

Can Essential Oils Be a Natural Alternative for the Control of Spodoptera frugiperda? A Review of Toxicity Methods and Their Modes of Action


Abstract
1. Introduction
2. Essential Oils Evaluated against Spodoptera frugiperda
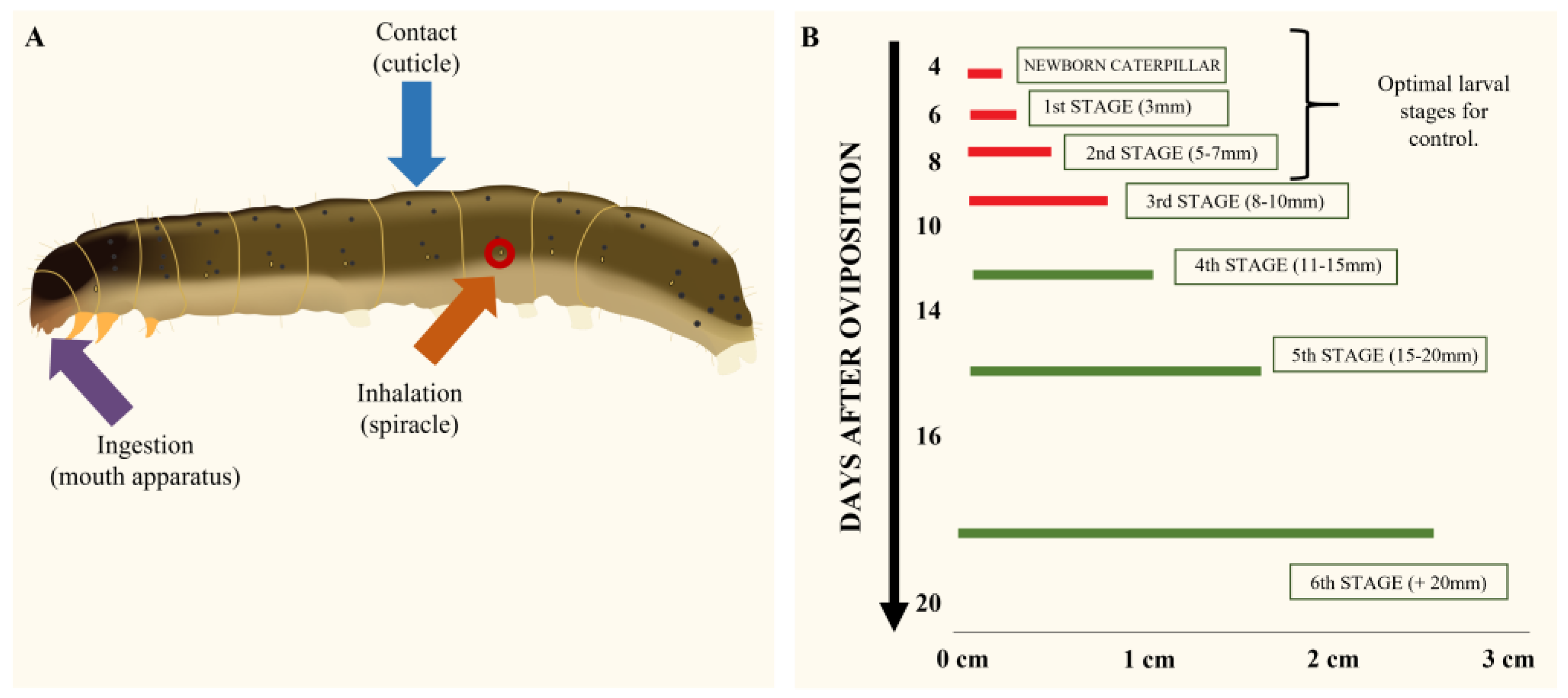
3. Routes of Entry of Essential Oils
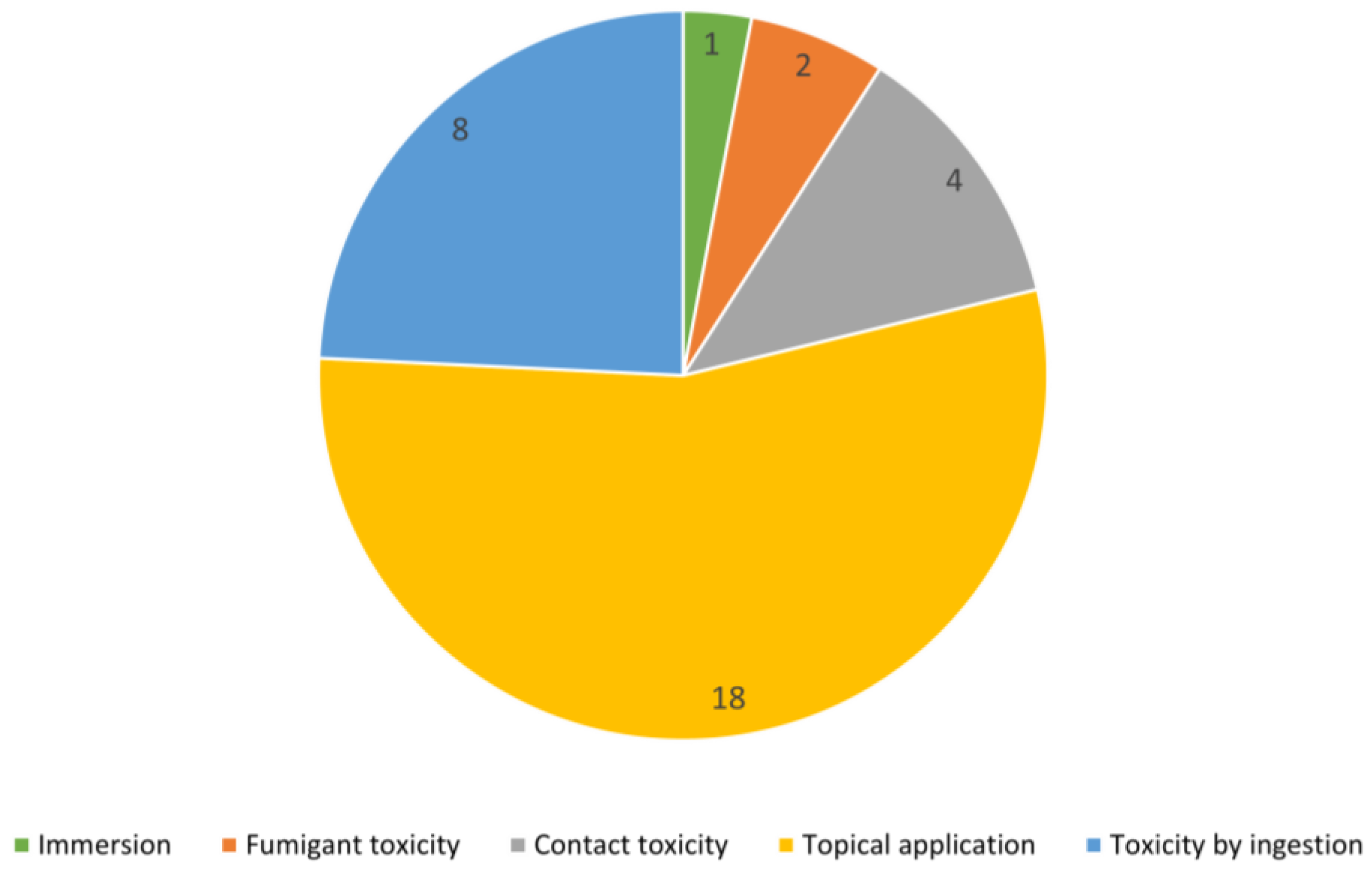
4. Toxicological Methods against Spodoptera frugiperda
5. Toxicity of Essential Oils against Spodoptera frugiperda
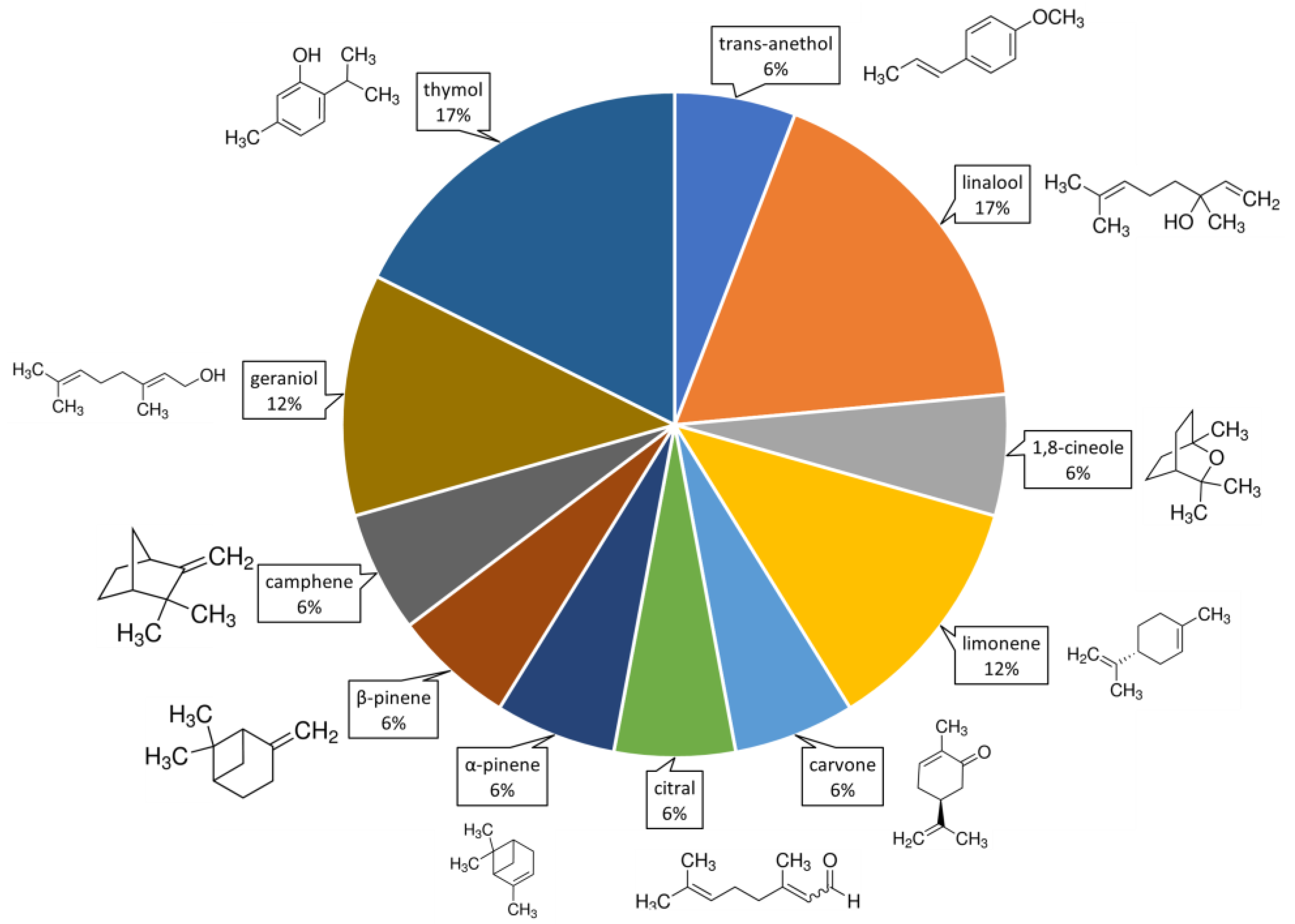
5.1. Volatile Organic Compounds Tested against Spodoptera frugiperda
5.2. Comparison of Insecticidal Effects of EOs between S. frugiperda and the S. littoralis-S. litura-S. exigua Complex
5.3. Structure–Activity Relationship
6. Mode of Action of Essential Oil in Insects
