

Amylase Trypsin Inhibitors (ATIs) in a Selection of Ancient and Modern Wheat: Effect of Genotype and Growing Environment on Inhibitory Activities





Abstract
:1. Introduction
2. Results and Discussion
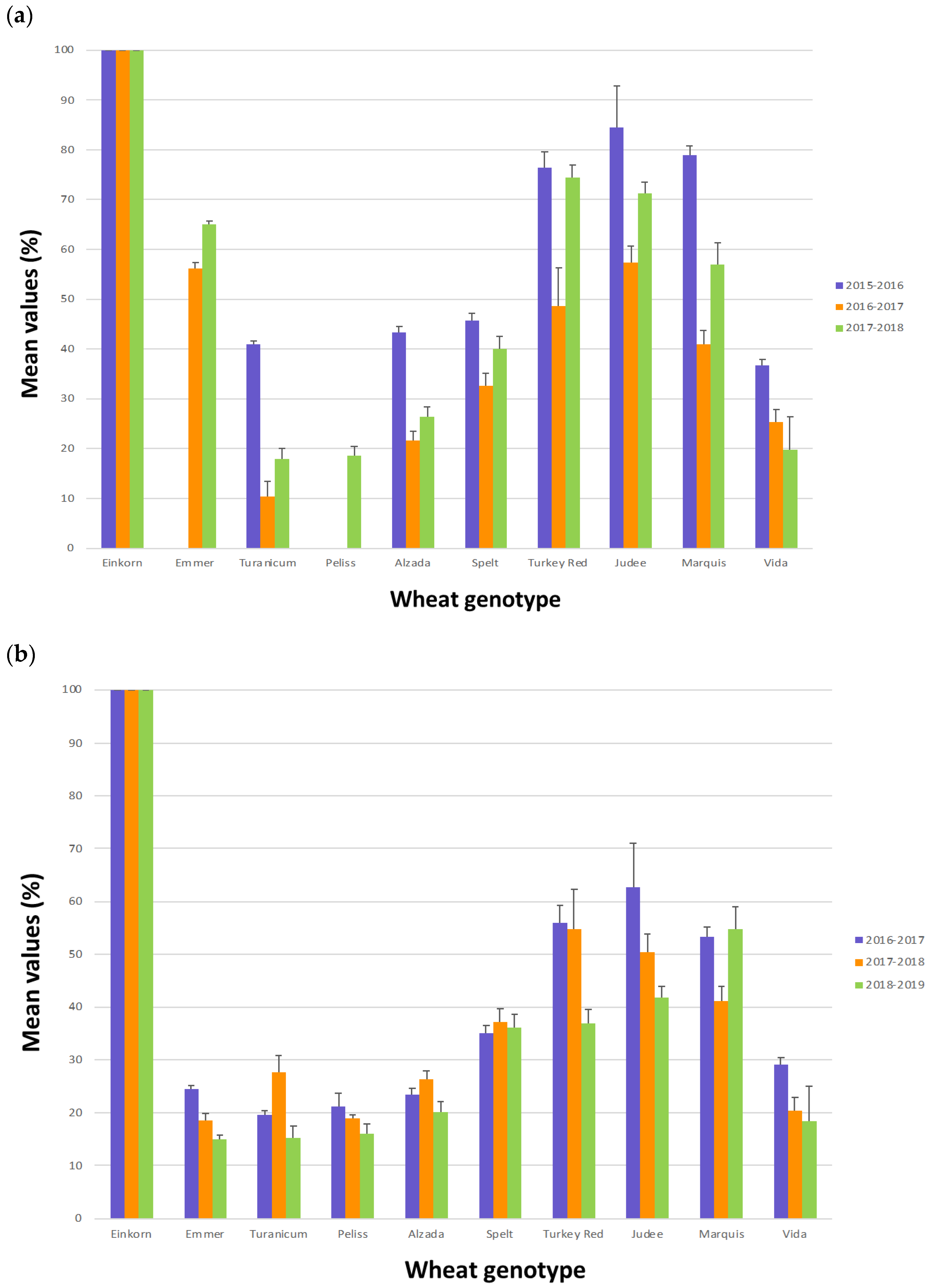
2.1. Characterization of Grain Samples
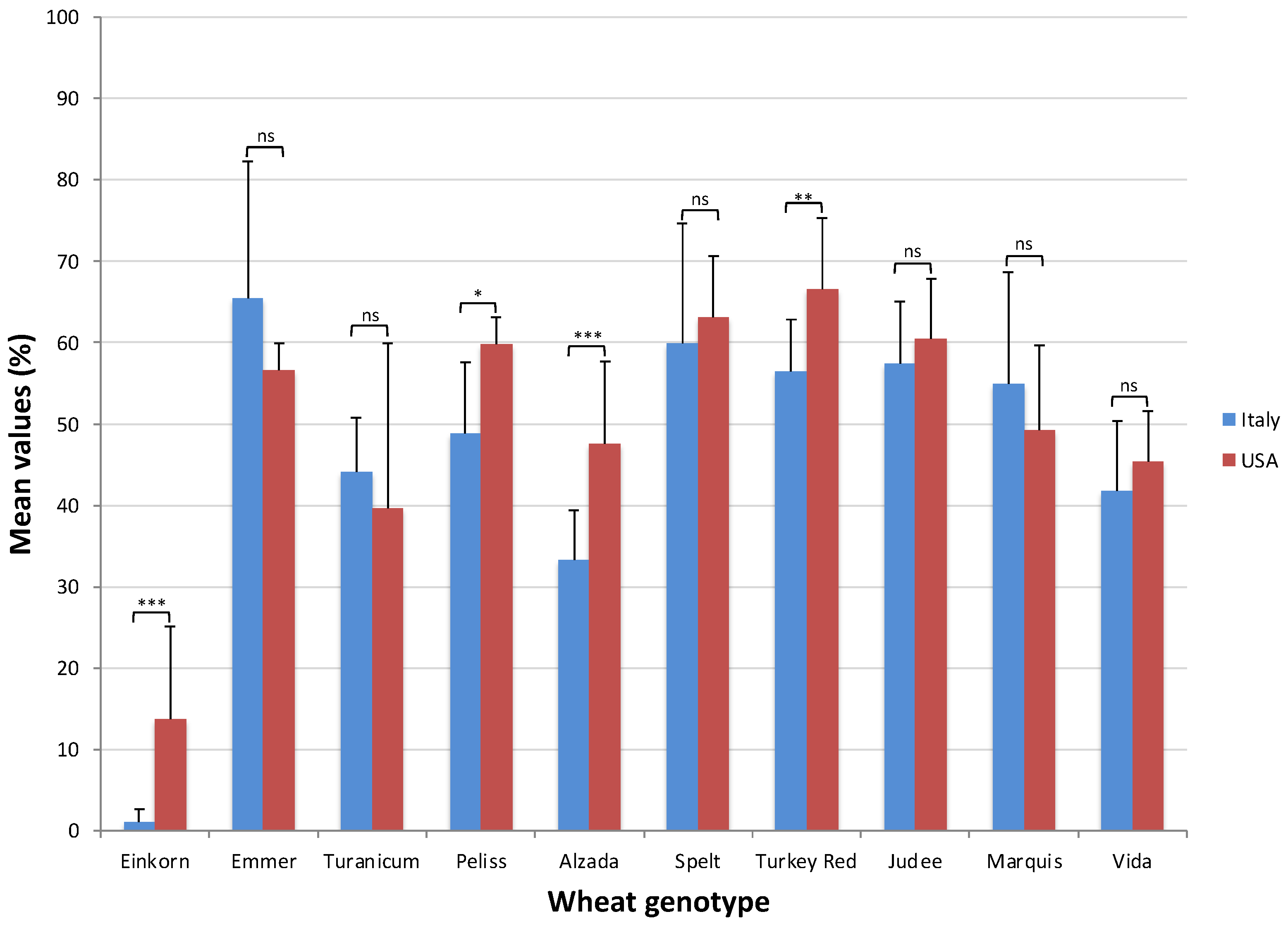
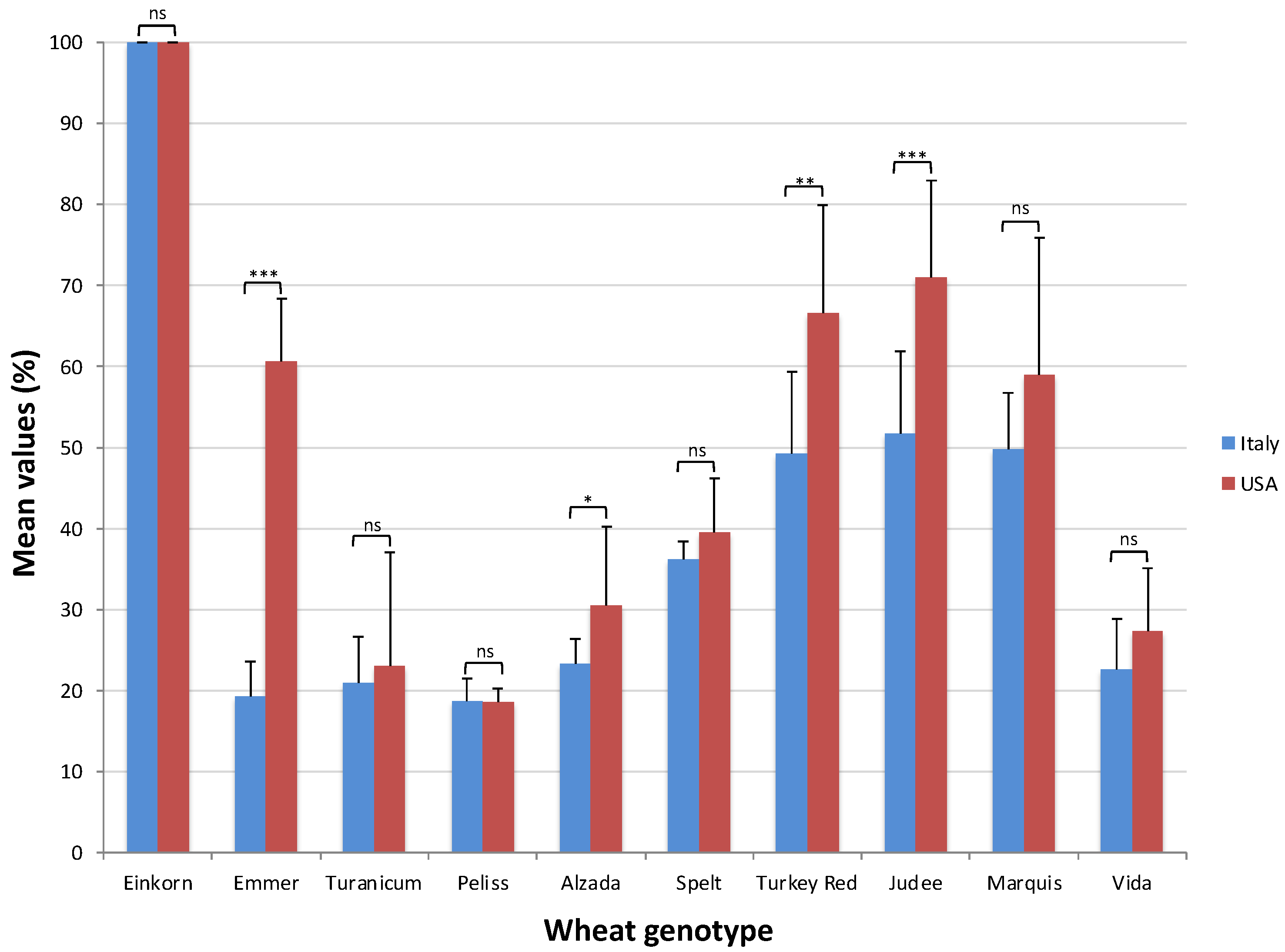
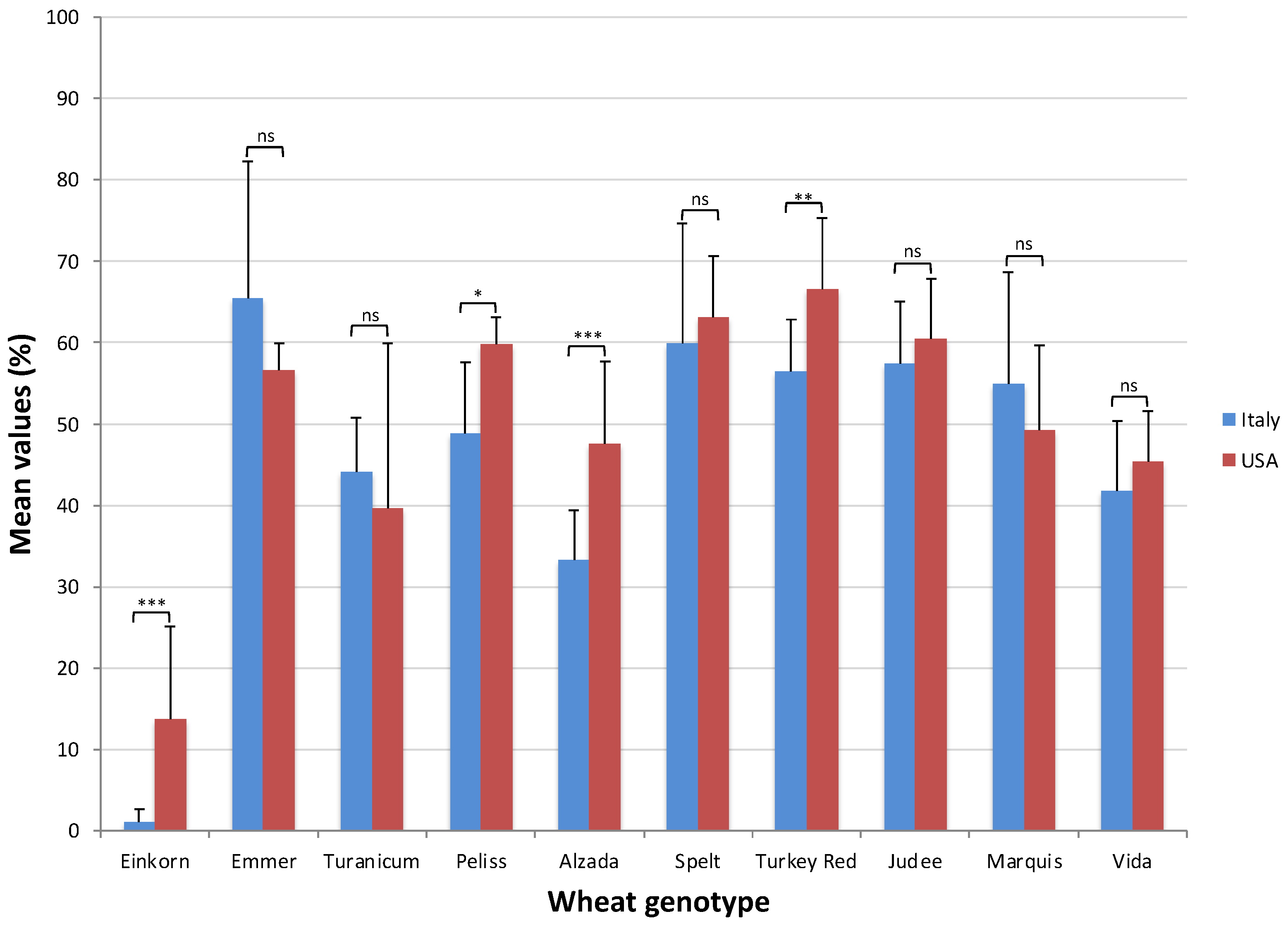
2.2. Alpha-Amylase Inhibitory Activity Assay
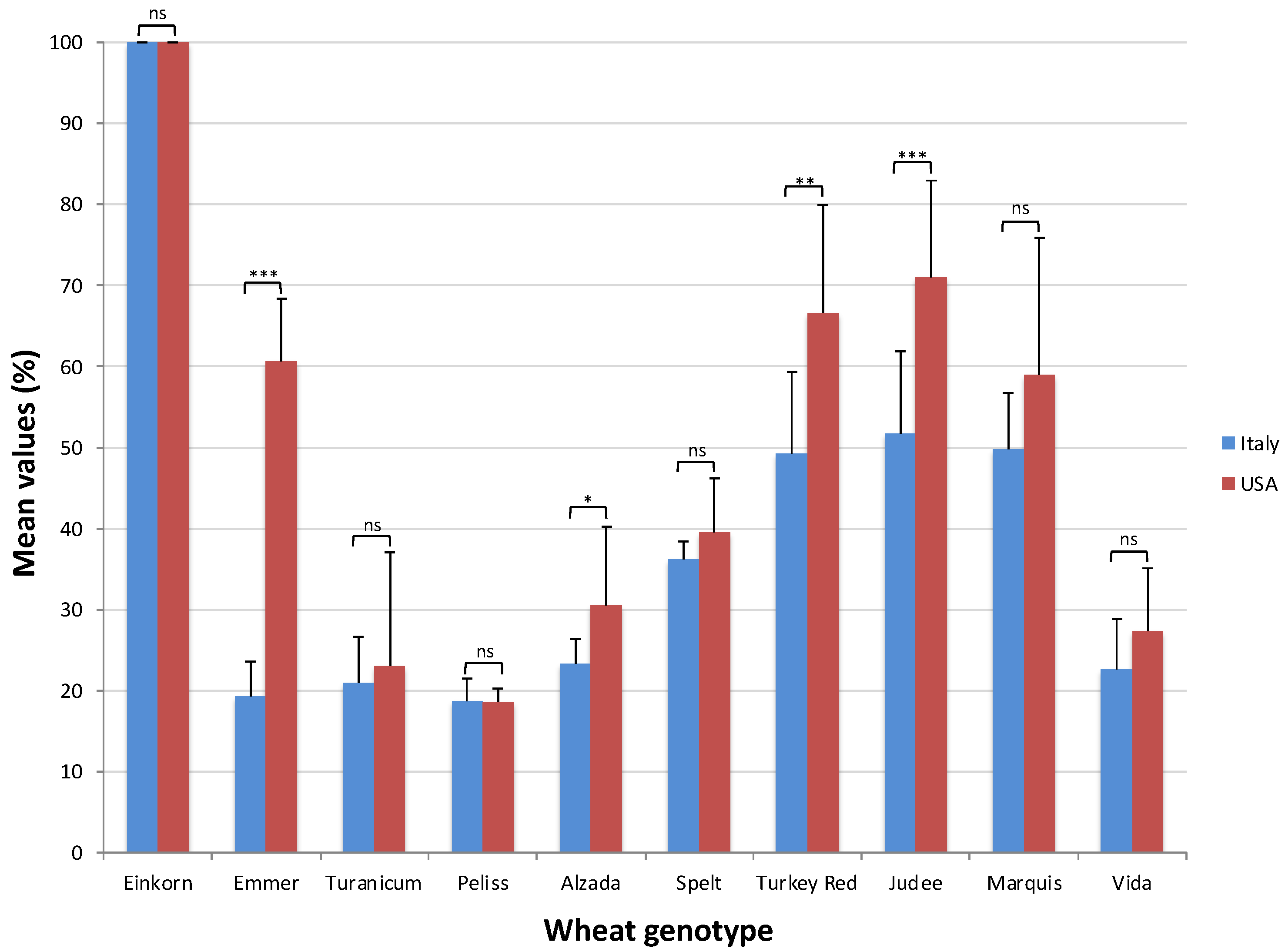
2.3. Trypsin Inhibitory Activity Assay
2.4. ATI Inhibitory Activity, Genotype and Growing Environment
2.5. ATI-Inhibitory Activity and Human Nutrition
3. Materials and Methods
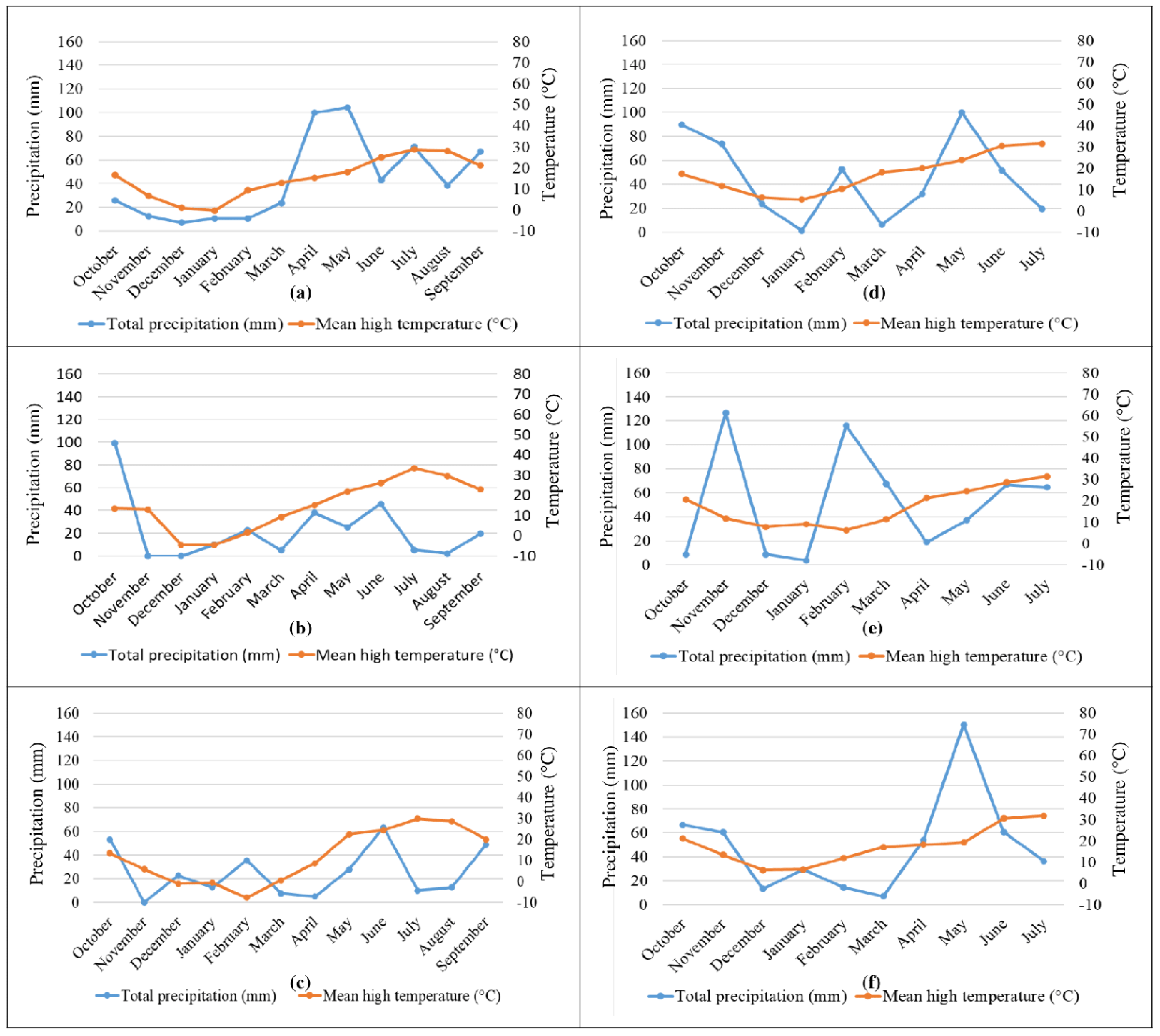
3.1. Grain Samples
3.2. Assessment of Mycotoxin Levels
3.3. Enzymatic Assays
3.3.1. Alpha-Amylase Inhibitory Activity Assay
3.3.2. Trypsin Inhibitory Activity Assay
3.3.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dupont, F.M.; Vensel, W.H.; Tanaka, C.K.; Hurkman, W.J.; Altenbach, S.B. Deciphering the Complexities of the Wheat Flour Proteome Using Quantitative Two-Dimensional Electrophoresis, Three Proteases and Tandem Mass Spectrometry. Proteome Sci. 2011, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonero, P.; García-Olmedo, F. A Multigene Family of Trypsin/α-Amylase Inhibitors from Cereals. In Seed Proteins; Springer: Dordrecht, The Netherlands, 1999; pp. 617–633. [Google Scholar] [CrossRef] [Green Version]
