
Musa Germplasm A and B Genomic Composition Differentially Affects Their Susceptibility to Banana Bunchy Top Virus and Its Aphid Vector, Pentalonia nigronervosa

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Planting Material
2.2. Screening of Musa Genotypes for Banana Aphid Population Growth in the Screenhouse
2.3. Evaluation of Musa Genotypes against Banana Aphid and BBTD under Field Conditions
2.4. Data Analysis
3. Results
3.1. Banana Aphid Abundance on Musa Genotypes in the Screenhouse
3.2. Banana Aphid and BBTD Occurrence on Musa Genotypes under Field Conditions
Abundance of Banana Aphid in the Field
3.3. Time to First BBTD Symptoms
3.4. BBTD Incidence and Area under Disease Progress Curve (AUDPC)
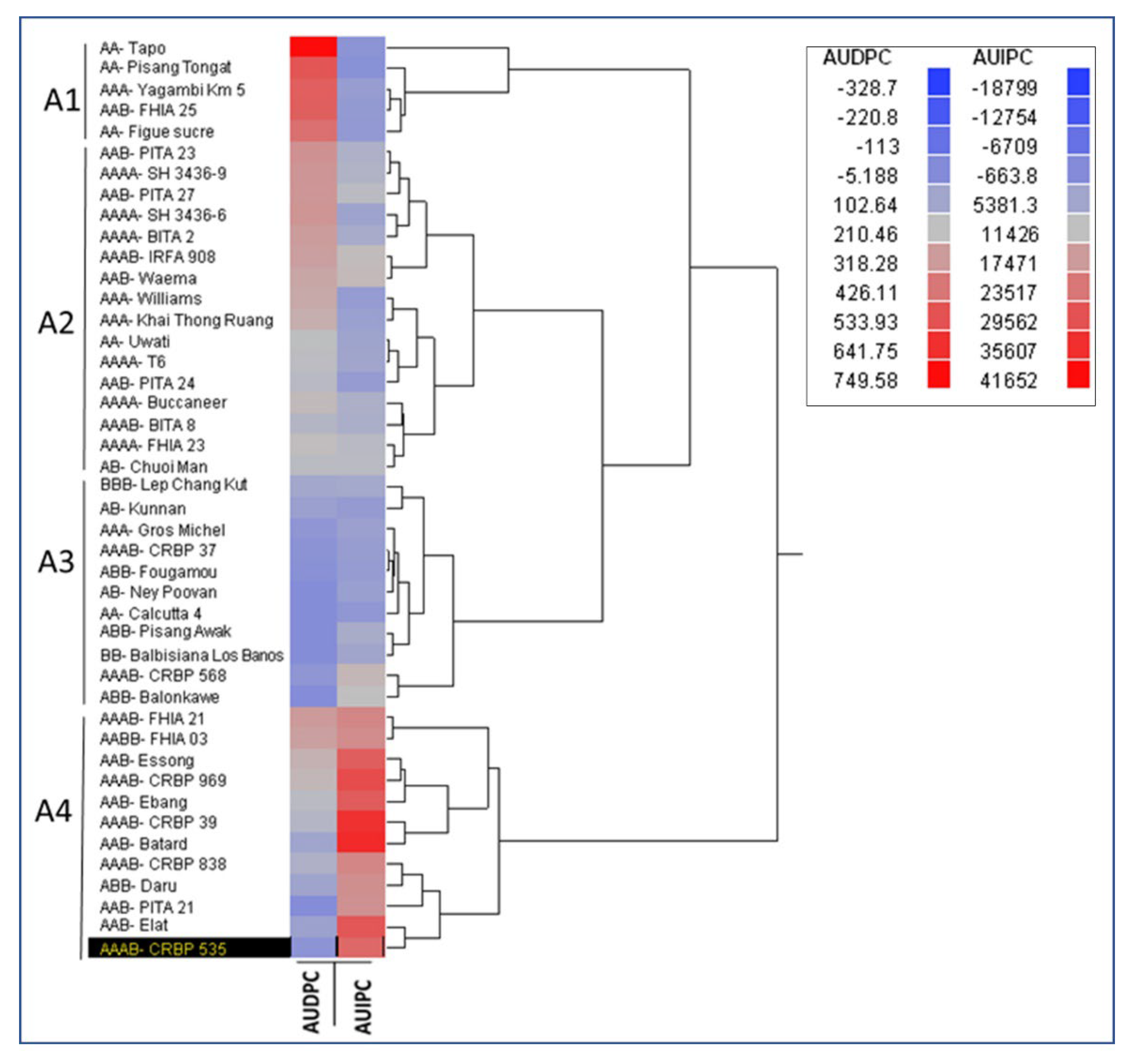
3.5. Classification of Musa Genotypes for the Response to Banana Aphid and BBTD in Field
3.6. Correlation for Aphid Performance in Screenhouse and Field
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Banana Market Review Preliminary Results 2020; FAO: Rome, Italy, 2021; 21, Available online: https://www.fao.org/3/cb5150en/cb5150en.pdf (accessed on 8 February 2022).