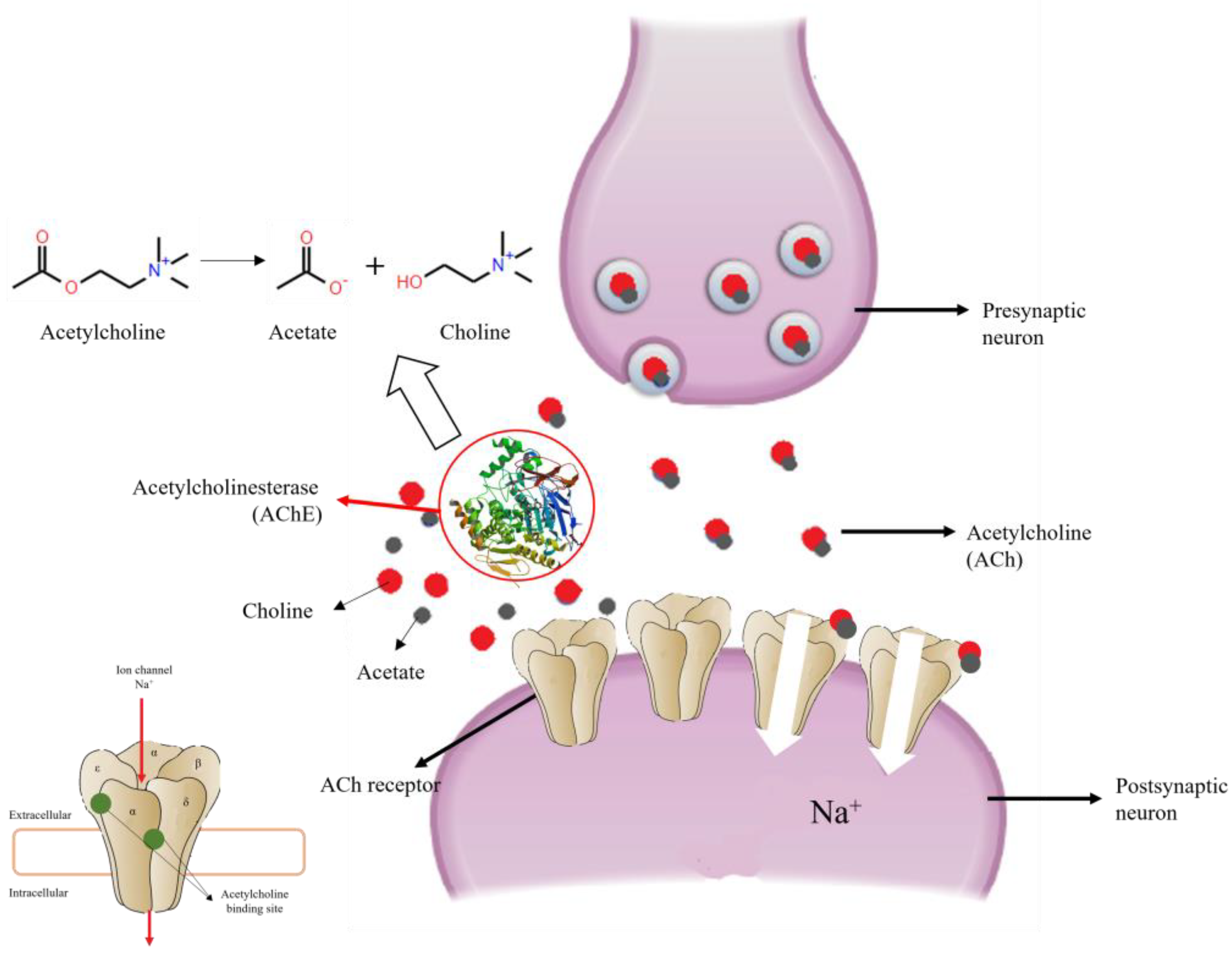
6.1. Acetylcholinesterase (AChE)
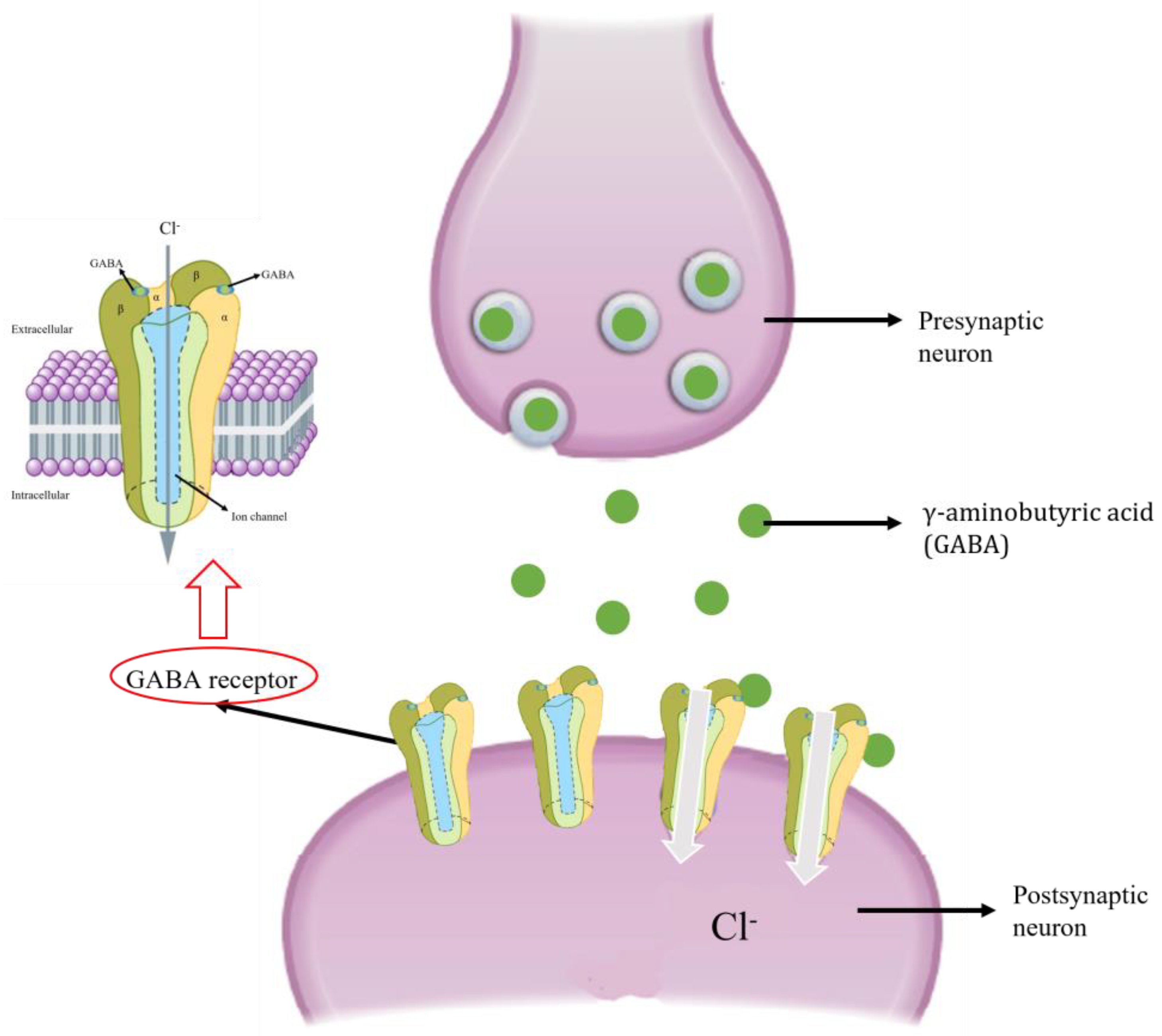
6.2. GABA Receptor
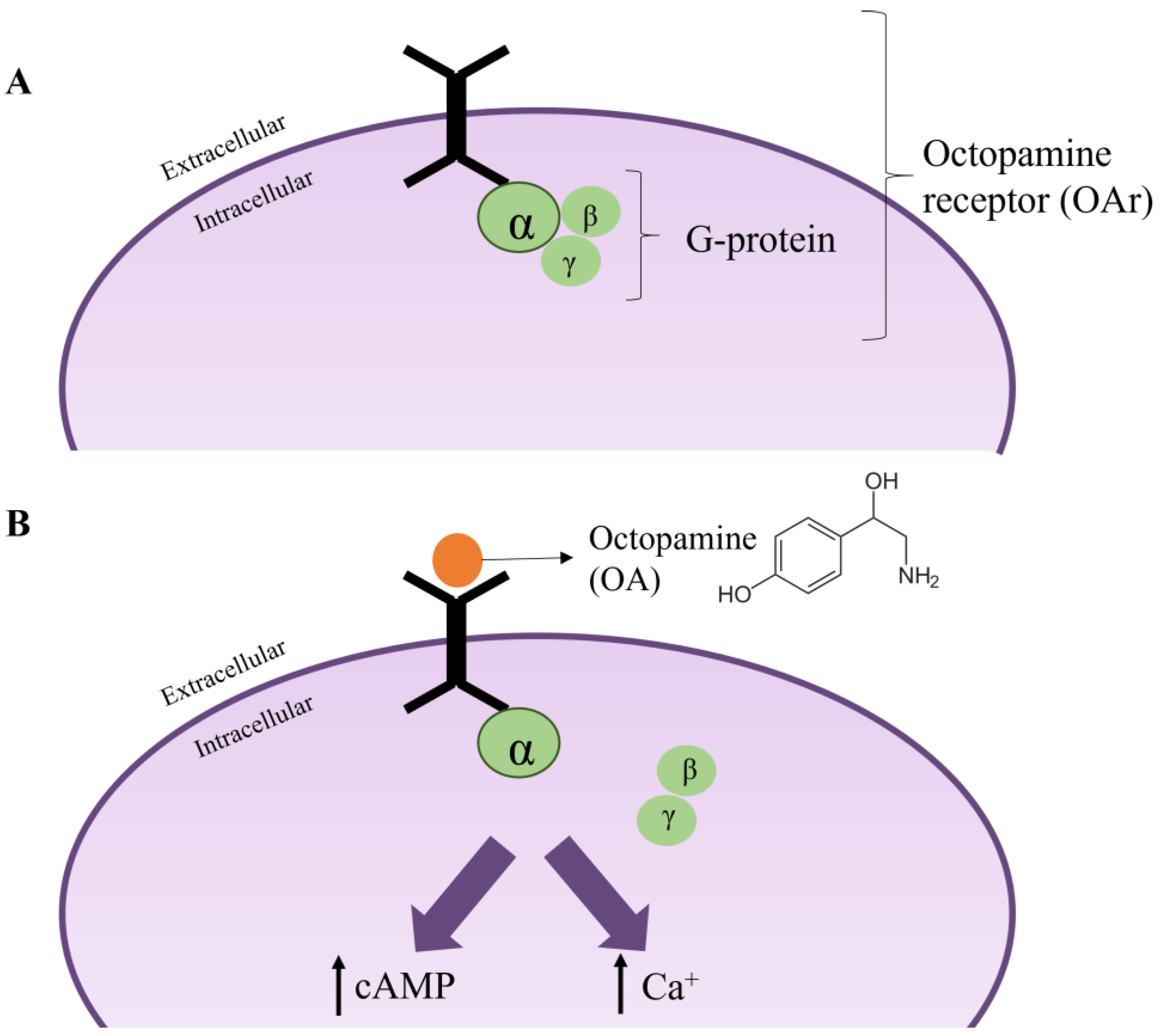
6.3. Octopamine
6.4. Other Modes of Action
7. Final Considerations
- The method most used for evaluating the toxicity of EOs on S. frugiperda was topical application, where the bioactive compound enters the organism through the cuticle. Considering the field application method of traditional insecticides, this method of topical application simulates what happens when the insect pest is found in the cultivar and is reached by traditional spraying. Thus, this testing method could be recommended for laboratory study using mainly third-instar larvae, in order to obtain comparable results with already published articles.
