
Accelerated Genetic Gains in Early-Maturing Maize Hybrids following Three Periods of Genetic Enhancement for Grain Yield under Low and High Soil-Nitrogen Environments



Abstract
:1. Introduction
2. Results
2.1. Analyses of Variance and Genotype by Environment Interaction of Grain Yield and Other Agronomic Traits
2.2. Repeatability Estimates of Grain Yield and Other Agronomic Traits
2.3. Genetic Improvements in Grain Yield and Other Measured Traits
2.4. Correlation of Grain Yield and Other Agronomic Traits
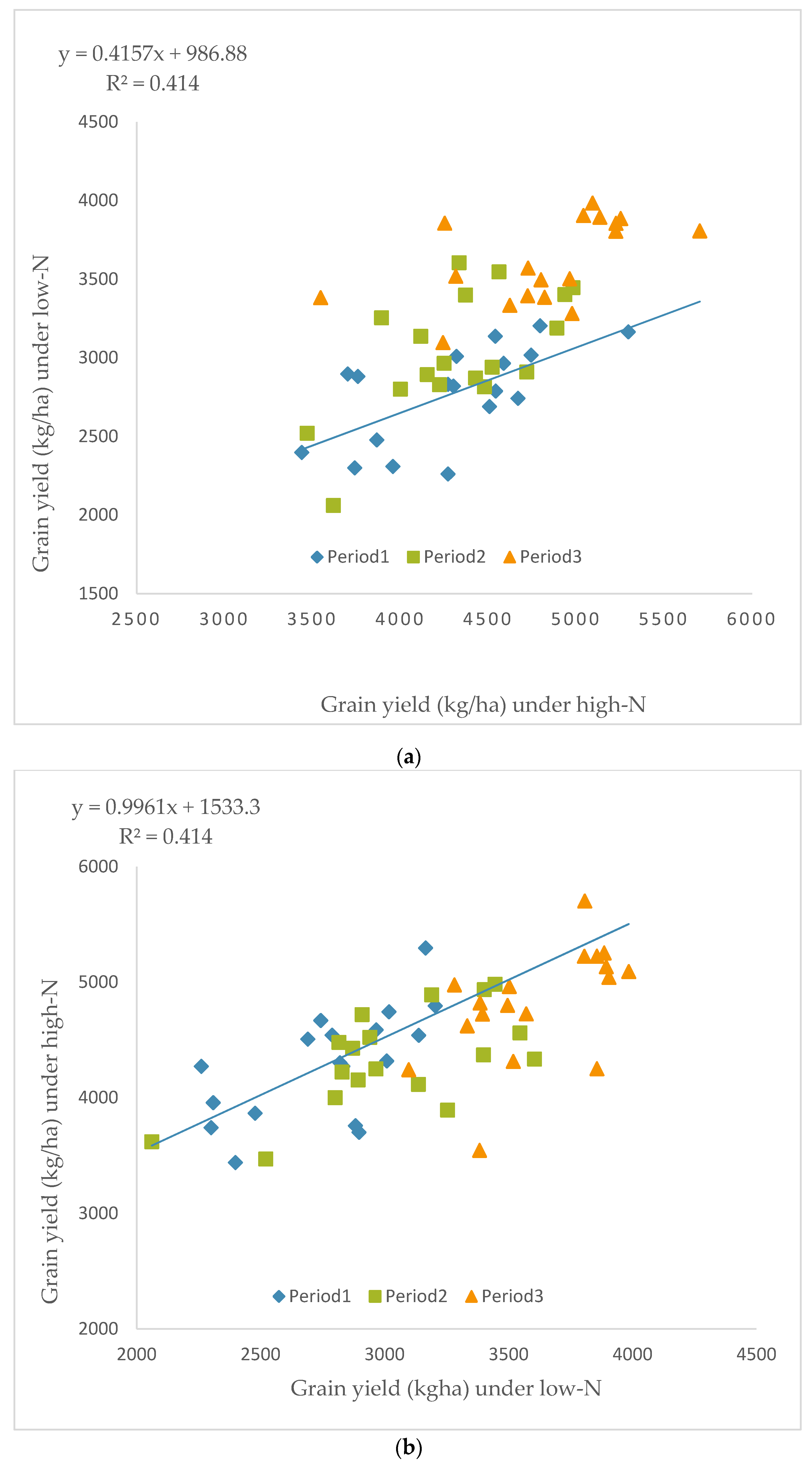
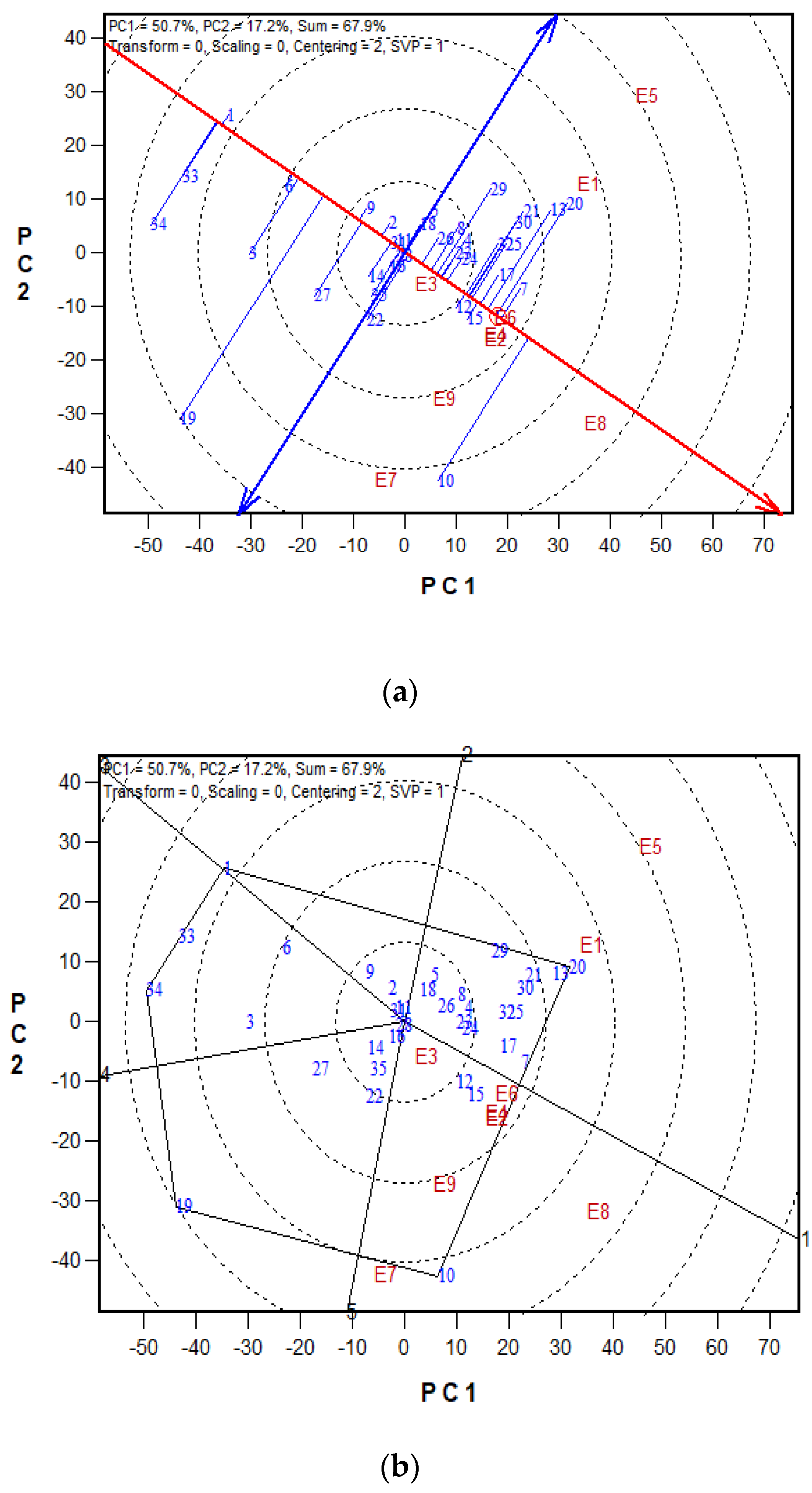
2.5. Grain Yield Performance and Stability of Maize Hybrid in Low- and High N Environments
3. Discussion
4. Materials and Methods
4.1. Development of Low NTolerant, Early-Maturing Inbreds and Hybrids for the Present Study
4.2. Field Evaluation
4.3. Data Collection
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Medium-Term Outlook for Raw Materials, Horticulture and Tropical; Food and Agriculture Organization Pub: Rome, Italy, 2016. [Google Scholar]
- Cairns, J.E.; Hellin, J.; Sonder, K.; Araus, J.L.; MacRobert, J.F.; Thierfelder, C.; Prasanna, B.M. Adapting maize production to climate change in sub-Saharan Africa. Food Sec. 2013, 5, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Badu-Apraku, B.; Samuel, A.; Agre, P.; Melaku, G.; Robert, A. Identification of QTLs Controlling Resistance/Tolerance to Striga hermonthica in an Extra-Early Maturing Yellow Maize Population. Agronomy 2020, 10, 1168. [Google Scholar] [CrossRef]
- Cechin, I.; Press, M.C. The influence of nitrogen on growth and photosynthesis of sorghum infected with Striga hermonthica from different provenances. Weed Res. 1993, 3, 289–298. [Google Scholar] [CrossRef]
- Kim, S.K.; Adetimirin, V.O. Responses of tolerant and susceptible maize varieties to timing and rate of nitrogen under Striga hermonthica infestation. J. Agron. 1997, 89, 38–44. [Google Scholar] [CrossRef]
- Banziger, M.; Edmeades, G.O.; Lafitte, H.R. Selection for drought tolerance increases maize yields across a range of nitrogen levels. Crop Sci. 1999, 39, 1035–1040. [Google Scholar] [CrossRef]