- Hooks, C.R.R.; Wright, M.G.; Kabasawa, D.S.; Manandhar, R.; Almeida, R.P.P. Effect of banana bunchy top virus infection on morphology and growth characteristics of banana. Ann. Appl. Biol. 2008, 153, 1–9. [Google Scholar] [CrossRef]
- Kumar, P.L.; Selvarajan, R.; Iskra-Caruana, M.L.; Chabannes, M.; Hanna, R. Biology, etiology, and control of virus diseases of banana and plantain. Adv. Vir. Res. 2015, 91, 229–269. [Google Scholar] [CrossRef]
- Dale, J.L. Banana bunchy top: An economically important tropical plant virus disease. Adv. Virus Res. 1987, 33, 301–325. [Google Scholar] [CrossRef]
- Hooks, C.R.R.; Manandhar, R.; Perez, E.P.; Wang, K.H.; Almeida, R.P.P. Comparative susceptibility of two banana cultivars to banana bunchy top virus under laboratory and field environments. J. Econ. Entomol. 2009, 102, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.L.; Hanna, R.; Alabi, O.J.; Soko, M.M.; Oben, T.T.; Vangu, G.H.P.; Naidu, R.A. Banana bunchy top virus in sub-Saharan Africa: Investigations on virus distribution and diversity. Virus Res. 2011, 159, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Qazi, J. Banana bunchy top virus and the bunchy top disease. J. Gen. Plant Pathol. 2016, 82, 2–11. [Google Scholar] [CrossRef]
- Eastop, V.F. A taxonomic study of australian aphidoidea (Homoptera). Aust. J. Zool. 1965, 14, 399–592. [Google Scholar] [CrossRef]
- Foottit, R.G.; Maw, H.E.L.; Pike, K.S.; Miller, R.H. The identity of Pentalonia nigronervosa Coquerel and P. caladii van der Goot (Hemiptera: Aphididae) based on molecular and morphometric analysis. Zootaxa 2010, 2358, 25–38. [Google Scholar] [CrossRef]
- Watanabe, S.; Greenwell, A.M.; Bressan, A. Localization, concentration, and transmission efficiency of banana bunchy top virus in four asexual lineages of Pentalonia aphids. Viruses 2013, 5, 758–776. [Google Scholar] [CrossRef] [Green Version]
- Hamim, I.; Green, J.C.; Borth, W.B.; Melzer, M.J.; Wang, Y.N.; Hu, J.S. First report of banana bunchy top virus in Heliconia spp. on Hawaii. Plant Dis. 2017, 101, 2153. [Google Scholar] [CrossRef]
- Rahayuniati, R.F.; Hartono, S.; Somowiyarjo, S.; Subandiyah, S.; Thomas, J.E. Characterization of banana bunchy top virus on Sumatra (Indonesia) wild banana. Biodivers. J. Biol. Divers. 2021, 22, 1243–1249. [Google Scholar] [CrossRef]
- Hu, J.S.; Wang, M.; Sether, D.; Xie, W.; Leonhardt, K.W. Use of polymerase chain reaction (PCR) to study transmission of banana bunchy top virus by the banana aphid (Pentalonia nigronervosa). Ann. Appl. Biol. 1996, 128, 55–64. [Google Scholar] [CrossRef]
- Anhalt, M.D.; Almeida, R.P.P. Effect of temperature, vector life stage, and plant access period on transmission of Banana bunchy top virus to banana. Phytopathology 2008, 98, 743–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magee, C.J.P. Investigation on the bunchy top disease of the banana. Bull. Counc. Sci. Ind. Res. Aust. 1927, 30, 64. [Google Scholar]
- Smith, M.C.; Holt, J.; Kenyon, L.; Foot, C. Quantitative epidemiology of Banana Bunchy Top Virus Disease and its control. Plant Pathol. 1998, 47, 177–187. [Google Scholar] [CrossRef]
- Allen, R.N. Epidemiological factors influencing the success of roguing for the control of bunchy top disease of bananas in New South Wales. Aust. J. Agric. Res. 1978, 29, 535–544. [Google Scholar] [CrossRef]
- Robson, J.D.; Wright, M.G.; Almeida, R.P.P. Effect of imidacloprid foliar treatment and banana leaf age on Pentalonia nigronervosa (Hemiptera, Aphididae) survival. N. Z. J. Crop Hortic. Sci. 2007, 35, 415–422. [Google Scholar] [CrossRef]
- Abiola, A.; Zandjanakou-Tachin, M.; Aoudji, K.N.A.; Avocevou-Ayisso, C.; Kumar, P.L. Adoption of roguing to contain banana bunchy top disease in south-east Bénin: Role of farmers’ knowledge and perception. Int. J. Fruit Sci. 2020, 20, 720–736. [Google Scholar] [CrossRef]
- Nkengla-Asi, L.; Eforuoku, F.; Olaosebikan, O.; Ladigbolu, T.A.; Amah, D.; Hanna, R.; Kumar, P.L. Gender roles in sourcing and sharing of banana planting material in communities with and without banana bunchy top disease in Nigeria. Sustainability 2021, 13, 3310. [Google Scholar] [CrossRef]
- Metcalf, R.L.; Luckmann, W.H. Introduction to Insect Pest Management; John Wiley & Sons, Inc.: New York, NY, USA, 1994; 672. [Google Scholar]
- Jones, R.A.C. Control of Plant Virus Diseases. Adv. Virus Res. 2006, 67, 205–244. [Google Scholar]
- Rodrigues, S.P.; Lindsey, G.G.; Bueno Fernandes, P.M. Biotechnological approaches for plant virus resistance: From general to the modern RNA silencing pathway. Braz. Arch. Biol. Technol. 2009, 52, 795–808. [Google Scholar] [CrossRef]
- Hill, J.H.; Whitham, S.A. Control of virus diseases in soybeans. Adv. Vir. Res. 2014, 90, 355–390. [Google Scholar] [CrossRef]