- The most effective EOs were Ocimum gratissimum, Siparuna guianesis, Piper marginatum, Piper septuplinervium, Cymbopogon citratus, Citrus limon, and Ageratum conyzoides for the methods of topical application, contact toxicity, immersion, fumigant and ingestion, respectively. In general, these essential oils presented a high percentage of non-oxygenated volatile compounds, with the exception of C. citratus EO, thereby allowing us to predict that against this insect, terpene hydrocarbon-type compounds would present a greater toxicity. However, mostly only pure oxygenated compounds have been tested, with anethole being the most toxic of these. We suggest that mixtures of lipophilic and hydrophilic compounds could have a greater toxic effect as the former act as vehicles for the latter to cross the insect cuticle and facilitate their arrival at the active site.
- EOs and their pure compounds are approximately 1000 to 100,000 fold less toxic than most insecticides. Although it is known that these synthetic insecticides produce health and environmental problems, it is important to highlight that the effect of natural compounds is significantly lower, so they may not be widely accepted by rural producers. However, alternatives could be implemented such as the formulation of synergistic mixtures between EOs, or their more bioactive components, and traditional synthetic insecticides, in order to reduce their applied concentrations.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Population Prospects: The 2019 Revision; United Nations Department of Economic and Social Affairs: New York, NY, USA, 2019. Available online: https://www.un.org/development/desa/pd/news/world-population-prospects-2019-0 (accessed on 25 October 2022).
- van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A Meta-Analysis of Projected Global Food Demand and Population at Risk of Hunger for the Period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- United States Departament of Agriculture. Foreign Agricultural Service. World Agricultural Production; United States Departament of Agriculture: Washington, DC, USA, 2022. Available online: https://apps.fas.usda.gov/psdonline/circulars/production.pdf (accessed on 25 October 2022).
- Pimentel, D. Integrated Pest Management: Innovation-Development Process. In Integrated Pest Management; Peshin, R., Dhawan, A.K., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 1, pp. 83–87. ISBN 978-1-4020-8991-6. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide Pesticide Usage and Its Impacts on Ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Andrews, K.L. The Wholworm Spodoptera frugiperda in Central America and Nerghboring Areas. Fla. Entomol. 1980, 63, 456–467. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular Methods to Detect Spodoptera frugiperda in Ghana, and Implications for Monitoring the Spread of Invasive Species in Developing Countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Q.; Zhang, H.; Wu, K. Spread of Invasive Migratory Pest Spodoptera frugiperda and Management Practices throughout China. J. Integr. Agric. 2021, 20, 637–645. [Google Scholar] [CrossRef]
- Shylesha, A.N.; Jalali, S.K.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K.; et al. Studies on New Invasive Pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) and Its Natural Enemies. J. Biol. Control 2018, 32, 145–151. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First Report of Outbreaks of the Fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a New Alien Invasive Pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- Deshmukh, S.; Pavithra, H.B.; Kalleshwaraswamy, C.M.; Shivanna, B.K.; Maruthi, M.S.; Mota-Sanchez, D. Field Efficacy of Insecticides for Management of Invasive Fall Armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) on Maize in India. Fla. Entomol. 2020, 103, 221–227. [Google Scholar] [CrossRef]
- Qi, G.J.; Ma, J.; Wan, J.; Ren, Y.L.; McKirdy, S.; Hu, G.; Zhang, Z.F. Source Regions of the First Immigration of Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) Invading Australia. Insects 2021, 12, 1104. [Google Scholar] [CrossRef]
- Lezaun, J. Gusano Cogollero: Una Plaga de Alto Impacto. Available online: https://www.croplifela.org/es/plagas/listado-de-plagas/gusano-cogollero (accessed on 25 October 2022).
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M.R. Recent Developments and Future Prospects in Insect Pest Control in Transgenic Crops. Trends Plant Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef]
- Machado, E.P.; Gerson, G.L.; Führ, F.M.; Zago, S.L.; Marques, L.H.; Santos, A.C.; Nowatzki, T.; Dahmer, M.L.; Omoto, C.; Bernardi, O. Cross-Crop Resistance of Spodoptera frugiperda Selected on Bt Maize to Genetically-Modified Soybean Expressing Cry1Ac and Cry1F Proteins in Brazil. Sci. Rep. 2020, 10, 10080. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, M.M.; Matos, J.M.L.; Santos-Amaya, O.F.; França, J.C.; Gonçalves, J.; Paula-Moraes, S.V.; Guedes, R.N.C.; Pereira, E.J.G. Bt-Toxin Susceptibility and Hormesis-like Response in the Invasive Southern Armyworm (Spodoptera eridania). Crop Prot. 2020, 132, 105129. [Google Scholar] [CrossRef]
- Szwarc, D.E. Dispersión y Mortalidad de Larvas de Spodoptera frugiperda S. (Lepidoptera: Noctuidae) en Híbridos de Maíz Convencional y Transgénico Bt. Potenciales Implicancias para el Manejo de la Resistencia. Master’s Thesis, Universidad Nacional de La Plata, La Plata, Argentina, 2018. [Google Scholar]
- Téllez-Rodríguez, P.; Raymond, B.; Morán-Bertot, I.; Rodríguez-Cabrera, L.; Wright, D.J.; Borroto, C.G.; Ayra-Pardo, C. Strong Oviposition Preference for Bt over Non-Bt Maize in Spodoptera frugiperda and Its Implications for the Evolution of Resistance. BMC Biol. 2014, 12, 48. [Google Scholar] [CrossRef]
- Santos-Amaya, O.F.; Rodrigues, J.V.C.; Souza, T.C.; Tavares, C.S.; Campos, S.O.; Guedes, R.N.C.; Pereira, E.J.G. Resistance to Dual-Gene Bt Maize in Spodoptera frugiperda: Selection, Inheritance, and Cross-Resistance to Other Transgenic Events. Sci. Rep. 2015, 5, 18243. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, D.; Salmeron, E.; Horikoshi, R.J.; Bernardi, O.; Dourado, P.M.; Carvalho, R.A.; Martinelli, S.; Head, G.P.; Omoto, C. Cross-Resistance between Cry1 Proteins in Fall Armyworm (Spodoptera frugiperda) May Affect the Durability of Current Pyramided Bt Maize Hybrids in Brazil. PLoS ONE 2015, 10, e0140130. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.A.; Chiaravalle, W.; Dalla-Rizza, M.; Farias, J.R.; García-Degano, M.F.; Gastaminza, G.; Mota-Sánchez, D.; Murúa, M.G.; Omoto, C.; Pieralisi, B.K.; et al. Current Situation of Pests Targeted by Bt Crops in Latin America. Curr. Opin. Insect Sci. 2016, 15, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Souza Ribas, N.; McNeil, J.N.; Días Araújo, H.; Souza Ribas, B.; Lima, E. The Effect of Resistance to Bt Corn on the Reproductive Output of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- Jacques, F.L.; Degrande, P.E.; Gauer, E.; Malaquias, J.B.; Scoton, A.M.N. Intercropped Bt and Non-Bt Corn with Ruzigrass (Urochloa ruziziensis) as a Tool to Resistance Management of Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae). Pest Manag. Sci. 2021, 77, 3372–3381. [Google Scholar] [CrossRef]
- Hanif, M.A.; Nisar, S.; Khan, G.S.; Mushtaq, Z.; Zubair, M. Essential Oils. In Essential Oil Research; Malik, S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 3–17. ISBN 9783030165468. [Google Scholar]
- Dambolena, J.S.; Gallucci, M.N.; Luna, A.; Gonzalez, S.B.; Guerra, P.E.; Zunino, M.P. Composition, Antifungal and Antifumonisin Activity of Pinus wallichiana, Pinus monticola and Pinus strobus Essential Oils from Patagonia Argentina. J. Essent. Oil Bear. Plants 2016, 19, 1769–1775. [Google Scholar] [CrossRef]
- Pizzolitto, R.P.; Jacquat, A.G.; Usseglio, V.L.; Achimón, F.; Cuello, A.E.; Zygadlo, J.A.; Dambolena, J.S. Quantitative-Structure-Activity Relationship Study to Predict the Antifungal Activity of Essential Oils against Fusarium verticillioides. Food Control 2020, 108, 106836. [Google Scholar] [CrossRef]
- Brito, V.D.; Achimón, F.; Pizzolitto, R.P.; Ramírez Sánchez, A.; Gómez Torres, E.A.; Zygadlo, J.A.; Zunino, M.P. An Alternative to Reduce the Use of the Synthetic Insecticide against the Maize Weevil Sitophilus zeamais through the Synergistic Action of Pimenta racemosa and Citrus sinensis Essential Oils with Chlorpyrifos. J. Pest Sci. 2021, 94, 409–421. [Google Scholar] [CrossRef]
- Arena, J.S.; Peschiutta, M.L.; Calvimonte, H.; Zygadlo, J.A. Fumigant and Repellent Activities of Different Essential Oils Alone and Combined Against the Maize Weevil (Sitophilus zeamais Motschulsky). MOJ Bioorg. Org. Chem. 2017, 1, 249–253. [Google Scholar] [CrossRef][Green Version]
- Rizvi, S.A.H.; Ling, S.; Zeng, X. Seriphidium brevifolium Essential Oil: A Novel Alternative to Synthetic Insecticides against the Dengue Vector Aedes albopictus. Environ. Sci. Pollut. Res. 2020, 27, 31863–31871. [Google Scholar] [CrossRef]
- Cantó-Tejero, M.; Pascual-Villalobos, M.J.; Guirao, P. Aniseed Essential Oil Botanical Insecticides for the Management of the Currant-Lettuce Aphid. Ind. Crop. Prod. 2022, 181, 114804. [Google Scholar] [CrossRef]
- Achimón, F.; Peschiutta, M.L.; Brito, V.D.; Beato, M.; Pizzolitto, R.P.; Zygadlo, J.A.; Zunino, M.P. Exploring Contact Toxicity of Essential Oils against Sitophilus zeamais through a Meta-Analysis Approach. Plants 2022, 11, 3070. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Spochacz, M.; Chowański, S.; Walkowiak-Nowicka, K.; Szymczak, M.; Adamski, Z. Plant-Derived Substances Used against Beetles–Pests of Stored Crops and Food–and Their Mode of Action: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1339–1366. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Jalali Sendi, J. A Review on Recent Research Results on Bio-Effects of Plant Essential Oils against Major Coleopteran Insect Pests. Toxin Rev. 2015, 34, 76–91. [Google Scholar] [CrossRef]
- Mossa, A.T.H. Green Pesticides: Essential Oils as Biopesticides in Insect-Pest Management. J. Environ. Sci. Technol. 2016, 9, 354–378. [Google Scholar] [CrossRef]
- Benelli, G.; Maggi, F. Insecticidal Activity of Plant Secondary Metabolites. Plants 2022, 11, 2804. [Google Scholar] [CrossRef]
