
Molecular Characterization of a Recombinant Isolate of Tomato Leaf Curl New Delhi Virus Associated with Severe Outbreaks in Zucchini Squash in Southern Italy


Abstract
:1. Introduction
2. Results
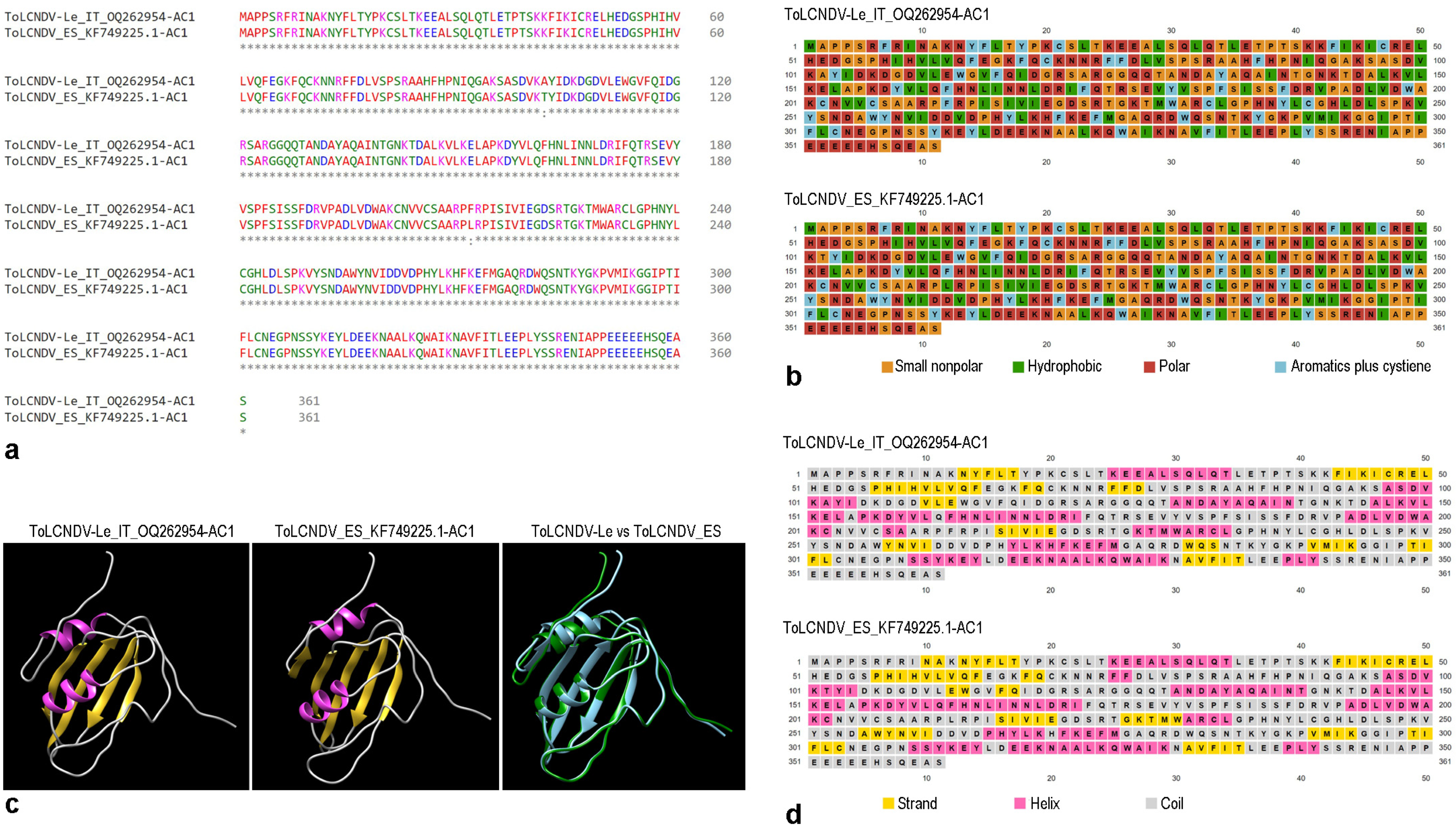
