
Vegetation Classification and Distribution Patterns in the South Slope of Yarlung Zangbo Grand Canyon National Nature Reserve, Eastern Himalayas

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Vertical Vegetation Belts
2.1.1. Lower Montane Seasonal Rainforest Belt (Group 1)
2.1.2. Lower Montane Evergreen Broadleaf Forest Belt (Group 2)
2.1.3. Middle Montane Semi-Evergreen Broadleaf Forest Belt (Group 3)
2.1.4. Subalpine Evergreen Needleleaf Forest Belt (Group 4)
2.1.5. Alpine Shrubland and Meadow Belt (Group 5)
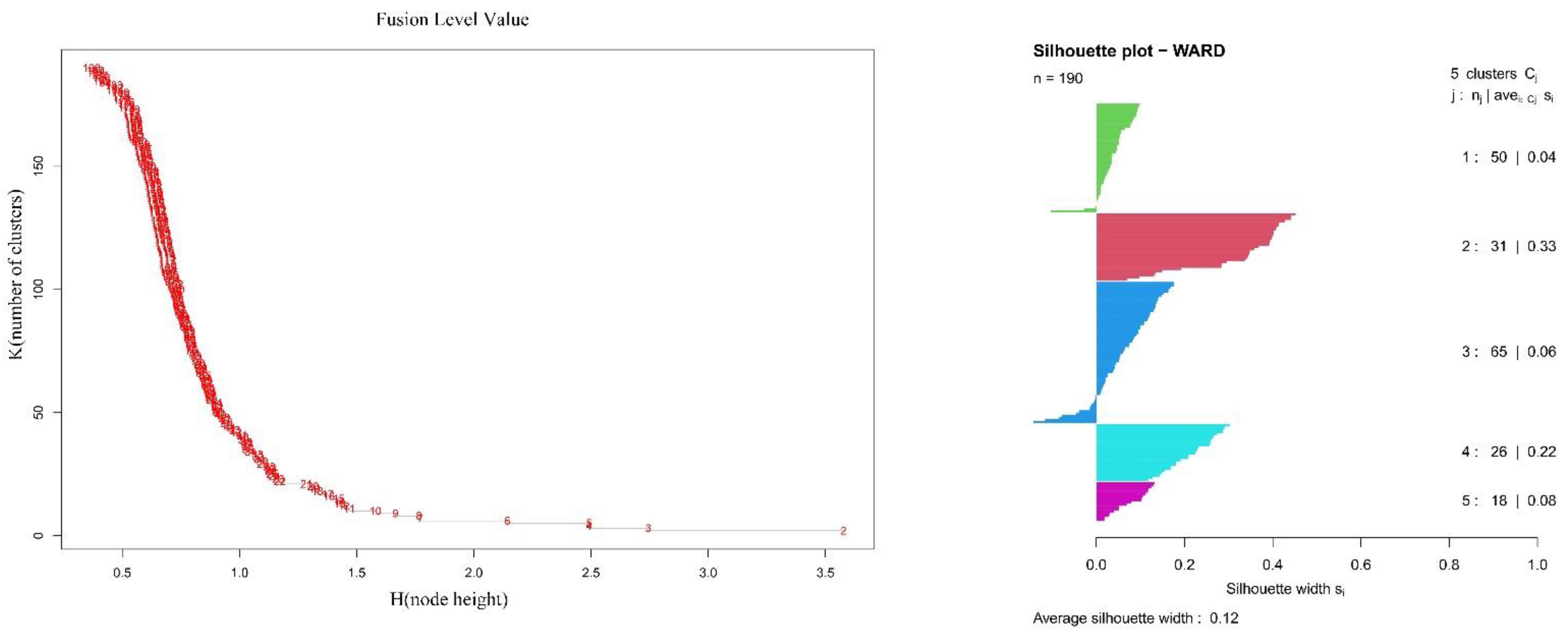
2.2. Ordination of Vegetation
2.3. Species Diversity
2.4. The New Division Scheme of Vertical Vegetation Belts
3. Discussion
3.1. Comparison of Vegetation Belts Distribution Schemes
3.2. The Unique Features of the Vertical Vegetation Belts
3.3. Vegetation Conservation
4. Materials and Methods
4.1. Fieldwork and Data Collection
4.2. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Group1 | Group2 | Group3 | Group4 | Group5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| IndVal | p | IndVal | p | IndVal | p | IndVal | p | IndVal | p | |
| Altingia excelsa | 0.411 | 0.001 | ||||||||
| Impatiens stenantha | 0.368 | 0.001 | ||||||||
| Phrynium placentarium | 0.339 | 0.001 | ||||||||
| Impatiens namchabarwensis | 0.314 | 0.002 | ||||||||
| Sambucus ebulus | 0.298 | 0.002 | ||||||||
| Mussaenda decipiensis | 0.291 | 0.004 | ||||||||
| Terminalia myriocarpa | 0.284 | 0.006 | ||||||||
| Boehmeria macrophylla var. rotundifolia | 0.279 | 0.004 | ||||||||
| Lagerstroemia minuticarpa | 0.276 | 0.008 | ||||||||
| Blumea balsamifera | 0.275 | 0.004 | ||||||||
| Cordia dichotoma | 0.26 | 0.006 | ||||||||
| Macaranga denticulata | 0.258 | 0.008 | ||||||||
| Trachelospermum jasminoides | 0.253 | 0.007 | ||||||||
| Dichrocephala benthamii | 0.252 | 0.014 | ||||||||
| Polygonum chinense | 0.245 | 0.007 | ||||||||
| Gynostemma pentaphyllum | 0.238 | 0.018 | ||||||||
| Poikilospermum suaveolens | 0.238 | 0.02 | ||||||||
| Equisetum diffusum | 0.231 | 0.035 | ||||||||
| Chrysopogon aciculatus | 0.225 | 0.032 | ||||||||
| Syngonium podophyllum | 0.224 | 0.029 | ||||||||
| Castanopsis indica | 0.901 | 0.001 | ||||||||
| Glochidion hirsutum | 0.462 | 0.001 | ||||||||
| Oplismenus undulatifolius | 0.404 | 0.001 | ||||||||
| Triumfetta cana | 0.368 | 0.001 | ||||||||
| Pteris cretica var. nervosa | 0.333 | 0.001 | ||||||||
| Desmodium sequax | 0.32 | 0.001 | ||||||||
| Colocasia antiquorum | 0.314 | 0.003 | ||||||||
| Strobilanthes dimorphotricha | 0.314 | 0.001 | ||||||||
| Polygonum capitatum | 0.307 | 0.002 | ||||||||
| Saurauia napaulensis | 0.296 | 0.002 | ||||||||
| Elatostema acuminatum | 0.287 | 0.009 | ||||||||
| Circaea cordata | 0.287 | 0.009 | ||||||||
| Chirita pumila | 0.287 | 0.013 | ||||||||
| Dioscorea hispida | 0.287 | 0.009 | ||||||||
| Solena amplexicaulis | 0.287 | 0.006 | ||||||||
| Hedyotis scandens | 0.287 | 0.004 | ||||||||
| Wallichia disticha | 0.257 | 0.006 | ||||||||
| Ophiorrhiza mungos | 0.253 | 0.019 | ||||||||
| Impatiens arguta | 0.25 | 0.017 | ||||||||
| Engelhardtia spicata | 0.249 | 0.009 | ||||||||
| Cyclobalanopsis lamellosa | 0.571 | 0.001 | ||||||||
| Exbucklandia populnea | 0.378 | 0.001 | ||||||||
| Cyclobalanopsis kiukiangensis | 0.367 | 0.001 | ||||||||
| Damnacanthus indicus | 0.361 | 0.001 | ||||||||
| Pholidota articulata | 0.352 | 0.001 | ||||||||
| Ficus sarmentosa | 0.348 | 0.001 | ||||||||
| Disporum bodinieri | 0.342 | 0.001 | ||||||||
| Arisaema concinum | 0.341 | 0.001 | ||||||||
| Ainsliaea latifolia | 0.315 | 0.001 | ||||||||
| Vaccinium kingdom-wardii | 0.31 | 0.001 | ||||||||
| Myrsine semiserrata | 0.306 | 0.002 | ||||||||
| Pinus bhutanica | 0.29 | 0.003 | ||||||||
| Remusatia vivipara | 0.289 | 0.003 | ||||||||