- Salcedo, G.; Snchez-Monge, R.; Garca-Casado, G.; Armentia, A.; Gomez, L.; Barber, D. The Cereal α-Amylase/Trypsin Inhibitor Family Associated with Bakers’ Asthma and Food Allergy. In Plant Food Allergens; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2004; pp. 70–86. [Google Scholar] [CrossRef]
- Altenbach, S.; Vensel, W.H.; Dupont, F.M. The Spectrum of Low Molecular Weight Alpha-Amylase/Protease Inhibitor Genes Expressed in the US Bread Wheat Cultivar Butte 86. BMC Res. Notes 2011, 4, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatham, A.S.; Shewry, P.R. Allergens to Wheat and Related Cereals. Clin. Exp. Allergy 2008, 38, 1712–1726. [Google Scholar] [CrossRef] [PubMed]
- Junker, Y.; Zeissig, S.; Kim, S.J.; Barisani, D.; Wieser, H.; Leffler, D.A.; Zevallos, V.; Libermann, T.A.; Dillon, S.; Freitag, T.L.; et al. Wheat Amylase Trypsin Inhibitors Drive Intestinal Inflammation via Activation of Toll-like Receptor 4. J. Exp. Med. 2012, 209, 2395–2408. [Google Scholar] [CrossRef] [PubMed]
- Geisslitz, S.; Shewry, P.; Brouns, F.; America, A.H.P.; Caio, G.P.I.; Daly, M.; D’Amico, S.; De Giorgio, R.; Gilissen, L.; Grausgruber, H.; et al. Wheat ATIs: Characteristics and Role in Human Disease. Front. Nutr. 2021, 8, 667370. [Google Scholar] [CrossRef] [PubMed]