- Nicaise, V. Crop immunity against viruses: Outcomes and future challenges. Front. Plant Sci. 2014, 5, 660. [Google Scholar] [CrossRef] [PubMed]
- Schädler, M.; Brandl, R.; Kempel, A. Host plant genotype determines bottom-up effects in an aphid-parasitoid-predator system. Entomol. Exp. Appl. 2010, 135, 162–169. [Google Scholar] [CrossRef]
- Hapsari, L.; Masrum, A. Preliminary screening resistance of Musa germplasms for banana bunchy top disease in Purwodadi Botanic Garden, Pasuruan, East Java. Bul. Kebun Raya 2012, 15, 57–70. [Google Scholar]
- Espino, R.C.; Magnaye, L.V.; Johns, A.P.; Juanillo, C. Evaluation of Philippine banana cultivars for resistance to bunchy top and fusarium wilt. In Proceedings of the International Symposium on Recent Developments in Banana Cultivation Technology, Pingtung, Taiwan, 14–18 December 1992; Valmayor, R.V., Hwang, S.C., Ploetz, R., Lee, R.C., Roa, N.V., Eds.; INIBAP/ASPNET: Los Baños, Philippines, 1993; pp. 89–102. [Google Scholar]
- Ngatat, S.; Hanna, R.; Kumar, P.L.; Gray, S.M.; Cilia, M.; Ghogomu, R.T.; Fontem, D.A. Relative susceptibility of Musa genotypes to banana bunchy top disease in Cameroon and implication for disease management. Crop Prot. 2017, 101, 116–122. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on World’s Crops: An Identification and Information Guide, 2nd ed.; Wiley: Chichester, UK, 2000. [Google Scholar]
- Robson, J.D.; Wright, M.G.; Almeida, R.P.P. Within-plant distribution and binomial sampling of Pentalonia nigronervosa (Hemiptera: Aphididae) on banana. J. Econ. Entomol. 2006, 99, 2185–2190. [Google Scholar] [CrossRef] [Green Version]
- Hanna, R.; Kumar, P.L. Banana aphid and banana bunchy top virus in Africa: Distribution and development of management options. In Proceedings of the Annual Meeting of the Entomological Society of America, Indianapolis, IN, USA, 13–16 December 2009. [Google Scholar]
- Shaner, G.; Finney, R. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in knox wheat. Phytopathology 1977, 77, 1051. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591. [Google Scholar] [CrossRef]
- Wang, Z.; Miao, H.; Liu, J.; Xu, B.; Yao, X.; Xu, C.; Zhao, S.; Fang, X.; Jia, C.; Wang, J.; et al. Musa balbisiana genome reveals subgenome evolution and functional divergence. Nat. Plants 2019, 5, 810–882. [Google Scholar] [CrossRef] [Green Version]
- Mobambo, K.N.; Pasberg-Gauhl; Gauhk, F.; Zuofa., K. Criblage précoce pour la résistance à la maladie des raies noires ou cercosporiose noire en conditions d’inoculation naturelle (Early screening for black leaf streak/black Sigatoka disease resistance under natural inoculation conditions). InfoMusa 1994, 3, 14–16. [Google Scholar]
- Karley, A.J.; Douglas, A.E.; Parker, W.E. Amino acid composition and nutritional quality of potato leaf phloem sap for aphids. J. Exp. Biol. 2002, 205, 3009–3018. [Google Scholar] [CrossRef] [PubMed]
- Powell, G.; Tosh, C.R.; Hardie, J. Host plant selection by aphids: Behavioral, evolutionary, and applied perspectives. Annu. Rev. Entomol. 2006, 51, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Meihls, L.N.; Handrick, V.; Glauser, G.; Barbier, H.; Kaur, H.; Haribal, M.M.; Lipka, A.E.; Gershenzon, J.; Buckler, E.S.; Erb, M.; et al. Natural variation in maize aphid resistance is associated with 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one glucoside methyltransferase activity. Plant Cell 2013, 25, 2341–2355. [Google Scholar] [CrossRef] [Green Version]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Döring, T.F.; Chittka, L. Visual ecology of aphids—A critical review on the role of colours in host finding. Arthropod-Plant. Interact. 2007, 1, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Murhububa, I.S.; Tougeron, K.; Bragard, C.; Fauconnier, M.-L.; Basengere, E.B.; Masamba, J.W.; Hance, T. Banana tree infected with banana bunchy top virus attracts Pentalonia nigronervosa aphids through increased volatile organic compounds emission. J. Chem. Ecol. 2021, 47, 755–767. [Google Scholar] [CrossRef]
- Pickett, J.A.; Birkett, M.A.; Bruce, T.J.A.; Chamberlain, K.; Gordon-Weeks, R.; Matthes, M.C.; Napier, J.A.; Smart, L.E.; Woodcock, C.M. Developments in aspects of ecological phytochemistry: The role of cis-jasmone in inducible defence systems in plants. Phytochemistry 2007, 68, 2937–2945. [Google Scholar] [CrossRef]
- Carlier, J.; De Waele, D.; Escalant, J.-V.; Vezina, A.; Picq., C. Global Evaluation of Musa Germplasm for Resistance to Fusarium Wilt, Mycosphaerella Leaf Spot Diseases and Nematodes: In-dept Evaluation; INIBAP Technical Guidelines No. 6; INIBAP: Montpellier, France, 2002; 64, Available online: https://hdl.handle.net/10568/105201 (accessed on 1 March 2022)ISBN 978-2-910810-52-8.