| Toxicodendron hookeri var. microcarpum | 0.288 | 0.001 | ||||||||
| Ilex wilsonii | 0.288 | 0.002 | ||||||||
| Viburnum cylindricum | 0.287 | 0.001 | ||||||||
| Betula cylindrostachya | 0.277 | 0.004 | ||||||||
| Pyrrosia lanceolata | 0.269 | 0.004 | ||||||||
| Tupistra aurantiaca | 0.268 | 0.005 | ||||||||
| Taxillus dalavayi | 0.267 | 0.004 | ||||||||
| Tsuga dumosa | 0.717 | 0.001 | ||||||||
| Abies delavayi var. motuoensis | 0.69 | 0.001 | ||||||||
| Acanthopanax evodiaefolius | 0.49 | 0.001 | ||||||||
| Ribes glaciale | 0.483 | 0.001 | ||||||||
| Lindera obtusiloba | 0.443 | 0.001 | ||||||||
| Pilea notata | 0.434 | 0.001 | ||||||||
| Acer campbellii | 0.426 | 0.001 | ||||||||
| Synotis alata | 0.375 | 0.001 | ||||||||
| Circaea alpina | 0.37 | 0.001 | ||||||||
| Smilacina fusca | 0.359 | 0.001 | ||||||||
| Lonicera tangutica | 0.353 | 0.002 | ||||||||
| Lunathyrium medogense | 0.353 | 0.001 | ||||||||
| Oxalis acetosella subsp. Leucolepis | 0.348 | 0.001 | ||||||||
| Gaultheria trichophylla | 0.347 | 0.001 | ||||||||
| Dysosma tsayuensis | 0.345 | 0.003 | ||||||||
| Acer sp. | 0.335 | 0.001 | ||||||||
| Arisaema rhizomatum | 0.333 | 0.002 | ||||||||
| Arisaema biauriculatum | 0.328 | 0.001 | ||||||||
| Mahonia fortunei | 0.32 | 0.003 | ||||||||
| Euonymus alatus | 0.316 | 0.001 | ||||||||
| Pleurospermum angelicoides | 0.534 | 0.001 | ||||||||
| Dryopteris barbigera | 0.511 | 0.001 | ||||||||
| Viola biflora | 0.486 | 0.001 | ||||||||
| Athyrium attenuatum | 0.482 | 0.001 | ||||||||
| Geranium polyanthes | 0.474 | 0.001 | ||||||||
| Cardamine macrophylla | 0.469 | 0.001 | ||||||||
| Pedicularis lineata | 0.467 | 0.001 | ||||||||
| Polygonum polystachyum | 0.466 | 0.001 | ||||||||
| Rosa sericea | 0.461 | 0.001 | ||||||||
| Rhododendron viridescens | 0.457 | 0.001 | ||||||||
| Ribes orientale | 0.456 | 0.001 | ||||||||
| Polygonum viviparum | 0.445 | 0.001 | ||||||||
| Cirsium eriophoroides | 0.445 | 0.001 | ||||||||
| Gaultheria dolichopoda | 0.444 | 0.001 | ||||||||
| Anaphalis margaritacea var. japonica | 0.44 | 0.001 | ||||||||
| Lonicera angustifolia var. myrtillus | 0.425 | 0.001 | ||||||||
| Rhodiola rosea | 0.416 | 0.001 | ||||||||
| Saxifraga sp. | 0.416 | 0.001 | ||||||||
| Polygonatum verticillatum | 0.408 | 0.001 | ||||||||
| Senecio lingianas | 0.396 | 0.001 | ||||||||
Appendix C
| Plot | Elevation | Longitude | Latitude | Aspect | Slope |
|---|---|---|---|---|---|
| CaIn32 | 635 | 95.051 | 29.185 | 153.43 | 25.84 |
| CaIn21 | 718 | 95.054 | 29.188 | 286.02 | 39.18 |
| made3 | 759 | 95.084 | 29.188 | 191.59 | 18.35 |
| CaIn10 | 815 | 95.103 | 29.215 | 247.61 | 20.95 |
| lixi2 | 664 | 95.004 | 29.182 | 202.69 | 13.94 |
| cain | 867 | 95.021 | 29.181 | 136.25 | 28.20 |
| made2 | 1518 | 95.131 | 29.256 | 187.35 | 38.00 |
| CaIn6 | 1557 | 95.129 | 29.253 | 7.41 | 43.13 |
| fise5 | 1415 | 95.14 | 29.252 | 157.28 | 21.22 |
| oxpa | 1472 | 95.156 | 29.269 | 116.68 | 42.57 |
| fiau | 1152 | 95.18 | 29.331 | 292.03 | 20.68 |
| expo8 | 2079 | 95.348 | 29.308 | 193.09 | 20.19 |
| pana | 2029 | 95.354 | 29.303 | 187.59 | 7.19 |
| dibu | 1615 | 95.179 | 29.224 | 85.31 | 14.30 |
| expi | 1621 | 95.182 | 29.228 | 270.00 | 28.07 |
| cyki6 | 1479 | 95.153 | 29.206 | 214.17 | 22.19 |
| CaIn5 | 1677 | 95.14 | 29.187 | 11.85 | 19.02 |
| expo7 | 1843 | 95.168 | 29.21 | 334.33 | 19.59 |
| cyla18 | 1863 | 95.168 | 29.21 | 317.68 | 20.68 |
| cyki1 | 1644 | 95.189 | 29.225 | 227.60 | 14.54 |
| expo6 | 1823 | 95.169 | 29.219 | 2.94 | 36.13 |
| sapo4 | 1679 | 95.178 | 29.226 | 90.90 | 28.07 |
| lipu | 1018 | 95.132 | 29.214 | 321.74 | 28.89 |
| alex19 | 669 | 95.127 | 29.222 | 310.33 | 16.15 |
| temy4 | 814 | 95.147 | 29.232 | 350.39 | 15.35 |
| alex8 | 1200 | 95.155 | 29.255 | 119.05 | 34.46 |
| temy3 | 595 | 94.999 | 29.183 | 203.19 | 31.08 |