- Mccown, R.L.; Keating, B.A.; Probert, M.E.; Jones, R.K. Strategies for sustainable crop production in semi-arid Africa. Outlook Agric. 1992, 21, 21–31. [Google Scholar] [CrossRef]
- Wolfe, D.W.; Henderson, D.W.; Hsiao, T.C.; Alvio, A. Interactive water and nitrogen effects on maize. II. Photosynthetic decline and longevity of individual leaves. J. Agron. 1998, 80, 865–870. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Yallou, C.G.; Oyekunle, M. Genetic gains from selection for high grain yield and Striga resistance in early maturing maize cultivars of three breeding periods under Striga-infested and Striga-free environments. Field Crops Res. 2013, 147, 54–67. [Google Scholar] [CrossRef]
- Castleberry, R.M.; Crum, C.W.; Krull, C.F. Genetic yield improvement of U.S. maize cultivars under varying fertility and climatic environments. Crop Sci. 1984, 24, 33–36. [Google Scholar] [CrossRef]
- Tollenaar, M.; Nissanka, S.P.; Rajcan, I.; Bruulsema, T.W. Yield response of old and new corn hybrids to nitrogen. Better Crop. 1997, 81, 3–5. [Google Scholar]
- Sangoi, L.; Ender, M.; Guidolin, A.F.; De Almeida, M.L.; Konflanz, V.A. Nitrogen fertilization impact on agronomic traits of maize hybrids released at different decades. Pesqui. Agropecu. Bras. 2001, 36, 757–764. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, P.M.; Shanahan, J.F.; Schepers, J.S.; Caldwell, B. Agronomic responses of corn hybrids from different eras to deficit and adequate levels of water and nitrogen. J. Agron. 2004, 96, 1660–1667. [Google Scholar] [CrossRef] [Green Version]
- Oyekunle, M.; Ado, S.G.; Usman, I.S.; Abdulmalik, R.O.; Ahmed, H.O.; Hassan, L.B.; Yahaya, M.A. Gains in grain yield of released maize (Zea mays L.) cultivars under drought and well-watered conditions. Exp. Agric. 2019, 55, 934–944. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Oyekunle, M.; Akinwale, R.O. Genetic gains in grain yield under nitrogen stress following three decades of breeding for drought tolerance and Striga resistance in early maturing maize. J. Agric. Sci. 2015, 154, 647–661. [Google Scholar] [CrossRef]