- Zevallos, V.F.; Raker, V.; Tenzer, S.; Jimenez-Calvente, C.; Ashfaq-Khan, M.; Rüssel, N.; Pickert, G.; Schild, H.; Steinbrink, K.; Schuppan, D. Nutritional Wheat Amylase-Trypsin Inhibitors Promote Intestinal Inflammation via Activation of Myeloid Cells. Gastroenterology 2017, 152, 1100–1113.e12. [Google Scholar] [CrossRef] [Green Version]
- Gélinas, P.; Gagnon, F. Inhibitory Activity towards Human α-Amylase in Wheat Flour and Gluten. Int. J. Food Sci. Technol. 2018, 53, 467–474. [Google Scholar] [CrossRef]
- Bosi, S.; Negri, L.; Fakaros, A.; Oliveti, G.; Whittaker, A.; Dinelli, G. GGE Biplot Analysis to Explore the Adaption Potential of Italian Common Wheat Genotypes. Sustainability 2022, 14, 897. [Google Scholar] [CrossRef]
- Dinu, M.; Whittaker, A.; Pagliai, G.; Benedettelli, S.; Sofi, F. Ancient Wheat Species and Human Health: Biochemical and Clinical Implications. J. Nutr. Biochem. 2018, 52, 1–9. [Google Scholar] [CrossRef]
- Call, L.; Kapeller, M.; Grausgruber, H.; Reiter, E.; Schoenlechner, R.; D’Amico, S. Effects of Species and Breeding on Wheat Protein Composition. J. Cereal Sci. 2020, 93, 102974. [Google Scholar] [CrossRef]
- Simonetti, E.; Bosi, S.; Negri, L.; Baffoni, L.; Masoni, A.; Marotti, I.; Benedettelli, S.; Dinelli, G. Molecular Phylogenetic Analysis of Amylase Trypsin Inhibitors (ATIs) from a Selection of Ancient and Modern Wheat. J. Cereal Sci. 2022, 105, 103441. [Google Scholar] [CrossRef]