| CaIn4 | 631 | 95.069 | 29.187 | 166.07 | 28.21 |
| CaIn3 | 895 | 95.174 | 29.24 | 320.42 | 43.08 |
| alex7 | 853 | 95.134 | 29.222 | 343.30 | 38.06 |
| alex6 | 859 | 95.175 | 29.247 | 234.59 | 22.99 |
| alex5 | 1648 | 95.174 | 29.214 | 52.25 | 18.09 |
| ensp2 | 722 | 95.162 | 29.255 | 123.08 | 33.28 |
| CaIn2 | 758 | 95.169 | 29.27 | 113.19 | 15.93 |
| alex4 | 1115 | 95.206 | 29.263 | 60.26 | 3.84 |
| CaIn1 | 1127 | 95.207 | 29.26 | 326.53 | 16.41 |
| CaIn31 | 1063 | 95.213 | 29.262 | 325.30 | 7.51 |
| CaIn30 | 1063 | 95.213 | 29.262 | 325.30 | 7.51 |
| CaIn29 | 789 | 95.246 | 29.268 | 38.66 | 3.05 |
| CaIn28 | 865 | 95.264 | 29.285 | 320.35 | 10.72 |
| tsdu11 | 2458 | 95.099 | 29.395 | 127.47 | 17.48 |
| tsdu8 | 2361 | 95.103 | 29.391 | 233.13 | 24.62 |
| cyts | 2377 | 95.108 | 29.387 | 232.98 | 34.15 |
| cyki5 | 2308 | 95.111 | 29.381 | 229.35 | 31.82 |
| cyla9 | 2202 | 95.117 | 29.373 | 180.00 | 15.37 |
| cyla8 | 2189 | 95.117 | 29.372 | 58.32 | 12.95 |
| cyla7 | 1990 | 95.135 | 29.358 | 70.11 | 11.76 |
| cyki4 | 1847 | 95.144 | 29.354 | 170.28 | 37.42 |
| expo5 | 1958 | 95.153 | 29.351 | 40.49 | 12.65 |
| cyki3 | 1877 | 95.162 | 29.347 | 82.21 | 21.77 |
| fise4 | 1139 | 95.179 | 29.331 | 127.14 | 9.79 |
| cyla6 | 1224 | 95.172 | 29.329 | 278.13 | 31.74 |
| CaIn27 | 842 | 95.244 | 29.264 | 62.30 | 20.63 |
| CaIn26 | 820 | 95.284 | 29.299 | 278.24 | 31.40 |
| CaIn25 | 820 | 95.279 | 29.325 | 339.53 | 16.59 |
| CaIn24 | 887 | 95.382 | 29.414 | 171.09 | 19.29 |
| figl | 797 | 95.358 | 29.39 | 132.31 | 20.68 |
| alex3 | 1407 | 95.225 | 29.252 | 18.43 | 4.52 |
| alex2 | 1487 | 95.226 | 29.258 | 136.24 | 15.16 |
| cahy2 | 1393 | 95.22 | 29.259 | 306.20 | 30.04 |
| CaIn23 | 1216 | 95.341 | 29.367 | 132.43 | 21.56 |
| CaIn22 | 856 | 95.347 | 29.366 | 115.49 | 16.70 |
| CaIn20 | 746 | 95.319 | 29.335 | 147.45 | 27.42 |
| CaIn19 | 802 | 95.304 | 29.326 | 161.00 | 15.04 |
| fise3 | 1125 | 95.265 | 29.32 | 65.67 | 35.67 |
| cete | 835 | 95.299 | 29.325 | 146.51 | 17.99 |
| lixi1 | 1830 | 95.19 | 29.296 | 188.13 | 32.95 |
| expo4 | 1617 | 95.184 | 29.277 | 61.93 | 15.81 |
| CaIn18 | 1407 | 95.242 | 29.297 | 202.59 | 31.72 |
| lami | 896 | 95.393 | 29.411 | 1.81 | 21.60 |
| temy2 | 849 | 95.387 | 29.41 | 347.59 | 23.10 |
| CaIn17 | 1014 | 95.384 | 29.406 | 316.12 | 16.72 |
| alex1 | 952 | 95.38 | 29.392 | 344.74 | 18.39 |
| cyki2 | 1998 | 95.381 | 29.323 | 108.68 | 31.44 |
| cyki8 | 2107 | 95.37 | 29.32 | 156.25 | 6.49 |
| cyla5 | 1984 | 95.38 | 29.313 | 60.12 | 12.72 |
| cyki7 | 2044 | 95.377 | 29.317 | 49.40 | 8.74 |
| CaIn16 | 2043 | 95.365 | 29.332 | 5.71 | 14.10 |
| cyla4 | 2136 | 95.359 | 29.298 | 303.99 | 12.19 |
| rhde | 1940 | 95.358 | 29.309 | 17.56 | 19.04 |
| coca2 | 1843 | 95.36 | 29.331 | 257.30 | 25.36 |
| alsh | 1084 | 95.362 | 29.356 | 188.91 | 40.70 |
| alex18 | 923 | 95.353 | 29.351 | 174.17 | 39.38 |
| alca | 2435 | 95.36 | 29.287 | 148.67 | 12.64 |
| cyla1 | 2303 | 95.347 | 29.291 | 23.59 | 38.34 |
| ospa | 1308 | 95.324 | 29.313 | 322.22 | 22.86 |
| alex17 | 1365 | 95.322 | 29.309 | 297.83 | 22.75 |
| sapo3 | 824 | 95.323 | 29.332 | 301.90 | 27.48 |
| alex16 | 754 | 95.31 | 29.324 | 326.04 | 13.82 |
| fise2 | 1261 | 95.343 | 29.343 | 287.14 | 25.01 |
| pibh6 | 1579 | 95.184 | 29.224 | 333.43 | 2.13 |
| made1 | 832 | 95.383 | 29.413 | 153.71 | 37.64 |
| alex15 | 947 | 95.38 | 29.416 | 190.00 | 26.72 |
| alex14 | 895 | 95.408 | 29.42 | 261.86 | 34.12 |
| pibh5 | 1997 | 95.349 | 29.303 | 62.10 | 4.58 |
| coca1 | 1812 | 95.359 | 29.33 | 260.86 | 25.29 |
| pibh4 | 1814 | 95.193 | 29.195 | 309.80 | 11.04 |
| alex13 | 1340 | 95.448 | 29.471 | 223.38 | 22.71 |
| caec2 | 1434 | 95.453 | 29.468 | 236.50 | 34.19 |
| alex12 | 1407 | 95.455 | 29.478 | 305.70 | 18.18 |
