlings with two leaves were subjected to priming treatment (T0P). Seedling phenotypes of control, 7d post priming and 3d post priming, and 4d post triggering are shown. (B) The bleaching cotyledon phenotype was quantified in seedlings shown in (A) and represented as a percentage. (C) Leaves observed at 4d post triggering. Error bars indicate SEM (n = 3). An amount of 10–12 seedlings constituted each biological replicate. For cotyledon bleaching, statistical analyses were performed using arcsine conversion of percentages. Different letters indicate significant differences calculated using one-way ANOVA with post-hoc Tukey’s test (p < 0.05).
To further investigate the effect of priming on seedling development and because of previous reports indicating that root growth is reduced in seedlings grown at 30 °C for 3–4 days [30], we determined the root length of thermoprimed seedlings. We observed that heat reduced root growth in WT, OEBDX, and bdx-1, but the length of the bdx-1 roots after the thermopriming was significantly reduced, with respect to the treated WT and OEBDX lines. At 24 h after priming (24HP), no difference in root length between lines was observed; at 72 h after priming (72HP), there was a significant increase in root length that was not observed in bdx-1 roots (Figure 3). These results suggest that BIIDXI is involved in thermotolerance acquisition.
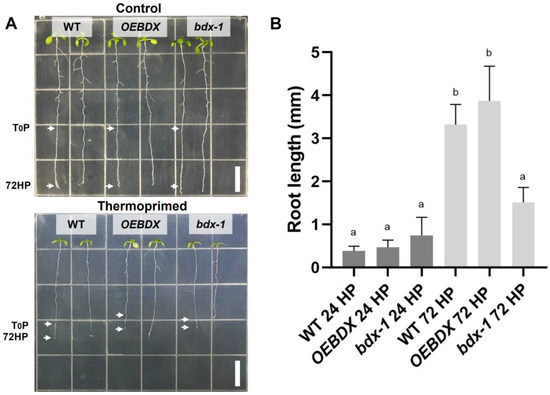
Figure 3.
Effect of thermopriming treatment on root growth. (A) Seedlings of WT, OEBDX, and bdx-1 were sown vertically on MS medium. The seedlings were exposed to priming treatment, and the comparisons of the root lengths between the lines were performed 72 h after priming. (B) Root length average after 24 and 72 h of priming. Error bars indicate SEM (n = 3). An amount of 30 seedlings constituted each biological replicate. Different letters indicate significant differences calculated using one-way ANOVA with post-hoc Tukey’s test (p < 0.05). White bars = 10 mm.
2.3. BIIDXI Participates in the Regulation of PME Activity in Thermotolerance
The proteins of the DUF642 family participate in different stages of plant development, mainly by increasing the PME activity [11,23]. In vitro interaction analyses indicate that a possible target is AtPME3, a PME that is highly expressed in seedlings [22]. Heat stress promotes an increase in total PME activity that is involved in cell wall remodeling, which is crucial for thermotolerance [3]. The total PME activity of T0P seedlings from bdx-1 corresponded to 50% of the total PME activity of WT seedlings at T0P (Figure 4A), although there were no differences in AtPME3 expression levels. To investigate if the bdx-1 heat sensitivity phenotype was related to PME activity modulation, we determined the total PME activity of the WT and bdx-1 seedlings during thermopriming and thermotriggering treatments (Figure 4). The difference in PME activity between WT and bdx-1 seedlings was observed during both treatments, except for 24 h after priming treatment; the decrease in PME activity detected in WT was similar to the PME activity levels in 24HP seedlings. As it was previously described [13], priming promoted a significant increase in total PME activity in WT seedlings. This increase was not detected in bdx-1 seedlings (Figure 4A). As expected, triggering also promoted an increase in PME activity in WT seedlings (Figure 4A). In seedlings, triggering treatment also promoted an increase in PME activity; this activity level was equivalent to the T0P seedling from WT. These results suggest that BIIDXI is mainly involved in the increase in PME activity caused by thermopriming.
