1. Introduction
Species-specific variations in response to stress fundamentally underlie community composition and ecosystem services; quantifying this interspecific variation can help forecast future distributions and predict community dynamics. For example, interspecific variation in stress tolerance drives succession [
1,
2], spatial patterning such as zonation [
3], ecological filters [
4], species interactions [
5,
6], and species distributions [
7,
8]. Stress events can be broadly categorized as press stressors (i.e., long-term changes in an environmental variable) and pulse stressors (i.e., shorter, punctuated events) [
9,
10].
Salinity is a near universal stressor for terrestrial plants. Elevated salt concentrations induce osmotic stress, mimicking drought conditions and making it more difficult for plants to uptake the water necessary to perform vital functions, including photosynthesis and maintaining turgor pressure [
11]. Additionally, salt can have toxic effects on plant tissues [
12]; it can reduce germination, nutrient uptake, seed production or crop yield, and growth, and can ultimately cause senescence and plant death [
12,
13,
14].
Salinity is often dynamic in the soil environment and can occur as a press or pulse stressor [
15]. Sea-level rise (SLR) is a press stressor that acts over longer timescales, as salinity steadily rises when tidal waters permeate the soil and groundwater [
10]. This primarily impacts stress intensity, as salt concentrations gradually rise over decadal timescales. Saltwater intrusion (SWI; defined as the movement of saltwater into inland areas via groundwater) and storm events act as pulse stressors, with seawater overtopping soil during acute flooding events and causing punctuated rises in groundwater and porewater salinity at local and regional scales [
16]. The frequency and strength of storm and SWI events are anticipated to increase due to elevated sea surface temperatures in tandem with gradually rising sea levels [
17]. It is therefore imperative to study the interaction between salinity intensity and stress duration in order to anticipate its impact on coastal plant communities in the face of pulse and press stressors [
17,
18]. This is the impetus for our study, which seeks to investigate, on a species-level, how plants with different salinity adaptations will respond to elevated stress along multiple axes of exposure.
Controlled greenhouse experiments and field observations have demonstrated that plant ecophysiological responses to increased salinity are species-specific, with marked differences in how species tolerate and recover from seawater inundation events [
10,
18,
19,
20]. Plant species can also vary in thresholds of tolerance based on the duration of salinity exposure [
10]. Some non-halophytic plant species can recover after short-term exposure to elevated salinity levels as long as post-pulse conditions allow for a recovery period, i.e., lowered interstitial salinities [
21,
22]. This is a source of resilience for coastal plant communities experiencing pulses of salinity. Yet, if elevated salinity persists over time and becomes a press stressor, or the post-pulse environment is not suitable for plant recovery, salt-sensitive species are filtered out of the community assemblage [
10,
21]. Interspecific variation in tolerance and recovery patterns between species can result in a shift in community composition. This, in turn, can alter ecosystem function [
23,
24,
25]. Understanding which plant species are tolerant to different combinations of salinity intensity and duration can offer insight into how communities are likely to shift in composition, and therefore function, over time with SLR and SWI [
24,
25].
To investigate how pulses and presses of salinity stress affect the survival and fitness of coastal plant species, we manipulated two variables of salinity exposure that represent the pulse and press stressors occurring in coastal ecosystems: (1) intensity, or the concentration of salt in water, measured in parts per thousand (PPT); and (2) duration, or the length of time that plants were exposed to salt water, measured in days per month (DPM) [
18]. We also calculate a novel stress index that integrates salinity stress intensity and duration, which we call the salt stress exposure (SSE) index.
The SSE index was inspired by the growing degree days index, which has intuitive units and is extremely useful in relating temperature and organismal physiology. The growing degree days index tabulates the time when plant growth can occur by summing the number of degrees over a baseline temperature which accumulate each day over a growing season, and the summation of which describes the total growing time that falls within benign operating conditions [
26]. Every degree over the threshold temperature contributes to the growth of the plant, and growing degree days are highly correlated with plant production and yield. Building on this framework of growing degree days, we estimate the SSE index by multiplying the intensity treatment by the duration treatment to accumulate salinity exposure “stress-time”. SSE combines intensity and duration into a single variable, as a simple representation of the interaction between intensity and duration.
As species vary greatly in their ability to tolerate salinity, this study aimed to test salinity tolerance in a group of coastal and upland species that exhibit a diverse array of adaptations to salinity [
27] (
Table 1). Species fall into two broad tolerance groups: halophytes and glycophytes. Halophytic plants have evolved specialized adaptations to cope with saline conditions, such as synthesizing organic solutes and accumulating stress metabolites such as proline, calcium, or potassium ions to maintain osmotic balance between internal and external environments [
12,
28]. Additional adaptations for salt tolerance include low rates of sodium and chlorine ion transport to the leaves and compartmentalization in vacuoles or cell walls to prevent toxic buildup in the cytoplasm [
11]. A select number of halophytes are also able to exude salt from salt glands in their leaves to prevent toxicity [
29,
30]. Having one or more of these adaptations allows halophytic species to avoid toxicity from an excessive buildup of salt ions. Glycophytes lack these adaptations and are therefore more susceptible to salinity stress.
