
Chrysosphaerella septentrionalis sp. nov. (Chrysophyceae, Chromulinales), a New Species from the Arctic Including the Description of Chrysosphaerellaceae, fam. nov.



Abstract
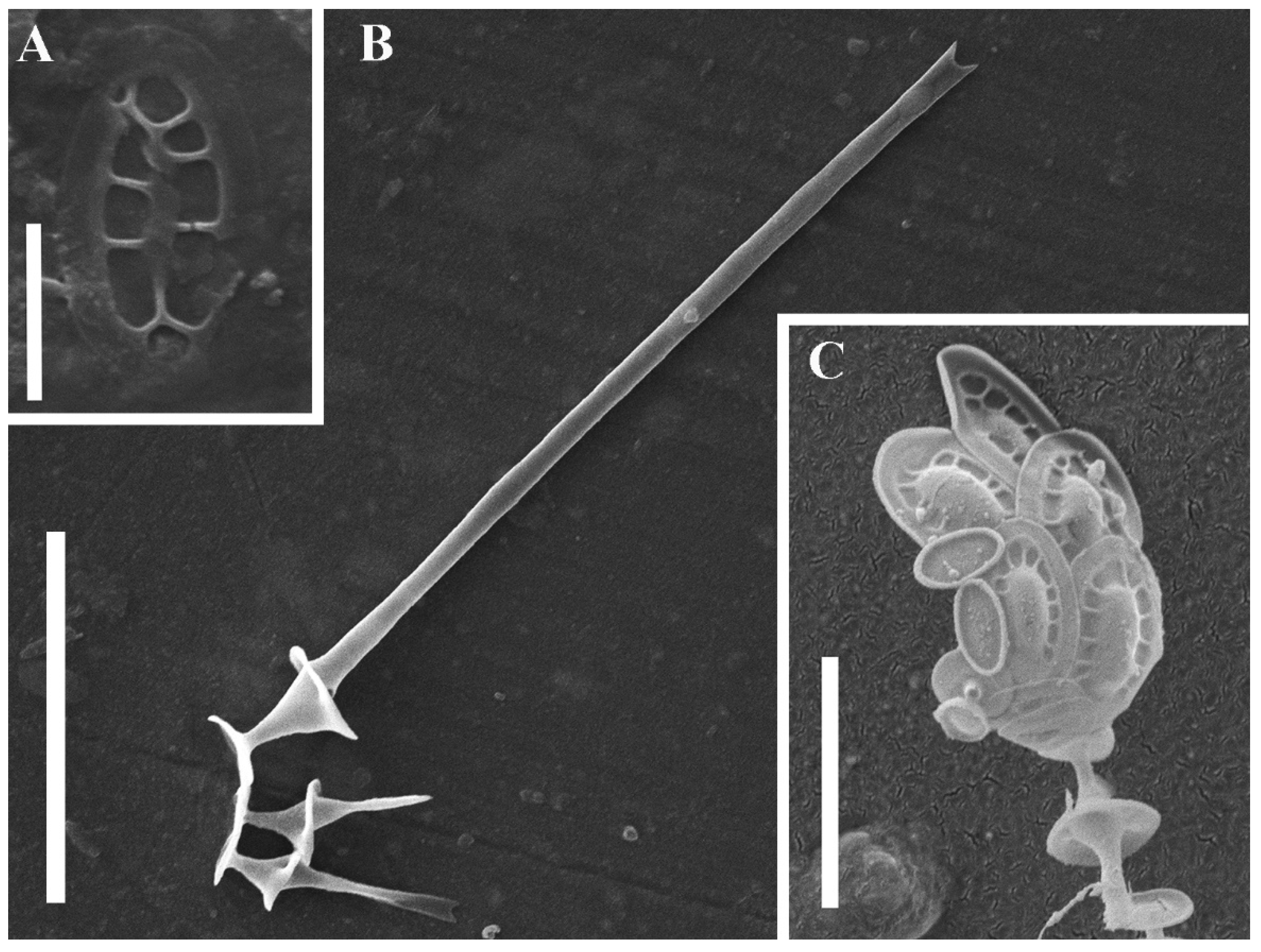
1. Introduction
2. Results
3. Discussion
3.1. Species Diversity of Chrysosphaerella
3.2. Infrageneric Classification of the Genus Chrysosphaerella
- Chrysosphaerella sect. Chrysosphaerella
- 2.