- Nerio, L.S.; Olivero-Verbel, J.; Stashenko, E. Repellent Activity of Essential Oils: A Review. Bioresour. Technol. 2010, 101, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Giller, K.E.; Delaune, T.; Silva, J.V.; Descheemaeker, K.; van de Ven, G.; Schut, A.G.T.; van Wijk, M.; Hammond, J.; Hochman, Z.; Taulya, G.; et al. The Future of Farming: Who Will Produce Our Food? Food Secur. 2021, 13, 1073–1099. [Google Scholar] [CrossRef]
- OECD-FAO. Perspectivas para el Medio Ambiente. In Agricultura Mundial: Hacia Los Años 2015/2030; Food and Agriculture Organization: Washington, DC, USA, 2015. [Google Scholar]
- Devrjna, N.; Milutinović, M.; Savić, J. When Scent Becomes a Weapon- Plant Essential Oils as Potent Bioinsecticides. Sustainability 2022, 14, 5847. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Stepnowski, P. Chemical Composition of Insect Surface Waxes: Biological Functions and Analytics. In Handbook of Bioanalytics; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–19. [Google Scholar]
- Stamm, K.; Saltin, B.D.; Dirks, J.H. Biomechanics of Insect Cuticle: An Interdisciplinary Experimental Challenge. Appl. Phys. A Mater. Sci. Process. 2021, 127, 329. [Google Scholar] [CrossRef]
- Singh, S.; Chaurasia, P.; Bharati, S.; Golla, U. A Mini-Review on the Safety Profile of Essential Oils. MOJ Biol. Med. 2022, 7, 33–36. [Google Scholar] [CrossRef]
- Dutra, K.; Wanderley-Teixeira, V.; Guedes, C.; Cruz, G.; Navarro, D.; Monteiro, A.; Agra, A.; Lapa Neto, C.; Teixeira, Á. Toxicity of Essential Oils of Leaves of Plants from the Genus Piper with Influence on the Nutritional Parameters of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). J. Essent. Oil Bear. Plants 2020, 23, 213–229. [Google Scholar] [CrossRef]
- Gad, H.A.; Hamza, A.F.; Abdelgaleil, S.A.M. Chemical Composition and Fumigant Toxicity of Essential Oils from Ten Aromatic Plants Growing in Egypt against Different Stages of Confused Flour Beetle, Tribolium confusum Jacquelin Du Val. Int. J. Trop. Insect Sci. 2022, 42, 697–706. [Google Scholar] [CrossRef]
- Renoz, F.; Demeter, S.; Degand, H.; Nicolis, S.C.; Lebbe, O.; Martin, H.; Deneubourg, J.L.; Fauconnier, M.L.; Morsomme, P.; Hance, T. The Modes of Action of Mentha arvensis Essential Oil on the Granary Weevil Sitophilus granarius Revealed by a Label-Free Quantitative Proteomic Analysis. J. Pest Sci. 2022, 95, 381–395. [Google Scholar] [CrossRef]
- Gerolt, P. Mode of Entry of Contact. J. Insect Physiol. 1969, 15, 563–580. [Google Scholar] [CrossRef]
- Sugiura, M.; Horibe, Y.; Kawada, H.; Takagi, M. Insect Spiracle as the Main Penetration Route of Pyrethroids. Pestic. Biochem. Physiol. 2008, 91, 135–140. [Google Scholar] [CrossRef]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect Cuticle: A Critical Determinant of Insecticide Resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N.; Ortiz-Urquiza, A.; Huarte-Bonnet, C.; Zhang, S.; Keyhani, N.O. Targeting of Insect Epicuticular Lipids by the Entomopathogenic Fungus Beauveria bassiana: Hydrocarbon Oxidation within the Context of a Host-Pathogen Interaction. Front. Microbiol. 2013, 4, 24. [Google Scholar] [CrossRef] [PubMed]
- Karunaratne, P.; De Silva, P.; Weeraratne, T.; Surendran, N. Insecticide Resistance in Mosquitoes: Development, Mechanisms and Monitoring. Ceylon J. Sci. 2018, 47, 299. [Google Scholar] [CrossRef]
- Tak, J.H.; Isman, M.B. Enhanced Cuticular Penetration as the Mechanism for Synergy of Insecticidal Constituents of Rosemary Essential Oil in Trichoplusia Ni. Sci. Rep. 2015, 5, 12690. [Google Scholar] [CrossRef] [PubMed]
- Mwamburi, L.A. Role of Plant Essential Oils in Pest Management. In New and Future Development in Biopesticide Research: Biotechnological Exploration; De Mandal, S., Ramkuamar, G., Karthi, S., Jin, F., Eds.; Springer: Singapore, 2022; ISBN 9789811639883. [Google Scholar]
- Papachristos, D.P.; Stamopoulos, D.C. Repellent, Toxic and Reproduction Inhibitory Effects of Essential Oil Vapours on Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res. 2002, 38, 117–128. [Google Scholar] [CrossRef]
- Programa Manejo de Resistencia de Insectos (MRI); Insecticide Resistance Action Committee (IRAC Argentina). Cogollero (Spodoptera frugiperda) en el Cultivo de Maíz: Bases para su Manejo y Control en Sistemas de Producción; REM AAPRESID: Rosario, Argentina, 2019; ISBN 9788578110796. [Google Scholar]
- Ianonne, N.; Leiva, P. Guia Practica para el Cultivo de Maiz; Instituto Nacional de Tecnología Agropecuaria: Buenos Aires, Argentina, 2011. [Google Scholar]
- International Plant Protection Convention (IPPC). Secretariat Prevention, Preparedness and Response Guidelines for Spodoptera frugiperda; International Plant Protection Convention: Rome, Italy, 2021. [Google Scholar]
- LG Semillas. GUÍA DE MANEJO: Spodoptera Frugiperda, Gusano Cogollero en Maíz; LG Semillas: Buenos Aires, Argentina, 2020. [Google Scholar]
- Monteiro, I.N.; Monteiro, O.S.; de Oliveira, A.K.M.; Favero, S.; Garcia, N.Z.T.; Fernandes, Y.M.L.; Jacinto, G.S.S.; Rivero-Wendt, C.L.G.; Matias, R. Chemical Analysis and Insecticidal Activity of Ocimum gratissimum Essential Oil and Its Major Constituent against Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae). Res. Soc. Dev. 2020, 9, e4999119787. [Google Scholar] [CrossRef]
- Cruz, G.S.; Wanderley-Teixeira, V.; Oliveira, J.V.; Lopes, F.S.C.; Barbosa, D.R.S.; Breda, M.O.; Dutra, K.A.; Guedes, C.A.; Navarro, D.M.A.F.; Teixeira, A.A.C. Sublethal Effects of Essential Oils from Eucalyptus staigeriana (Myrtales: Myrtaceae), Ocimum gratissimum (Lamiales: Laminaceae), and Foeniculum vulgare (Apiales: Apiaceae) on the Biology of Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2016, 109, 660–666. [Google Scholar] [CrossRef]
- Cruz, G.S.; Wanderley-Teixeira, V.; da Silva, L.M.; Dutra, K.A.; Guedes, C.A.; de Oliveira, J.V.; Navarro, D.M.A.F.; Araújo, B.C.; Teixeira, Á.A.C. Chemical Composition and Insecticidal Activity of the Essential Oils of Foeniculum vulgare Mill., Ocimum basilicum L., Eucalyptus staigeriana F. Muell. Ex Bailey, Eucalyptus citriodora Hook and Ocimum gratissimum L. and Their Major Components on Spodoptera. J. Essent. Oil Bear. Plants 2017, 20, 1360–1369. [Google Scholar] [CrossRef]
- Aćimović, M.; Rat, M.; Pezo, L.; Lončar, B.; Pezo, M.; Miljković, A.; Lazarević, J. Biological and Chemical Diversity of Angelica archangelica L.—Case Study of Essential Oil and Its Biological Activity. Agronomy 2022, 12, 1570. [Google Scholar] [CrossRef]
- Barra, A. Factors Affecting Chemical Variability of Essential Oils: A Review of Recent Developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef]
- da Cruz, E.N.S.; Peixoto, L.S.; da Costa, J.S.; Mourão, R.H.V.; Do Nascimento, W.M.O.; Maia, J.G.S.; Setzer, W.N.; da Silva, J.K.; Figueiredo, P.L.B. Seasonal Variability of a Caryophyllane Chemotype Essential Oil of Eugenia patrisii Vahl Occurring in the Brazilian Amazon. Molecules 2022, 27, 2417. [Google Scholar] [CrossRef] [PubMed]
- Miladinović, D.L.; Dimitrijević, M.V.; Miladinović, L.C.; Marković, M.S.; Stojanović, G.S. Seasonal Variation in the Essential Oil of Satureja kitaibelii Determines Chemotypes. J. Essent. Oil Res. 2022, 34, 567–575. [Google Scholar] [CrossRef]
- Alves-Silva, J.M.; Piras, A.; Porcedda, S.; Falconieri, D.; Maxia, A.; Gonçalves, M.J.; Cruz, M.T.; Salgueiro, L. Chemical Characterization and Bioactivity of the Essential Oil from Santolina insularis, a Sardinian Endemism. Nat. Prod. Res. 2022, 36, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Servi, H. Chemical Composition and Biological Activities of Essential Oils of Two New Chemotypes of Glebionis Cass. Turk. J. Chem. 2021, 45, 1559–1566. [Google Scholar] [CrossRef]
- De Martino, L.; Amato, G.; Caputo, L.; Nazzaro, F.; Scognamiglio, M.R.; De Feo, V. Variations in Composition and Bioactivity of Ocimum basilicum Cv ‘Aroma 2′ Essential Oils. Ind. Crop. Prod. 2021, 172, 114068. [Google Scholar] [CrossRef]
- Lima, R.K.; Cardoso, M.G.; Moraes, J.C.; Melo, B.A.; Rodrigues, V.G.; Guimarães, P.L. Atividade Inseticida Do Óleo Essencial de Pimenta Longa (Piper hispidinervum C. DC.) Sobre Lagarta-Do-Cartucho Do Milho Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae). Acta Amaz. 2009, 39, 377–382. [Google Scholar] [CrossRef]
- Monteiro, I.N.; Ferreira, L.O.G.; de Oliveira, A.K.M.; Favero, S.; Figueiredo, P.L.B.; Maia, J.G.S.; Monteiro, O.S.; Matias, R. Toxicity of the Lippia gracilis Essential Oil Chemotype, Pinene-Cineole-Limonene, on Spodoptera frugiperda (Lepidoptera: Noctuidae). Int. J. Trop. Insect Sci. 2021, 41, 181–187. [Google Scholar] [CrossRef]
- Knaak, N.; Wiest, S.L.F.; Andreis, T.F.; Fiuza, L.M. Toxicity of Essential Oils to the Larvae of Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Biopestic. 2013, 6, 49–53. [Google Scholar]
- Silva Bibiano, C.; Alves, D.S.; Freire, B.C.; Vilela Bertolucci, S.K.; Carvalho, G.A. Toxicity of Essential Oils and Pure Compounds of Lamiaceae Species against Spodoptera frugiperda (Lepidoptera: Noctuidae) and Their Safety for the Nontarget Organism Trichogramma pretiosum (Hymenoptera: Trichogrammatidae). Crop Prot. 2022, 158, 106011. [Google Scholar] [CrossRef]
- Niculau, E.; Alves, P.B.; Paulo, P.C.; Valéria, V.R.; Matos, A.P.; Bernardo, A.R.; Volante, A.C.; Fernandes, J.B.; Da Silva, M.F.G.F.; Corrêa, A.G.; et al. Atividade Inseticida de Óleos Essenciais de Pelargonium graveolens l’Herit E Lippia alba (Mill) N. E. Brown Sobre Spodoptera frugiperda (J. E. Smith). Quim. Nova 2013, 36, 1391–1394. [Google Scholar] [CrossRef]
- Lima, A.P.S.; Santana, E.D.R.; Santos, A.C.C.; Silva, J.E.; Ribeiro, G.T.; Pinheiro, A.M.; Santos, Í.T.B.F.; Blank, A.F.; Araújo, A.P.A.; Bacci, L. Insecticide Activity of Botanical Compounds against Spodoptera frugiperda and Selectivity to the Predatory Bug Podisus nigrispinus. Crop Prot. 2020, 136, 105230. [Google Scholar] [CrossRef]
- Dutra, K.A.; Wanderley Teixeira, V.; Cruz, G.S.; Silva, C.T.S.; D’Assunção, C.G.; Ferreira, C.G.M.; Monteiro, A.L.B.; Agra Neto, A.C.; Lapa Neto, C.J.C.; Teixeira, A.A.C.; et al. Morphological and Immunohistochemical Study of the Midgut and Fat Body of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) Treated with Essential Oils of the Genus Piper. Biotech. Histochem. 2019, 94, 498–513. [Google Scholar] [CrossRef]