- Belderok, B.; Mesdag, J.; Mesdag, H.; Donner, D.A. Bread-Making Quality of Wheat: A Century of Breeding in Europe; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2000; 416p. [Google Scholar]
- Piasecka-Kwiatkowska, D.; Madaj, D.; Warchalewski, J.R. The Biological Activity of Wheat, Rye and Triticale Varieties Harvested in Four Consecutive Years. Acta Sci. Pol. Technol. Aliment. 2007, 6, 55–65. [Google Scholar]
- Franco, O.L.; Rigden, D.J.; Melo, F.R.; Bloch, C.; Silva, C.P.; Grossi De Sá, M.F. Activity of Wheat α-Amylase Inhibitors towards Bruchid α-Amylases and Structural Explanation of Observed Specificities. Eur. J. Biochem. 2000, 267, 2166–2173. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Maeda, K.; Murayama, R.; DiMagno, E.P. Character of a Wheat Amylase Inhibitor Preparation and Effects on Fasting Human Pancreaticobiliary Secretions and Hormones. Gastroenterology 1996, 111, 1313–1320. [Google Scholar] [CrossRef]
- Bedetti, C. Amylase Protein Inhibitors and the Role of Aegilops Species in Polyploid Wheat Speciation. Biochim. Biophys. Acta 1974, 362, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Geisslitz, S.; Ludwig, C.; Scherf, K.A.; Koehler, P. Targeted LC-MS/MS Reveals Similar Contents of α-Amylase/Trypsin-Inhibitors as Putative Triggers of Nonceliac Gluten Sensitivity in All Wheat Species except Einkorn. J. Agric. Food Chem. 2018, 66, 12395–12403. [Google Scholar] [CrossRef]