- Kamara, A.Y.; Menkir, A.; Fakorede, M.A.B.; Ajala, S.O.; Badu-Apraku, B.; Kureh, I. Agronomic Performance of maize cultivars representing three decades of breeding in the Guinea savannas of West and Central Africa. J. Agric. Sci. 2004, 142, 567–575. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Yallou, C.G.; Obeng-Antwi, K.; Alidu, H.; Talabi, A.O.; Annor, B.; Oyekunle, M.; Akaogu, I.C.; Aderounmu, M. Yield gains in extra- early maize cultivars of three breeding eras under multiple environments. J. Agron. 2017, 109, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Badu-Apraku, B.; Oyekunle, M.; Menkir, A.; Haruna, A. Comparative Performance of Early-maturing Maize Cultivars Developed in Three Eras under Drought Stress and Well-watered Environments in West Africa. Crop Sci. 2013, 53, 1298–1311. [Google Scholar] [CrossRef]
- Chen, K.; Camberato, J.J.; Tuinstra, M.R.; Kumudini, S.V.; Tollenaar, M.; Vyn, T.J. Genetic improvement in density and nitrogen stress tolerance traits over 38 years of commercial maize hybrid release. Field Crop. Res. 2016, 196, 438–451. [Google Scholar] [CrossRef] [Green Version]
- Bänziger, M.; Lafitte, H.R. Efficiency of secondary traits for improving maize for low nitrogen target environments. Crop Sci. 1997, 37, 1110–1117. [Google Scholar] [CrossRef]
- Oyekunle, M.; Haruna, A.; Badu-Apraku, B.; Usman, I.S.; Mani, H.; Ado, S.G.; Olaoye, G.; Obeng-Antwi, K.; Abdulmalik, R.O.; Ahmed, H.O. Assessment of early-maturing maize hybrids and testing sites using GGE biplot analysis. Crop Sci. 2017, 57, 2942–2950. [Google Scholar] [CrossRef] [Green Version]
- Badu-Apraku, B.; Oyekunle, M. Genetic analysis of grain yield and other traits of extra-early yellow maize inbreds and hybrid performance under contrasting environments. Field Crops Res. 2012, 129, 99–110. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Gloria, B.A.; Abdoul-Madjidou, Y.; Johnson, T.; Samuel, A. Gains in Genetic Enhancement of Early Maturing Maize Hybrids Developed during Three Breeding Periods under Striga-Infested and Striga-Free Environments. J. Agron. 2020, 10, 1188. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Oyekunle, M. Agronomic traits associated with genetic gains in maize yield during three breeding eras in West Africa. Maydica 2014, 59, 49–57. [Google Scholar]