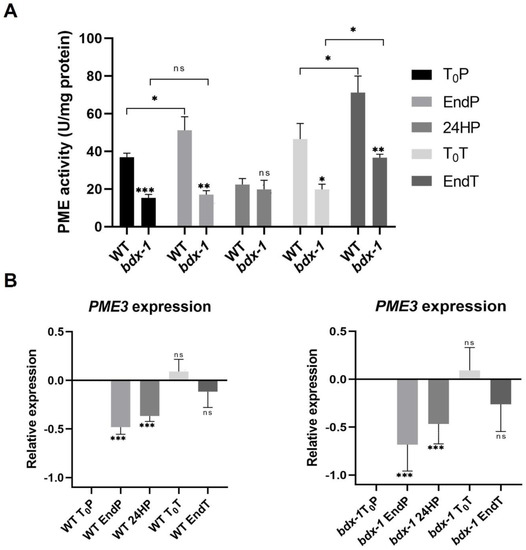
Figure 4.
PME activity and AtPME3 expression of WT and bdx-1 lines during thermotolerance and thermomemory. (A) Effect of priming and triggering on the total PME activity of WT and bdx-1 seedlings. Error bars indicate SEM (n = 4). Statistical analyses were performed using Student’s t-test. Asterisks indicate statistically significant differences between WT and bdx-1 and between treatments (* p < 0.05, ** p < 0.01, *** p < 0.001). (B) Effect of priming and triggering on expression levels of AtPME3 in WT and bdx-1 seedlings. The expression values are relative to T0P expression. Error bars indicate S.D. (n = 3). Statistical analyses were performed using Student’s t-test. Asterisks indicate statistically significant differences to T0P (* p < 0.05, ** p < 0.01, *** p < 0.001, ns, not significant).
In general, it has been described that the expression levels of different PME isoforms decrease in response to heat shock or thermopriming treatment [13]. AtPME3 expressions in response to priming and triggering in WT and bdx-1 seedlings exhibit similar dynamics (Figure 4B). The AtPME3 expression decreased because of priming until after 24 h. At 72 h of treatment, the expression levels had recovered, and the triggering treatment did not affect this expression level.
2.4. The HSFA2-Mediated Signaling Pathway Is Activated in bdx-1 Seedlings
It has been proposed that PMEs and consequently, cell wall remodeling in response to heat shock, could activate the signaling pathway involved in heat shock response [3]; thus, we determined the expressions of HSFA2 and HSP22 during priming and triggering treatments on the WT and bdx-1 lines. HSFA2 expression increased markedly in both lines in response to priming (Figure 5A). HSP22 expression increased in response to priming in both lines and remained at high levels up to 72 h after treatment. Triggering slightly promoted expressions in the WT and bdx-1 lines (Figure 5B).
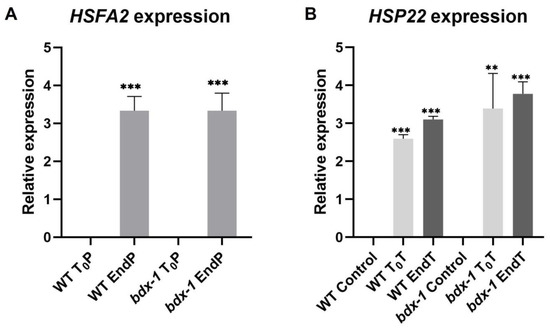
Figure 5.
Expression of thermomemory genes in the bdx-1 line. (A) HSFA2 expression at the end of priming compared to T0P in each line. The expression level is similar in WT and bdx-1. (B) HSP22 expression at the beginning and the end of triggering, compared with plants without treatment. For HSFA2, the expression values are relative to T0P expression and for HSP22, to WT control. Error bars indicate S.D. (n = 3). Statistical analyses were performed using Student’s t-test. Asterisks indicate statistically significant differences to T0P and to WT control (** p < 0.01, *** p < 0.001).
