
Role of LOC_Os01g68450, Containing DUF2358, in Salt Tolerance Is Mediated via Adaptation of Absorbed Light Energy Dissipation

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
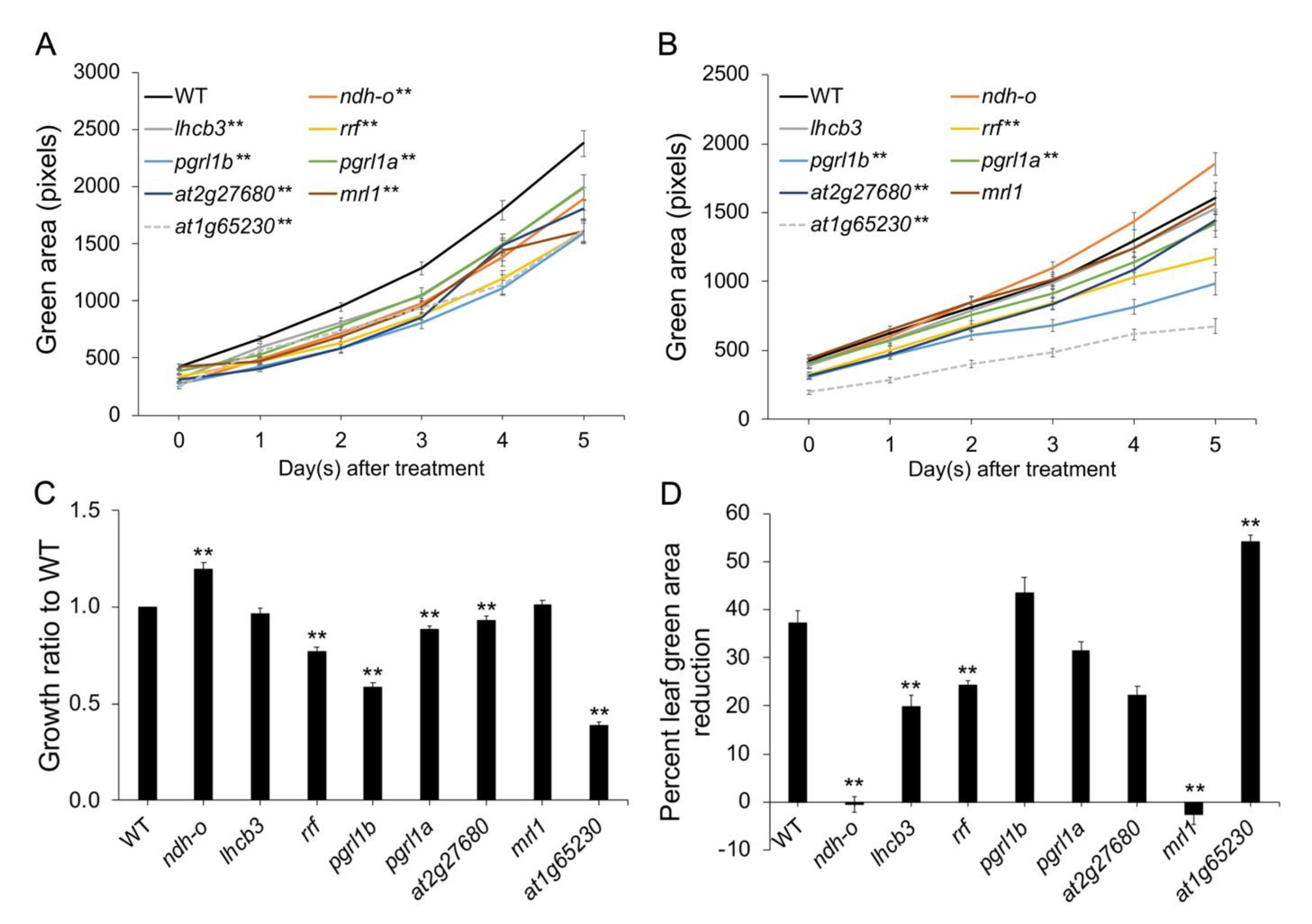
2.1. Mutation in At1g65230 Strongly Inhibited Plant Growth under Salt Stress
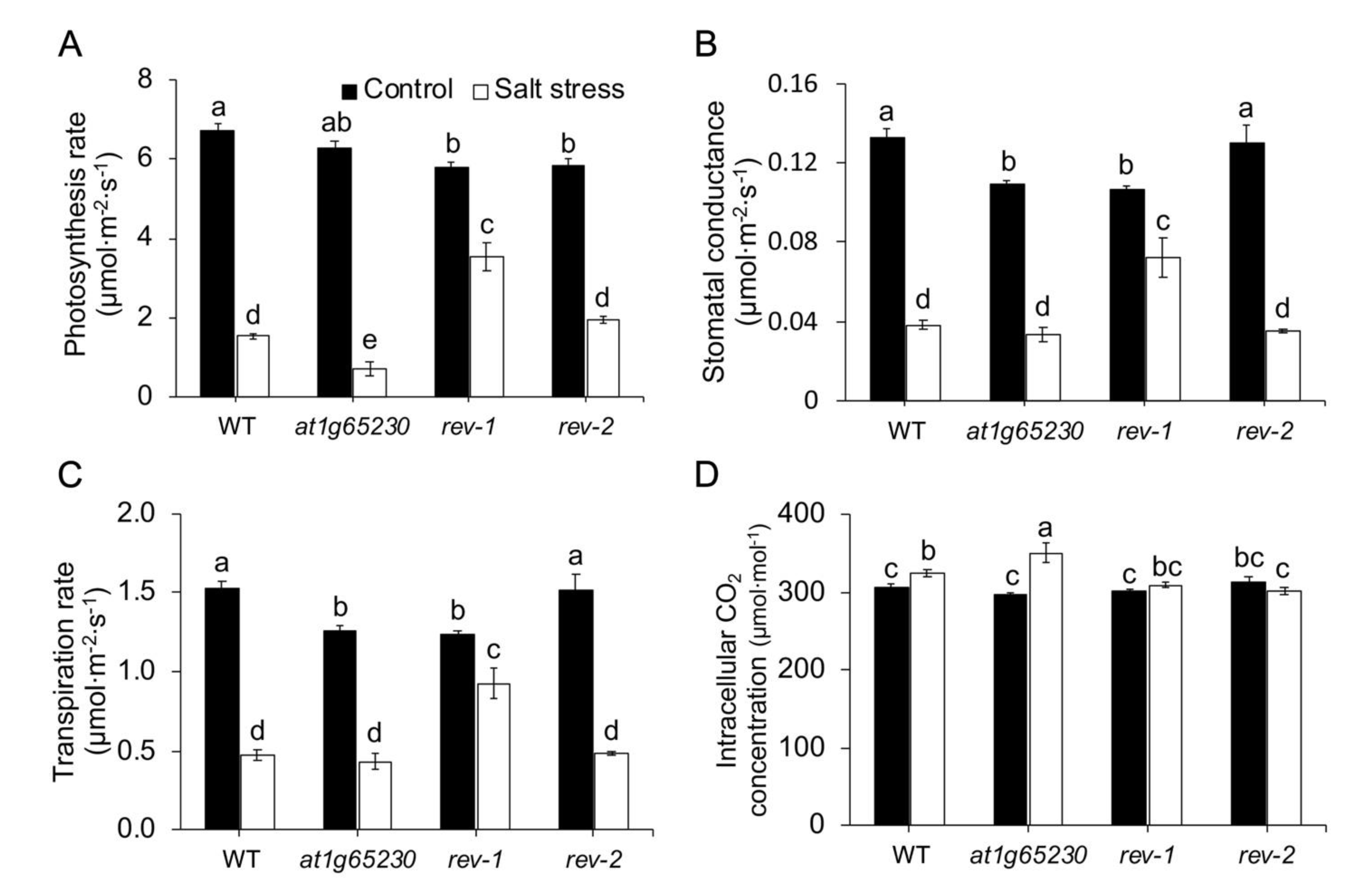
2.2. Ectopic Expression of LOC_Os01g68450 Reverted the Susceptibility of the At1g65230 Mutant to Salt Stress via the Light Reaction Adaptation
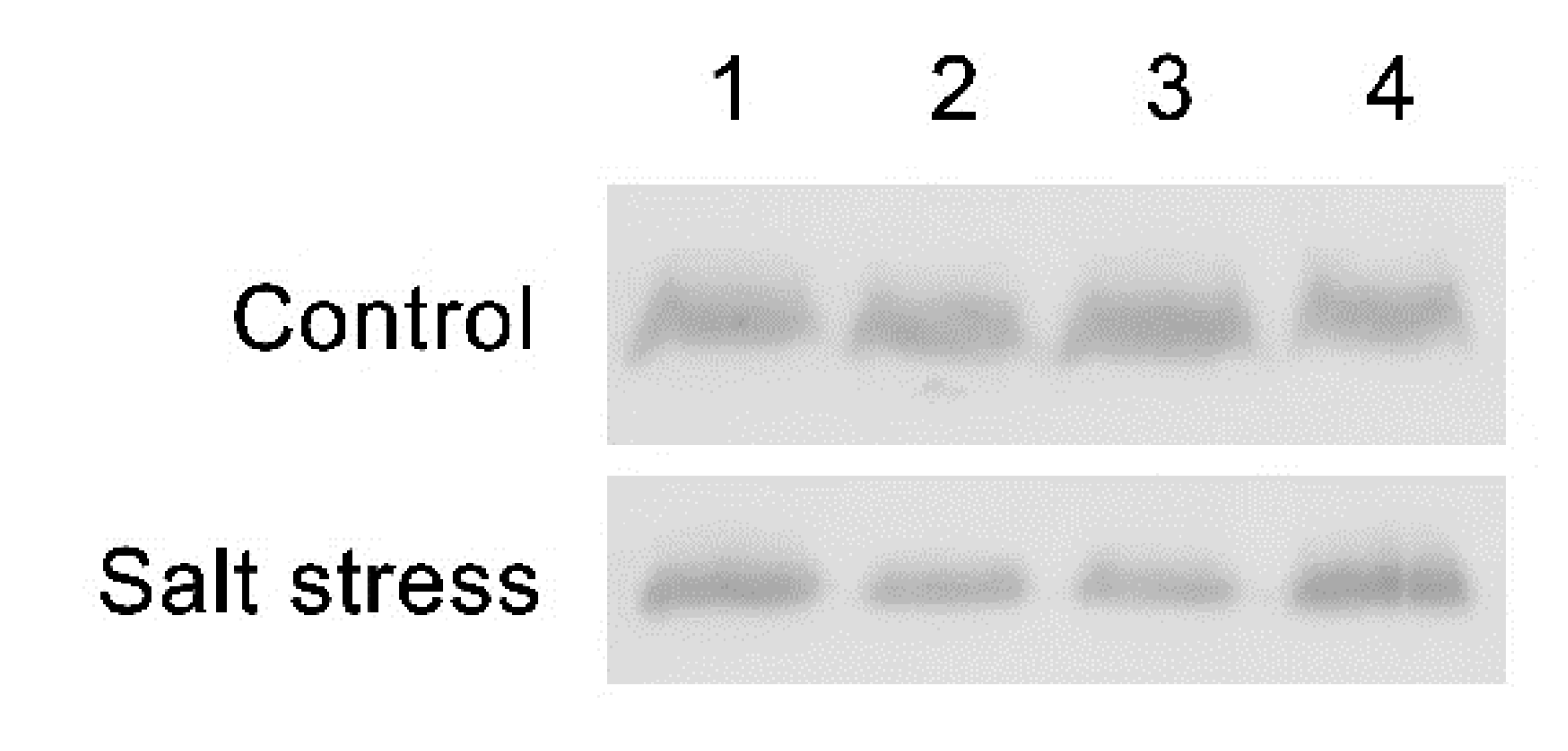
2.3. Investigation of At1g65230 and LOC_Os01g68450 Genes on PsbD Protein Stability under Salt Stress Condition
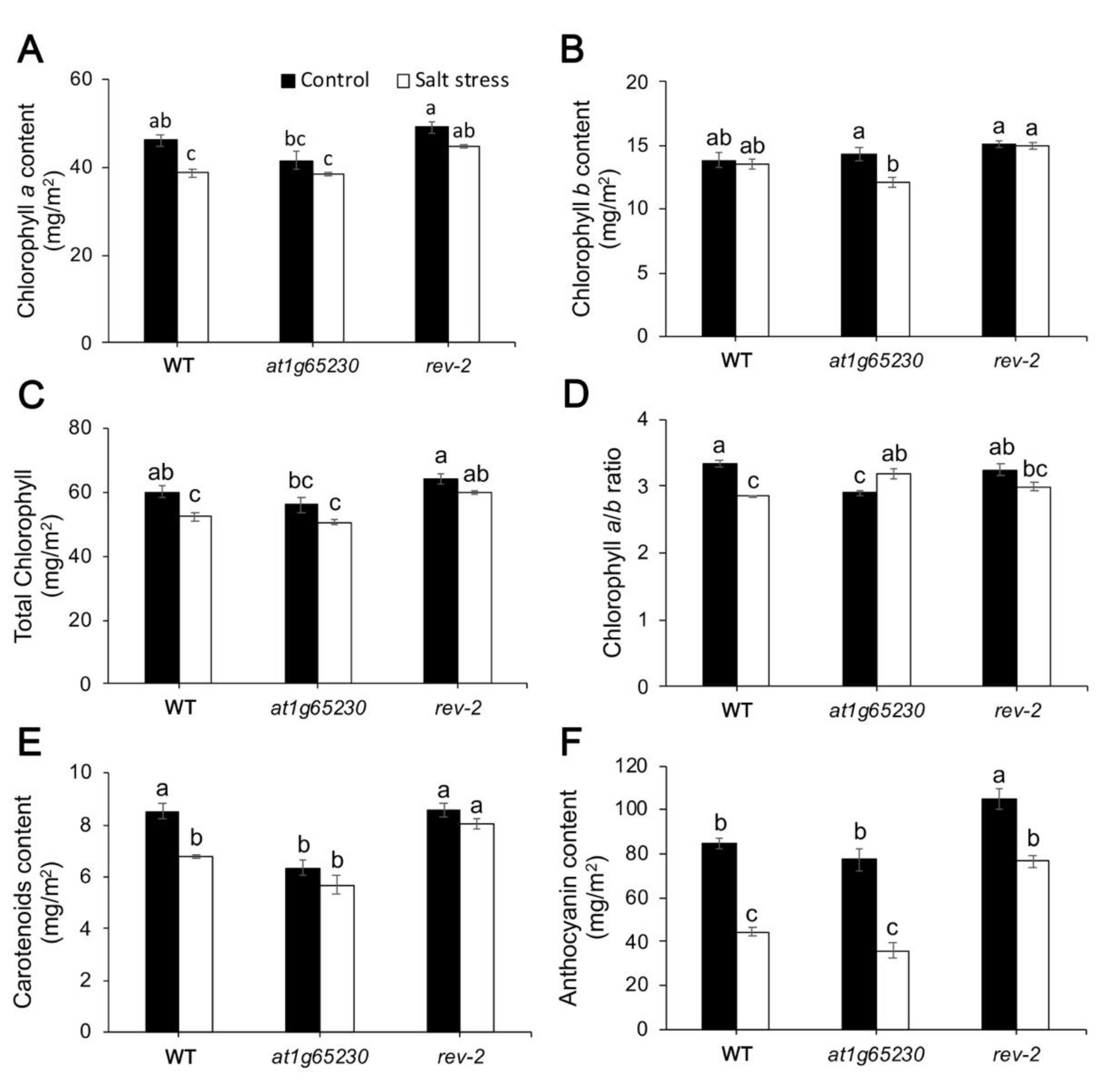
2.4. Ectopic Expression of LOC_Os01g68450 Gene Affected Photosynthetic Pigment Levels
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Ectopic Expression and Revertant Line Generation
4.3. Estimating the Green Area of Leaves
4.4. Estimating Physiological Parameters
4.4.1. Growth Conditions
4.4.2. Photosynthetic Parameters
4.4.3. Photosynthetic Pigment and Anthocyanin Content
4.4.4. Chlorophyll Fluorescence Induction Curves
4.5. Quantification of Gene Expression
4.5.1. RNA Extraction
4.5.2. Gene Quantification
| at1g65230-F | AATCCGCAAAAGATGTCGCC |
| at1g65230-R | GACAATGGCTAGAAACAGTGCAA |
| LOC_Os01g68450-F | AGGGTACAGCATCTCGGCTA |
| LOC_Os01g68450-R | CAGGAAGAGCGCAATCTGGT |
| EF1α-F | TGCCGCAGGTGAATCAAAGG |
| EF1α-R | CCCAATTACGAGAACAACGCTCTG |
4.6. Immunoblotting
4.6.1. Extraction of Thylakoid Membranes
4.6.2. Western Blot Analysis