Coastal and tidal wetland species are halophytic and adapted to repeated inundation events and elevated levels of salinity. We expected halophytic species (
Table 1) to maintain biomass and chlorophyll production at elevated levels of salinity (both intensity and duration). Upland plant species tested within this study included native and introduced herbs as well as several common crop species (
Table 1). We expected to see less salinity tolerance in upland, glycophytic species, resulting in lower biomass and lower chlorophyll production in response to increasing salinity intensity and duration.
Table 1.
Known halophytic adaptations of the focal species in this study.
Table 1.
Known halophytic adaptations of the focal species in this study.
| Species | Tolerance Class | Adaptations to Saline Conditions |
|---|
| Kosteletzkya virginica | higher tolerance, native | Accumulates inorganic compounds and synthesizes organic compounds to maintain osmotic balance [31], including proline [31,32]. Mucilage in shoot, stems, and roots assists in regulating water ascent and ion transport [33]. |
| Panicum amarum | higher tolerance, native | Adapted to dune systems, where plants are subjected to salt spray, storm surges, inundation, and other stresses; can withstand extended drought [34]. |
| Panicum virgatum | higher tolerance, native | Salt excretion glands [35]; synthesizes organic compounds (e.g., proline) to maintain osmotic balance. |
| Paspalum floridanum | higher tolerance, native | Observed in saline ditches adjacent to farm fields (pers. comm., Chris Miller). |
| Phragmites australis | higher tolerance, invasive | Salt excretion glands [36]; organic and inorganic compounds raise osmotic pressure [27]; oxidoreductase activity and glutathione metabolism [37]; free amino acids and sugars as osmolytes (namely proline and glutamine) [38]; ions (K+, Na+, Ca2+, Mg2+ and Cl−) in bundle sheath and mesophyll cells [39]. |
| Brassica napus | lower tolerance, native | Accumulates proteins associated with protein metabolism and damage repair in saline conditions [40]. |
| Glycine max | lower tolerance, crop, a genetically modified, chloride-excluding variety | Root length unaffected and root water content increased in mild to moderate salinity (3 to 12 PPT) [41]. |
| Persicaria punctata | lower tolerance, native | Found in low-salinity (~5 PPT) tidal marshes [42]. |
| Persicaria maculosa | lower tolerance, invasive | No published salt-tolerance information. |
| Sorghum bicolor | lower tolerance, crop | Acclimation at low salinity allowed internal regulation of Na+ and Cl− concentrations during extended exposure to higher salt concentrations [43]. |
By developing physiological response curves based on exposure to various levels of salinity intensity and duration, this study investigated which metric(s) of salinity exposure explain variation in survival and plant ecophysiology (biomass and chlorophyll production) of halophytic and glycophytic species. In addition, we test the ability of a simplified salinity stress metric, the SSE index, to describe species tolerance to salinity stress, and compare this index to the alternative of models that include separate predictor variables of intensity and duration. The research significance of this study is to identify the metric of salinity that best explains plant physiology and mortality, detect whether this metric differs between glycophytes and halophytes, and identify, from amongst the tested crops, those which perform best in elevated salinity.
4. Materials and Methods
4.1. Species Selection and Sourcing
We selected five glycophytes,
Brassica napus,
Glycine max,
Persicaria maculosa,
Persicaria punctata, and
Sorghum bicolor, and five halophytes,
Kosteletzkya virginica,
Panicum amarum,
Panicum virgatum,
Paspalum floridanum, and
Phragmites australis, for salinity stress exposure experiments. These species were selected based on their importance in coastal and agro-ecosystems in the region, as well as seed availability.
P. australis is a pervasive invasive species that is dominant in many coastal wetland systems in the eastern United States; all other halophytes were native species in the Mid-Atlantic region. Among the glycophytes,
P. maculosa is an introduced species, and
B. napus,
G. max, and
S. bicolor are important crops in the region. We intentionally included two sets of congeners, the two
Persicaria species in the glycophyte experiment and the two
Panicum species in the halophyte experiment. Additionally, the selected study species encompass a variety of ecological niches, habitat preferences, and known halophytic adaptations (
Table 1). Therefore, we expected to see different responses to salinity stress between species, as well as between the glycophytic and halophytic tolerance classes.
Seeds for the glycophyte experiment were sourced from the same sources as in de la Reguera [
57], with the exception of
P. maculosa, which was collected from an urban, roadside population in northwest Washington, D.C., and
P. punctata, which was collected from the understory of a maritime forest in The Nature Conservancy’s Brownsville Preserve in Nassawadox, Virginia, USA. Seeds for the halophyte experiment were provided by the USDA-NRCS Cape May Plant Materials Center, with the exception of
P. australis, which was collected from the forest edge of Moneystump Swamp in Blackwater National Wildlife Refuge.
4.2. Experimental Design
Two salt stress exposure experiments were conducted in the Harlan Greenhouse in George Washington University’s Science and Engineering Hall. The first experiment, run from 15 March to 9 May 2021, tested the five glycophytes, and also included the moderately salt-tolerant P. virgatum. The second greenhouse experiment was run from 15 May to 16 July 2021 and tested the five halophytes. P. virgatum was included in both experiments to bridge results.