3. Discussion
The cell walls of different plant cells have complex and dynamic structures that actively participate in the perception of environmental conditions and in the promotion of cellular responses [1]. Pectin modifications could be involved in the response to different abiotic stresses such as salinity, osmostic, heat, and cold, although the mechanisms involved in perceiving the cell wall damage are not yet known. It has been proposed that salt and heat stresses influence the mechanical properties of the cell wall, modifying the pectin degree of esterification [27]. Recently, Gigli–Bisceglia et al. [4] reported that PME activation by salt is a requirement for the activation of salt-induced responses. In our study, we focused on investigating the function of the DUF642 protein BIIDXI during thermotolerance acquisition and in thermomemory. BIIDXI gene expression decreased when the seedlings were subjected to 37 °C and to 44 °C during the priming treatment (Figure 1B). In tissues of different plants, it has been determined that heat shock inhibits the gene expression of the DUF642 family [27,31,32]. BIIDXI expression increased 24 h after priming (Figure 1C). This increase in expression, unaffected by the second heat shock, could be regulated by HSFA2, a heat stress response factor involved in maintaining a high expression of genes, over several days; it could be involved in thermomemory [33]. The promoter region of BIIDXI presents several regulatory regions related to the response to heat stress, and the positive regulation of the expression of a gene of the DUF642 family by HSFA2 has been previously described in tomato [6].
BIIDXI participates in seed development and germination by modifying the pectin degree of esterification by promoting PME activity. An abnormal embryo elongation, resulting from a high methyl esterification of pectins in the cell wall of endosperm cells, was observed in approximately 40% of bdx-1 seeds. These misshapen seeds present altered germination and seedling development [24,25]. For this study, we used seedlings from normal seeds. These bdx-1 seedlings presented a high sensitivity to the priming treatment, determined by a higher percentage of seedlings with cotyledon bleaching and by the retarded growth of the main root (Figure 2 and Figure 3). An increase in the testa rupture rate was observed in OEBDX seeds, but seedling development was not affected. No difference in heat sensitivity was observed (Figure 2). The demethyl esterification rate of PME activity was increased substantially with increasing temperature, but the mechanism for temperature activation is less understood [3]. WT seedlings showed an increase in PME activity because of priming, as previously described (Figure 4; [13]), while at 24 h after treatment, the activity dropped considerably to similar levels determined in T0P seedlings from the bdx-1 mutant. This decrease in PME activity observed in WT seedlings could be related to the decrease in BIIDXI expression resulting from priming treatment (Figure 1B). bdx-1 seedlings did not show any changes in PME activity during priming; the PME activity peak present in WT seedlings was not observed (Figure 4). Heat shock also causes decreases in the expressions of different PMEs. In winter oilseed rape, heat shock induced a nearly 10-fold reduction in PME35 gene expression [34]. In Arabidopsis, the expressions of AtPM34, AtPM28, and AtPM7 were reduced at the end of the priming treatment but recovered after 3 h [13]. In the current study, priming promoted a decrease in the expression of AtPME3, the PME isoform that interacts in vitro with BIIDXI, in both WT and bdx-1 seedlings (Figure 4). Huang et al. [13] suggested that a regulator of PME activity might be involved in the increased activity during the response to heat stress. Our results support the hypothesis that BIIDXI is involved in the acquisition of thermotolerance by promoting an increase in PME activity. Furthermore, the mechanism by which BIIDXI participates in the response to heat is through promoting PME activity; this was supported by the identification of the thermotolerance impairment of Arabidopsis bdx-1 mutant plants, which was independent from the expression of heat stress-responsive genes. The gene expression patterns of heat-responsive genes in bdx-1, such as HSFA2 and HSP22, were shown to be similar to those of WT plants (Figure 5).
In response to triggering, PME activity was significantly increased in WT seedlings, while AtPME3 and BIIDXI expression levels were not affected (Figure 4). These results suggest that BIIDXI could also be involved in thermomemory by promoting cell wall remodeling. The relevance of cell wall remodeling in thermomemory is also suggested by the HSFA2 upregulation of the expression of a high number of genes involved in cell wall dynamics, including several PMEs [6]. However, triggering also promoted a PME activity increase in bdx-1, suggesting that in thermomemory, there could be alternate mechanisms to promote PME activation in the absence of BIIDXI. These results can be partially explained by the maintenance of AtPME3 expression levels during triggering treatment (Figure 4).
The participation of cell wall proteins of the DUF642 family in plant development, especially in the response to stress, has been poorly studied. However, studies in Arabidopsis and rice indicate that this family of proteins actively participates in increasing the degree of demethyl esterification of pectins in the cell wall of different cell types by promoting PME activity, although the biochemical mechanism has not been described. The results obtained suggest that BIIDXI participates in the response to heat stress by the activation of PMEs, a process necessary for the acquisition of thermotolerance.