| CaIn15 | 1343 | 95.453 | 29.489 | 333.43 | 14.62 |
| cyla3 | 2028 | 95.487 | 29.501 | 125.44 | 31.11 |
| pibh3 | 1528 | 95.512 | 29.496 | 43.85 | 16.41 |
| expo3 | 1657 | 95.558 | 29.458 | 28.44 | 12.81 |
| expo2 | 1798 | 95.571 | 29.452 | 357.66 | 42.66 |
| caec1 | 1691 | 95.563 | 29.46 | 199.35 | 18.09 |
| deti | 1797 | 95.444 | 29.442 | 338.11 | 45.16 |
| muba | 876 | 95.444 | 29.462 | 347.08 | 24.98 |
| cahy1 | 1651 | 95.558 | 29.459 | 210.96 | 17.52 |
| lixy | 1785 | 95.59 | 29.454 | 160.01 | 5.57 |
| pibh2 | 1750 | 95.606 | 29.441 | 298.30 | 7.01 |
| saps2 | 1977 | 95.66 | 29.429 | 346.84 | 24.60 |
| pibh1 | 2155 | 95.668 | 29.447 | 180.83 | 40.78 |
| expo1 | 2001 | 95.685 | 29.439 | 323.23 | 24.34 |
| jusi3 | 2336 | 95.686 | 29.459 | 180.00 | 4.29 |
| saps1 | 2151 | 95.721 | 29.462 | 159.37 | 22.49 |
| jusi2 | 2641 | 95.71 | 29.469 | 128.75 | 20.77 |
| powi | 2192 | 95.776 | 29.468 | 189.36 | 21.02 |
| saps4 | 2821 | 95.739 | 29.553 | 296.56 | 8.48 |
| saps3 | 2788 | 95.737 | 29.548 | 218.29 | 5.76 |
| tsdu7 | 2621 | 95.743 | 29.515 | 290.46 | 16.59 |
| tsdu6 | 2605 | 95.747 | 29.508 | 198.43 | 15.46 |
| cyla2 | 2492 | 95.753 | 29.5 | 264.55 | 9.97 |
| temy1 | 826 | 95.441 | 29.461 | 296.38 | 31.35 |
| rhsa3 | 3498 | 94.971 | 29.481 | 81.53 | 11.19 |
| rhsa2 | 3498 | 94.971 | 29.481 | 81.53 | 11.19 |
| rhsa1 | 3498 | 94.971 | 29.481 | 81.53 | 11.19 |
| safl3 | 3613 | 94.966 | 29.481 | 74.95 | 21.86 |
| safl2 | 3613 | 94.966 | 29.481 | 74.95 | 21.86 |
| safl1 | 3613 | 94.966 | 29.481 | 74.95 | 21.86 |
| rhch | 4255 | 94.947 | 29.487 | 9.32 | 15.79 |
| sare4 | 3834 | 94.963 | 29.486 | 144.21 | 48.74 |
| sare3 | 3834 | 94.963 | 29.486 | 144.21 | 48.74 |
| sare2 | 3834 | 94.963 | 29.486 | 144.21 | 48.74 |
| abmo10 | 3233 | 95.009 | 29.463 | 237.65 | 16.48 |
| abmo8 | 3186 | 95.014 | 29.458 | 231.34 | 36.76 |
| abmo7 | 2969 | 95.021 | 29.45 | 231.65 | 19.59 |
| ensp1 | 2023 | 95.493 | 29.655 | 124.87 | 18.53 |
| sapo2 | 1770 | 95.485 | 29.637 | 258.94 | 20.27 |
| cyla17 | 2229 | 95.499 | 29.678 | 106.69 | 27.56 |
| tsdu5 | 2454 | 95.518 | 29.694 | 136.34 | 26.61 |
| tsdu4 | 2602 | 95.522 | 29.703 | 253.55 | 24.52 |
| tsdu3 | 2813 | 95.594 | 29.713 | 187.12 | 15.04 |
| abmo6 | 3007 | 95.614 | 29.716 | 207.47 | 6.70 |
| abmo5 | 3213 | 95.645 | 29.726 | 203.62 | 8.28 |
| fise1 | 1576 | 95.478 | 29.613 | 96.43 | 29.14 |
| abmo4 | 3424 | 95.673 | 29.737 | 154.23 | 15.02 |
| abmo3 | 3589 | 95.681 | 29.741 | 181.07 | 41.72 |
| jusi1 | 3712 | 95.696 | 29.753 | 158.42 | 43.92 |
| CaIn14 | 1721 | 95.138 | 29.179 | 336.80 | 1.82 |
| CaIn13 | 1551 | 95.15 | 29.171 | 71.57 | 12.62 |
| CaIn12 | 1540 | 95.156 | 29.193 | 242.35 | 30.65 |
| cyla16 | 2048 | 95.384 | 29.309 | 7.43 | 5.52 |
| cyla15 | 2034 | 95.382 | 29.31 | 1.00 | 8.53 |
| cyla14 | 2280 | 95.71 | 29.444 | 335.44 | 20.35 |
| cyla13 | 2218 | 95.708 | 29.453 | 305.21 | 9.84 |
| abmo2 | 3273 | 95.976 | 29.488 | 51.73 | 49.16 |
| abmo1 | 3151 | 95.971 | 29.475 | 116.80 | 24.36 |
| abmo9 | 2918 | 95.959 | 29.459 | 111.50 | 16.46 |
| tsdu2 | 2827 | 95.955 | 29.453 | 156.54 | 13.53 |
| tsdu1 | 2626 | 95.888 | 29.447 | 220.03 | 7.75 |
| tsdu10 | 2519 | 95.843 | 29.466 | 194.56 | 28.66 |
| tsdu9 | 2404 | 95.827 | 29.469 | 191.30 | 9.65 |
| cyla12 | 2288 | 95.808 | 29.467 | 198.43 | 18.23 |
| alex11 | 1152 | 95.374 | 29.682 | 223.31 | 11.33 |
| sapo1 | 1723 | 95.372 | 29.646 | 296.20 | 41.27 |
| cace | 1569 | 95.391 | 29.639 | 155.19 | 28.64 |
| rhme12 | 4219 | 95.696 | 29.753 | 136.63 | 31.75 |
| rhme1 | 4130 | 95.698 | 29.752 | 138.57 | 10.69 |
| saan2 | 4000 | 95.698 | 29.747 | 191.18 | 36.14 |
| saan1 | 3907 | 95.695 | 29.746 | 208.51 | 37.53 |
| sare1 | 3789 | 95.695 | 29.744 | 187.76 | 36.50 |
| cyla11 | 2134 | 95.32 | 29.738 | 258.56 | 36.33 |
| cyla10 | 2462 | 95.389 | 29.692 | 225.60 | 28.98 |