- Chrysosphaerella sect. Brevispinae Kapustin, sect. nov.
- 3.
- Chrysosphaerella sect. Septispinae Kapustin, sect. nov.
3.3. Taxonomic Placement of the Genus Chrysosphaerella
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lauterborn, R. Diagnosen neuer Protozoen aus dem Gebiete des Oberrheins. Zool. Anz. 1896, 19, 14–18. [Google Scholar]
- Lauterborn, R. Protozoen-Studien. IV. Flagellaten aus dem Gebiete des Oberrheins. Z. Wiss. Zool. 1899, 65, 369–391. [Google Scholar]
- Korshikov, A.A. On some new or little known flagellates. Arch. Protistenkd. 1941, 95, 22–44. [Google Scholar]
- Fott, B.; Ludvík, J. Elektronenoptische Untersuchung der Kieselstrukturen bei Chrysosphaerella (Chrysomonadineae). Preslia 1956, 28, 276–278. [Google Scholar]
- Harris, K.; Bradley, D.E. Some Unusual Chrysophyceae Studied in the Electron Microscope. J. Gen. Microbiol. 1958, 18, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Skuja, H. Taxonomie des Phytoplanktons einiger Seen in Uppland, Schweden. Symb. Bot. Upsal. 1948, 9, 1–399. [Google Scholar]
- Bradley, D.E. A study of the Mallomonas, Synura and Chrysosphaerella of Northern Iceland. J. Gen. Microbiol. 1964, 37, 321–333. [Google Scholar] [CrossRef]
- Kristiansen, J. Chrysosphaerella multispina Bradley and some other remarkable Chrysophyceae from Lake Stråken, Aneboda, Sweden. Österr. Bot. Z. 1969, 116, 70–84. [Google Scholar] [CrossRef]
- Wujek, D.E.; Hamilton, R. Studies on Michigan Chrysophyceae. I. Mich. Bot. 1972, 11, 51–59. [Google Scholar]
- Asmund, B. Survey of the genus Chrysosphaerella (Chrysophyceae) as studied in the electron microscope with description of a new species. Bot. Tidsskr. 1973, 68, 132–139. [Google Scholar]
- Kristiansen, J. Chrysophyceae from Alberta and British Columbia. Syesis 1975, 8, 97–108. [Google Scholar]
- Nicholls, K.H. A Reassessment of Chrysosphaerella lonqispina and C. multispina, and a Revised Key to Related Genera in the Synuraceae (Chrysophyceae). Plant Syst. Evol. 1980, 135, 95–106. [Google Scholar] [CrossRef]
- Wujek, D.E.; Gretz, M.; Wujek, M.G. Studies on Michigan Chrysophyceae. IV. Mich. Bot. 1977, 16, 191–195. [Google Scholar]
- Preisig, H.R.; Takahashi, E. Chrysosphaerella (Pseudochrysosphaerella) solitaria, spec. nova (Chrysophyceae). Plant Syst. Evol. 1978, 129, 135–142. [Google Scholar] [CrossRef]
- Beech, P.L.; Moestrup, Ø. Light and electron microscopical observations on the heterotrophic protist Thaumatomastix salina comb. nov. (syn. Chrysosphaerella salina) and its allies. Nord. J. Bot. 1986, 6, 865–877. [Google Scholar] [CrossRef]
- Takahashi, E. Studies on genera Mallomonas and Synura, and other plankton in freshwater with the electron microscope. VII. New genus Spiniferomonas of the Synuraceae (Chrysophyceae). Bot. Mag. 1973, 86, 75–88. [Google Scholar] [CrossRef]
- Nicholls, K.H. Spiniferomonas septispina and S. enigmata, two new algal species confusing the distinction between Spiniferomonas and Chrysosphaerella (Chrysophyceae). Plant Syst. Evol. 1984, 148, 103–117. [Google Scholar] [CrossRef]