4. Methods
4.1. Plant Growth Conditions
Arabidopsis thaliana seeds of the WT (Col-0), bdx-1, and OEBDX lines were sown on sterile Murashige and Skoog (MS) medium with 1% sucrose and a pH of 5.7. Seeds were stratified at 4 °C for 48 h and subsequently placed in a CONVIRON growth chamber (Winnipeg, Canada) at 22 °C under long photoperiod conditions (16 h light; 8 h dark).
Transgenic Plants
bdx-1: 142260 SALK line with the T-DNA insertion at the end of the second exon of BIIDXI. In the homozygous line, the expression of BIIDXI was approximately reduced to a 60% [24].
OEBDX: Transgenic plants were transformed with a pBIN vector that contained the full-length coding BIIDXI sequence under the control of the cauliflower mosaic virus 35S promoter [24]. Expression of BIIDXI was highly increased (1000-fold change) in the homozygous transgenic lines [35].
4.2. Thermotolerance and Thermomemory Analyses
Thermopriming and thermotriggering treatments were performed based on Olas et al., [29] and is briefly described as follows: Seedlings with two full leaves were subjected to priming stimulus at 6 h after dawn (90 min at 37 °C in a water bath; recovery at 22 °C for 90 min in the growth chamber; 45 min at 44 °C in a water bath), then returned to normal growth conditions (22 °C) for 72 h of the recovery phase, and then subjected to the triggering treatment (45 min at 44 °C) (Figure 1A). To have enough material for subsequent analyses, the treatment time was reduced to promote seedling survival, especially for bdx-1 seedlings.
Whole seedling samples were collected and frozen immediately at the beginning and at the end of priming (T0P and EndP), 24 h after the end of priming (24HP), 72 h after priming/start of triggering (T0T), at the end of triggering (EndT), and 4 days after triggering (4dpT).
The analysis of the percentage of seedlings with cotyledon bleaching in the lines WT, bdx-1, and OEBDX after priming and triggering was carried out in three independent biological replicates with 10–12 seedlings. The effects of priming on root lengths were also performed in the lines WT, bdx-1, and OEBDX in three independent biological replicates with 30 seedlings.
All the data analyses were performed using GraphPad Prism version 8.0.0 for windows, Graph Pad software, San Diego, CA, USA, www.graphpad.com.
4.3. RNA Isolation, cDNA Synthesis, and Quantitative RT-PCR
Total RNA was extracted from 50 mg of collected tissue at each thermomemory treatment point of WT and bdx-1 plants using the TRIzol extraction technique (Invitrogen) according to the manufacturer’s recommendations. RNA was quantified by means of a NanoDrop Lite spectrophotometer (Thermo Fisher Scientific). The cDNA synthesis was performed from 200 ng of total RNA using the Im Prom-II Reverse Transcription System kit (Promega Corporation). The qRT-PCR measurements were performed using SYBR Green Master Mix and detected with an Applied Biosystems StepOne platform (Applied Biosystems). Three or four independent biological replicates with three technical replicates were performed, and either TUB2 or ACT7 was used as endogenous controls using the primers listed in Table S1. The specificity of BIIDXI primers was confirmed by cloning and sequencing different clones [24]. In silico analysis also confirmed the primer specificity (Table S2).
Expression analyses were performed using the method described by [36]. Student’s t-tests were performed using GraphPad Prism version 8.0.0 for windows, Graph Pad software, San Diego, CA, USA, www.graphpad.com.
4.4. Determination of Pectin Methyl Esterase Activity
The assay was performed based on Huang et al., [13]. Seedlings samples (200 mg) were ground and resuspended in phosphate–citrate buffer (0.1 M citric acid; Na2HPO4 0.2 M; NaCl 1 M; pH 5) at 4 °C, with a 3:1 ratio. Then, the samples were centrifuged at 12,000 rpm at 4 °C, the supernatants were recovered, and the total proteins were quantified with the Bradford method.