- Fogain, R.; Price, N.S. Varietal screening of some Musa cultivars for susceptibility to the banana weevil, Cosmopolites sordidus (Coleoptera: Curculionidae). Fruits 1994, 49, 247–251. [Google Scholar]
- Jones, D.R. (Ed.) Diseases of Banana, Abacá and Enset; CABI: Wallingford, UK, 2000; ISBN 0-85199-355-9. [Google Scholar]
- Tripathi, L.; Odipio, J.; Tripathi, J.N.; Tusiime, G. A rapid technique for screening banana cultivars for resistance to Xanthomonas wilt. Eur. J. Plant Pathol. 2008, 121, 9–19. [Google Scholar] [CrossRef]
- Ravi, I.; Uma, S.; Vaganan, M.M.; Mustaffa, M.M. Phenotyping bananas for drought resistance. Front. Physiol. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenkouano, A.; Swennen, R.L. Plantain and banana: Progress in breeding and delivering improved plantain and banana to African farmers. Chron. Hortic. 2004, 44, 9–15. [Google Scholar]



{kind=link}
{kind=link}
| Ploidy Level | Genomic Group | Genotype | Source |
|---|---|---|---|
| Diploid | AA | Calcutta 4 | ITC |
| Figue Sucrée | ITC | ||
| Pisang Tongat | ITC | ||
| Uwati | ITC | ||
| Tapo | ITC | ||
| AB | Ney Poovan | ITC | |
| Auko * | ITC | ||
| Vunapope * | ITC | ||
| Kunnan | ITC | ||
| Chuoi Man | ITC | ||
| BB | Balbisiana Los Banos | ITC | |
| Triploid | AAA | Gros Michel | ITC |
| Khai Thong Ruang | ITC | ||
| Yangambi Km5 | ITC | ||
| Williams | IITA | ||
| AAB | Batard # | IITA | |
| Ebang | IITA | ||
| Elat # | IITA | ||
| Essong | IITA | ||
| FHIA 25 | IITA | ||
| PITA 21 | IITA | ||
| PITA 23 | IITA | ||
| PITA 24 # | IITA | ||
| PITA 27 # | IITA | ||
| Waema | ITC | ||
| ABB | Ice cream * | ITC | |
| Pisang Awak | ITC | ||
| Balonkawe | ITC | ||
| Daru | ITC | ||
| Fougamou # | IITA | ||
| BBB | Lep Chang Kut | ITC | |
| Tetraploid | AAAA | T6 | ITC |
| Buccaneer | ITC | ||
| SH3436-9 | ITC | ||
| SH3436-6 | ITC | ||
| FHIA 23 | IITA | ||
| BITA-2 | ITC | ||
| AAAB | BITA 8 # | IITA | |
| IRFA 908 | ITC | ||
| FHIA-21 | ITC | ||
| CRBP 37 | ITC | ||
| CRBP 39 | ITC | ||
| CRBP 568 # | CARBAP | ||
| CRBP 535 # | CARBAP | ||
| CRBP 838 # | CARBAP | ||
| CRBP 969 # | CARBAP | ||
| AABB | FHIA-03 | ITC | |
| ABBT | Yawa 2 * | ITC |
| Genomic Group | Genotype | Apterous Aphid Abundance | Alate Aphid Abundance |
|---|---|---|---|
| AA | Calcutta 4 | 9.7 ± 4.6 a | 0 a |
| AA | Figue sucrée | 120.9 ± 35.5 a | 1.3 ± 0.2 a |
| AA | Pisang Tongat | 73.2 ± 16.6 a | 2.1 ± 1.0 a |
| AA | Tapo | 58.4 ± 20.6 a | 4.1 ± 3.3 a |
| AA | Uwati | 82.8 ± 28.9 a | 1.0 ± 0.6 a |
| AB | Auko | 118.3 ± 48.4 a | 0 a |
| AB | Chuoi Man | 40.3 ± 19.9 a | 0.2 ± 0.2 a |
| AB | Kunnan | 136.7 ± 66.6 a | 2.9 ± 1.4 a |
| AB | Ney Poovan | 17.9 ± 7.5 a | 0.1 ± 0.0 a |
| AB | Vunapope | 67.4 ± 23.8 a | 2.8 ± 2.1 a |
| BB | Balbisiana | 111.4 ± 18.4 a | 0.2 ± 0.2 a |
| Chisq | 17.66 | 13.9 | |
| df | 10 | 10 | |