2.1. Sequence Analysis
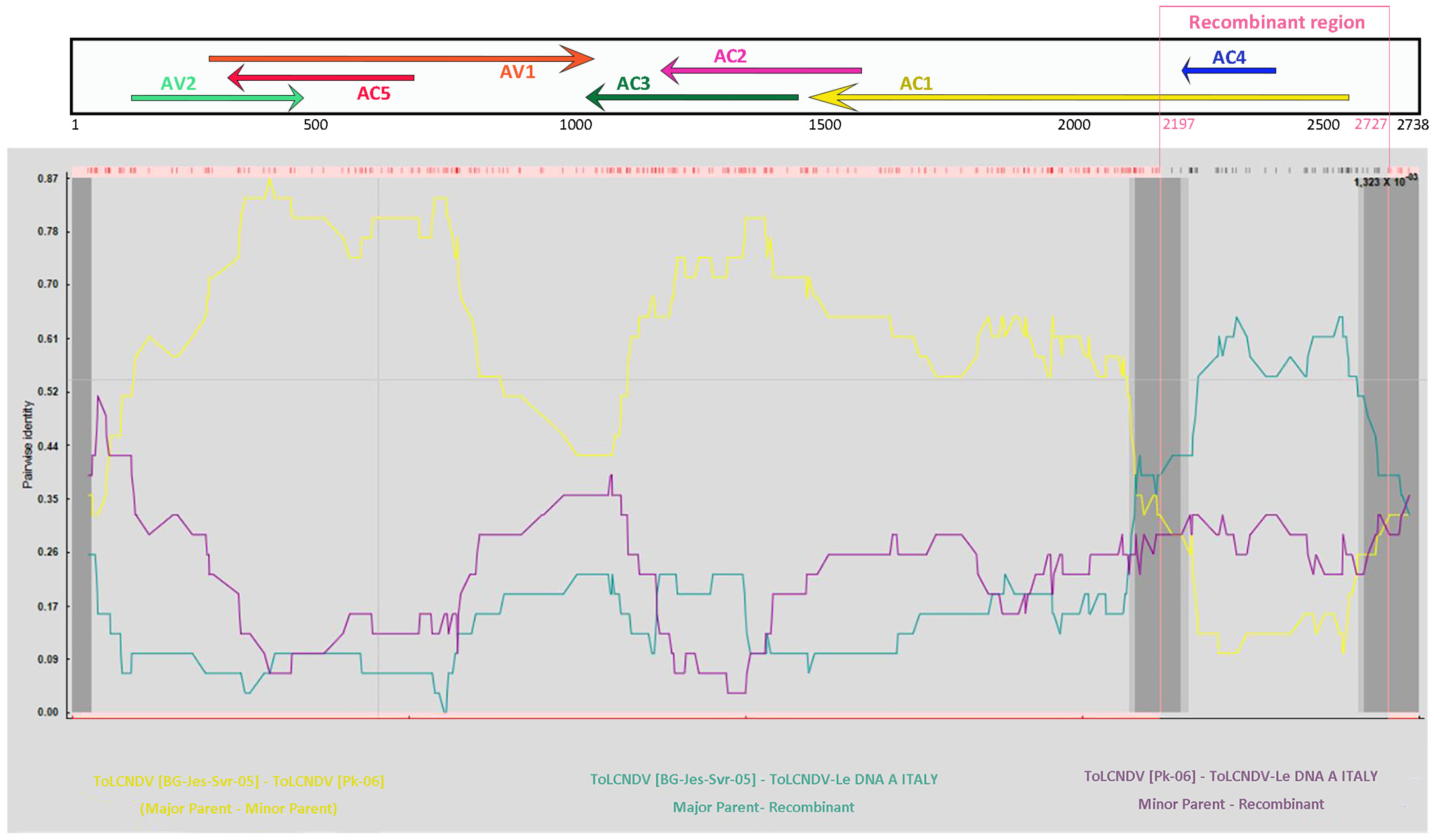
2.2. Recombination Analysis
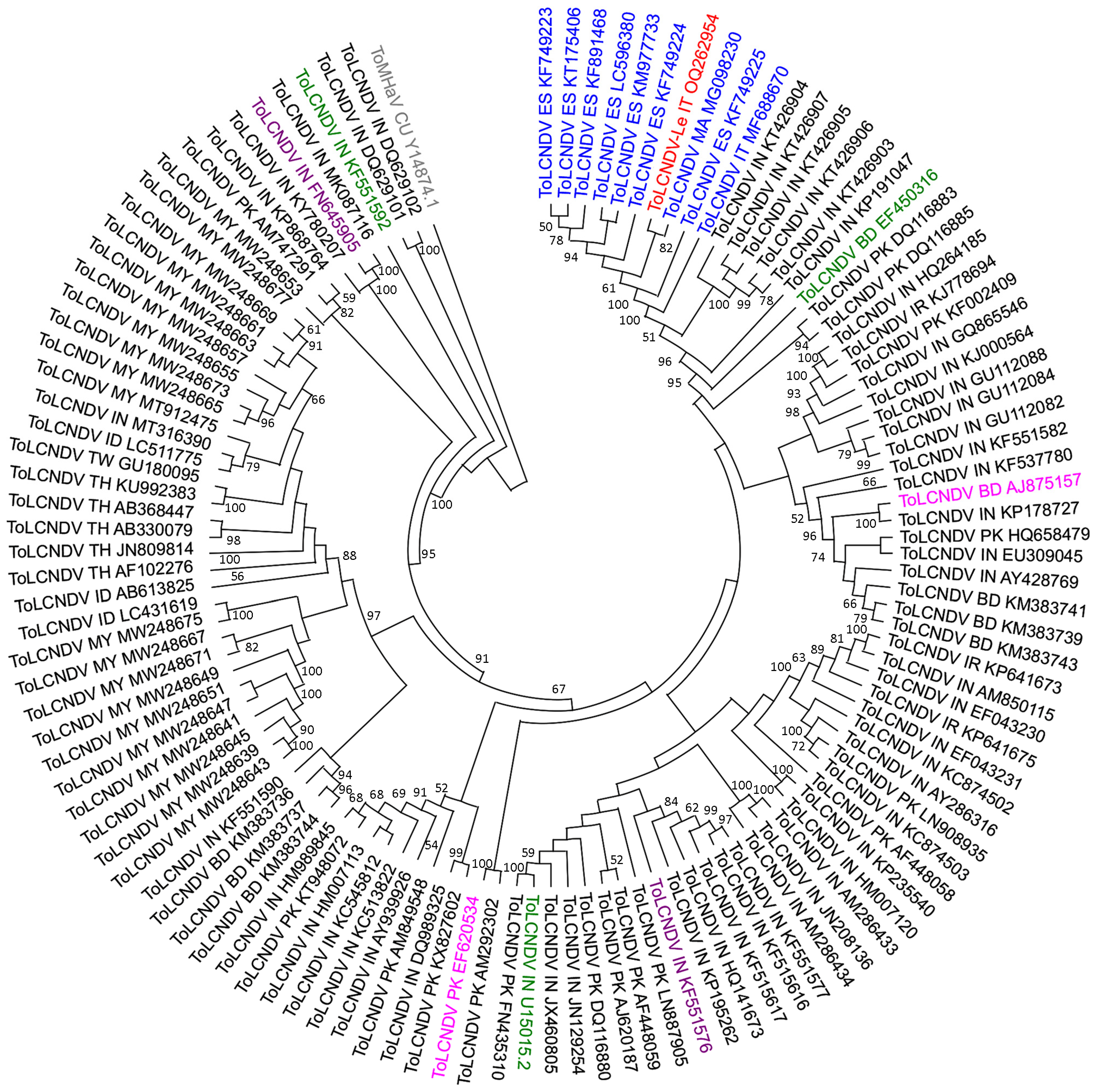
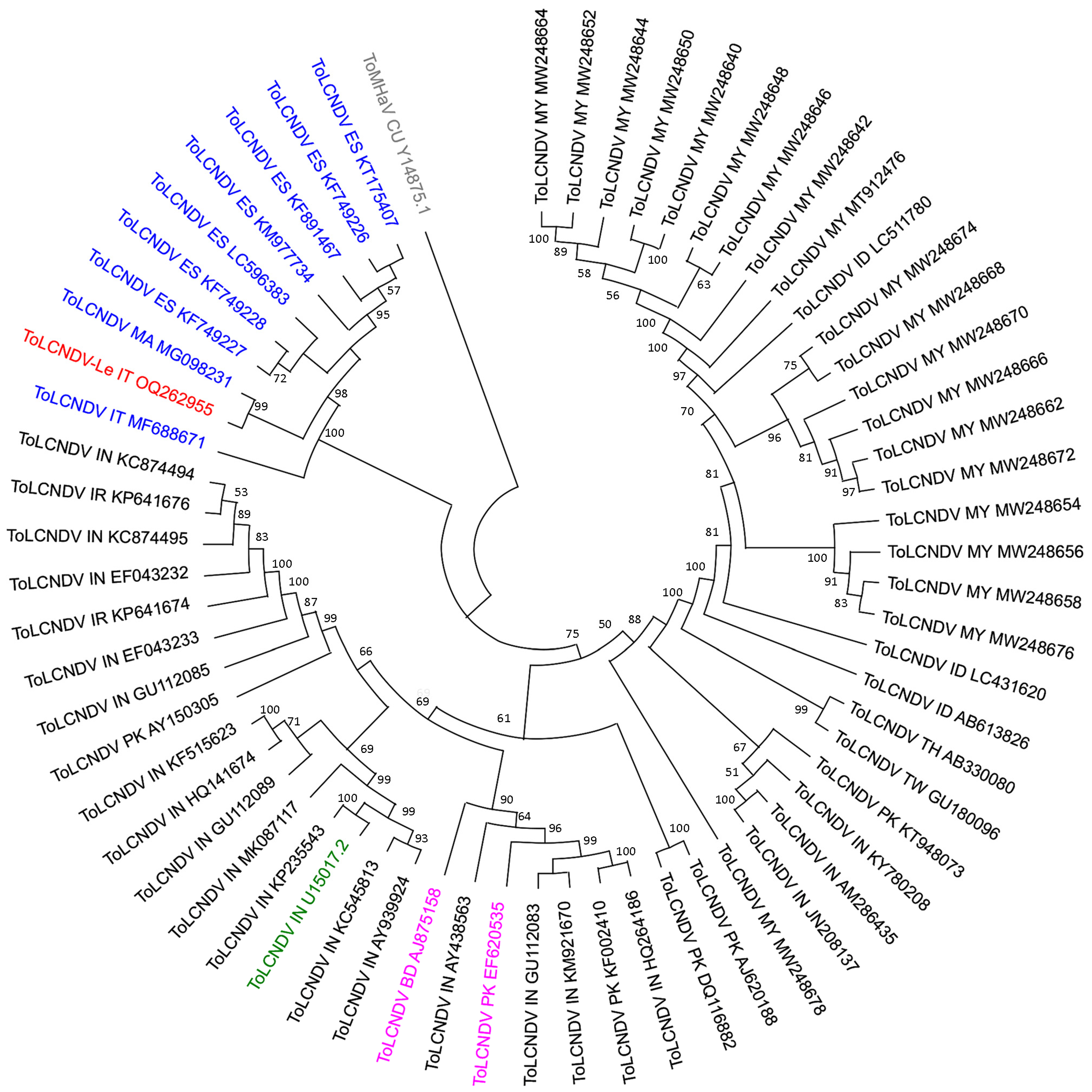
2.3. Phylogenetic Affinities of ToLCNDV-Le Complete Genome
3. Discussion
4. Materials and Methods
4.1. Whole Genome Sequencing (WGS) and Assembly
4.2. Betasatellite Detection in ToLCNDV-Le Infected Plants
4.3. Sequence Validation through Polymerase Chain Reaction (PCR) and Sanger Sequencing
4.4. Phylogeny Relationships and Evidence of Recombination
4.5. Protein Structural Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virus Taxonomy. Available online: https://ictv.global/report/chapter/geminiviridae/geminiviridae/begomovirus (accessed on 13 January 2023).