- Badu-Apraku, B.; Akinwale, R.O.; Ajala, S.O.; Menkir, A.; Fakorede, M.A.B.; Oyekunle, M. Relationships among traits of tropical early maize cultivars in contrasting environments. J. Agron. 2011, 103, 717–729. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Akinwale, R.O.; Franco, J.; Oyekunle, M. Assessment of reliability of secondary traits in selecting for improved grain yield in drought and low-nitrogen environments. Crop Sci. 2012, 52, 2050–2062. [Google Scholar] [CrossRef]
- Yan, W.; Fregeau-Reid, J.; Pageau, D.; Martin, R.; Mitchell-Fetch, J.; Etienne, M.; Rowsell, J.; Scott, P.; Price, M.; de Haan, B. Identifying essential test locations for oat breeding in Eastern Canada. Crop Sci. 2010, 50, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Soil Survey Staff. Soil taxonomy: A basic system of soil classification for making and interpreting soil surveys. In USDA-NRCS Agriculture Handbook No. 436, 2nd ed.; Gov. Print. Office: Washington, DC, USA, 1999; p. 869. [Google Scholar]
- SAS Institute Inc. SAS User’s Guide: Statistics; Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman: New York, NY, USA, 1996. [Google Scholar]
- Yan, W. GGE biplot: A Windows application for graphical analysis of multi-environment trial data and other types of two-way data. J. Agron. 2001, 93, 1111–1118. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Source of Variation | DF | Grain Yield, (kg/ha) | Days to Anthesis | Days to Silk | Anthesis Silking Interval | Plant Height (cm) | Ear Height (cm) | Root Lodging (%) | Stalk Lodging (%) | Husk Cover | Plant Aspect | Ear Aspect | Ear Rot | EPP | Stay-Green Characteristic |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low N Conditions | |||||||||||||||
| Environment (E) | 3 | 486,250 ** | 342.9 ** | 269.20 ** | 9.10 ** | 5616.10 ** | 3808.70 ** | 26.40 ** | 22,860.60 ** | 150.20 ** | 1.50 ** | 33.80 ** | 18,428.00 ** | 2.05 ** | 107.50 ** |
| Block (E×Rep) | 60 | 1,402,441 ** | 3.6 ** | 4.30 ** | 0.50 ns | 503.10 ** | 262.30 ** | 8.50 * | 15.90 ns | 0.40 ** | 1.00 ** | 1.20 ** | 79.50 ** | 0.01 ns | 1.10 ** |
| Rep (E) | 8 | 4,932,208 ** | 11.5 ** | 16.00 ** | 1.00 * | 1721.20 ** | 781.20 ** | 43.40 ** | 140.30 ** | 0.10 ns | 2.90 ** | 4.50 ** | 238.70 ** | 0.03 ** | 4.20 ** |