For the PME activity analysis, 5 µg of protein was added to 1 mL of 0.1% esterified pectin (SigmaAldrich) in 50 mM phosphate buffer (Na2HPO4 [0.2 M]; NaH2PO4 [0.2 M]; pH 7) and incubated at 37 °C for 1 h. At the end of the incubation, 200 µL of 0.02% ruthenium red was added and incubated at room temperature for 15 min. Finally, 200 µL of 0.6 M CaCl2 was added and centrifuged at 12,000 rpm for 10 min. Supernatant absorbance at 534 nm was recorded. Four independent biological replicates for each sample, with three technical replicates, were performed. The activity of commercially purifying PME (SigmaAldrich) was performed using the protocol previously described. The standard curve was used to calculate the specific activity for each replicate (U/mg protein). Student’s t-tests were performed using GraphPad Prism version 8.0.0 for windows, Graph Pad software, San Diego, CA, USA, www.graphpad.com.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11223049/s1, Figure S1: At3g08030 and DGR2 (At5g25460) gene expression in WT Col. during thermopriming and thermo-triggering treatments. Table S1: List of primers used for qRT-PCR experiment. Table S2: Alignment of BIIDXI primers to DUF642 gene sequences of Arabidopsis thaliana.
Author Contributions
Conceptualization, A.G.-d. and E.P.-H.; methodology, E.P.-H., J.E.C.-V. and X.G.-M.; analysis, E.P.-H., J.E.C.-V., A.G.-d. and E.M.-B.; writing—original draft preparation, E.P.-H. and A.G.-d.; writing—review and editing, J.E.C.-V., X.G.-M. and E.M.-B.; visualization, E.P.-H. and A.G.-d.; supervision, A.G.-d.; project administration, A.G.-d.; funding acquisition, A.G.-d. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT UNAM) [grant IN210621] to A.G.-d., and E.P.-H. was supported by the scholarship provided by the Consejo Nacional de Ciencia y Tecnología (CONACyT) [grant 800940].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Marco Tulio Solano de la Cruz for his technical assistance at the “Unidad de Genética Molecular” Instituto de Ecología, UNAM. We thank Teresa Caudillo and Q. Blanca Hernández Bautista for their technical assistance. E.P.-H. thanks Programa de Posgrado en Ciencias Biológicas (Universidad Nacional Autónoma de México).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wolf, S. Cell Wall Signaling in Plant Development and Defense. Annu. Rev. Plant Biol. 2022, 73, 323–353. [Google Scholar] [CrossRef] [PubMed]
- Balazadeh, S. A ‘hot’ cocktail: The multiple layers of thermomemory in plants. Curr. Opin. Plant Biol. 2022, 65, 102147. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-C.; Bulgakov, V.P.; Jinn, T.-L. Pectin methylesterases: Cell wall remodeling proteins are required for plant response to heat stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [PubMed]
- Gigli-Bisceglia, N.; Van Zelm, E.; Huo, W.; Lamers, J.; Testerink, C. Arabidopsis root responses to salinity depend on pectin modification and cell wall sensing. Development 2022, 149, dev200363. [Google Scholar] [CrossRef] [PubMed]
- Fabri, J.H.T.M.; Rocha, M.C.; Fernandes, C.M.; Persinoti, G.F.; Ries, L.N.A.; Cunha, A.F.D.; Goldman, G.H.; Del Poeta, M.; Malavazi, I. The heat shock transcription factor HsfA is essential for thermotolerance and regulates cell wall integrity in Aspergillus fumigatus. Front. Microbiol. 2021, 12, 656548. [Google Scholar] [CrossRef]
- Fragkostefanakis, S.; Mesihovic, A.; Simm, S.; Paupière, M.J.; Hu, Y.; Paul, P.; Mishra, S.K.; Tschiersch, B.; Theres, K.; Bovy, A.; et al. HsfA2 controls the activity of developmentally and stress-regulated heat stress protection mechanisms in tomato male reproductive tissues. Plant Physiol. 2016, 170, 2461–2477. [Google Scholar] [CrossRef]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K.L. Hitting the wall—Sensing and signaling pathways involved in plant cell wall remodeling in response to abiotic stress. Plants 2018, 7, 89. [Google Scholar] [CrossRef]