| p | 0.06 | 0.17 |
| Genomic Group | Genotype | Apterous Aphid Abundance | Alate Aphid Abundance |
|---|---|---|---|
| AAA | Gros Michel | 115.6 ± 55.8 ab | 2.9 ± 2.8 ab |
| AAA | Khai Tang Wang | 86.7 ± 34.8 ab | 1.0 ± 0.8 ab |
| AAA | William | 79.5 ± 18.3 ab | 2.7 ± 1.4 ab |
| AAA | Yangambi Km5 | 80.3 ± 9.3 ab | 0 b |
| AAB | Ebang | 366.1 ± 24.7 ab | 34.7 ±11.7 a |
| AAB | Essong | 262.3 ± 49.3 ab | 22.0 ± 8.4 a |
| AAB | FHIA 25 | 138.9 ± 43.9 ab | 2.1 ± 1.2 ab |
| AAB | PITA 23 | 96.8 ± 13.6 ab | 2.6 ± 0.6 ab |
| AAB | Waema | 521.0 ± 126.0 a | 31.8 ± 8.1 a |
| ABB | Daru | 114.2 ± 43.3 ab | 6.5 ± 5.2 ab |
| ABB | Fougamou | 124.9 ± 56.1 ab | 3.4 ± 2.0 ab |
| ABB | Ice Cream | 63.5 ± 5.9 b | 0.5 ± 0.3 ab |
| ABB | Pisang Awak | 117.9 ±25.7 ab | 6.8 ± 3.0 ab |
| ABB | Balonkawe | 97.7 ± 44.1 ab | 0.3 ± 0.3 b |
| BBB | Lep Chang Kut | 64.6 ± 22.0 ab | 0.1 ± 0.0 b |
| Chisq | 24.38 | 32.22 | |
| df | 14 | 14 | |
| p | 0.04 | 0.004 |
| Genomic Group | Genotype | Apterous Aphid Abundance | Alate Aphid Abundance |
|---|---|---|---|
| AAAA | BITA 2 | 190.4 ± 34.6 ab | 8.0 ± 4.4 ab |
| AAAA | Buccaneer | 153.0 ± 35.6 ab | 2.5 ± 0.5 ab |
| AAAA | FHIA 23 | 101.6 ± 38.9 ab | 1.3 ± 1.0 b |
| AAAA | SH 3436-6 | 117.0 ± 14.6 ab | 0.3 ± 0.1 b |
| AAAA | SH 3436-9 | 90.0 ± 34.8 ab | 0.6 ± 0.4 b |
| AAAA | T6 | 50.7 ± 11.0 b | 0.3 ± 0.2 b |
| AAAB | CRBP 37 | 82.1 ± 7.8 b | 0.6 ± 0.4 b |
| AAAB | CRBP 39 | 115.5 ± 18.2 ab | 3.9 ± 1.8 ab |
| AAAB | FHIA 21 | 167.2 ± 55.1 ab | 5.6 ± 2.7 ab |
| AAAB | IRFA 908 | 153.1 ± 25.2 ab | 7.4 ± 3.9 ab |
| AABB | FHIA 03 | 327.0 ± 40.1 a | 27.3 ± 6.6 a |
| ABBT | Yawa 2 | 128.8 ± 19.4 ab | 5.6 ± 2.5 ab |
| Chisq | 21.49 | 25.19 | |
| df | 11 | 11 | |
| p | 0.03 | 0.009 |
| Genomic Group | Genotypes | Apterous Aphid | Alate Aphid |
|---|---|---|---|
| Genotypes with high aphid densities | |||
| AAB | Waema | 395.6 ± 20.8 a | 24.9 ± 7.1 a |
| AAB | Ebang | 366.1 ± 24.7 ab | 34.7 ± 11.7 a |
| AABB | FHIA 03 | 327.0 ± 40.1 ab | 27.3 ± 6.6 a |
| AAAA | BITA 2 | 190.4 ± 34.6 abc | 8.0 ± 4.4 ab |
| AB | Kunnan | 136.7 ± 66.7 bcd | 2.9 ± 1.4 abc |
| AA | Figue sucrée | 120.9 ± 35.5 abcd | 1.3 ± 0.2 abc |
| Genotypes with low aphid densities | |||
| AAAB | CRBP 37 | 82.1 ± 7.8 bcde | 0.6 ± 0.4 bcd |
| BBB | Lep Chang Kut | 64.6 ± 22.0 cde | 0.1 ± 0.0 cd |
| ABB | Ice Cream | 63.5 ± 5.9 cde | 0.5 ± 0.3 bcd |
| AAAA | T6 | 50.7 ± 11.0 cde | 0.3 ± 0.2 bcd |
| AB | Ney Poovan | 17.9 ± 7.6 de | 0.1 ± 0.0 cd |
| AA | Calcutta 4 | 9.7 ± 4.6 e | 0 d |
| Chisq | 30.0 | 30.1 | |
| Df | 11 | 11 | |
| p | 0.002 | 0.002 | |
| Genomic Group | Genotype | Total Plants Planted | Month to First Symptoms | Apterous Aphid Abundance | Alate Aphid Abundance | BBTD Incidence | AUDPC | Total Tested for BBTV | BBTV Incidence (%) |
|---|---|---|---|---|---|---|---|---|---|