- Kristiansen, J.; Tong, D. Chrysosphaerella annulala n. sp., a new scale-bearing chrysophyte. Nord. J. Bot. 1989, 9, 329–332. [Google Scholar] [CrossRef]
- Preisig, H.R.; Hibberd, D.J. Ultrastructure and taxonomy of Paraphysomonas (Chrysophyceae) and related genera 3. Nord. J. Bot. 1983, 3, 695–723. [Google Scholar] [CrossRef]
- Škaloud, P.; Kristiansen, J.; Škaloudová, M. Developments in the taxonomy of silica-scaled chrysophytes—From morphological and ultrastructural to molecular approaches. Nord. J. Bot. 2013, 31, 385–402. [Google Scholar] [CrossRef]
- Scoble, J.M.; Cavalier-Smith, T. Scale evolution in Paraphysomonadida (Chrysophyceae): Sequence phylogeny and revised taxonomy of Paraphysomonas, new genus Clathromonas, and 25 new species. Eur. J. Protistol. 2014, 50, 551–592. [Google Scholar] [CrossRef] [PubMed]
- Kapustin, D.A.; Gusev, E.S.; Lilitskaya, G.G.; Kulikovskiy, M.S. Silica-scaled chrysophytes from the Ukrainian Polissia. Cryptogam. Algol. 2020, 41, 121–135. [Google Scholar] [CrossRef]
- Dürrschmidt, M.; Croome, R. Mallomonadaceae (Chrysophyceae) from Malaysia and Australia. Nord. J. Bot. 1985, 5, 285–298. [Google Scholar] [CrossRef]
- Bessudova, A.Y.; Domysheva, V.M.; Firsova, A.D.; Likhoshway, Y.V. Silica-scaled chrysophytes of Lake Baikal. Acta Biol. Sib. 2017, 3, 47–56. [Google Scholar] [CrossRef][Green Version]
- Balonov, I.M. Golden algae of the family Synuraceae from the waterbodies of Karelia. In Flora and Vegetation of the Waterbodies of the Upper Volga Basin; Rybinsk, Russia, 1979; pp. 3–26. [Google Scholar]
- Kapustin, D.A.; Gusev, E.S. Silica-scaled chrysophytes from West Java (Indonesia) including description of a new Chrysosphaerella species. Nova Hedwig. Beih. 2019, 148, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Balonov, I.M. Electron microscopic study of the genus Spiniferomonas Takahashi (Chrysophyta). Bot. Zhurnal 1978, 63, 1639–1647. (In Russian) [Google Scholar]
- Škaloudová, M.; Škaloud, P. A new species of Chrysosphaerella (Chrysophyceae: Chromulinales), Chrysosphaerella rotundata sp. nov., from Finland. Phytotaxa 2013, 130, 34–42. [Google Scholar] [CrossRef]
- Cronberg, G.; Kristiansen, J. Synuraceae and other Chrysophyceae from central Småland, Sweden. Bot. Not. 1980, 133, 595–618. [Google Scholar]
- Nicholls, K.H. New and little-known marine and freshwater species of the silica-scaled genera Thaumatomastix and Reckertia (Cercozoa: Thaumatomonadida). J. Mar. Biol. Assoc. U.K. 2013, 93, 1231–1244. [Google Scholar] [CrossRef]
- Siver, P.A. Morphological and ecological characteristics of Chrysosphaerella longispina and C. brevispina (Chrysophyceae). Nord. J. Bot. 1993, 13, 343–351. [Google Scholar] [CrossRef]
- Siver, P.A.; Voloshko, L.N.; Gavrilova, O.V.; Getsen, M.V. The scaled chrysophyte flora of the Bolshezemelskaya tundra (Russia). Nova Hedwig. Beih. 2005, 128, 125–150. [Google Scholar]
- Bessudova, A.Y.; Bukin, Y.S.; Sorokovikova, L.M.; Firsova, A.D.; Tomberg, I.V. Silica-scaled chrysophytes in small lakes of the lower Yenisei basin, the Arctic. Nova Hedwig. 2018, 107, 315–336. [Google Scholar] [CrossRef]