| Period | 2 | 10,464,454 ** | 0.7 ns | 2.50 ns | 1.20 ns | 4699.30 ** | 659.80 ** | 3.70 ns | 84.10 ns | 2.80 ** | 5.30 ** | 6.80 ** | 9.00 ns | 0.14 ** | 1.80 * |
| Hybrid (Period) | 51 | 1,169,502 ** | 10.4 ** | 11.80 ** | 0.60 * | 967.30 ** | 395.50 ** | 7.50 ** | 27.70 ns | 0.70 ** | 0.80 ** | 1.30 ** | 55.80 ** | 0.01 ns | 1.50 ** |
| E×Hybrid (Period) | 153 | 677,545 ** | 16 ** | 2.40 ** | 0.70 ** | 261.70 ** | 116.10 ns | 6.70 ns | 26.90 ns | 0.50 ** | 0.40 * | 0.60 ** | 34.10 ** | 0.01 ns | 0.50 ns |
| E×Period | 6 | 807,687 * | 2.2 * | 2.90 * | 0.90 ns | 210.00 ns | 63.30 ns | 7.00 ns | 81.20 * | 0.90 ** | 0.70 * | 0.20 ns | 27.50 ns | 0.03 * | 0.70 ns |
| Error | 364 | 361,686 | 1 | 1.1 | 0.4 | 165.5 | 96.1 | 5.7 | 30.1 | 0.2 | 0.3 | 0.4 | 17.6 | 0.01 | 0.4 |
| Repeatability | 0.56 | 0.84 | 0.8 | 0.44 | 0.79 | 0.75 | 0.14 | 0.23 | 0.41 | 0.6 | 0.63 | 0.37 | 0.22 | 0.68 | |
| High N Conditions | |||||||||||||||
| Environment (E) | 4 | 497,614,744 ** | 546.36 ** | 122.15 ** | 234.61 ** | 42,144.65 ** | 12,789.53 ** | 4901.94 ** | 699.24 ** | 180.14 ** | 167.00 ** | 155.36 ** | 11,843.41 ** | 2.33 ** | |
| Block (E×Rep) | 75 | 1,120,561 ** | 2.05 ** | 2.62 ** | 0.89 * | 345.10 ** | 222.56 * | 48.37 ns | 20.21 * | 0.58 ** | 0.68 ** | 0.69 ** | 37.12 ** | 0.02 * | |
| Rep (E) | 10 | 1,484,870 ** | 3.66 ** | 1.67 ns | 0.89 ns | 293.65 ns | 463.66 ** | 58.80 ns | 14.79 ns | 1.07 ** | 1.13 ** | 0.71 ns | 452.26 ** | 0.07 ** | |
| Period | 2 | 7,534,475 ** | 19.31 ** | 3.27 ns | 5.04 ** | 6695.97 ** | 1607.07 ** | 42.45 ns | 18.81 ns | 2.18 ** | 3.49 ** | 3.16 ** | 9.23 ns | 0.12 ** | |
| Hybrid (Period) | 51 | 2,836,305 ** | 18.87 ** | 21.29 ** | 1.46 ** | 1763.46 ** | 657.48 ** | 145.77 ** | 30.30 ** | 0.88 ** | 0.94 ** | 1.55 ** | 33.29 ** | 0.03 ** | |
| E×Hybrid (Period) | 204 | 1,251,778 ** | 2.89 ** | 3.27 ** | 1.12 ** | 274.62 * | 175.43 ns | 70.86 ** | 21.68 ** | 0.61 ** | 0.57 *8 | 0.67 ** | 22.76 ** | 0.02 ** | |
| E×Period | 8 | 637,817 ns | 4.23 ** | 4.98 ** | 3.08 ** | 233.82 ns | 83.93 ns | 30.43 ns | 10.15 ns | 1.08 ** | 1.65 ** | 1.23 ** | 26.49 ** | 0.03 * | |
| Error | 455 | 479,051 | 1.34 | 1.59 | 0.63 | 214.61 | 152.06 | 44.87 | 15.55 | 0.36 | 0.28 | 0.39 | 11.29 | 0.02 | |