- Negrini, M.; Fidelis, E.G.; Schurt, D.A.; Silva, F.S.; Pereira, R.S.; Bizzo, H.R. Insecticidal Activity of Essential Oils in Controlling Fall Armyworm, Spodoptera frugiperda. Arq. Inst. Biol. 2019, 86, e1112018. [Google Scholar] [CrossRef]
- Silva, S.M.; Rodrigues da Cunha, J.P.A.; de Carvalho, S.M.; Zandonadi, C.H.S.; Martins, R.C.; Chang, R. Ocimum basilicum Essential Oil Combined with Deltamethrin to Improve the Management of Spodoptera frugiperda. Ciênc. Agrotecnol. 2017, 41, 665–675. [Google Scholar] [CrossRef]
- Fazolin, M.; Estrela, J.L.V.; Medeiros, A.F.M.; da Silva, I.M.; Gomes, L.P.; Silva, M.S.F. Potencial Sinérgico Do Óleo Essencial Rico em Dilapiol para Inseticidas Piretroides Sintéticos Frente à Lagarta-Do-Cartucho. Cienc. Rural 2016, 46, 382–388. [Google Scholar] [CrossRef]
- Lourenço, A.M.; Haddi, K.; Ribeiro, B.M.; Corrêia, R.F.T.; Tomé, H.V.V.; Santos-Amaya, O.; Pereira, E.J.G.; Guedes, R.N.C.; Santos, G.R.; Oliveira, E.E.; et al. Essential Oil of Siparuna guianensis as an Alternative Tool for Improved Lepidopteran Control and Resistance Management Practices. Sci. Rep. 2018, 8, 7215. [Google Scholar] [CrossRef]
- Lima, R.K.; Cardoso, M.G.; Moraes, J.C.; Andrade, M.A.; Melo, B.A.; Rodrigues, V.G. Chemical Characterization and Inseticidal Activity of the Essential Oil Leaves of Ageratum conyzoides L. on Fall Armyworm Spodoptera frugiperda (Smith, 1797) (Lepidoptera: Noctuidae). Biosci. J. 2010, 26, 1–5. [Google Scholar]
- Villafañe, E.; Tolosa, D.; Bardón, A.; Neske, A. Toxic Effects of Citrus aurantium and C. limon Essential Oils on Spodoptera frugiperda (Lepidoptera: Noctuidae). Nat. Prod. Commun. 2011, 6, 1389–1392. [Google Scholar] [CrossRef]
- Aguiar, M.C.S.; Denadae, B.E.; Silva, M.F.G.F.; Fernandes, J.B.; Bittencourt, P.R.S.; Scremin, F.R.; Forim, M.R. Quality Control for Lignin and Gelatin Microcapsules Loaded with Orange Essential Oil. J. Braz. Chem. Soc. 2022, 33, 85–95. [Google Scholar] [CrossRef]
- de Menezes, C.W.G.; Carvalho, G.A.; Alves, D.S.; de Carvalho, A.A.; Aazza, S.; de Oliveira Ramos, V.; Pinto, J.E.B.P.; Bertolucci, S.K.V. Biocontrol Potential of Methyl Chavicol for Managing Spodoptera frugiperda (Lepidoptera: Noctuidae), an Important Corn Pest. Environ. Sci. Pollut. Res. 2020, 27, 5030–5041. [Google Scholar] [CrossRef]
- Ávila Murillo, M.C.; Suarez, L.E.; Cerón Salamanca, J.A. Chemical Composition and Insecticidal Properties of Essential Oils of Piper septuplinervium and P. subtomentosum (Piperaceae). Nat. Prod. Commun. 2014, 9, 1527–1530. [Google Scholar] [CrossRef]
- Németh-Zámbori, É. Natural Variability of Essential Oil Components. In Handbook of Essential Oils; Husnu Can Baser, K., Buchbauer, G., Eds.; Taylor & Francis Group: Boca Raton, FL, USA, 2020; p. 40. ISBN 9781351246460. [Google Scholar]
- Ilardi, V.; Badalamenti, N.; Bruno, M. Chemical Composition of the Essential Oil from Different Vegetative Parts of Foeniculum vulgare Subsp. Piperitum (Ucria) Coutinho (Umbelliferae) Growing Wild in Sicily. Nat. Prod. Res. 2022, 36, 3587–3597. [Google Scholar] [CrossRef] [PubMed]
- Chalchat, J.C.; Garry, R.P.; Muhayimana, A. Essential Oil of Tagetes Minuta from Rwanda and France: Chemical Composition According to Harvesting Location, Growth Stage and Part of Plant Extracted. J. Essent. Oil Res. 1995, 7, 375–386. [Google Scholar] [CrossRef]
- Guedes, C.A.; Teixeira, V.W.; Dutra, K.A.; Navarro, D.M.A.F.; Cruz, G.S.; Lapa Neto, C.J.C.; Correia, A.A.; Sandes, J.M.; Brayner, F.A.; Alves, L.C.; et al. Evaluation of Piper marginatum (Piperales: Piperaceae) Oil and Geraniol on the Embryonic Development of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Comparison to Formulated Products. J. Econ. Entomol. 2020, 113, 239–248. [Google Scholar] [CrossRef]
- Alkan, M.; Erturk, S. Insecticidal Efficacy and Repellency of Trans-Anethole against Four Stored-Product Insect Pests. J. Agric. Sci. 2020, 26, 64–70. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, Y.; Sabier, M.; Zhang, T.; Deng, J.; Song, X.; Liao, Z.; Li, Q.; Yang, S.; Cao, Y.; et al. Trans-Anethole Is a Potent Toxic Fumigant That Partially Inhibits Rusty Grain Beetle (Cryptolestes ferrugineus) Acetylcholinesterase Activity. Ind. Crop. Prod. 2021, 161, 113207. [Google Scholar] [CrossRef]
- Johnson, A.J.; Venukumar, V.; Varghese, T.S.; Viswanathan, G.; Leeladevi, P.S.; Remadevi, R.K.S.; Baby, S. Insecticidal Properties of Clausena austroindica Leaf Essential Oil and Its Major Constituent, Trans-anethole, against Sitophilus oryzae and Tribolium castaneum. Ind. Crop. Prod. 2022, 182, 114854. [Google Scholar] [CrossRef]
- Pour, S.A.; Shahriari, M.; Zibaee, A.; Mojarab-Mahboubkar, M.; Sahebzadeh, N.; Hoda, H. Toxicity, Antifeedant and Physiological Effects of Trans-anethole against Hyphantria cunea Drury (Lep: Arctiidae). Pestic. Biochem. Physiol. 2022, 185, 105135. [Google Scholar] [CrossRef]
- Silva, S.M.; Da Cunha, J.P.A.R.; Zandonadi, C.H.S.; De Assunção, H.H.T.; Marques, M.G. Synergistic Effects of Binary Mixtures of Linalool with Pyrethroids against Fall Armyworm. Biosci. J. 2020, 36, 228–237. [Google Scholar] [CrossRef]
- Zamani Fard, S.; Hemmati, S.A.; Shishehbor, P.; Stelinski, L.L. Growth, Consumption and Digestive Enzyme Activities of Spodoptera littoralis (Boisd) on Various Mung Bean Cultivars Reveal Potential Tolerance Traits. J. Appl. Entomol. 2022, 146, 1145–1154. [Google Scholar] [CrossRef]
- Xia, X.; Lan, B.; Tao, X.; Lin, J.; You, M. Characterization of Spodoptera litura Gut Bacteria and Their Role in Feeding and Growth of the Host. Front. Microbiol. 2020, 11, 1492. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, M.; Ullah, F.; Khan, M.M.; Li, X.; Zhang, Z.; Shah, S.; Imran, M.; Assiri, M.A.; Fernández-Grandon, G.M.; Desneux, N.; et al. Metabolic-Based Insecticide Resistance Mechanism and Ecofriendly Approaches for Controlling of Beet Armyworm Spodoptera exigua: A Review. Environ. Sci. Pollut. Res. 2022, 29, 1746–1762. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Beato, M.; Usseglio, V.L.; Camina, J.; Zygadlo, J.A.; Dambolena, J.S.; Zunino, M.P. Phenolic Compounds as Controllers of Sitophilus zeamais: A Look at the Structure-Activity Relationship. J. Stored Prod. Res. 2022, 99, 102038. [Google Scholar] [CrossRef]
- Zaio, Y.P.; Gatti, G.; Ponce, A.; Saavedra Larralde, N.A.; Martinez, M.J.; Zunino, M.P.; Zygadlo, J.A. Cinnamaldehyde and Related Phenylpropanoids, Natural Repellents, and Insecticides against Sitophilus zeamais (Motsch.). A Chemical Structure-Bioactivity Relationship. J. Sci. Food Agric. 2018, 98, 5822–5831. [Google Scholar] [CrossRef]
- Pang, Y.P. Insect Acetylcholinesterase as a Target for Effective and Environmentally Safe Insecticides, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; Volume 46, ISBN 9780124170100. [Google Scholar]
- Abubakar, M.U.; Abubakar, D. Characterization of Acetylcholinesterase from Various Sources: A Mini Review. J. Environ. Bioremediat. Toxicol. 2021, 4, 24–30. [Google Scholar] [CrossRef]
- Chaudhari, A.K.; Singh, V.K.; Kedia, A.; Das, S.; Dubey, N.K. Essential Oils and Their Bioactive Compounds as Eco-Friendly Novel Green Pesticides for Management of Storage Insect Pests: Prospects and Retrospects. Environ. Sci. Pollut. Res. 2021, 28, 18918–18940. [Google Scholar] [CrossRef]
- de Oliveira, E.R.; Alves, D.S.; Carvalho, G.A.; de Oliveira, B.M.R.G.; Aazza, S.; Bertolucci, S.K.V. Toxicity of Cymbopogon flexuosus Essential Oil and Citral for Spodoptera frugiperda. Ciênc. Agrotecnol. 2018, 42, 408–419. [Google Scholar] [CrossRef]
- Herrera, J.M.; Goñi, M.L.; Gañan, N.A.; Zygadlo, J.A. An Insecticide Formulation of Terpene Ketones against Sitophilus zeamais and Its Incorporation into Low Density Polyethylene Films. Crop Prot. 2017, 98, 33–39. [Google Scholar] [CrossRef]
- Goñi, M.L.; Gañán, N.A.; Herrera, J.M.; Strumia, M.C.; Andreatta, A.E.; Martini, R.E. Supercritical CO2 Iof LDPE Films with Terpene Ketones as Biopesticides against Corn Weevil (Sitophilus zeamais). J. Supercrit. Fluids 2017, 122, 18–26. [Google Scholar] [CrossRef]
- Herrera, J.M.; Zunino, M.P.; Dambolena, J.S.; Pizzolitto, R.P.; Gañan, N.A.; Lucini, E.I.; Zygadlo, J.A. Terpene Ketones as Natural Insecticides against Sitophilus zeamais. Ind. Crop. Prod. 2015, 70, 435–442. [Google Scholar] [CrossRef]
- Peschiutta, M.L.; Brito, V.D.; Achimón, F.; Zunino, M.P.; Usseglio, V.L.; Zygadlo, J.A. New Insecticide Delivery Method for the Control of Sitophilus zeamais in Stored Maize. J. Stored Prod. Res. 2019, 83, 185–190. [Google Scholar] [CrossRef]
- Xie, F.; Rizvi, S.A.H.; Zeng, X. Fumigant Toxicity and Biochemical Properties of (α + β) Thujone and 1,8-Cineole Derived from Seriphidium brevifolium Volatile Oil against the Red Imported Fire Ant Solenopsis invicta (Hymenoptera: Formicidae). Rev. Bras. Farmacogn. 2019, 29, 720–727. [Google Scholar] [CrossRef]
- Ruttanaphan, T.; Bullangpoti, V. The Potential Use of Thymol and (R)-(+)-Pulegone as Detoxifying Enzyme Inhibitors against Spodoptera litura (Lepidoptera: Noctuidae). Phytoparasitica 2022, 50, 913–920. [Google Scholar] [CrossRef]
- Sousa, P.A.S.; Neto, J.; Bastos, M.M.S.M.; Aguiar, A.A.R.M. Eugenol and Pulegone as Potential Biorational Alternatives for Trioza erytreae (Hemiptera: Triozidae) Control: Preliminary Results on Nymphal Toxicity and Applicability on Citrus limon. J. Nat. Pestic. Res. 2022, 1, 100004. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, J.; Zhang, N.; Yu, W.; Jiang, J.; Dai, G. Insecticidal Activities of Salvia hispanica L. Essential Oil and Combinations of Their Main Compounds against the Beet Armyworm Spodoptera exigua. Ind. Crop. Prod. 2021, 162, 113271. [Google Scholar] [CrossRef]
- Pérez-Garrido, A.; Helguera, A.M.; Rodríguez, F.G.; Cordeiro, M.N.D.S. QSAR Models to Predict Mutagenicity of Acrylates, Methacrylates and α,β-Unsaturated Carbonyl Compounds. Dent. Mater. 2010, 26, 397–415. [Google Scholar] [CrossRef] [PubMed]
- Lucia, A.; Guzmán, E. Emulsions Containing Essential Oils, Their Components or Volatile Semiochemicals as Promising Tools for Insect Pest and Pathogen Management. Adv. Colloid Interface Sci. 2021, 287, 102330. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review. Molecules 2018, 23, 34. [Google Scholar] [CrossRef]
- Basicmedical Key. Available online: https://basicmedicalkey.com/acetylcholine-receptor-agonists-2/%0A (accessed on 23 September 2022).