- Geisslitz, S.; Longin, C.F.H.; Koehler, P.; Scherf, K.A. Comparative Quantitative LC–MS/MS Analysis of 13 Amylase/Trypsin Inhibitors in Ancient and Modern Triticum Species. Sci. Rep. 2020, 10, 14570. [Google Scholar] [CrossRef]
- Zoccatelli, G.; Sega, M.; Bolla, M.; Cecconi, D.; Vaccino, P.; Rizzi, C.; Chignola, R.; Brandolini, A. Expression of α-Amylase Inhibitors in Diploid Triticum Species. Food Chem. 2012, 135, 2643–2649. [Google Scholar] [CrossRef]
- García-Maroto, F.; Maraña, C.; Mena, M.; García-Olmedo, F.; Carbonero, P. Cloning of CDNA and Chromosomal Location of Genes Encoding the Three Types of Subunits of the Wheat Tetrameric Inhibitor of Insect α-Amylase. Plant Mol. Biol. 1990, 14, 845–853. [Google Scholar] [CrossRef]
- Payan, F. Structural Basis for the Inhibition of Mammalian and Insect α-Amylases by Plant Protein Inhibitors. Biochim. Biophys. Acta-Proteins Proteom. 2004, 1696, 171–180. [Google Scholar] [CrossRef]
- Climate-Data.org. Big Sandy Climate (United States of America). Available online: https://en.climate-data.org/north-america/united-states-of-america/montana/big-sandy-124832/ (accessed on 2 September 2022).