- Němcová, Y.; Pusztai, M.; Škaloudová, M.; Neustupa, J. Silica-scaled chrysophytes (Stramenopiles, Ochrophyta) along a salinity gradient: A case study from the Gulf of Bothnia western shore (northern Europe). Hydrobiologia 2016, 764, 187–197. [Google Scholar] [CrossRef]
- Lemmermann, E. Das Phytoplankton sächsischer Teiche. Forschungsber. Biol. Stat. Plön 1899, 7, 96–135. [Google Scholar]
- Pascher, A. Chrysomonadinae. In Die Süsswasser-Flora Deutschlands, Österreichs und der Schweiz; Heft 2, S. 7–95; Pascher, A., Ed.; G. Fischer Verlag: Jena, Germany, 1913. [Google Scholar]
- Pascher, A. Der Grossteich bei Hirschberg in Nord-Böhmen. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1910, 1, 1–66. [Google Scholar]
- Pascher, A. Über Rhizopoden- und Palmellastadien bei Flagellaten (Chrysomonaden), nebst einer Übersicht über die braunen Flagellaten. Arch. Protistenkd. 1912, 25, 153–200. [Google Scholar]
- Bourrelly, P. Recherches sur les Chrysophycées. Morphologie, Phylogénie, Systématique. Rev. Algol. Mém. Hors Sér. 1957, 1, 1–412. [Google Scholar]
- Starmach, K. Chrysophyceae und Haptophyceae. In Süßwasserflora von Mitteleuropa; Band 1, S. 1–515; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Stuttgart, Gerrmany, 1985. [Google Scholar]
- Cavalier-Smith, T.; Chao, E.E.; Thompson, C.E.; Hourihane, S.L. Oikomonas, a distinctive zooflagellate related to chrysomonads. Arch. Protistenkd. 1996, 146, 273–279. [Google Scholar] [CrossRef]
- Andersen, R.A.; Van de Peer, Y.; Potter, D.; Sexton, J.P.; Kawachi, M.; LaJeunesse, T. Phylogenetic analysis of the SSU rRNA from members of the Chrysophyceae. Protist 1999, 150, 71–84. [Google Scholar] [CrossRef]
- Kristiansen, J.; Škaloud, P. Chrysophyta. In Handbook of the Protists, 2nd ed.; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 331–366. [Google Scholar] [CrossRef]
- Andersen, R.A. Molecular systematics of the Chrysophyceae and Synurophyceae. In Unravelling the Algae: The Past, Present, and Future of Algal Systematics; Brodie, J., Lewis, J., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 285–313. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Guiry, M.D. Reinstatement of Lepidochromonas Kristiansen (Lepidochromonadaceae fam. nov., Chrysophyceae). Phytotaxa 2019, 413, 49–53. [Google Scholar] [CrossRef]
- Pascher, A. Über Flagellaten und Algen. Ber. Dtsch. Bot. Ges. 1914, 32, 136–160. [Google Scholar]



{kind=link}
{kind=link}
| Taxon | Taxonomic Status | Reference(s) |
|---|---|---|
| Chrysosphaerella annulata Kristiansen & D. Tong | Accepted | [18] |
| Chrysosphaerella astrea Dürrschmidt | Accepted | [23] |
| Chrysosphaerella baikalensis Popovskaya | Accepted (Probably, it is conspecific with C. brevispina) | [24] |
| Chrysosphaerella brevispina Korshikov | Accepted | [5] |
| Chrysosphaerella conradii Bourrelly | Synonym of C. brevispina | [10] |
| Chrysosphaerella coronacircumspina Wujek & Kristiansen var. coronacircumspina | Accepted | [13] |
| Chrysosphaerella coronacircumspina var. grandibasa Balonov | Accepted | [25] |