| Repeatability | 0.59 | 0.86 | 0.85 | 0.47 | 0.86 | 0.77 | 0.51 | 0.35 | 0.33 | 0.38 | 0.56 | 0.30 | 0.24 | ||
| Trait | Period | Low N Conditions | High N Conditions |
|---|---|---|---|
| Grain yield (kg/ha) | 2008–2010 | 2784.03 ± 75.11 | 4294.33 ± 111.79 |
| 2011–2013 | 2932.54 ± 91.12 | 4330.10 ± 99.38 | |
| 2014–2016 | 3196.79 ± 66.59 | 4614.44 ± 116.52 | |
| Days to anthesis | 2008–2010 | 53.19 ± 0.17 | 51.28 ± 0.21 |
| 2011–2013 | 53.06 ± 0.23 | 51.29 ± 0.27 | |
| 2014–2016 | 53.09 ± 0.28 | 51.77 ± 0.35 | |
| Days to silking | 2008–2010 | 53.99 ± 0.16 | 52.42 ± 0.21 |
| 2011–2013 | 53.96 ± 0.23 | 52.52 ± 0.30 | |
| 2014–2016 | 53.78 ± 0.32 | 52.65 ± 0.37 | |
| Anthesis-silking interval | 2008–2010 | 0.85 ± 0.07 | 1.33 ± 0.07 |
| 2011–2013 | 0.91 ± 0.78 | 1.23± 0.07 | |
| 2014–2016 | 0.78 ± 0.06 | 1.05 ± 0.09 | |
| Plant height (cm) | 2008–2010 | 157.03 ± 1.95 | 161.76 ± 2.42 |
| 2011–2013 | 165.42 ± 2.55 | 170.53 ± 3.20 | |
| 2014–2016 | 164.18 ± 2.20 | 171.49 ± 2.17 | |
| Ear height (cm) | 2008–2010 | 70.86 ± 1.46 | 73.96 ± 1.60 |
| 2011–2013 | 74.58 ± 1.60 | 79.29 ± 1.91 | |
| 2014–2016 | 71.94 ± 1.11 | 76.64 ± 1.39 | |
| Root lodging % | 2008–2010 | 0.90 ± 0.17 | 4.75 ± 0.89 |
| 2011–2013 | 0.70 ± 0.16 | 5.58 ± 0.77 | |
| 2014–2016 | 0.99 ± 0.22 | 4.98 ± 0.68 | |
| Stalk lodging % | 2008–2010 | 6.17 ± 0.34 | 2.89 ± 0.32 |
| 2011–2013 | 6.77 ± 0.41 | 3.10 ± 0.40 | |
| 2014–2016 | 5.14 ± 0.35 | 2.55 ± 0.33 | |
| Husk cover | 2008–2010 | 3.11 ± 0.06 | 3.41 ± 0.05 |
| 2011–2013 | 3.20 ± 0.08 | 3.41 ± 0.06 | |
| 2014–2016 | 2.94 ± 0.05 | 3.25 ± 0.07 | |
| Plant aspect | 2008–2010 | 4.76 ± 0.05 | 4.47 ± 0.06 |
| 2011–2013 | 4.61 ± 0.08 | 4.35 ± 0.06 | |
| 2014–2016 | 4.39 ± 0.06 | 4.23 ± 0.06 | |
| Ear aspect | 2008–2010 | 4.65 ± 0.08 | 3.92 ± 0.09 |
| 2011–2013 | 4.34 ± 0.10 | 3.76 ± 0.07 | |
| 2014–2016 | 4.42 ± 0.07 | 3.69 ± 0.08 | |
| Ear rot | 2008–2010 | 8.48 ± 0.51 | 6.04 ± 0.33 |
| 2011–2013 | 8.51 ± 0.51 | 5.64 ± 0.42 | |
| 2014–2016 | 8.15 ± 0.58 | 5.81 ± 0.34 | |
| Ears per plant | 2008–2010 | 0.80 ± 0.01 | 0.82 ± 0.01 |
| 2011–2013 | 0.82 ± 0.01 | 0.86 ± 0.01 | |
| 2014–2016 | 0.84 ± 0.01 | 0.86 ± 0.01 | |
| Stay-green characteristic | 2008–2010 | 3.31 ± 0.09 | − |
| 2011–2013 | 3.22 ± 0.08 | − | |
| 2014–2016 | 3.14 ± 0.09 | − |