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, M.S. Present Scenario of Global Salt Affected Soil, its Management and Importance of Salinity Research. Int. Res. J. Biol. Sci. 2019, 1, 1–3. [Google Scholar]
- Li, W.; Li, Q. Effect of Environment Salt Stress on Plant and the Molecular Mechanism of Salt Stress Tolerance. Int. J. Environ. Sci. Nat. Resour. 2017, 7, 81–86. [Google Scholar]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanjoo, V.; Jearakongman, S.; Punyawaew, K.; Siangliw, J.L.; Siangliw, M.; Vanavichit, A.; Toojinda, T. Co-Location of Quantitative Trait Loci for Drought and Salinity Tolerance in Rice. Thai J. Genet. 2011, 4, 126–138. [Google Scholar]
- Punchkhon, C.; Plaimas, K.; Buaboocha, T.; Siangliw, J.L.; Toojinda, T.; Comai, L.; De Diego, N.; Spíchal, L.; Chadchawan, S. Drought-tolerance gene identification using genome comparison and co-expression network analysis of chromosome substitution lines in rice. Genes 2020, 11, 1197. [Google Scholar] [CrossRef]
- Chutimanukul, P.; Kositsup, B.; Plaimas, K.; Buaboocha, T.; Siangliw, M.; Toojinda, T.; Comai, L.; Chadchawan, S. Photosynthetic Responses and Identification of Salt Tolerance Genes in a Chromosome Segment Substitution Line of ‘Khao Dawk Mali 105′ Rice. Environ. Exp. Bot. 2018, 155, 497–508. [Google Scholar] [CrossRef]
- Chutimanukul, P. Salt Tolerant Gene Identification in Rice Using Chromosome Substitution Lines with ‘Khao Dawk Mali 105′ Rice Genetic Background. Doctoral Dissertation, Chulalongkorn University, Bangkok, Thailand, 30 June 2018. [Google Scholar]