- Fergani, Y.; Elbanna, H.; Hamama, H. Genotoxicity of Some Plant Essential Oils in Cotton Leafworm, Spodoptera littoralis (Lepidoptera: Noctuidae): The Potential Role of Detoxification Enzymes. Egypt. J. Zool. 2020, 73, 53–66. [Google Scholar] [CrossRef]
- El-Aw, M.; Draz, K.; El Naggar, E.; Abd Elsalam, F. Ovicidal, Larvicidal and Biochemical Effects of Thyme, Thymus vulgaris, on the Cotton Leafworm, Spodoptera littoralis (Boisd.). J. Plant Prot. Pathol. 2021, 12, 389–394. [Google Scholar] [CrossRef]
- Shahriari, M.; Zibaee, A.; Sahebzadeh, N.; Shamakhi, L. Effects of α-Pinene, Trans-Anethole, and Thymol as the Essential Oil Constituents on Antioxidant System and Acetylcholine Esterase of Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). Pestic. Biochem. Physiol. 2018, 150, 40–47. [Google Scholar] [CrossRef]
- Bloomquist, J.R. GABA and Glutamate Receptors as Biochemical Sites for Insecticide Action. In Biochemical Sites of Insecticide Action and Resistance; Ishaaya, I., Ed.; Springerg: Berlin/Heidelberg, Germany, 2001; pp. 17–41. [Google Scholar]
- Mustard, J.A.; Jones, L.; Wright, G.A. GABA Signaling Affects Motor Function in the Honey Bee. J. Insect Physiol. 2020, 120, 103989. [Google Scholar] [CrossRef] [PubMed]
- Urtasum, M.A.; Mordujovich-buschiazzo, P.; Cañás, M. Benzodiazepinas: Uso Crónico y Deprescripción. Folia Doc. 2021, 2, 1–12. [Google Scholar]
- Ozoe, Y.; Akamatsu, M. Non-Competitive GABA Antagonists: Probing the Mechanisms of Their Selectivity for Insect versus Mammalian Receptors. Pest Manag. Sci. 2001, 57, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Zhan, E.L.; Wang, Y.; Jiang, J.; Jia, Z.Q.; Tang, T.; Song, Z.J.; Han, Z.J.; Zhao, C.Q. Influence of Three Insecticides Targeting GABA Receptor on Fall Armyworm Spodoptera frugiperda: Analyses from Individual, Biochemical and Molecular Levels. Pestic. Biochem. Physiol. 2021, 179, 104973. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.M.; Gad, H.A.; Ramadan, G.R.M.; El-Bakry, A.M.; El-Sabrout, A.M. Monoterpenes: Chemistry, Insecticidal Activity against Stored Product Insects and Modes of Action—A Review. Int. J. Pest Manag. 2021, 1–23. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.M.; Badawy, M.E.I.; Mahmoud, N.F.; Marei, A.S.M. Acaricidal Activity, Biochemical Effects and Molecular Docking of Some Monoterpenes against Two-Spotted Spider Mite (Tetranychus urticae Koch). Pestic. Biochem. Physiol. 2019, 156, 105–115. [Google Scholar] [CrossRef]
- Cui, G.; Yuan, H.; He, W.; Deng, Y.; Sun, R.; Zhong, G. Synergistic Effects of Botanical Curcumin-Induced Programmed Cell Death on the Management of Spodoptera litura Fabricius with Avermectin. Ecotoxicol. Environ. Saf. 2022, 229, 113097. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, H.; Zhang, Y.; Chen, J.; Upadhyay, A.; Bhowmick, B.; Hang, J.; Wu, S.; Liao, C.; Han, Q. Functional Analysis Reveals Ionotropic GABA Receptor Subunit RDL Is a Target Site of Ivermectin and Fluralaner in the Yellow Fever Mosquito, Aedes aegypti. Pest Manag. Sci. 2022, 78, 4173–4182. [Google Scholar] [CrossRef]
- Taktak, N.E.M.; Badawy, M.E.I.; Awad, O.M.; Abou El-Ela, N.E. Nanoemulsions Containing Some Plant Essential Oils as Promising Formulations against Culex pipiens (L.) Larvae and Their Biochemical Studies. Pestic. Biochem. Physiol. 2022, 185, 105151. [Google Scholar] [CrossRef]
- Taillebois, E.; Thany, S.H. The Use of Insecticide Mixtures Containing Neonicotinoids as a Strategy to Limit Insect Pests: Efficiency and Mode of Action. Pestic. Biochem. Physiol. 2022, 184, 105126. [Google Scholar] [CrossRef] [PubMed]
- Gad, H.A.; Ramadan, G.R.M.; El-Bakry, A.M.; El-Sabrout, A.M.; Abdelgaleil, S.A.M. Monoterpenes: Promising Natural Products for Public Health Insect Control—A Review. Int. J. Trop. Insect Sci. 2022, 42, 1059–1075. [Google Scholar] [CrossRef]
- Classen, D.E.; Arnason, J.T.; Serratos, J.A.; Lambert, J.D.H.; Nozzolillo, C.; Philogene, B.J.R. Correlation of Phenolic Acid Content of Maize to Resistance To Sitophilus zeamais, the Maize Weevil, in CIMMYT’S Collections. J. Chem. Ecol. 1990, 16, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.N.; Brent, C.S.; Breed, M.D. Octopamine and Tyramine Modulate the Thermoregulatory Fanning Response in Honey Bees (Apis mellifera). J. Exp. Biol. 2017, 220, 1925–1930. [Google Scholar] [CrossRef] [PubMed]
- David, J.; Lafon-cazal, M. Octopamine Distribution in the Locusta migratoria Nervous and Non-Nervous, Systems. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1979, 64, 161–164. [Google Scholar] [CrossRef]
- Evans, P.D.; Talamo, B.R.; Kravitz, E.A. Octopamine Neurons: Morphology, Release of Octopamine and Possible Physiological Role. Brain Res. 1975, 90, 340–347. [Google Scholar] [CrossRef]
- Evans, P.D. Octopamine: A High-Affinity Uptake Mechanism in the Nervous System of the Cockroach. J. Neurochem. 1978, 30, 1015–1022. [Google Scholar] [CrossRef]
- Fussnecker, B.L.; Smith, B.H.; Mustard, J.A. Octopamine and Tyramine Influence the Behavioral Profile of Locomotor Activity in the Honey Bee (Apis mellifera). J. Insect Physiol. 2006, 52, 1083–1092. [Google Scholar] [CrossRef]
- Selcho, M.; Pauls, D. Linking Physiological Processes and Feeding Behaviors by Octopamine. Curr. Opin. Insect Sci. 2019, 36, 125–130. [Google Scholar] [CrossRef]
- Wamberg, C.; Christensen, S.A.; Jakobsen, I. Interaction between Foliar-Feeding Insects, Mycorrhizal Fungi, and Rhizosphere Protozoa on Pea Plants. Pedobiologia 2003, 47, 281–287. [Google Scholar] [CrossRef]
- Ubuka, T. Octopamine. In Handbook of Hormones; Ando, H., Ukena, K., Nagata, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1045–1047. [Google Scholar]
- Yan, T.K.; Asari, A.; Salleh, S.A.; Azmi, W.A. Eugenol and Thymol Derivatives as Antifeedant Agents against Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) Larvae. Insects 2021, 12, 551. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, M.; Lapied, B.; Jankowski, W.; Stankiewicz, M. The Unusual Action of Essential Oil Component, Menthol, in Potentiating the Effect of the Carbamate Insecticide, Bendiocarb. Pestic. Biochem. Physiol. 2019, 158, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Mohanny, K.; Mohamed, G.; Allam, R. Efficacy of Some Promising Plant Essential Oils to Control the Red Palm Weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae) under Laboratory Conditions. SVU-Int. J. Agric. Sci. 2019, 1, 12–45. [Google Scholar] [CrossRef]
- Toledo, P.F.S.; Viteri Jumbo, L.O.; Rezende, S.M.; Haddi, K.; Silva, B.A.; Mello, T.S.; Della Lucia, T.M.C.; Aguiar, R.W.S.; Smagghe, G.; Oliveira, E.E. Disentangling the Ecotoxicological Selectivity of Clove Essential Oil against Aphids and Non-Target Ladybeetles. Sci. Total Environ. 2020, 718, 137328. [Google Scholar] [CrossRef]
- Ahmed, M.A.I.; Vogel, C.F.A. The Synergistic Effect of Octopamine Receptor Agonists on Selected Insect Growth Regulators on Culex quinquefasciatus Say (Diptera: Culicidae) Mosquitoes. One Health 2020, 10, 100138. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, T.; Ahmed, N.; Shahjeer, K.; Ahmed, S.; Awadh Al-Mutairi, K.; Fathy Khater, H.; Fathey Ali, R. Botanical Insecticides and Their Potential as Anti-Insect/Pests: Are They Successful against Insects and Pests? In Global Decline of Insects; El-Shafie, H.A.F., Ed.; IntechOpen: London, UK, 2022. [Google Scholar]
- Castillo-Morales, R.M.; Carreño Otero, A.L.; Mendez-Sanchez, S.C.; Da Silva, M.A.N.; Stashenko, E.E.; Duque, J.E. Mitochondrial Affectation, DNA Damage and AChE Inhibition Induced by Salvia officinalis Essential Oil on Aedes aegypti Larvae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 221, 29–37. [Google Scholar] [CrossRef]
- Yooboon, T.; Kuramitsu, K.; Bullangpoti, V.; Kainoh, Y.; Furukawa, S. Cytotoxic Effects of β-Asarone on Sf9 Insect Cells. Arch. Insect Biochem. Physiol. 2019, 102, e21596. [Google Scholar] [CrossRef]
- Olmedo, R.; Herrera, J.M.; Lucini, E.I.; Zunino, M.P.; Pizzolitto, R.P.; Dambolena, J.S.; Zygadlo, J.A. Essential Oil of Tagetes filifolia against the Flour Beetle Tribolium castaneum and Its Relation to Acetylcholinesterase Activity and Lipid Peroxidation. Agriscientia 2015, 32, 113–121. [Google Scholar] [CrossRef]
- Moustafa, M.; Awad, M.; Amer, A.; Hassan, N.; Ibrahim, E.; Ali, H.; Akrami, M.; Salem, M. Insecticidal Activity of Lemongrass Essential Oil as an Eco-Friendly Agent against the Black Cutworm Agrotis ipsilon (Lepidoptera: Noctuidae). Insects 2021, 12, 737. [Google Scholar] [CrossRef]
- Chintalchere, J.M.; Dar, M.A.; Shaha, C.; Pandit, R.S. Impact of Essential Oils on Musca domestica Larvae: Oxidative Stress and Antioxidant Responses. Int. J. Trop. Insect Sci. 2021, 41, 821–830. [Google Scholar] [CrossRef]