- Wolf, S.; Mouille, G.; Pelloux, J. Homogalacturonan methyl-esterification and plant development. Mol. Plant 2009, 2, 851–860. [Google Scholar] [CrossRef]
- Willats, W.G.T.; Orfila, C.; Limberg, G.; Buchholti, H.C.; van Alebeek, G.J.W.M.; Voragen, A.G.J.; Marcus, S.E.; Mikkelsen, J.D.; Murray, B.S.; Knox, J.P. Modulation of the degree and pattern of methyl-esterification of pectic homogalacturonan in plant cell walls: Implications for pectin methyl esterase action, matrix properties, and cell adhesion. J. Biol. Chem. 2001, 276, 19404–19413. [Google Scholar] [CrossRef]
- Wormit, A.; Usadel, B. The multifaceted role of pectin methylesterase inhibitors (PMEIs). Int. J. Mol. Sci. 2018, 19, 2878. [Google Scholar] [CrossRef]
- Cruz-Valderrama, J.E.; Gómez-Maqueo, X.; Salazar-Iribe, A.; Zúñiga-Sánchez, E.; Hernández-Barrera, A.; Quezada-Rodríguez, E.; Gamboa-deBuen, A. Overview of the role of cell wall DUF642 proteins in plant development. Int. J. Mol. Sci. 2019, 20, 3333. [Google Scholar] [CrossRef] [PubMed]
- Pelloux, J.; Rusterucci, C.; Mellerowicz, E.J. New insights into pectin methylesterase structure and function. Trends Plant Sci. 2007, 12, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-C.; Wu, H.C.; Wang, Y.D.; Liu, C.H.; Lin, C.C.; Luo, D.L.; Jinn, T.L. PECTIN METHYLESTERASE34 contributes to heat tolerance through its role in promoting stomatal movement. Plant Physiol. 2017, 174, 748–763. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Yu, S.Y.; Wang, Y.D.; Jinn, T.L. Guard Cell-Specific Pectin METHYLESTERASE53 Is Required for Abscisic Acid-Mediated Stomatal Function and Heat Response in Arabidopsis. Front. Plant Sci. 2022, 13, 346. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Hsu, S.F.; Luo, D.L.; Chen, S.J.; Huang, W.D.; Lur, H.S.; Jinn, T.L. Recovery of heat shock-triggered released apoplastic Ca2+ accompanied by pectin methylesterase activity is required for thermotolerance in soybean seedlings. J. Exp. Bot. 2010, 61, 2843–2852. [Google Scholar] [CrossRef]
- Bosch, M.; Hepler, P.K. Pectin methylesterases and pectin dynamics in pollen tubes. Plant Cell 2005, 17, 3219–3226. [Google Scholar] [CrossRef]
- Balestrieri, C.; Castaldo, D.; Giovane, A.; Quagliuolo, L.; Servillo, L. A glycoprotein inhibitor of pectin methylesterase in kiwi fruit (Actinidia chinensis). Eur. J. Biochem. 1990, 193, 183–187. [Google Scholar] [CrossRef]
- Coculo, D.; Lionetti, V. The Plant Invertase/Pectin Methylesterase Inhibitor Superfamily. Front. Plant Sci. 2022, 13, 863892. [Google Scholar] [CrossRef]
- Chen, J.; Chen, X.; Zhang, Q.; Zhang, Y.; Ou, X.; An, L.; Feng, H.; Zhao, Z. A cold-induced pectin methyl-esterase inhibitor gene contributes negatively to freezing tolerance but positively to salt tolerance in Arabidopsis. J. Plant Physiol. 2018, 222, 67–78. [Google Scholar] [CrossRef]
- Vázquez-Lobo, A.; Roujol, D.; Zuñiga-Sánchez, E.; Albenne, C.; Piñero, D.; de Buen, A.G.; Jamet, E. The highly conserved spermatophyte cell wall DUF642 protein family: Phylogeny and first evidence of interaction with cell wall polysaccharides in vitro. Mol. Phylogenetics Evol. 2012, 63, 510–520. [Google Scholar] [CrossRef]
- Jamet, E.; Canut, H.; Boudart, G.; Pont-Lezica, R.F. Cell wall proteins: A new insight through proteomics. Trends Plant Sci. 2006, 11, 33–39. [Google Scholar] [CrossRef]