- Zaidi, S.S.; Martin, D.P.; Amin, I.; Farooq, M.; Mansoor, S. Tomato leaf curl New Delhi virus: A widespread bipartite begomovirus in the territory of monopartite begomoviruses. Mol. Plant Pathol. 2017, 18, 901–911. [Google Scholar] [CrossRef]
- Hussain, M.; Mansoor, S.; Iram, S.; Zafar, Y.; Briddon, R.W. First report of tomato leaf curl New Delhi virus affecting chilli pepper in Pakistan. Plant Pathol. 2004, 53, 794. [Google Scholar] [CrossRef]
- Pratap, D.; Kashikar, A.; Mukherjee, S. Molecular characterization and infectivity of a tomato leaf curl New Delhi virus variant associated with newly emerging yellow mosaic disease of eggplant in India. Virol. J. 2011, 8, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, A.K.; Sneshi, S.K.; Khan, M.S.; Sharma, P.K.; Raj, S.K.; Rao, G.P. Molecular detection and identification of Tomato leaf curl New Delhi virus associated with yellow mosaic and leaf curl disease of Luffa cylindrica in India. Indian Phytopathol. 2012, 65, 80–84. [Google Scholar]
- Maruthi, M.N.; Rekha, A.R.; Cork, A.; Colvin, J.; Alam, S.N.; Kader, K.A. First report of Tomato leaf curl New Delhi virus infecting tomato in Bangladesh. Plant Dis. 2005, 9, 1011. [Google Scholar] [CrossRef]
- Ruiz, M.L.; Simón, A.; Velasco, L.; García, M.C.; Janssen, D. First report of Tomato leaf curl New Delhi virus infecting tomato in Spain. Plant Dis. 2015, 99, 894. [Google Scholar] [CrossRef]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First detection of Tomato leaf curl New Delhi virus infecting zucchini in Spain. Plant Dis. 2014, 98, 857. [Google Scholar] [CrossRef]
- López, C.; Ferriol, M.; Picó, M. Mechanical transmission of Tomato leaf curl New Delhi virus to cucurbit germplasm: Selection of tolerance sources in Cucumis melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Ruiz, L.; Simón, A.; Velasco, L.; Janssen, D. Biological characterization of Tomato leaf curl New Delhi virus from Spain. Plant Pathol. 2017, 66, 376–382. [Google Scholar] [CrossRef]
- Fortes, I.; Sánchez-Campos, S.; Fiallo-Olivé, E.; Díaz-Pendón, J.; Navas-Castillo, J.; Moriones, E. A novel strain of Tomato leaf curl New Delhi virus has spread to the Mediterranean basin. Viruses 2016, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Mnari-Hattab, M.; Zammouri, S.; Belkadhi, M.; Doña, D.B.; Ben Nahia, E.; Hajlaoui, M. First report of tomato leaf curl New Delhi virus infecting cucurbits in Tunisia. New Dis. Rep. 2015, 31, 21. [Google Scholar] [CrossRef] [Green Version]
- Panno, S.; Iacono, G.; Davino, M.; Marchione, S.; Zappardo, V.; Bella, P.; Tomassoli, L.; Accotto, G.P.; Davino, S. First report of Tomato leaf curl New Delhi virus affecting zucchini squash in an important horticultural area of southern Italy. New Dis. Rep. 2016, 33, 6. [Google Scholar] [CrossRef] [Green Version]
- Parrella, G.; Troiano, E.; Formisano, G.; Accotto, G.P.; Giorgini, M. First Report of Tomato leaf curl New Delhi virus Associated with Severe Mosaic of Pumpkin in Italy. Plant Dis. 2017, 102, 459. [Google Scholar] [CrossRef]
- Luigi, M.; Bertin, S.; Manglli, A.; Troiano, E.; Davino, S.; Tomassoli, L.; Parrella, G. First Report of Tomato Leaf Curl New Delhi Virus Causing Yellow Leaf Curl of Pepper in Europe. Plant Dis. 2019, 103, 2970. [Google Scholar] [CrossRef]
- Luigi, M.; Manglli, A.; Bertin, S.; Donati, L.; Tomassoli, L.; Ferretti, L.; Faggioli, F. Development and validation of a specific real-time PCR protocol for the detection of tomato leaf curl New Delhi virus. Eur. J. Plant Pathol. 2020, 157, 969–974. [Google Scholar] [CrossRef]
- Sifres, A.; Sáez, C.; Ferriol, M.; Selmani, E.A.; Riado, J.; Picó, B.; López, C. First report of Tomato leaf curl New Delhi virus infecting zucchini in Morocco. Plant Dis. 2018, 102, 1045. [Google Scholar] [CrossRef]
- Kheireddine, A.; Sifres, A.; Sáez, C.; Picó, B.; López, C. First report of Tomato leaf curl New Delhi virus infecting cucurbit plants in Algeria. Plant Dis. 2019, 103, 3291. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef] [PubMed]