| AA | Calcutta 4 | 9 | NS | 1.4 ± 0.4 bcd | 0.2 ± 0.1 a | 0 c | 0 c | 7 | 0 |
| AA | Figue sucree | 14 | 3 | 2.1 ± 0.6 bcd | 0.2 ± 0.1 a | 56.7 ± 12.0 abc | 449 ± 110 ab | NA | - |
| AA | Pisang Tongat | 13 | 3 | 0.4 ± 0.1 d | 0.2 ± 0.1 a | 61.7 ± 7.3 ab | 527 ± 70.4 ab | 3 | 0 |
| AA | Tapo | 9 | 3 | 1.4 ± 1.1 cd | 0.2 ± 0.1 a | 100 ± 0.0 a | 869 ± 37.5 a | NA | - |
| AA | Uwati | 12 | 7 | 3.9 ± 0.9 abc | 0.1 ± 0.0 a | 33.3 ± 17.6 abc | 202 ± 117 bc | 24 | 0 |
| AB | Chuoi Man | 14 | 2 | 9.2 ± 1.6 a | 0.4 ± 0.1 a | 21.7 ± 11.7 bc | 187 ± 107 bc | 47 | 0 |
| AB | Kunnan | 15 | 18 | 2.5 ± 0.6 abc | 0.1 ± 0.0 a | 33.3 ± 6.7 abc | 80.6 ± 64.4 bc | 45 | 15.6 |
| AB | Ney Poovan | 15 | 36 | 2.5 ± 0.9 abc | 0.3 ± 0.1 a | 6.7 ± 6.7 c | 1.1 ± 1.1 c | 99 | 0 |
| BB | Balbisiana Los Banos | 8 | NS | 5.2 ± 1.9 ab | 0.1 ± 0.1 a | 0 c | 0 c | 7 | 0 |
| Chisq | 15.84 | 12.1 | 19.55 | 20.15 | - | - | |||
| Df | 8 | 8 | 8 | 8 | - | - | |||
| p | 0.04 | 0.15 | 0.01 | 0.009 | - | - | |||
| Genomic Group | Genotypes | Total Plants Planted | Month to First Infection | Apterous Aphid Abundance | Alate Aphid Abundance | BBTD Incidence (%) | AUDPC | Samples Tested for BBTV | BBTV Incidence (%) |
|---|---|---|---|---|---|---|---|---|---|
| AAA | Gros Michel | 10 | 21 | 5.4 ± 2.5 bcde | 0.22 ± 0.05 abc | 8.3 ± 8.3 a | 38.8 ± 38.8 ab | 23 | 4.2 |
| AAA | Khai Thong Ruang | 15 | 6 | 3.9 ± 0.9 bcde | 0.19 ± 0.05 bc | 40.0 ± 11.5 a | 261.8 ± 84.8 ab | 76 | 2.6 |
| AAA | Williams | 13 | 3 | 2.4 ± 0.5 de | 0.10 ± 0.03 c | 60.0 ± 10.0 a | 278.2 ± 108.0 ab | 6 | 16.7 |
| AAA | Yagambi Km5 | 7 | 11 | 2.4 ± 1.3 de | 0.05 ± 0.03 c | 55.0 ± 5.0 a | 498.5 ± 51.8 a | 12 | 0 |
| AAB | Batard | 15 | 6 | 29.2 ± 6.7 a | 0.44 ± 0.05 ab | 20.0 ± 0.0 a | 98.5 ± 51.3 ab | 36 | 0 |
| AAB | Ebang | 15 | 16 | 21.4 ± 8.6 abc | 0.45 ± 0.12 ab | 40.0 ± 11.5 a | 186.6 ± 42.5 ab | 21 | 0 |
| AAB | Elat | 14 | 4 | 20.7 ± 3.8 ab | 0.37 ± 0.12 abc | 26.7 ± 17.6 a | 83.6 ± 76.4 ab | 32 | 0 |
| AAB | Essong | 15 | 4 | 23.5 ± 9.6 abcd | 0.55 ± 0.11 a | 33.3 ± 17.6 a | 253.1 ± 127.1 ab | 15 | 0 |
| AAB | FHIA 25 | 15 | 4 | 2.0 ± 0.6 e | 0.11 ± 0.04 c | 66.7 ±17.6 a | 497.1 ± 128.7 a | 10 | 0 |
| AAB | PITA 21 | 14 | NS | 19.9 ± 8.0 abcd | 0.34 ± 0.07 abc | 0 a | 0 b | 30 | 0 |
| AAB | PITA 23 | 15 | 3 | 5.8 ± 1.5 abcde | 0.31 ± 0.09 abc | 53.3 ± 24.0 a | 348.4 ± 244.6 ab | 22 | 4.8 |
| AAB | PITA 24 | 15 | 10 | 2.4 ± 0.8 e | 0.15 ± 0.04 bc | 53.3 ± 17.6 a | 183.0 ± 89.9 ab | 23 | 4.2 |
| AAB | PITA 27 | 12 | 7 | 9.5 ± 1.3 abcde | 0.27 ± 0.06 abc | 50.0 ± 14.4 a | 332.6 ± 69.4 ab | 16 | 0 |
| AAB | Waema | 15 | 2 | 9.3 ± 2.6 abcde | 0.28 ± 0.08 abc | 40.0 ± 11.5 a | 287.9 ± 53.7 ab | 44 | 0 |
| ABB | Daru | 15 | 4 | 13.9 ± 1.3 abcd | 0.32 ± 0.07 abc | 13.3 ± 6.7 a | 93.8 ± 56.3 ab | 48 | 4.2 |