| Trait | Relative Gain (% per Year) | R2 | a | b |
|---|---|---|---|---|
| Low N | ||||
| Grain yield (kg/ha) | 2.91 | 0.26 | 2588.70 | 75.37 ** |
| Days to anthesis | 0.02 | 0.01 | 53.06 | 0.01 ns |
| Days to silk | −0.02 | 0.00 | 53.95 | −0.01 ns |
| Anthesis-silking interval | −1.88 | 0.03 | 0.94 | −0.02 ns |
| Plant height (cm) | 1.07 | 0.17 | 153.89 | 1.64 ** |
| Ear height (cm) | 0.57 | 0.03 | 70.42 | 0.40 ns |
| Root lodging (%) | 3.12 | 0.01 | 0.75 | 0.02 ns |
| Stalk lodging (%) | 0.50 | 0.00 | 5.88 | 0.03 ns |
| Husk cover | −0.33 | 0.01 | 3.14 | −0.01 ns |
| Plant aspect | −1.07 | 0.18 | 4.85 | −0.05 ** |
| Ear aspect | −1.31 | 0.17 | 4.79 | −0.06 ** |
| Ear rot | −0.80 | 0.01 | 8.74 | −0.07 ns |
| Stay-green characteristic | −0.89 | 0.04 | 3.38 | −0.03 ns |
| Ears/plant | 1.08 | 0.27 | 0.78 | 0.01 ** |
| High N | ||||
| Grain yield (kg/ha) | 1.33 | 0.08 | 4133.30 | 55.12 * |
| Days to anthesis | 0.19 | 0.04 | 50.95 | 0.10 ns |
| Days to silk | 0.12 | 0.02 | 52.21 | 0.06 ns |
| Anthesis silking interval | −3.15 | 0.11 | 1.43 | −0.05 * |
| Plant height (cm) | 1.11 | 0.14 | 159.00 | 1.76 ** |
| Ear height (cm) | 0.84 | 0.05 | 73.51 | 0.62 ns |
| Root lodging (%) | 1.87 | 0.00 | 4.66 | 0.09 ns |
| Stalk lodging (%) | −0.92 | 0.00 | 2.99 | −0.03 ns |
| Husk cover | −0.68 | 0.05 | 3.47 | −0.02 ns |
| Plant aspect | −0.95 | 0.15 | 4.57 | −0.04 ** |
| Ear aspect | −0.99 | 0.08 | 3.99 | −0.04 * |
| Ear rot | −1.16 | 0.01 | 6.20 | −0.07 ns |
| Ears/plant | 0.81 | 0.13 | 0.82 | 0.01 ** |
| Grain Yield, (kg/ha) | Days to Anthesis | Days to Silk | ASI | Plant Height (cm) | Ear Height (cm) | Husk Cover | Plant Aspect | Ear Aspect | Ear Rot | Ears Per Plant | Stay-Green Characteristic | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grain yield | 0.17 ns | 0.11 ns | −0.04 ns | 0.47 ** | 0.26 * | −0.21 ns | −0.65 ** | −0.61 ** | −0.20 ns | 0.49 ** | −0.29 * | |
| Days to anthesis | −0.02 ns | 0.96 ** | 0.03 ns | 0.36 ** | 0.41 ** | −0.28 * | −0.32 * | −0.11 ns | −0.48 ** | −0.17 ns | 0.37 ** | |
| Days to silk | −0.13 ns | 0.95 ** | 0.28 * | 0.33 ** | 0.39 ** | −0.24 ns | −0.27 ns | −0.02 ns | −0.41 ** | −0.17 ns | 0.35 ** | |