- Simón, A.; Ruiz, L.; Velasco, L.; Janssen, D. Absolute quantification of Tomato leaf curl New Delhi virus Spain strain ToLCNDV-ES: Virus accumulation in a host-specific manner. Plant Dis. 2018, 102, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Van derWalt, E.; Rybicki, E.P.; Varsani, A.; Polston, J.E.; Billharz, R.; Donaldson, L.; Monjane, A.L.; Martin, D.P. Rapid host adaptation by extensive recombination. J. Gen. Virol. 2009, 90, 734–746. [Google Scholar] [CrossRef]
- Moriones, E.; Shelly, P.; Chakraborty, S. Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses 2017, 9, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangeetha, B.; Malathi, V.; Alice, D.; Suganthy, M.; Renukadevi, P. A distinct seed-transmissible strain of tomato leaf curl New Delhi virus infecting Chayote in India. Virus Res. 2018, 258, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.-J.; Vo, T.T.B.; Fadhila, C.; Ho, P.T.; Lal, A.; Troiano, E.; Parrella, G.; Lee, S. Seed-Transmission of Tomato Leaf Curl New Delhi Virus from Zucchini Squash in Italy. Plants 2020, 9, 563. [Google Scholar] [CrossRef]
- Chen, Y.J.; Lai, H.C.; Lin, C.C.; Neoh, Z.Y.; Tsai, W.S. Genetic Diversity, Pathogenicity and Pseudorecombination of Cucurbit-Infecting Begomoviruses in Malaysia. Plants 2021, 10, 2396. [Google Scholar] [CrossRef] [PubMed]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. The role of AV2 (“precoat”) and coat protein in viral replication and movement in tomato leaf curl geminivirus. Virology 1996, 224, 390–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snehi, S.K.; Purvia, A.S.; Parihar, S.S.; Gupta, G.; Singh, V.; Raj, S.K. Overview of Begomovirus genomic organization and its impact. Int. J. Curr. Res. 2017, 9, 61368–61380. [Google Scholar]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. Tomato leaf curl geminivirus from India has a bipartite genome and coat protein is not essential for infectivity. J. Gen. Virol. 1995, 76, 25–35. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 105–140. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Mastrochirico, M.; Spanò, R.; Mascia, T. Grafting to Manage Infections of the Emerging Tomato Leaf Curl New Delhi Virus in Cucurbits. Plants 2023, 12, 37. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Mansoor, S.; Amin, I.; Markham, P.G. Universal primers for the PCR-mediated amplification of DNA beta: A molecule associated with some monopartite begomoviruses. Mol. Biotechnol. 2002, 20, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Open Reading Frame Finder. Available online: https://www.ncbi.nlm.nih.gov/orffinder/ (accessed on 20 July 2022).
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Clustal Omega. Available online: https://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 3 April 2023).
- Psipred. UCL Department of Computer Science: Bioinformatics Group. Available online: http://bioinf.cs.ucl.ac.uk/psipred/ (accessed on 4 April 2023).
- Campos-Olivas, R.; Louis, J.M.; Clerot, D.; Gronenborn, A.M. The structure of a replication initiator unites diverse aspects of nucleic acid metabolism. Proc. Natl. Acad. Sci. USA 2002, 99, 10310–10315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefeuvre, P.; Martin, D.P.; Hoareau, M.; Naze, F.; Delatte, H.; Thierry, M.; Varsani, A.; Becker, N.; Reynaud, B.; Lett, J.M. Begomovirus ‘melting pot’ in the south-west Indian Ocean islands: Molecular diversity and evolution through recombination. J. Gen. Virol. 2007, 88, 3458–3468. [Google Scholar] [CrossRef] [PubMed]
- Chellappan, P.; Vanitharani, R.; Fauquet, C.M. MicroRNA-binding viral protein interferes with Arabidopsis development. Proc. Natl. Acad. Sci. USA 2005, 2, 10381–10386. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 96, 14147–14152. [Google Scholar] [CrossRef] [Green Version]
- Lefeuvre, P.; Lett, J.M.; Reynaud, B.; Martin, D.P. Avoidance of protein fold disruption in natural virus recombinants. PLoS Pathog. 2007, 3, 181. [Google Scholar] [CrossRef] [Green Version]
- Dalmay, T.; Ruhino, L.; Burgyan, J.; Kollar, A.; Russo, M. Functional Analysis of Cymbidium Ringspot Virus Genome. Virology 1993, 194, 697–704. [Google Scholar] [CrossRef]
- Babraham Bioinformatics. Available online: www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 5 July 2022).