- Tóth, S.Z.; Oukarroum, A.; Schansker, G. Probing the Photosynthetic Apparatus Noninvasively in the Laboratory of Reto Stresser in the Countryside of Geneva between 2001 and 2009. Photosynthetica 2020, 58, 560–572. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Zhu, W.; Hamilton, J.; Lin, H.; Campbell, M.; Childs, K.; Thibaud-Nissen, F.; Malek, R.L.; Lee, Y.; Zheng, L.; et al. The TIGR Rice Genome Annotation Resource: Improvements and New Features. Nucleic Acids Res. 2007, 35, D883–D887. [Google Scholar] [CrossRef] [Green Version]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The Arabidopsis Information Resource: Making and Mining the ‘Gold Standard’ Annotated Reference Plant Genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef] [Green Version]
- Moradi, F.; Ismail, A.M. Responses of Photosynthesis, Chlorophyll Fluorescence and ROS-Scavenging Systems to Salt Stress during Seedling and Reproductive Stages in Rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [Green Version]
- Soussi, M.; Ocana, A.; Lluch, C. Effects of Salt Stress on Growth, Photosynthesis and Nitrogen Fixation in Chick-Pea (Cicer arietinum L.). J. Exp. Bot. 1998, 49, 1329–1337. [Google Scholar] [CrossRef]
- Helena, H.; František, H.; Jaroslava, M.; Kamil, K. Effects of Salt Stress on Water Status, Photosynthesis and Chlorophyll Fluorescence of Rocket. Plant Soil Environ. 2017, 63, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of Exogenous Selenium Supply on Photosynthesis, Na+ Accumulation and Antioxidative Capacity of Maize (Zea mays L.) under Salinity Stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taj, Z.; Challabathula, D. Protection of Photosynthesis by Halotolerant Staphylococcus sciuri ET101 in Tomato (Lycoperiscon esculentum) and Rice (Oryza sativa) Plants during Salinity Stress: Possible Interplay between Carboxylation and Oxygenation in Stress Mitigation. Front. Microbiol. 