- Zúñiga-Sánchez, E.; Gamboa-de Buen, A. The two DUF642 At5g11420 and At4g32460-encoded proteins interact in vitro with the AtPME3 catalytic domain. Protein Interact. 2012, 7, 119–142. [Google Scholar]
- Wang, M.; Zhu, X.; Peng, G.; Liu, M.; Zhang, S.; Chen, M.; Liao, S.; Wei, X.; Xu, P.; Tan, X.; et al. Methylesterification of cell-wall pectin controls the diurnal flower-opening times in rice. Mol. Plant 2022, 15, 956–972. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga-Sánchez, E.; Soriano, D.; Martínez-Barajas, E.; Orozco-Segovia, A.; Gamboa-deBuen, A. BIIDXI, the At4g32460 DUF642 gene, is involved in pectin methyl esterase regulation during Arabidopsis thaliana seed germination and plant development. BMC Plant Biol. 2014, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Valderrama, J.E.; Jiménez-Durán, K.; Zúñiga-Sánchez, E.; Salazar-Iribe, A.; Márquez-Guzmán, J.; Gamboa-deBuen, A. Degree of pectin methyl esterification in endosperm cell walls is involved in embryo bending in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2018, 495, 639–645. [Google Scholar] [CrossRef]
- Salazar-Iribe, A.; Agredano-Moreno, L.T.; Zúñiga-Sánchez, E.; Jiménez-Garcia, L.F.; Gamboa-deBuen, A. The cell wall DUF642 At2g41800 (TEB) protein is involved in hypocotyl cell elongation. Plant Sci. 2016, 253, 206–214. [Google Scholar] [CrossRef]
- Shin, Y.; Chane, A.; Jung, M.; Lee, Y. Recent advances in understanding the roles of pectin as an active participant in plant signaling networks. Plants 2021, 10, 1712. [Google Scholar] [CrossRef]
- Pinski, A.; Betekhtin, A.; Skupien-Rabian, B.; Jankowska, U.; Jamet, E.; Hasterok, R. Changes in the cell wall proteome of leaves in response to high temperature stress in Brachypodium distachyon. Int. J. Mol. Sci. 2021, 22, 6750. [Google Scholar] [CrossRef]
- Olas, J.J.; Apelt, F.; Annunziata, M.G.; John, S.; Richard, S.I.; Gupta, S.; Kragler, F.; Balazadeh, S.; Mueller-Roeber, B. Primary carbohydrate metabolism genes participate in heat-stress memory at the shoot apical meristem of Arabidopsis thaliana. Mol. Plant 2021, 14, 1508–1524. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Wang, S.; Cui, Y.; Yan, D. Heat Stress Reduces Root Meristem Size via Induction of Plasmodesmal Callose Accumulation Inhibiting Phloem Unloading in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 2063. [Google Scholar] [CrossRef]
- Bustamante, C.A.; Budde, C.O.; Borsani, J.; Lombardo, V.A.; Lauxmann, M.A.; Andreo, C.S.; Lara, M.V.; Drincovich, M.F. Heat treatment of peach fruit: Modifications in the extracellular compartment and identification of novel extracellular proteins. Plant Physiol. Biochem. 2012, 60, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Ngcala, M.G.; Goche, T.; Brown, A.P.; Chivasa, S.; Ngara, R. Heat stress triggers differential protein accumulation in the extracellular matrix of sorghum cell suspension cultures. Proteomes 2020, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J. 2016, 35, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.; Fan, C.; Yang, Q.; Li, X.; Wan, B.; Dong, Y.; Wang, X.; Zhou, Y. Identification of heat responsive genes in Brassica napus siliques at the seed-filling stage through transcriptional profiling. PLoS ONE 2014, 9, e101914. [Google Scholar] [CrossRef]
- Salazar-Iribe, A.; Cruz-Valderrama, J.E.; Jímenez-Durán, K.; Gómez-Maqueo, X.; Gamboa-deBuen, A. BIIDXI, a DUF642 cell wall protein, is involved in hypocotyl growth via auxin efflux. J. Plant. Physiol. 2018, 231, 105–109. [Google Scholar] [CrossRef]
- Willems, E.; Leyns, L.; Vandesompele, J. Standardization of real-time PCR gene expression data from independent biological replicates. Anal. Biochem. 2008, 379, 127–129. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).