- Climate-Data.org. Grafico Temperatura Argelato. Available online: https://it.climate-data.org/europa/italia/emilia-romagna/argelato-14258/#temperature-graph (accessed on 2 September 2022).
- Priya, S.; Kumar, S.; Kaur, N.; Gupta, A.K. Specificity of α-Amylase and Trypsin Inhibitor Proteins in Wheat against Insect Pests. N. Z. J. Crop Hortic. Sci. 2013, 41, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Prandi, B.; Faccini, A.; Tedeschi, T.; Galaverna, G.; Sforza, S. LC/MS Analysis of Proteolytic Peptides in Wheat Extracts for Determining the Content of the Allergen Amylase/Trypsin Inhibitor CM3: Influence of Growing Area and Variety. Food Chem. 2013, 140, 141–146. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, S.; Call, L.; Edlmayr, M.; Marzak, S.; Jäger, H.; Domig, K.; Reiter, E.; Grausgruber, H. Characterization of Non-Gluten Proteins in Austrian Wheat Varieties from 19th Century until Today. Cereal Technol. 2018, 1, 1–10. [Google Scholar] [CrossRef]
- EL Hassouni, K.; Sielaff, M.; Curella, V.; Neerukonda, M.; Leiser, W.; Würschum, T.; Schuppan, D.; Tenzer, S.; Longin, C.F.H. Genetic Architecture Underlying the Expression of Eight α-Amylase Trypsin Inhibitors. Theor. Appl. Genet. 2021, 134, 3427–3441. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-R.; Tsen, C.C. Note on Trypsin Inhibitor Activity in the Acetate Extract of Cereal Samples. Cereal Chem. 1979, 56, 493–494. [Google Scholar]
- Taddei, F.; Gazza, L.; Conti, S.; Muccilli, V.; Foti, S.; Pogna, N.E. Starch-bound 2S proteins and kernel texture in einkorn, Triticum monococcum ssp. monococcum. Theor. Appl. Genet. 2009, 119, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Sielaff, M.; Curella, V.; Neerukonda, M.; Afzal, M.; El Hassouni, K.; Distler, U.; Schuppan, D.; Longin, C.F.H.; Tenzer, S. Hybrid QconCAT-Based Targeted Absolute and Data-Independent Acquisition-Based Label-Free Quantification Enables In-Depth Proteomic Characterization of Wheat Amylase/Trypsin Inhibitor Extracts. J. Proteome Res. 2021, 20, 1544–1557. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Kothari, K.M. Transcript Profiles of Genes Expressed in Endosperm Tissue Are Altered by High Temperature during Wheat Grain Development. J. Cereal Sci. 2004, 40, 115–126. [Google Scholar] [CrossRef]
- Juhász, A.; Belova, T.; Florides, C.G.; Maulis, C.; Fischer, I.; Gell, G.; Birinyi, Z.; Ong, J.; Keeble-Gagnère, G.; Maharajan, A.; et al. Genome Mapping of Seed-Borne Allergens and Immunoresponsive Proteins in Wheat. Sci. Adv. 2018, 4, eaar8602. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Jørgensen, A.D.; Li, H.; Søndergaard, I.; Finnie, C.; Svensson, B.; Jiang, D.; Wollenweber, B.; Jacobsen, S. Implications of High-Temperature Events and Water Deficits on Protein Profiles in Wheat (Triticum aestivum L. Cv. Vinjett) Grain. Proteomics 2011, 11, 1684–1695. [Google Scholar] [CrossRef]
- Gu, A.; Hao, P.; Lv, D.; Zhen, S.; Bian, Y.; Ma, C.; Xu, Y.; Zhang, W.; Yan, Y. Integrated Proteome Analysis of the Wheat Embryo and Endosperm Reveals Central Metabolic Changes Involved in the Water Deficit Response during Grain Development. J. Agric. Food Chem. 2015, 63, 8478–8487. [Google Scholar] [CrossRef] [PubMed]
- Cuccioloni, M.; Mozzicafreddo, M.; Ali, I.; Bonfili, L.; Cecarini, V.; Eleuteri, A.M.; Angeletti, M. Interaction between Wheat Alpha-Amylase/Trypsin Bi-Functional Inhibitor and Mammalian Digestive Enzymes: Kinetic, Equilibrium and Structural Characterization of Binding. Food Chem. 2016, 213, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Kawamori, R.; Tajima, N.; Iwamoto, Y.; Kashiwagi, A.; Shi-mamoto, K.; Kaku, K. Voglibose for prevention of type 2 diabetes mellitus: A randomised, double-blind trial in Japanese individuals with impaired glucose tolerance. Lancet 2009, 373, 1607–1614. [Google Scholar] [CrossRef]
- Bardella, M.T.; Elli, L.; Ferretti, F. Non Celiac Gluten Sensitivity. Curr. Gastroenterol. Rep. 2016, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Kodama, T.; Miyazaki, T.; Kitamura, I.; Suzuki, Y.; Namba, Y.; Sakurai, J.; Torikai, Y.; Inoue, S. Effects of Single and Long-Term Administration of Wheat Albumin on Blood Glucose Control: Randomized Controlled Clinical Trials. Eur. J. Clin. Nutr. 2005, 59, 384–392. [Google Scholar] [CrossRef]
- Birk, Y. Plant Protease Inhibitors: Significance in Nutrition. Plant Protection, Cancer Prevention and Genetic Engineering, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 52–96. [Google Scholar]