| alex10 | 1473 | 95.404 | 29.621 | 287.96 | 17.95 |
| CaIn11 | 1623 | 95.399 | 29.603 | 345.59 | 38.21 |
| alex9 | 1561 | 95.387 | 29.59 | 228.86 | 41.15 |
| CaIn9 | 1505 | 95.402 | 29.569 | 214.00 | 39.84 |
| CaIn8 | 1929 | 95.402 | 29.542 | 343.94 | 48.85 |
| CaIn7 | 1707 | 95.427 | 29.523 | 247.68 | 40.55 |
| povi | 4255 | 94.947 | 29.487 | 9.32 | 15.79 |
| bepu | 4255 | 94.947 | 29.487 | 9.32 | 15.79 |
| amhi | 1253 | 95.17 | 29.331 | 222.39 | 14.54 |
| fise6 | 1314 | 95.171 | 29.337 | 247.14 | 28.68 |
| expo9 | 1754 | 95.163 | 29.345 | 132.31 | 37.05 |
| tsdu12 | 2875 | 95.047 | 29.43 | 174.55 | 9.97 |
| tsdu13 | 2764 | 95.059 | 29.42 | 173.89 | 33.67 |
| tsdu14 | 2703 | 95.067 | 29.416 | 180.00 | 15.37 |
| tsdu15 | 2603 | 95.079 | 29.411 | 121.60 | 7.25 |
| tsdu16 | 2486 | 95.092 | 29.401 | 113.74 | 6.49 |
References
- Fattorini, S.; Di Biase, L.; Chiarucci, A. Recognizing and interpreting vegetational belts: New wine in the old bottles of a von Humboldt’s legacy. J. Biogeogr. 2019, 46, 1643–1651. [Google Scholar] [CrossRef]
- Von Humboldt, A.; Bonpland, A. Essai Sur la Géographie des Plantes; Levrault, Schoell et Compagnie: Paris, France, 1805. [Google Scholar]
- Morueta-Holme, N.; Engemann, K.; Sandoval-Acuna, P.; Jonas, J.D.; Segnitz, R.M.; Svenning, J.-C. Strong upslope shifts in Chimborazo’s vegetation over two centuries since Humboldt. Proc. Natl. Acad. Sci. USA 2015, 112, 12741–12745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daubenmire, R.F. Vegetational zonation in the Rocky Mountains. Bot. Rev. 1943, 6, 326–393. [Google Scholar] [CrossRef]
- Shu-Qing, Z.; Jing-Yun, F.; Zhan-Jiang, Z.; Biao, Z.; Hai-Hua, S. Composition, structure and species diversity of plant communities along an altitudinal gradient on the northern slope of Mt.Changbai, Northeast China. Biodivers. Sci. 2004, 12, 164–173. [Google Scholar]
- Just, T.H. The vegetation of the Eastern Alps. Ecology 1940, 21, 270–271. [Google Scholar] [CrossRef]
- Van der Plas, G.W.; Rucina, S.M.; Hemp, A.; Marchant, R.A.; Hooghiemstra, H.; Schueler, L.; Verschuren, D. Climate-human-landscape interaction in the eastern foothills of Mt. Kilimanjaro (equatorial East Africa) during the last two millennia. Holocene 2021, 31, 556–569. [Google Scholar] [CrossRef]
- Hemp, A. Continuum or zonation? Altitudinal gradients in the forest vegetation of Mt. Kilimanjaro. Plant Ecol. 2006, 184, 27–42. [Google Scholar] [CrossRef]
- Bo-Sheng, L. The vertical spectra of vegetation in the MT. Namjabarwa region. Mt. Res. 1984, 2, 174–181. [Google Scholar]
- Lu, Z. Add Himalayas’ Grand Canyon to China’s first national parks. Nature 2021, 592, 353. [Google Scholar]
- Yan, H.R.; Huang, J.P.; He, Y.L.; Liu, Y.Z.; Wang, T.H.; Li, J.M. Atmospheric Water Vapor Budget and Its Long-Term Trend Over the Tibetan Plateau. J. Geophys. Res.-Atmos. 2020, 125, 17. [Google Scholar] [CrossRef]
- Ma, Y.Z.; Lu, M.Q.; Bracken, C.; Chen, H.N. Spatially coherent clusters of summer precipitation extremes in the Tibetan Plateau: Where is the moisture from? Atmos. Res. 2020, 237, 7. [Google Scholar] [CrossRef]
- Li, B. The zonation of vegetation in the MT.Namjagbarwa region. Mt. Res. 1985, 3, 291–298. [Google Scholar]
- Zhu, H. Geographical elements of seed plants suggest the boundary of the tropical zone in China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 386, 16–22. [Google Scholar] [CrossRef]
- Zhu, H. suggestions for the northern boundary of the tropical zone in China. Plant Sci. J. 2018, 36, 893–898. [Google Scholar]