| ABB | Fougamou | 11 | 36 | 3.3 ± 1.5 cde | 0.22 ± 0.06 abc | 8.3 ± 8.3 a | 19.2 ± 19.2 ab | 47 | 0 |
| ABB | Pisang Awak | 13 | 28 | 6.6 ± 1.7 abcde | 0.23 ± 0.06 abc | 6.7 ± 6.7 a | 1.1 ± 1.1 ab | 81 | 4.8 |
| ABB | Balonkawe | 10 | NS | 10.9 ± 3.6 abcde | 0.5 ± 0.2 abc | 0 a | 0 a | 9 | 0 |
| BBB | Lep Chang Kut | 7 | 15 | 6.2 ± 2.5 abcde | 0.01 ±0.01 c | 12.5 ± 12.5 a | 106.1 ± 106.1 ab | 3 | 0 |
| Chisq | 40.47 | 39.95 | 32.22 | 34.92 | - | - | |||
| Df | - | 18 | 18 | 18 | 18 | - | - | ||
| p | - | 0.002 | 0.002 | 0.02 | 0.01 | - | - | ||
| Genomic Group | Genotypes | Total Plants Planted | Month to First Infection | Apterous Aphid Abundance | Alate Aphid Abundance | BBTD Incidence | AUDPC | Total Samples Tested for BBTV | BBTV Incidence (%) |
|---|---|---|---|---|---|---|---|---|---|
| AAAA | BITA2 | 9 | 14 | 4.9 ± 1.1 bc | 0.3 ± 0. 1 a | 55.6 ± 29.4 a | 317.6 ± 191.8 a | 6 | 0 |
| AAAA | Buccaneer | 14 | 3 | 7.2 ± 1.5 abc | 0.1 ± 0.0 a | 35.0 ± 5.0 a | 228.0 ± 46.5 a | 51 | 0 |
| AAAA | FHI A23 | 15 | 4 | 6.5 ± 2.5 abc | 0.3 ± 0.1 a | 33.3 ± 6.7 a | 217.3 ± 51.0 a | 25 | 0 |
| AAAA | SH 3436-6 | 13 | 4 | 3.7 ± 0.7 c | 0.3 ± 0.1 a | 48.3 ± 15.9 a | 338.1 ± 136 a | 15 | 17.8 |
| AAAA | SH 3436-9 | 14 | 3 | 5.6 ± 2.0 bc | 0.2 ± 0.1 a | 50.0 ± 5.8 a | 335.6 ± 73.7 a | 11 | 8.3 |
| AAAA | T6 | 14 | 3 | 3.0 ± 1.0 c | 0.1 ± 0.1 a | 33.3 ± 24.0 a | 197 ± 144 a | 9 | 0 |
| AAAB | BITA 8 | 12 | 7 | 6.6 ± 1.3 abc | 0.4 ± 0.1 a | 25.0 ± 14.4 a | 169 ± 87.0 a | 17 | 0 |
| AAAB | CRBP 37 | 15 | 26 | 1.97 ± 0.6 c | 0.1 ± 0.1 a | 6.7 ± 6.7 a | 23.1 ± 23.1 a | 19 | 0 |
| AAAB | CRBP 39 | 15 | 11 | 24.0 ± 5.5 a | 0.3 ± 0.1 a | 33.3 ± 6.7 a | 168 ± 75.7 a | 14 | 6.7 |
| AAAB | CRBP 535 | 14 | 28 | 21.0 ± 4.5 a | 0.5 ± 0.1 a | 8.3 ± 8.3 a | 28.9 ± 28.9 a | 25 | 0 |
| AAAB | CRBP 568 | 15 | 18 | 9.2 ± 2.0 abc | 0.2 ± 0.1 a | 6.7 ± 6.7 a | 37.8 ± 37.8 a | 24 | 0 |
| AAAB | CRBP 838 | 14 | 13 | 17.1 ± 4.1 ab | 0.3 ± 0.1 a | 25.0 ± 25.0 a | 146 ± 146 a | 35 | 2.6 |
| AAAB | CRBP 969 | 15 | 4 | 24.9 ± 6.5 abc | 0.3 ± 0.1 a | 33.3 ± 17.6 a | 237 ± 119 a | 21 | 4.8 |
| AAAB | FHIA 21 | 15 | 4 | 15.0 ± 3.2 abc | 0.4 ± 0.1 a | 53.3 ± 6.7 a | 323 ± 38.5 a | 9 | 0 |
| AAAB | IRFA 908 | 15 | 4 | 7.9 ± 2.4 abc | 0.7 ± 0.4 a | 66.7 ± 13.3 a | 308 ± 84.9 a | 30 | 0 |
| AABB | FHIA 03 | 7 | 3 | 14.5 ± 3.2 abc | 0.6 ± 0.1 a | 62.5 ± 37.5 a | 303 ± 46.8 a | 7 | 0 |
| Df | - | 16 | 16 | 16 | 16 | - | - | ||
| p | 0.003 | 0.18 | 0.14 | 0.16 | - | - |
| Genomic Groups | Genotypes | Apterous Aphids | Alate Aphids |
|---|---|---|---|
| Genotypes with high aphid densities | |||
| AAB | Batard | 29.2 ± 6.7 a | 0.4 ± 0.0 a |
| AAAB | CRBP 969 | 24.9 ± 10.8 ab | 0.3 ± 0.1 ab |
| AAAB | CRBP 39 | 24.0 ± 4.2 ab | 0.3 ± 0.1 ab |
| AAB | Essong | 23.5 ± 9.6 ab | 0.5 ± 0.0 ab |
| AB | Chuoi Man | 9.2 ± 2.2 abc | 0.4 ± 0.1 ab |