| ASI | −0.38 ** | 0.11 ns | 0.36 ** | −0.13 ns | −0.10 ns | 0.17 ns | 0.11 ns | 0.23 ns | 0.09 ns | 0.04 ns | −0.02 ns | |
| Plant height | 0.32 ** | 0.27 * | 0.29 * | 0.02 ns | 0.80 ** | −0.06 ns | −0.55 ** | −0.25 ns | 0.00 ns | 0.19 ns | 0.40 ** | |
| Ear height | 0.09 ns | 0.36 ** | 0.42 ** | 0.21 ns | 0.80 ** | 0.00 ns | −0.45 ns | −0.20 ns | −0.01 ns | 0.03 ns | 0.38 ** | |
| Husk cover | −0.07 ns | −0.32 * | −0.33 ** | −0.07 ns | −0.35 ** | −0.26 * | 0.45 ** | 0.10 ns | 0.34 ** | −0.15 ns | 0.16 ns | |
| Plant aspect | −0.59 ** | −0.27 * | −0.25 * | 0.00 ns | −0.59 ** | −0.49 ** | 0.60 ** | 0.46 ** | 0.26 * | −0.36 ** | 0.11 ns | |
| Ear aspect | −0.80 ** | 0.02 ns | 0.10 ns | 0.25 * | −0.22 ns | −0.04 ns | 0.11 na | 0.58 ** | 0.28 * | −0.36 ** | 0.22 ns | |
| Ear rot | 0.04 ns | −0.42 ** | −0.42 ** | −0.18 ns | 0.03 ns | −0.09 ns | 0.37 ** | 0.33 * | 0.20 ns | 0.05 ns | 0.13 ns | |
| EPP | 0.60 ** | 0.12 ns | 0.07 ns | −0.19 ns | 0.20 ns | 0.08 ns | −0.17 ns | −0.18 ns | −0.19 ns | −0.14 ns | −0.42 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badu-Apraku, B.; Fakorede, M.A.B.; Abubakar, A.M. Accelerated Genetic Gains in Early-Maturing Maize Hybrids following Three Periods of Genetic Enhancement for Grain Yield under Low and High Soil-Nitrogen Environments. Plants 2022, 11, 1208. https://doi.org/10.3390/plants11091208
Badu-Apraku B, Fakorede MAB, Abubakar AM. Accelerated Genetic Gains in Early-Maturing Maize Hybrids following Three Periods of Genetic Enhancement for Grain Yield under Low and High Soil-Nitrogen Environments. Plants. 2022; 11(9):1208. https://doi.org/10.3390/plants11091208
Chicago/Turabian StyleBadu-Apraku, Baffour, Morakinyo Abiodun Bamidele Fakorede, and Adamu Masari Abubakar. 2022. "Accelerated Genetic Gains in Early-Maturing Maize Hybrids following Three Periods of Genetic Enhancement for Grain Yield under Low and High Soil-Nitrogen Environments" Plants 11, no. 9: 1208. https://doi.org/10.3390/plants11091208
APA StyleBadu-Apraku, B., Fakorede, M. A. B., & Abubakar, A. M. (2022). Accelerated Genetic Gains in Early-Maturing Maize Hybrids following Three Periods of Genetic Enhancement for Grain Yield under Low and High Soil-Nitrogen Environments. Plants, 11(9), 1208. https://doi.org/10.3390/plants11091208