- Brooks, T.M.; Mittermeier, R.A.; da Fonseca, G.A.B.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S.L. Global Biodiversity Conservation Priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, C. Biodiversity hotspots: A shortcut for a more complicated concept. Glob. Ecol. Conserv. 2015, 3, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.C.; Cheng, S.Z. The Flora of the Vascular Plants in MT. Namjagbarwa Region; Beijing Science and Technology Press: Beijing, China, 1992; pp. 1–96. [Google Scholar]
- Wu, P.P.; Wang, Z.; Jia, N.X.; Guo, S.W.; Liu, C.C.; Jin, X.H.; Guo, K. Calanthe x yarlungzangboensis, a new natural hybrid in genus Calanthe (Orchidaceae) from China. Phytotaxa 2021, 518, 167–174. [Google Scholar] [CrossRef]
- Bi, W.-X.; Chen, C.-C.; Lin, M.-Y. First record of Jacobsoniidae (Coleoptera) from China with description of a new species of Sarothrias Grouvelle. ZooKeys 2015, 496, 53–60. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Gorochov, A.V. A new genus for a new species of Podoscirtini from southeast Tibet (Orthoptera: Gryllidae; Podoscirtinae; Podoscirtini). Zootaxa 2015, 4033, 259–264. [Google Scholar] [CrossRef]
- Liu, C.; Ya, J.-D.; Tan, Y.-H.; He, H.-J.; Dong, G.-J.; Li, D.-Z. Marsdenia yarlungzangboensis (Apocynaceae, Asclepiadoideae), a new species from Xizang, China. Phytokeys 2019, 130, 85–92. [Google Scholar] [CrossRef]
- Wang, W.-G.; Lang, X.-A.; Yang, L.-L.; Wu, H.; Zhang, S.-Z. Begonia zhongyangiana, a new species of Begonia (Begoniaceae) from western China. Phytotaxa 2019, 407, 51–58. [Google Scholar] [CrossRef]
- Ya, J.-D.; Guo, Y.-J.; Liu, C.; Cai, J.; Dong, G.-J.; Jiang, H.; Li, D.-Z. Bulbophyllum reflexipetalum (Orchidaceae, Epidendroideae, Malaxideae), a new species from Xizang, China. Phytokeys 2019, 130, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Han, H.L.; Kononenko, V.S. Three new species of the genus Araeopteron Hampson, 1893 (Lepidoptera, Erebidae, Boletobiinae) from the Xizang Autonomous Region, China with an updated list of the world species. Zookeys 2021, 1060, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Y.; Li, W. A new species of Neoperla (Plecoptera: Perlidae) from Motuo County of Tibet, China and redescription of Neoperla perspicillata Zwick, 1980. Zootaxa 2021, 4964, 169–178. [Google Scholar] [CrossRef]
- Tong, Y.-H.; Zhao, W.-L.; Wang, B.-M.; Liu, E.-D.; Cai, J.; Guo, Y.-J. Vaccinium motuoense (Ericaceae), a new species from Xizang, China. Phytokeys 2021, 181, 105–111. [Google Scholar] [CrossRef]
- Du, C.; Liu, J.; Ye, W.; Liao, S.; Ge, B.; Liu, B.; Ma, J. Annual report of new taxa and new names for Chinese plants in 2020. Biodivers. Sci. 2021, 29, 1011–1020. [Google Scholar] [CrossRef]
- Du, Z.; Wei-Lie, C. A preliminary study on the vertical belts of vegetation of the eastern himalayas. Acta Bot. Sin. 1981, 23, 228–234. [Google Scholar]
- Hang, S.; Zhe-Kun, Z. Seed Plants of the Big Bend Gorge of Yalu Tsangpo in Tibet, E Himalayas; Yunnan Science and Technology Press: Yunan, China, 2002. [Google Scholar]
- Peng, B.; Pu, L.; Bao, H.; Higgitt, D.L. Vertical Zonation of Landscape Characteristics in the Namjagbarwa Massif of Tibet, China. Mt. Res. Dev. 1997, 17, 43–48. [Google Scholar]
- Chi, Y.; Shi, H.H.; Zheng, W.; Sun, J.K.; Fu, Z.Y. Spatiotemporal characteristics and ecological effects of the human interference index of the Yellow River Delta in the last 30 years. Ecol. Indic. 2018, 89, 880–892. [Google Scholar] [CrossRef]
- Han, D.; Gao, C.; Yu, Z.; Yu, X.; Li, Y.; Cong, J.; Wang, G. Late Holocene vegetation and climate changes in the Great Hinggan Mountains, northeast China. Quat. Int. 2019, 532, 138–145. [Google Scholar] [CrossRef]
- Prokop, P.; PLoSkonka, D. Natural and human impact on the land use and soil properties of the Sikkim Himalayas piedmont in India. J. Environ. Manag. 2014, 138, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Lotterhos, K.E.; Laruson, A.J.; Jiang, L.-Q. Novel and disappearing climates in the global surface ocean from 1800 to 2100. Sci. Rep. 2021, 11, 15535. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. The plateau zonality of vegetation in Xizang. Acta Bot. Sin. 1978, 20, 140–149. [Google Scholar]