- Call, L.; Reiter, E.V.; Wenger-Oehn, G.; Strnad, I.; Grausgruber, H.; Schoenlechner, R.; D’Amico, S. Development of an Enzymatic Assay for the Quantitative Determination of Trypsin Inhibitory Activity in Wheat. Food Chem. 2019, 299, 125038. [Google Scholar] [CrossRef] [PubMed]
- Sofi, F.; Whittaker, A.; Gori, A.M.; Cesari, F.; Surrenti, E.; Abbate, R.; Gensini, G.F.; Benedettelli, S.; Casini, A. Effect of Triticum turgidum Subsp. Turanicum Wheat on Irritable Bowel Syndrome: A Double-Blinded Randomised Dietary Intervention Trial. Br. J. Nutr. 2014, 111, 1992–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacomino, G.; Rotondi Aufiero, V.; Di Stasio, L.; Picascia, S.; Iannaccone, N.; Giardullo, N.; Troncone, R.; Gianfrani, C.; Mamone, G.; Mazzarella, G. Triticum monococcum Amylase Trypsin Inhibitors Possess a Reduced Potential to Elicit Innate Immune Response in Celiac Patients Compared to Triticum aestivum. Food Res. Int. 2021, 145, 110386. [Google Scholar] [CrossRef]
- Huang, X.; Schuppan, D.; Tovar, L.E.R.; Zevallos, V.F.; Loponen, J.; Gänzle, M. Sourdough Fermentation Degrades Wheat Alpha-Amylase/Trypsin Inhibitor (ATI) and Reduces Pro-Inflammatory Activity. Foods 2020, 9, 943. [Google Scholar] [CrossRef]
- Kostekli, M.; Karakaya, S. Protease Inhibitors in Various Flours and Breads: Effect of Fermentation, Baking and in Vitro Digestion on Trypsin and Chymotrypsin Inhibitory Activities. Food Chem. 2017, 224, 62–68. [Google Scholar] [CrossRef]
- Mamone, G.; Nitride, C.; Picariello, G.; Addeo, F.; Ferranti, P.; Mackie, A. Tracking the Fate of Pasta (T. durum Semolina) Immunogenic Proteins by in Vitro Simulated Digestion. J. Agric. Food Chem. 2015, 63, 2660–2667. [Google Scholar] [CrossRef]
- Gélinas, P.; McKinnon, C.; Gagnon, F. Inhibitory Activity towards Human α-Amylase in Cereal Foods. LWT-Food Sci. Technol. 2018, 93, 268–273. [Google Scholar] [CrossRef]
- National Center of Environmental Information. Climate Data Online Data Tools. Available online: https://www.ncdc.noaa.gov/cdo-web/datatools/ (accessed on 9 October 2022).
- Arpae Emila Romagna. Available online: https://www.arpae.it/it (accessed on 9 October 2022).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| USA | 2015–2016 | 2016–2017 | 2017–2018 | |||
|---|---|---|---|---|---|---|
| Wheat Genotype | Protein (g/100g dm) | DON (ppb) | Protein (g/100g dm) | DON (ppb) | Protein (g/100g dm) | DON (ppb) |
| Einkorn | 26.7 (1.0) | <200 | 20.8 (1.1) | <200 | 20.8 (1.1) | <200 |
| Emmer | Sample not available | 19.7 (0.6) | <200 | 19.1 (0.8) | <200 | |
| Turanicum | 19.4 (0.6) | 253 (18.4) | 15.8 (0.6) | <200 | 17.1 (0.4) | <200 |
| Peliss | Sample not available | Sample not available | 18.5 (0.7) | <200 | ||
| Alzada | 24.7 (1.0) | 376 (22.6) | 17.3 (1.1) | <200 | 19.7 (0.7) | <200 |
| Spelt | 14.8 (0.4) | 204 (5.7) | 18.6 (0.1) | <200 | 19.5 (0.7) | <200 |
| Turkey Red | 14.6 (0.4) | <200 | 17.0 (0.7) | <200 | 16.8 (0.1) | <200 |
| Judee | 13.7 (0.1) | <200 | 17.0 (0.3) | <200 | 16.4 (0.6) | <200 |
| Marquis | 22.3 (1.4) | <200 | 18.5 (0.6) | <200 | 19.2 (0.3) | <200 |
| Vida | 18.6 (0.8) | 296 (8.5) | 17.8 (0.3) | <200 | 17.9 (0.3) | <200 |
| Italy | 2016–2017 | 2017–2018 | 2018–2019 | |||
| Wheat Genotype | Protein (g/100g dm) | DON (ppb) | Protein (g/100g dm) | DON (ppb) | Protein (g/100g dm) | DON (ppb) |
| Einkorn | 15.2 (0.3) | <200 | 14.0 (0.7) | <200 | 17.3 (0.4) | <200 |
| Emmer | 12.3 (0.4) | <200 | 12.5 (0.7) | <200 | 11.1 (0.1) | 219 (12.7) |
| Turanicum | 12.7 (0.3) | <200 | 12.8 (0.3) | 593 (18.4) | 10.3 (0.4) | 969 (12.7) |
| Peliss | 12.3 (0.4) | <200 | 13.2 (0.6) | 383 (18.4) | 10.1 (0.1) | 342 (17.0) |
| Alzada | 12.6 (0.1) | 299 (19.1) | 14.2 (0.4) | 547 (23.3) | 11.5 (0.7) | 1122 (45.3) |
| Spelt | 12.0 (0.1) | <200 | 13.8 (0.3) | <200 | 14.0 (0.7) | 260 (14.1) |
| Turkey Red | 11.3 (0.7) | <200 | 11.4 (0.1) | <200 | 10.1 (0.1) | 271 (15.6) |
| Judee | 11.4 (0.4) | <200 | 11.3 (0.1) | <200 | 10.1 (0.1) | 920 (14.1) |
| Marquis | 12.5 (0.1) | <200 | 12.9 (0.6) | <200 | 12.5 (0.6) | <200 |
| Vida | 12.7 (0.3) | <200 | 13.3 (0.4) | <200 | 13.1 (0.1) | 373 (11.3) |
| USA | Italy | ||
|---|---|---|---|
| Year (Y) | ** | Year (Y) | ** |
| 2015–2016 | 40.83 (b) | 2016–2017 | 53.43 (a) |
| 2016–2017 | 56.26 (a) | 2017–2018 | 47.06 (b) |
| 2017–2018 | 49.75 (a) | 2018–2019 | 38.52 (c) |
| Genotype (G) | ** | Genotype (G) | ** |