- Ibrahim, S.S. Essential Oil Nanoformulations as a Novel Method for Insect Pest Control in Horticulture. In Horticultural Crops; Baimey, H.K., Hamamouch, N., Kolimbia, Y.A., Eds.; IntechOpen: London, UK, 2020; pp. 195–209. ISBN 978-1-83880437-4. [Google Scholar]
- Lemus de la Cruz, A.; Barrera-Cortés, J.; Lina-García, L.P.; Ramos-Valdivia, A.C.; Santillán, R. Nanoemulsified Formulation of Cedrela Odorata Essential Oil and Its Larvicidal Effect against Spodoptera frugiperda (J.E. Smith). Molecules 2022, 27, 2975. [Google Scholar] [CrossRef] [PubMed]
- Dinesh, D.; Murugan, K.; Subramaniam, J.; Paulpandi, M.; Chandramohan, B.; Pavithra, K.; Anitha, J.; Vasanthakumaran, M.; Fraceto, L.F.; Wang, L.; et al. Salvia leucantha Essential Oil Encapsulated in Chitosan Nanoparticles with Toxicity and Feeding Physiology of Cotton Bollworm Helicoverpa armigera. In Biopesticides; Elsevier: Amsterdam, The Netherlands, 2022; pp. 159–181. [Google Scholar]
- Ibrahim, S.S.; Abou-elseoud, W.S.; Elbehery, H.H.; Hassan, M.L. Chitosan-Cellulose Nanoencapsulation Systems for Enhancing the Insecticidal Activity of Citronella Essential Oil against the Cotton Leafworm Spodoptera littoralis. Ind. Crop. Prod. 2022, 184, 115089. [Google Scholar] [CrossRef]
- Tortorici, S.; Cimino, C.; Ricupero, M.; Musumeci, T.; Biondi, A.; Siscaro, G.; Carbone, C.; Zappala, L. Nanostructured Lipid Carriers of Essential Oils as Potential Tools for the Sustainable Control of Insect Pests. Ind. Crop. Prod. 2022, 181, 114766. [Google Scholar] [CrossRef]
- Bhan, S.; Mohan, L.; Srivastava, C.N. Nanopesticides: A Recent Novel Ecofriendly Approach in Insect Pest Management. J. Entomol. Res. 2018, 42, 263–270. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Fierascu, I.C.; Dinu-Pirvu, C.E.; Fierascu, I.; Paunescu, A. The Application of Essential Oils as a Next-Generation of Pesticides: Recent Developments and Future Perspectives. Z. Fur Naturforsch.—Sect. C J. Biosci. 2020, 75, 183–204. [Google Scholar] [CrossRef]
- Massuh, L.A. Plan de Negocios para La Producción y Comercialización de Aceite Esencial de Tagetes minuta. Master’s Thesis, Universidad Católica de Córdoba, Córdoba Argentina, 2016. [Google Scholar]
- Li, Y.; Fabiano-Tixier, A.S.; Chemat, F. Essential Oils as Synthons for Green Chemistry. In Essential Oils as Reagents in Green Chemistry; Li, Y., Fabiano-Tixier, A.S., Chemat, F., Eds.; Springer: Cham, Switzerland, 2014; pp. 63–71. ISBN 978-3-319-08449-7. [Google Scholar]
- Cipolatti, E.P.; Cerqueira Pinto, M.C.; Oliveira Henriques, R.; Pinto, J.C.C.S.; Manchado de Castro, A.; Guimarāes Freire, D.M.; Manoel, E.A. Enzymes in Green Chemistry: The State of the Art in Chemical Transformations. In Biomass, Biofuels, Biochemicals: Advances in Enzyme Technology; Pandey, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 137–151. ISBN 9780444641144. [Google Scholar]
- Murcia-Meseguer, A.; Alves, T.J.S.; Budia, F.; Ortiz, A.; Medina, P. Insecticidal Toxicity of Thirteen Commercial Plant Essential Oils against Spodoptera exigua (Lepidoptera: Noctuidae). Phytoparasitica 2018, 46, 233–245. [Google Scholar] [CrossRef]
- Polatoğlu, K.; Karakoç, Ö.C.; Yücel Yücel, Y.; Gücel, S.; Demirci, B.; Başer, K.H.C.; Demirci, F. Insecticidal Activity of Edible Crithmum maritimum L. Esse164tial Oil against Coleopteran and Lepidopteran Insects. Ind. Crops Prod. 2016, 89, 383–389. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal Activity of Some Essential Oils against Larvae of Spodoptera littoralis. Fitoterapia 2005, 76, 691–696. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Maggi, F.; Nkuimi Wandjou, J.G.; Yvette Fofie, N.G.B.; Koné-Bamba, D.; Sagratini, G.; Vittori, S.; Caprioli, G. Insecticidal Activity of the Essential Oil and Polar Extracts from Ocimum gratissimum Grown in Ivory Coast: Efficacy on Insect Pests and Vectors and Impact on Non-Target Species. Ind. Crops Prod. 2019, 132, 377–385. [Google Scholar] [CrossRef]
- Badalamenti, N.; Ilardi, V.; Bruno, M.; Pavela, R.; Boukouvala, M.C.; Kavallieratos, N.G.; Maggi, F.; Canale, A.; Benelli, G. Chemical Composition and Broad-Spectrum Insecticidal Activity of the Flower Essential Oil from an Ancient Sicilian Food Plant, Ridolfia segetum. Agriculture 2021, 11, 304. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Canale, A.; Senthil-Nathan, S.; Maggi, F. Not Just Popular Spices! Essential Oils from Cuminum cyminum and Pimpinella anisum Are Toxic to Insect Pests and Vectors without Affecting Non-Target Invertebrates. Ind. Crops Prod. 2018, 124, 236–243. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Bartolucci, F.; Canale, A.; Maggi, F. Origanum syriacum Subsp. Syriacum: From an Ingredient of Lebanese ‘Manoushe’ to a Source of Effective and Eco-Friendly Botanical Insecticides. Ind. Crops Prod. 2019, 134, 26–32. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Ricciutelli, M.; Lupidi, G.; Maggi, F. Efficacy of the Volatile Oil from Water Celery (Helosciadium nodiflorum, Apiaceae) against the Filariasis Vector Culex quinquefasciatus, the Housefly Musca domestica, and the African Cotton Leafworm Spodoptera littoralis. Chem. Biodivers. 2017, 14, e1700376. [Google Scholar] [CrossRef] [PubMed]
- Wandjou, J.G.N.; Baldassarri, C.; Ferrati, M.; Maggi, F.; Pavela, R.; Tsabang, N.; Petrelli, R.; Ricciardi, R.; Desneux, N.; Benelli, G. Essential Oils from Cameroonian Aromatic Plants as Effective Insecticides against Mosquitoes, Houseflies, and Moths. Plants 2022, 11, 2353. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Santini, G.; Fiorini, D.; Sut, S.; Dall’Acqua, S.; Canale, A.; Maggi, F. The Essential Oil from Industrial Hemp (Cannabis sativa L.) by-Products as an Effective Tool for Insect Pest Management in Organic Crops. Ind. Crops Prod. 2018, 122, 308–315. [Google Scholar] [CrossRef]
- Javier, A.M.V.; Ocampo, V.R.; Ceballo, F.A.; Javier, P.A. Insecticidal Activity of Selected Essential Oil Extracts against Common Cutworm, Spodoptera litura Fabricius (Lepidoptera: Noctuidae). Philipp. J. Sci. 2017, 146, 247–256. [Google Scholar]
- Kim, S.; Yoon, J.; Tak, J.H. Synergistic Mechanism of Insecticidal Activity in Basil and Mandarin Essential Oils against the Tobacco Cutworm. J. Pest Sci. 2021, 94, 1119–1131. [Google Scholar] [CrossRef]
- Pavela, R. Acute, Synergistic and Antagonistic Effects of Some Aromatic Compounds on the Spodoptera littoralis Boisd. (Lep., Noctuidae) Larvae. Ind. Crops Prod. 2014, 60, 247–258. [Google Scholar] [CrossRef]
- Abbassy, M.A.; Abdelgaleil, S.A.M.; Rabie, R.Y.A. Insecticidal and Synergistic Effects of Majorana hortensis Essential Oil and Some of Its Major Constituents. Entomol. Exp. Appl. 2009, 131, 225–232. [Google Scholar] [CrossRef]
- Isman, M.B. Plant Essential Oils for Pest and Disease Management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Family | Occurrence |
|---|---|
| Piperaceae | 13 |
| Lamiaceae | 12 |
| Verbenaceae | 10 |
| Myrtaceae | 5 |
| Asteraceae | 5 |
| Rutaceae | 3 |
| Poaceae | 3 |
| Zingiberaceae | 2 |
| Apiaceae | 1 |
| Siparunaceae | 1 |
| Geraniaceae | 1 |
| Total | 56 |
| Genera | Occurrence in Literature |
|---|---|
| Piper | 13 |
| Ocimum | 9 |
| Lippia | 9 |
| Eucalyptus | 5 |
| Hyptis | 3 |
| Cymbopogon | 3 |
| Foeniculum | 2 |
| Corymbia | 2 |
| Citrus | 2 |
| Siparuna | 1 |
| Ruta | 1 |
| Pelargonium | 1 |
| Mentha | 1 |
| Malva | 1 |
| Hyptis | 1 |
| Eremanthus | 1 |
| Tanacetum | 1 |
| Artemisia | 1 |
| Ageratum | 1 |
| Zingiber | 1 |
| Vanillosmopsis | 1 |
| Total | 60 |
| Plant Species | Occurrences in the Literature |
|---|---|
| Ocimum basilicum | 4 |
| Lippia alba | 3 |
| Piper marginatum | 3 |
| Corymbia citriodora | 2 |
| Cymbopogon citratus | 2 |
| Eucalyptus staigeriana | 2 |
| Foeniculum vulgare | 2 |
| Hyptis marrubioides | 2 |
| Lippia microphylla | 2 |
| Lippia sidoides | 2 |
| Ocimum gratissimum | 2 |
| Piper arboreum | 2 |
| Piper corcovadensis | 2 |
| Piper hispidinervum | 2 |
| Ageratum conyzoides | 1 |
| Artemisia absinthium | 1 |
| Citrus aurantium | 1 |
| Citrus limon | 1 |
| Citrus sinensis | 1 |
| Cymbopogon winterianus | 1 |
| Eremanthus erythropappus | 1 |
| Eucalyptus citriodora | 1 |
| Eucalyptus urograndis | 1 |
| Eucalyptus urophylla | 1 |
| Hyptis suaveolens | 1 |
| Lippia gracilis | 1 |
| Lippia origanoides | 1 |
| Malva sp. | 1 |
| Mentha sp. | 1 |
| Ocimum selloi | 1 |
| Pelargonium graveolens | 1 |
| Piper aduncum | 1 |
| Piper septuplinervium | 1 |
| Piper subtomentosum | 1 |
| Ruta graveolens | 1 |
| Siparuna guianensis | 1 |
| Tanacetum vulgare | 1 |
| Vanillosmopsis arborea | 1 |
| Zingiber officinale | 1 |
| Total | 57 |
| Positive Control Used in Bibliography | Occurrences in the Literature |
|---|---|
| neem extract (Azadirachta indica) | 2 |
| deltamethrin | 3 |