| BB | Balbisiana Los Banos | 5.2 ± 3.0 abcd | 0.1 ±0.0 b |
| Genotypes with low aphid densities | |||
| AAAA | T6 | 3.0 ± 1.0 bcd | 0.1 ± 0.1 ab |
| AAA | Yagambi Km5 | 2.4 ± 1.4 bcd | 0.0 ± 0.0 b |
| AAB | FHIA 25 | 2.0 ± 0.3 cd | 0.1 ± 0.0 b |
| AAAB | CRBP 37 | 2.0 ± 0.8 cd | 0.1 ± 0.1 b |
| AA | Tapo | 1.2 ± 0.9 cd | 0.1 ± 0.1 b |
| AA | Pisang Tongat | 0.4 ± 0.2 d | 0.2 ± 0.1 ab |
| Chisq | 27.7 | 23.5 | |
| Df | 11 | 11 | |
| p | 0.004 | 0.02 | |
| Genomic Group | Genotypes | Total Plants Planted | BBTD Incidence | AUDPC |
|---|---|---|---|---|
| AA | Tapo | 9 | 100 ± 0.0 a | 869 ± 37.5 a |
| AAB | FHIA 25 | 15 | 66.7 ± 17.6 a | 497 ± 128.7 ab |
| AAAB | IRFA 908 | 15 | 66.7 ± 13.3 a | 308 ± 84.9 bc |
| AABB | FHIA 03 | 7 | 62.5 ± 37.5 ab | 303 ± 46.8 abc |
| AA | Pisang Tongat | 13 | 61.7 ± 7.3 ab | 527 ± 70.4 ab |
| AAA | Williams | 13 | 60.0 ± 10.0 ab | 278 ± 108 bc |
| AAAB | CRBP 568 | 15 | 6.7 ± 6.7 bc | 37.8 ± 37.8 cd |
| AAAB | CRBP 37 | 15 | 6.7 ± 6.7 bc | 23.1 ± 23.1 cd |
| AA | Calcutta 4 | 9 | 0 bc | 0 d |
| BB | Balbisiana Los Banos | 8 | 0 bc | 0 d |
| ABB | Bolankawe | 10 | 0 bc | 0 d |
| AAB | PITA 21 | 14 | 0 bc | 0 d |
| Chisq | 26.9 | 28.5 | ||
| Df | 11 | 11 | ||
| p | 0.005 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngatat, S.; Hanna, R.; Lienou, J.; Ghogomu, R.T.; Nguidang, S.P.K.; Enoh, A.C.; Ndemba, B.; Korie, S.; Fotso Kuate, A.; Nanga Nanga, S.; et al. Musa Germplasm A and B Genomic Composition Differentially Affects Their Susceptibility to Banana Bunchy Top Virus and Its Aphid Vector, Pentalonia nigronervosa. Plants 2022, 11, 1206. https://doi.org/10.3390/plants11091206
Ngatat S, Hanna R, Lienou J, Ghogomu RT, Nguidang SPK, Enoh AC, Ndemba B, Korie S, Fotso Kuate A, Nanga Nanga S, et al. Musa Germplasm A and B Genomic Composition Differentially Affects Their Susceptibility to Banana Bunchy Top Virus and Its Aphid Vector, Pentalonia nigronervosa. Plants. 2022; 11(9):1206. https://doi.org/10.3390/plants11091206
Chicago/Turabian StyleNgatat, Sergine, Rachid Hanna, Jules Lienou, Richard T. Ghogomu, Sidonie Prisca K. Nguidang, Aime C. Enoh, Bertrand Ndemba, Sam Korie, Apollin Fotso Kuate, Samuel Nanga Nanga, and et al. 2022. "Musa Germplasm A and B Genomic Composition Differentially Affects Their Susceptibility to Banana Bunchy Top Virus and Its Aphid Vector, Pentalonia nigronervosa" Plants 11, no. 9: 1206. https://doi.org/10.3390/plants11091206
APA StyleNgatat, S., Hanna, R., Lienou, J., Ghogomu, R. T., Nguidang, S. P. K., Enoh, A. C., Ndemba, B., Korie, S., Fotso Kuate, A., Nanga Nanga, S., Fiaboe, K. K. M., & Kumar, P. L. (2022). Musa Germplasm A and B Genomic Composition Differentially Affects Their Susceptibility to Banana Bunchy Top Virus and Its Aphid Vector, Pentalonia nigronervosa. Plants, 11(9), 1206. https://doi.org/10.3390/plants11091206