| Einkorn | 13.76 (d) | Einkorn | 1.12 (f) |
| Emmer | 56.62 (ab) | Emmer | 65.42 (a) |
| Turanicum | 39.6 (c) | Turanicum | 44.14 (cd) |
| Peliss | 59.73 (ab) | Peliss | 48.80 (c) |
| Alzada | 47.61 (bc) | Alzada | 33.28 (e) |
| Spelt | 63.15 (a) | Spelt | 59.95 (b) |
| Turkey Red | 66.63 (a) | Turkey Red | 56.42 (b) |
| Judee | 60.50 (a) | Judee | 57.47 (b) |
| Marquis | 49.24 (bc) | Marquis | 55.00 (b) |
| Vida | 45.35 (bc) | Vida | 41.77 (d) |
| G × Y | ** | G × Y | ** |
| USA | Italy | ||
|---|---|---|---|
| Year (Y) | ** | Year (Y) | ** |
| 2015–2016 | 63.31 (a) | 2016–2017 | 42.53 (a) |
| 2016–2017 | 43.72 (c) | 2017–2018 | 39.57 (b) |
| 2017–2018 | 49.04 (b) | 2018–2019 | 35.47 (c) |
| Genotype (G) | ** | Genotype (G) | ** |
| Einkorn | 100.00 (a) | Einkorn | 100.00 (a) |
| Emmer | 60.55 (d) | Emmer | 19.34 (e) |
| Turanicum | 23.08 (h) | Turanicum | 20.93 (d) |
| Peliss | 18.61 (i) | Peliss | 18.77 (e) |
| Alzada | 30.45 (f) | Alzada | 23.30 (d) |
| Spelt | 39.46 (e) | Spelt | 36.18 (c) |
| Turkey Red | 66.54 (c) | Turkey Red | 49.27 (b) |
| Judee | 71.06 (b) | Judee | 51.70 (b) |
| Marquis | 59.00 (d) | Marquis | 49.76 (b) |
| Vida | 27.26 (g) | Vida | 22.64 (d) |
| G × Y | ns | G × Y | ns |
| (a) | |||
|---|---|---|---|
| Year (Y) | Genotype (G) | G × Y | |
| USA | 37.5 ** | 55.3 ** | 7.2 ** |
| Italy | 34.1 ** | 62.9 ** | 2.3 ** |
| (b) | |||
| Year (Y) | Genotype (G) | G × Y | |
| USA | 32.8 ** | 64.7 ** | 1.8 ns |
| Italy | 6.0 * | 91.5 ** | 1.4 ns |
| Ploidy Level | Triticum Species | Genotype | Year of Release | Classification | Name Used in This Study |
|---|---|---|---|---|---|
| Diploid | T. monococcum L. subsp. monococcum | Local landrace a | / | Hulled, ancient wheat | Einkorn |
| Tetraploid | T. turgidum L. subsp. dicoccum | Local landrace a | / | Hulled, ancient wheat | Emmer |
| T. turgidum L. subsp. turanicum * | QK-77 b | / | Naked, ancient wheat | Turanicum | |
| T. turgidum L. subsp. durum | Peliss c | 1900 | Naked, heritage durum wheat | Peliss | |
| T. turgidum L. subsp. durum | Alzada a | 2004 | Naked, modern durum wheat | Alzada | |
| Hexaploid | T. aestivum L. subsp. spelta | Local landrace a | / | Hulled, ancient wheat | Spelt |
| T. aestivum L. subsp. aestivum | Turkey Red d | 1873 | Naked, heritage hard red common wheat | Turkey Red | |
| T. aestivum L. subsp. aestivum | Judee a | 2011 | Naked, modern hard red common wheat | Judee | |
| T. aestivum L. subsp. aestivum | Marquis a | 1913 | Naked, heritage hard red common wheat | Marquis | |
| T. aestivum L. subsp. aestivum | Vida a | 2006 | Naked, modern hard red common wheat | Vida |
| (a) | |||
|---|---|---|---|
| Soil Characteristics before Sowing | 2016–2017 | 2017–2018 | 2018–2019 |
| pH | 7.6 | 6.6 | 7.6 |
| Organic matter (% dm) | 1.2 | 3.3 | 3.7 |
| Total N (‰ dm) | 0.7 | 1.4 | 1.5 |
| C/N ratio | 9.7 | 13.6 | 14.7 |
| Assimilable p (mg/kg dm) | 16.9 | 8.3 | 13.8 |
| Assimilable K (mg/kg dm) | 190.1 | 178.5 | 182.3 |
| (b) | |||
| Soil Characteristics before Sowing | 2016–2017 | 2017–2018 | 2018–2019 |
| pH | 7.3 | 7.0 | 7.4 |
| Organic matter (% dm) | 1.5 | 1.8 | 2.4 |
| Total N (‰ dm) | 0.9 | 0.8 | 0.5 |
| C/N ratio | 9.6 | 13.1 | 27.9 |
| Assimilable p (mg/kg dm) | 14.2 | 12.1 | 11.6 |
| Assimilable K (mg/kg dm) | 261.1 | 239.4 | 211.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simonetti, E.; Bosi, S.; Negri, L.; Dinelli, G. Amylase Trypsin Inhibitors (ATIs) in a Selection of Ancient and Modern Wheat: Effect of Genotype and Growing Environment on Inhibitory Activities. Plants 2022, 11, 3268. https://doi.org/10.3390/plants11233268
Simonetti E, Bosi S, Negri L, Dinelli G. Amylase Trypsin Inhibitors (ATIs) in a Selection of Ancient and Modern Wheat: Effect of Genotype and Growing Environment on Inhibitory Activities. Plants. 2022; 11(23):3268. https://doi.org/10.3390/plants11233268
Chicago/Turabian StyleSimonetti, Emanuela, Sara Bosi, Lorenzo Negri, and Giovanni Dinelli. 2022. "Amylase Trypsin Inhibitors (ATIs) in a Selection of Ancient and Modern Wheat: Effect of Genotype and Growing Environment on Inhibitory Activities" Plants 11, no. 23: 3268. https://doi.org/10.3390/plants11233268
APA StyleSimonetti, E., Bosi, S., Negri, L., & Dinelli, G. (2022). Amylase Trypsin Inhibitors (ATIs) in a Selection of Ancient and Modern Wheat: Effect of Genotype and Growing Environment on Inhibitory Activities. Plants, 11(23), 3268. https://doi.org/10.3390/plants11233268