- Flora Reipublicae Popularis Sinicae. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1959–2004. [Google Scholar]
- Li, B. A semi-evergreen broadleaf forest on the south slope of the eatern hmimalayas. Acta Bot. Sin. 1985, 27, 334–336. [Google Scholar]
- Zhang, J.W.; Jiang, S. A primary study on the vertical vegetation belt of Mt. Jolmo-Lungma (Everest) region and its relationship with horizontal zone. Acta Bot. Sin. 1973, 15, 221–236. [Google Scholar]
- Rodwell, J.S.; Evans, D.; Schaminee, J.H.J. Phytosociological relationships in European Union policy-related habitat classifications. Rend. Lincei-Sci. Fis. E Nat. 2018, 29, 237–249. [Google Scholar] [CrossRef]
- You, Q.L.; Fraedrich, K.; Ren, G.Y.; Pepin, N.; Kang, S.C. Variability of temperature in the Tibetan Plateau based on homogenized surface stations and reanalysis data. Int. J. Climatol. 2013, 33, 1337–1347. [Google Scholar] [CrossRef] [Green Version]
- Duan, A.M.; Xiao, Z.X. Does the climate warming hiatus exist over the Tibetan Plateau? Sci. Rep. 2015, 5, 9. [Google Scholar] [CrossRef]
- Cai, D.L.; You, Q.L.; Fraedrich, K.; Guan, Y.N. Spatiotemporal Temperature Variability over the Tibetan Plateau: Altitudinal Dependence Associated with the Global Warming Hiatus. J. Clim. 2017, 30, 969–984. [Google Scholar] [CrossRef]
- Chakraborty, A.; Saha, S.; Sachdeva, K.; Joshi, P.K. Vulnerability of forests in the Himalayan region to climate change impacts and anthropogenic disturbances: A systematic review. Reg. Environ. Change 2018, 18, 1783–1799. [Google Scholar] [CrossRef]
- Oorthuis, R.; Vaunat, J.; Hurlimann, M.; Lloret, A.; Moya, J.; Puig-Polo, C.; Fraccica, A. Slope Orientation and Vegetation Effects on Soil Thermo-Hydraulic Behavior. An Experimental Study. Sustainability 2021, 13, 14. [Google Scholar] [CrossRef]
- Legendre, P.; Mi, X.C.; Ren, H.B.; Ma, K.P.; Yu, M.J.; Sun, I.F.; He, F.L. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.N.; Yang, Q.P.; Zhou, D.Q.; Xu, W.G.; Gao, J.; Wang, Z. How evergreen and deciduous trees coexist during secondary forest succession: Insights into forest restoration mechanisms in Chinese subtropical forest. Glob. Ecol. Conserv. 2021, 25, e01418. [Google Scholar] [CrossRef]
- Qu, S.; Wang, L.C.; Lin, A.W.; Zhu, H.J.; Yuan, M.X. What drives the vegetation restoration in Yangtze River basin, China: Climate change or anthropogenic factors? Ecol. Indic. 2018, 90, 438–450. [Google Scholar] [CrossRef]
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Curtis, J.T.; McIntosh, R.P. The interrelations of certain analytic and synthetic phytosociological characters. Ecology 1950, 31, 434–455. [Google Scholar] [CrossRef]
- Kunming Institute of Botany, Chinese Academy of Sicence. Flora Yunnan; Science Press: Beijing, China, 2006; Volume 1–21. [Google Scholar]
- Wu, Z.Y. Flora of Tibet; Science Press: Beijing, China, 1987; Volume 1–5. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Guo, K.; Liu, C.C.; Xie, Z.Q.; Li, F.Y.; Franklin, S.B.; Lu, Z.J.; Ma, K.P. China Vegetation Classification: Concept, approach and applications. Phytocoenologia 2018, 48, 113–120. [Google Scholar] [CrossRef]
- Guo, K.; Fang, J.; Wang, G.; Tang, Z.; Xie, Z.; Shen, Z.; Wang, R.; Qiang, S.; Liang, C.; Da, L.; et al. A revised scheme of vegetation classification system of China. Chin. J. Plant Ecol. 2020, 44, 111–127. [Google Scholar] [CrossRef]
- Pakgohar, N.; Rad, J.E.; Gholami, G.; Alijanpour, A.; Roberts, D.W. A comparative study of hard clustering algorithms for vegetation data. J. Veg. Sci. 2021, 32, e13042. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 1 October 2021).