2021, 11, 1–24. [Google Scholar] [CrossRef]
- Vernotte, C.; Etienne, A.L.; Briantais, J.M. Quenching of the System II Chlorophyll Fluorescence by the Plastoquinone Pool. Biochim. Biophys. Acta 1979, 545, 519–527. [Google Scholar]
- Velthuys, B.R. Electron-Dependent Competition Between Plastoquinone and Inhibitors for Binding to Photosystem II. FEBS Lett. 1981, 126, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Latowski, D.; Kuczyńska, P.; Strzałka, K. Xanthophyll Cycle—A Mechanism Protesting Plants Against Oxidative Stress. Redox Rep. 2011, 16, 78–90. [Google Scholar] [CrossRef]
- Cebrián, G.; Iglesias-Moya, J.; García, A.; Martínez, J.; Romero, J.; Regalado, J.J.; Martínez, C.; Valenzuela, J.L.; Jamilena, M. Involvement of Ethylene Receptors in the Salt Tolerance Response of Cucurbita pepo. Hortic. Res. 2021, 8, 73. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Mahmood, K.; Rothstein, S.J. ROS Induces Anthocyanin Production via Late Biosynthetic Genes and Anthocyanin Deficiency Confers the Hypersensitivity to ROS-Generating Stresses in Arabidopsis. Plant Cell Physiol. 2017, 58, 1364–1377. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, J.; Gu, R.; Yue, L.; Wang, H.; Zhan, X.; Xing, B. Carotenoid and Superoxide Dismutase Are the Most Effective Antioxidants Participating in ROS Scavenging in Phenanthrene Accumulated Wheat Leaf. Chemosphere 2018, 197, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, X.; Ma, B.; Wu, Z.; Zheng, L.; Qi, Z.; Wang, Y. A Leucoanthocyanidin Dioxygenase Gene (RtLDOX2) from the Feral Forage Plant Reaumuria trigyna Promotes the Accumulation of Flavonoids and Improves Tolerance to Abiotic Stresses. J. Plant Res. 2021, 134, 1121–1138. [Google Scholar] [CrossRef] [PubMed]
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M.; et al. The Arabidopsis Information Resource (TAIR): Improved Gene Annotation and New Tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef]
- De Diego, N.; Fürst, T.; Humplík, J.F.; Ugena, L.; Podlešáková, K.; Spíchal, L. An Automated Method for High-Throughput Screening of Arabidopsis Rosette Growth in Multi-Well Plates and Its Validation in Stress Condition. Front. Plant Sci. 2017, 8, 1702. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Tanaka, A.; Tanaka, R. Simple Extraction Methods That Prevent the Artifactual Conversion of Chlorophyll to Chlorophyllide during Pigment Isolation from Leaf Samples. Plant Methods 2013, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Neff, M.M.; Chory, J. Genetic Interactions between Phytochrome A, Phytochrome B, and Cryptochrome 1 during Arabidopsis Development. Plant Physiol. 1998, 118, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Frankel, L.K.; Bricker, T. Functional Analysis of Photosystem II in a PsbO-1-Deficient Mutant in Arabidopsis thaliana. Biochemistry 2007, 46, 7607–7613. [Google Scholar] [CrossRef]