| Chrysosphaerella enigmata (K.H. Nicholls) Kristiansen & D. Tong | Accepted | [18] |
| Chrysosphaerella longispina Lauterborn | Accepted | [12] |
| Chrysosphaerella multispina Bradley | Invalid Synonym of C. longispina | [12] |
| Chrysosphaerella nichollsii D. Kapustin & E.S. Gusev | Accepted | [26] |
| Chrysosphaerella parva Asmund | Synonym of two Spiniferomonas taxa, S. abei E. Takahashi and S. bilacunosa E. Takahashi | [27] |
| Chrysosphaerella patelliformis E. Takahashi & Hara | Synonym of Thaumatomastix patelliformis (E. Takahashi & Hara) Beech & Moestrup | [15] |
| Chrysosphaerella rodhei Skuja | Synonym of C. brevispina | [10] |
| Chrysosphaerella rotundata Škaloudová & Škaloud | Accepted | [28] |
| Chrysosphaerella salina Birch-Andersen | Synonym of Thaumatomastix salina (Birch-Andersen) P.L. Beech & Moestrup | [15] |
| Chrysosphaerella septispina (K.H. Nicholls) Kristiansen & D. Tong | Accepted | [18] |
| Chrysosphaerella setifera Schiller | Insufficiently described | [10] |
| Chrysosphaerella solitaria Preisig & E. Takahashi | Synonym of C. coronacircumspina | [29] |
| Chrysosphaerella triangulata Balonov | Synonym of Thaumatomastix triangulata (Balonov) P.L. Beech & Moestrup emend. K.H. Nicholls | [15,30] |
| Chrysosphaerella tripus E. Takahashi & Hara | Synonym of Thaumatomastix tripus (E. Takahashi & Hara) P.L. Beech & Moestrup | [15] |
| Species | Plate-like Scales (Length × Width, µm) | Spine-like Scales (Length, µm) | Reference |
|---|---|---|---|
| Chrysosphaerella septentrionalis | 2.4–3.0 × 1.8–3.0 unpatterned scales: 2.1 × 1.1 | 4.3–8.3 13.0–31.3 | This study |
| Chrysosphaerella longispina (=C. multispina) | 0.6–6.0 | 5–10 20–25 35–40 | [7] |
| 2.0–2.5 × 1.25–1.6 4.6–6.0 × 2.1–3.3 unpatterned scales: 1.3–1.7 × 0.9–1.0 | 3–4 up to 50 | [8] | |
| 3.5–6.0 × 2.2–3.0 unpatterned scales: 1.8–2.5 × 1.0–1.6 | 3–85 | [12] | |
| 4.2–2.1 (mean size) | 3.7–6.8 13–53 up to 71 | [31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapustin, D.; Kulikovskiy, M. Chrysosphaerella septentrionalis sp. nov. (Chrysophyceae, Chromulinales), a New Species from the Arctic Including the Description of Chrysosphaerellaceae, fam. nov. Plants 2022, 11, 3166. https://doi.org/10.3390/plants11223166
Kapustin D, Kulikovskiy M. Chrysosphaerella septentrionalis sp. nov. (Chrysophyceae, Chromulinales), a New Species from the Arctic Including the Description of Chrysosphaerellaceae, fam. nov. Plants. 2022; 11(22):3166. https://doi.org/10.3390/plants11223166
Chicago/Turabian StyleKapustin, Dmitry, and Maxim Kulikovskiy. 2022. "Chrysosphaerella septentrionalis sp. nov. (Chrysophyceae, Chromulinales), a New Species from the Arctic Including the Description of Chrysosphaerellaceae, fam. nov." Plants 11, no. 22: 3166. https://doi.org/10.3390/plants11223166
APA StyleKapustin, D., & Kulikovskiy, M. (2022). Chrysosphaerella septentrionalis sp. nov. (Chrysophyceae, Chromulinales), a New Species from the Arctic Including the Description of Chrysosphaerellaceae, fam. nov. Plants, 11(22), 3166. https://doi.org/10.3390/plants11223166