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K.; Studer, M.; Roudier, P.; Gonzalez, J.; Kozlowski, K.; Schubert, E.; et al. Cluster: Cluster Analysis Basics and Extensions. R Package Version 2.1.1. Available online: https://CRAN.R-project.org/package=cluster (accessed on 1 October 2021).
- De Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 October 2021).







| NMDS1 | NMDS2 | R2 | Pr (>r) | |
|---|---|---|---|---|
| Elevation | 0.92596 | 0.37763 | 0.8681 | 0.001 |
| Aspect | −0.87558 | −0.48307 | 0.0745 | 0.001 |
| Slope | −0.09324 | −0.99564 | 0.0287 | 0.073 |
| Bio12 | 0.94077 | −0.33905 | 0.0817 | 0.003 |
| Bio14 | −0.39677 | 0.91792 | 0.0315 | 0.058 |
| Bio15 | 0.0542 | −0.99853 | 0.0728 | 0.002 |
| Diff | Lwr | Upr | P adj | |
|---|---|---|---|---|
| Group 2—Group 1 | 34.86226 | −174.121 | 243.845 | 0.990756 |
| Group 3—Group 1 | 820.0123 | 647.5891 | 992.4354 | * |
| Group 4—Group 1 | 1677.859 | 1466.588 | 1889.13 | * |
| Group 5—Group 1 | 2718.129 | 2468.598 | 2967.66 | * |
| Group 3—Group 2 | 785.15 | 580.621 | 989.679 | * |
| Group 4—Group 2 | 1642.997 | 1404.8 | 1881.194 | * |
| Group 5—Group 2 | 2683.267 | 2410.561 | 2955.972 | * |
| Group 4—Group 3 | 857.8466 | 650.98 | 1064.713 | * |
| Group 5—Group 3 | 1898.117 | 1652.303 | 2143.93 | * |
| Group 5—Group 4 | 1040.27 | 765.8073 | 1314.733 | * |
| Cluster Group | Number of Sites | Species Richness per Group (Total) | Species Richness per Site (Mean) | Shannon Diversity Index, per Site (Mean) | Simpson Diversity Index, per Site (Mean) | Pielou Diversity Index, per Site (Mean) |
|---|---|---|---|---|---|---|
| group 1 | 53 | 618 | 42.79 a | 3.16 a | 0.91 a | 0.86 a |
| group 2 | 31 | 423 | 38.74 b | 3.08 b | 0.91 b | 0.85 b |
| group 3 | 60 | 823 | 46.43 c | 3.26 c | 0.92 c | 0.86 c |
| group 4 | 28 | 459 | 39.32 d | 3.03 d | 0.91 d | 0.83 d |
| group 5 | 18 | 185 | 7.02 e | 2.72 e | 0.89 e | 0.86 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.-P.; Wang, Z.; Jia, N.-X.; Dong, S.-Q.; Qu, X.-Y.; Qiao, X.-G.; Liu, C.-C.; Guo, K. Vegetation Classification and Distribution Patterns in the South Slope of Yarlung Zangbo Grand Canyon National Nature Reserve, Eastern Himalayas. Plants 2022, 11, 1194. https://doi.org/10.3390/plants11091194
Wu P-P, Wang Z, Jia N-X, Dong S-Q, Qu X-Y, Qiao X-G, Liu C-C, Guo K. Vegetation Classification and Distribution Patterns in the South Slope of Yarlung Zangbo Grand Canyon National Nature Reserve, Eastern Himalayas. Plants. 2022; 11(9):1194. https://doi.org/10.3390/plants11091194
Chicago/Turabian StyleWu, Po-Po, Zi Wang, Ning-Xia Jia, Shao-Qiong Dong, Xiao-Yun Qu, Xian-Guo Qiao, Chang-Cheng Liu, and Ke Guo. 2022. "Vegetation Classification and Distribution Patterns in the South Slope of Yarlung Zangbo Grand Canyon National Nature Reserve, Eastern Himalayas" Plants 11, no. 9: 1194. https://doi.org/10.3390/plants11091194
APA StyleWu, P.-P., Wang, Z., Jia, N.-X., Dong, S.-Q., Qu, X.-Y., Qiao, X.-G., Liu, C.-C., & Guo, K. (2022). Vegetation Classification and Distribution Patterns in the South Slope of Yarlung Zangbo Grand Canyon National Nature Reserve, Eastern Himalayas. Plants, 11(9), 1194. https://doi.org/10.3390/plants11091194