- Giardine, B.; Riemer, C.; Hardison, R.C.; Burhans, R.; Elnitski, L.; Shah, P.; Zhang, Y.; Blankenberg, D.; Albert, I.; Taylor, J.; et al. Galaxy: A platform for interactive large-scale genome analysis. Genome Res. 2005, 15, 1451–1455. [Google Scholar] [CrossRef] [Green Version]
- Blankenberg, D.; Von Kuster, G.; Coraor, N.; Ananda, G.; Lazarus, R.; Mangan, M.; Nekrutenko, A.; Taylor, J. Galaxy: A web-based genome analysis tool for experimentalists. Curr. Protoc. Mol. Biol. 2010, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Panno, S.; Caruso, A.G.; Davino, S. First Report of Tomato Brown Rugose Fruit Virus on Tomato Crops in Italy. Plant Dis. 2019, 103, 1443. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Martin, D.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Lemey, P.; Lott, M.; Moulton, V.; Posada, D.; Lefeuvre, P. RDP3: A flexible and fast computer program for analyzing recombination. Bioinformatics 2010, 26, 2462–2463. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF * | Protein Function | Strand | Start | Stop | ORF Length (nt) | ORF Coding Capacity |
|---|---|---|---|---|---|---|
| AV1 | Coat | V | 279 | 1049 | 771 | 256 |
| AV2 | Precoat | V | 119 | 457 | 339 | 112 |
| AC1 | Replication initiation | C | 2583 | 1498 | 1086 | 361 |
| AC2 | Transcription activator | C | 1595 | 1191 | 405 | 134 |
| AC3 | Replication enhancer | C | 1456 | 1046 | 411 | 136 |
| AC4 | Pathogenesis-related | C | 2426 | 2250 | 117 | 58 |
| AC5 | Unknown | C | 671 | 309 | 363 | 120 |
| BV1 | Nuclear shuttle | V | 430 | 1236 | 807 | 268 |
| BC1 | Movement | C | 2135 | 1290 | 846 | 281 |
| Recombinant 1 | Potential Parents 2 | Recombination Breakpoint | Average p Values in Detecting Algorithms 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Minor | Major | R | M | C | S | T | L | ||
| ToLCNDV-Le DNA-A | ToLCNDV [PK-05] (EF620534) | ToLCNDV [BG-Jes-Svr-05] (AJ875157) | 2197–2727 | 1.323 × 10−3 | 1.724 × 10−2 | 7.821 × 10−3 | 4.982 × 10−12 | 8.523 × 10−6 | 7.144 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastrochirico, M.; Spanò, R.; De Miccolis Angelini, R.M.; Mascia, T. Molecular Characterization of a Recombinant Isolate of Tomato Leaf Curl New Delhi Virus Associated with Severe Outbreaks in Zucchini Squash in Southern Italy. Plants 2023, 12, 2399. https://doi.org/10.3390/plants12132399
Mastrochirico M, Spanò R, De Miccolis Angelini RM, Mascia T. Molecular Characterization of a Recombinant Isolate of Tomato Leaf Curl New Delhi Virus Associated with Severe Outbreaks in Zucchini Squash in Southern Italy. Plants. 2023; 12(13):2399. https://doi.org/10.3390/plants12132399
Chicago/Turabian StyleMastrochirico, Mariarosaria, Roberta Spanò, Rita Milvia De Miccolis Angelini, and Tiziana Mascia. 2023. "Molecular Characterization of a Recombinant Isolate of Tomato Leaf Curl New Delhi Virus Associated with Severe Outbreaks in Zucchini Squash in Southern Italy" Plants 12, no. 13: 2399. https://doi.org/10.3390/plants12132399
APA StyleMastrochirico, M., Spanò, R., De Miccolis Angelini, R. M., & Mascia, T. (2023). Molecular Characterization of a Recombinant Isolate of Tomato Leaf Curl New Delhi Virus Associated with Severe Outbreaks in Zucchini Squash in Southern Italy. Plants, 12(13), 2399. https://doi.org/10.3390/plants12132399