- Bevitori, R.; Oliveira, M.; Grossi-de-Sá, M.; Lanna, A.; Silveira, R. Selection of Optimized Candidate Reference Genes for qRT-PCR Normalization in Rice (Oryza sativa L.) During Magnaporthe oryzae Infection and Drought. GMR 2014, 13, 9795–9805. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Järvi, S.; Suorsa, M.; Paakkarinen, V.; Aro, E.M. Optimized Native Gel Systems for Separation of Thylakoid Protein Complexes: Novel Super- and Mega-Complexes. Biochem. J. 2011, 439, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, J.J. Chapter 6 Immunoblotting. In Methods in Cell Biology; Asai, D.J., Ed.; Academic Press: Cambridge, MA, USA, 1993; Volume 37, pp. 105–117. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Punchkhon, C.; Chutimanukul, P.; Chokwiwatkul, R.; Saputro, T.B.; Grennan, A.K.; Diego, N.D.; Spíchal, L.; Chadchawan, S. Role of LOC_Os01g68450, Containing DUF2358, in Salt Tolerance Is Mediated via Adaptation of Absorbed Light Energy Dissipation. Plants 2022, 11, 1233. https://doi.org/10.3390/plants11091233
Punchkhon C, Chutimanukul P, Chokwiwatkul R, Saputro TB, Grennan AK, Diego ND, Spíchal L, Chadchawan S. Role of LOC_Os01g68450, Containing DUF2358, in Salt Tolerance Is Mediated via Adaptation of Absorbed Light Energy Dissipation. Plants. 2022; 11(9):1233. https://doi.org/10.3390/plants11091233
Chicago/Turabian StylePunchkhon, Chutarat, Panita Chutimanukul, Ratchata Chokwiwatkul, Triono Bagus Saputro, Aleel K. Grennan, Nuria De Diego, Lukáš Spíchal, and Supachitra Chadchawan. 2022. "Role of LOC_Os01g68450, Containing DUF2358, in Salt Tolerance Is Mediated via Adaptation of Absorbed Light Energy Dissipation" Plants 11, no. 9: 1233. https://doi.org/10.3390/plants11091233