| α-cypermethrin | 1 |
| β-cypermethrin | 1 |
| fenpropathrin | 1 |
| δ-cyhalothrin | 1 |
| Indoxacarb | 1 |
| chlorpyrifos | 1 |
| Essential Oil | 50% Lethal Doses (LD50; mg/g Insect, CI) | Larval Stage | Reference |
|---|---|---|---|
| Ocimum gratissimum | 2.5 (1.7–2.6) × 10−4 *2 | 3rd | [59] |
| Lippia gracilis | 1.2 (0.9–1.6) × 10−3 *2 | 3rd | [70] |
| Artemisia absinthium | 7.1 (5.3–7.2) × 10−2 *2 | 2nd | [71] |
| Hyptis marrubioides | 0.24 (0.21–0.26) *2 | 2nd | [72] |
| Ocimum basilicum | 0.49 (0.45–0.53) *2 | 2nd | [72] |
| Pelargonium graveolens | 1.13 (0.083–0.145) 2 | 3rd | [73] |
| Lippia alba (LA-10) | 1.2 (0.84–1.57) *2 | 3rd | [73] |
| Lippia alba (LA-57) | 1.21 (0.90–1.57) *2 | 3rd | [73] |
| Ocimum gratissimum | 1.52 (1.36–1.67) 2 | 3rd | [60] |
| Ocimum gratissimum (White wild basil) | 1.52 (1.36–1.67) 2 | 3rd | [61] |
| Lippia alba (LA-22) | 1.56 (1.18–2.02) *2 | 3rd | [73] |
| Ocimum gratissimum (Wild basil) | 2.84 (2.34–3.38) 2 | 3rd | [61] |
| Eucalyptus staigeriana | 3.2 (2.41–4.07) 2 | 3rd | [60] |
| Lippia sidoides | 3.21 (2.95–3.49) 2 | 3rd | [74] |
| Piper hispidinervum | 3.39 (3.42–4.15) *2 | 3rd | [69] |
| Piper hispidinervum | 3.56 (3.22–3.91) *3 | 3rd | [69] |
| Piper corcovadensis | 3.58 (nd) 2 | 3rd | [75] |
| Piper marginatum | 4.18 (nd) 2 | 3rd | [75] |
| Eucalyptus citriodora | 4.58 (4.09–5.08) 2 | 3rd | [61] |
| Corymbia citriodora | 4.59 (4.15–5.03) 2 | 2nd | [76] |
| Piper hispidinervum | 4.62 (4.10–5.22) *1 | 3rd | [69] |
| Ocimum basilicum | 4.86 (4.02–6.13) 2 | 3rd | [61] |
| Foeniculum vulgare | 5.05 (4.13–5.96) 2 | 3rd | [61] |
| Lippia microphylla | 5.35 (4.65–6.05) 2 | 2nd | [76] |
| Ocimum basilicum | 6.27 (5.80–6.73) *2 | 3rd | [77] |
| Piper arboreum | 10.91 (nd) 2 | 3rd | [75] |
| Piper aduncum | 12 (7.1–18.0) 2 | 3rd | [78] |
| Vanillosmopsis arborea | 172.86 (152.8–200.0) 2 | 3rd | [66] |
| Essential Oil | 50% Lethal Concentration (LC50; μL/cm2, CI) | Larval Stage | Reference |
|---|---|---|---|
| Siparuna guianensis | 0.034 (0.033–0.034) μL/cm2 *1 | 3rd | [79] |
| Siparuna guianensis | 0.038 (0.036–0.047) μL/cm2 *2 | 3rd | [79] |
| Ocimum gratissimum | 0.171 (0.150–0.193) μL/cm2 | 3rd | [59] |
| Lippia gracilis | 1.55 (1.51–1.59) μL/cm2 | 3rd | [70] |
| Essential Oils | EO Application Method | Lethal Concentration 50 (LC50) | Concentration | Mortality (%) at 96 h | Larval Stage | Reference |
|---|---|---|---|---|---|---|
| Cymbopogon citratus | On ventral part of the leaf | 0.19 (0.13–0.38) μL/cm2 | 1st | [71] | ||
| Zingiber officinale | 0.25 (0.20–0.35) μL/cm2 | 1st | [71] | |||
| Mentha sp. | 0.33 (0.16–1.93) μL/cm2 | 1st | [71] | |||
| Ruta graveolens | 0.62 (0.49–1.02) μL/cm2 | 1st | [71] | |||
| Malva sp. | 0.67 (0.58–0.82) μL/cm2 | 1st | [71] | |||
| Artemisia absinthium | 2.09 (1.64–2.96) μL/cm2 | 1st | [71] | |||
| Citrus limon | Mixed in artificial diet | 98.29 ppm * | 2nd | [81] | ||
| Citrus aurantium | 100 ppm * | 2nd | [81] | |||
| Ocimum selloi | 600 (580–620) ppm * | 3rd | [83] | |||
| Citrus sinensis | ND | 0.1 mg/g of diet | 0 | 2nd | [82] | |
| 1 mg/g of diet | 5 | 2nd | ||||
| 10 mg/g of diet | 0 | 2nd | ||||
| 0.1 mg/g of diet | 10 | 2nd | ||||
| 1 mg/g of diet | 5 | 2nd | ||||
| 10 mg/g of diet | 0 | 2nd | ||||
| Ageratum conyzoides | By immersion of the maize or rice leaf in EOs solutions | 3430 ppm * | 1st | [80] | ||
| Piper hispidinervum | 9400 (7900–11,100) ppm | 1st | [69] | |||
| 16,200 (14,400–18,400) ppm | 1st | |||||
| 17000 (13,700–21,100) ppm | 1st | |||||
| 17,900 (15,900–20,200) ppm | 1st | |||||
| 18,200 (16,800–19,700) ppm | 1st | |||||
| 28,300 (24,300–32,900) ppm | 1st |
| Essential Oil | 50% Lethal Concentration (LC50; μL/L of Air) | Larval Stage | Reference |
|---|---|---|---|
| Piper septuplinervium1 | 9.4 (7.72–11.4) * | 2nd | [84] |
| Piper subtomentosum2 | 13.2 (10.3–16.6) * | 2nd | [84] |
| Corymbia citriodora3 | 44.85 (36.89–52.81) * | 2nd | [76] |
| Lippia microphylla3 | 116.52 (95.77–137.27) * | 2nd | [76] |
| Piper subtomentosum3 | 146 (116–180) * | 2nd | [84] |
| Compound | 50% Lethal Doses (LD50) (mg/g of Insect, CI) | Mortality (% ± SD) 1 | Larval Stage | LogP | Reference |
|---|---|---|---|---|---|
| γ-cyhalothrin 2 | 1.4 (1.08–1.78) × 10−5 * | 3rd | 6.20 | [78] | |
| Chlorpyrifos 2 | 2.4 (0.83–4) × 10−4 * | 2nd | 4.77–3.71 | [72] | |
| Deltamethrin 2 | 2.45 (1.13–3.76) × 10−4 * | 3rd | 6.20 | [93] | |
| Deltamethrin 2 | 2.46 (1.14–3.78) × 10−4 * | 3rd | 6.20 | [77] | |
| Deltamethrin 2 | 3.07 (2.58–3.35) × 10−3 | 3rd | 6.20 | [74] | |
| Decis 25 2 (Deltamethrin) | 3.17 (2.20–4.57) × 10−3 * | 3rd | 6.20 | [93] | |
| Commercial product 2 | 3.2 (2.2–4.6) × 10−3 * | 3rd | [77] | ||
| trans-anethole | 0.027 (0.021–0.032) | 3rd | 3.17 | [61] | |
| citronellal | 0.07 (0.06–0.08) | 3rd | 3.48 | [61] | |
| Fenpropathrin 2 | 0.18 (0.17–0.23) * | 3rd | 5.48 | [78] | |
| α-cypermethrin 2 | 0.19 (0.12–0.28) * | 3rd | 6.27 | [78] | |
| β-cypermethrin 2 | 1.03 (0.016–1.37) * | 3rd | 6.27 | [78] | |
| linalool | 2.10 (1.65–2.56) * | 3rd | 3.28 | [93] | |
| α-pinene | 2.40 (2.06–2.67) * | 3rd | 4.37 | [59] | |
| thymol | 3.19 (2.93–3.45) * | 3rd | 3.28 | [59] | |
| thymol | 4.91 (4.35–5.56) | 3rd | 3.28 | [74] | |
| linalool | 5.20 (4.21–6.27) | 3rd | 3.28 | [61] | |
| limonene | 32.24 (27.73–36.55) | 3rd | 4.45 | [61] | |
| 1,8-cineole | 2.0 ± 2.0 | 3rd | 2.82 | [73] | |
| limonene | 4.00 ± 2.44 | 3rd | 4.45 | [73] | |
| Azamax 2 | 14.00 ± 5.09 | 3rd | [73] | ||
| geraniol | 30.00 ± 8.84 | 3rd | 3.28 | [73] | |
| citral | 64.00 ± 7.07 | 3rd | 3.17 | [73] | |
| carvone | 84.00 ± 5.09 | 3rd | 2.27 | [73] | |
| linalool | 90.00 ± 3.16 | 3rd | 3.28 | [73] |
| Compound | 50% Lethal Concentration (LC50, CI) | Larval Stage | LogP | Reference |
|---|---|---|---|---|
| Contact toxicity per se | ||||
| Indoxacarb (Rumo 300 g a.i./L; DuPont do Brasil S.A.) | 0.0009 (0.0006–0.0018) μL/cm2 *1 | 3rd | 2.77 | [79] |
| 0.0015 (0.0009–0.0021) μL/cm2 *2 | 3rd | [79] | ||
| thymol | 0.255 (0.195–0.317) μL/cm2 | 3rd | 3.28 | [59] |
| α-pinene | 2.5 (2.11–2.91) μL/cm2 | 3rd | 4.37 | [59] |
| Fumigant toxicity | ||||
| camphene | 0.00067 μL/L * | 2nd | 4.37 | [84] |
| α-pinene | 0.0066 (0.0056–0.0079) μL/L * | 2nd | 4.37 | [84] |
| β-pinene | 0.016 (0.011–0.032) μL/L * | 2nd | 4.37 | [84] |
| Immersion | ||||
| Geraniol | 3793 (173–1281) ppm | 2nd 3 | 3.28 | [88] |
| Essential Oil | Main Compounds (Relative Percentage) | Reference | ||
|---|---|---|---|---|
| Ingestion | ||||
| Cymbopogon citratus | Geranial (47.53%) | Neral (32.5%) | nd | [71] |
| Citrus limon | Limonene * (nd) | nd | nd | [81] |
| Ageratum conyzoides | Precocene (87%) | β- caryophyllene (7.1%) | α-humulene (1.2%) | [80] |
| Fumigation | ||||
| Piper septuplinervium | α-pinene * (21%) | β-pinene * (13.8%) | Citronellal * (10.3%) | [84] |
| Topical application | ||||
| Ocimum gratissimum | Thymol * (33.2%) | p-cymene (22.5%) | γ-terpinene (21%) | [59] |
| Contact toxicity | ||||
| Siparuna guianesis | β-myrcene (74.94%) | 2-undecanone (9.36%) | bicyclo-germacrene (1.52%) | [79] |
| Immersion | ||||
| Piper marginatum | Exalatacin (9.12%) | α-pinene * (8.45%) | α-phellandrene (6.97%) | [75] |
| Essential Oils | Non-Oxygenated Terpenes (%) | Oxygenated Terpenes (%) | Reference |
|---|---|---|---|
| Ingestion | |||
| Ageratum conyzoides | 98.2 | nd | [80] |
| Cymbopogon citratus | nd | 79.03 | [71] |
| Fumigation | |||
| Piper septulinervium | 81.4 | 11.7 | [84] |
| Topical application | |||
| Ocimum gratissimum | 60.2 | 37.6 | [59] |
| Contact toxicity | |||
| Siparuna guianesis | 80.83 | 10.39 | [79] |
| Immersion | |||
| Piper marginatum | 52.72 | 18.35 | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usseglio, V.L.; Dambolena, J.S.; Zunino, M.P. Can Essential Oils Be a Natural Alternative for the Control of Spodoptera frugiperda? A Review of Toxicity Methods and Their Modes of Action. Plants 2023, 12, 3. https://doi.org/10.3390/plants12010003
Usseglio VL, Dambolena JS, Zunino MP. Can Essential Oils Be a Natural Alternative for the Control of Spodoptera frugiperda? A Review of Toxicity Methods and Their Modes of Action. Plants. 2023; 12(1):3. https://doi.org/10.3390/plants12010003
Chicago/Turabian StyleUsseglio, Virginia L., José S. Dambolena, and María P. Zunino. 2023. "Can Essential Oils Be a Natural Alternative for the Control of Spodoptera frugiperda? A Review of Toxicity Methods and Their Modes of Action" Plants 12, no. 1: 3. https://doi.org/10.3390/plants12010003
APA StyleUsseglio, V. L., Dambolena, J. S., & Zunino, M. P. (2023). Can Essential Oils Be a Natural Alternative for the Control of Spodoptera frugiperda? A Review of Toxicity Methods and Their Modes of Action. Plants, 12(1), 3. https://doi.org/10.3390/plants12010003