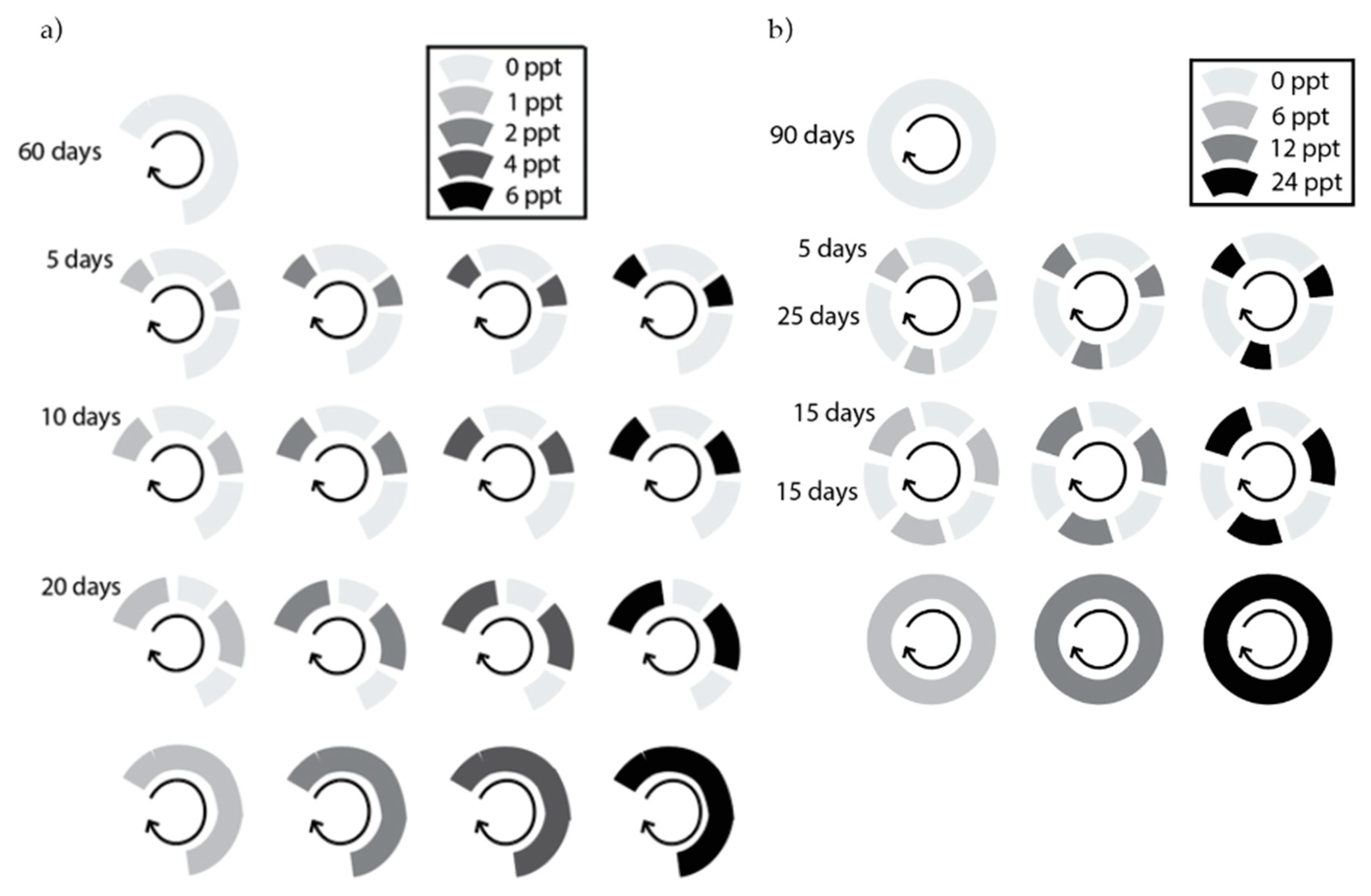
Salinity duration was controlled by the amount of time plants were subjected to salt water versus fresh water. Salinity intensity was controlled by the salt concentration of the saltwater. We exposed species to a range of salinity intensities appropriate to their salt stress tolerance class (maximum 6 PPT for the glycophyte experiment; maximum 24 PPT for the halophyte experiment). In the glycophyte experiment, intensity treatments were 0 (freshwater control), 1, 2, 4, and 6 PPT, and duration treatments were 0, 5, 10, 20, and 30 days of salinity exposure per month (DPM) (
Figure 4a). In the halophyte experiment, intensity treatments were 0 (freshwater control), 6, 12, and 24 PPT, and duration treatments were 0, 5, 15, and 30 DPM (
Figure 4b). Therefore, the glycophyte experiment included 17 combinations of salinity intensity and duration, while the halophyte experiment included 10 combinations (
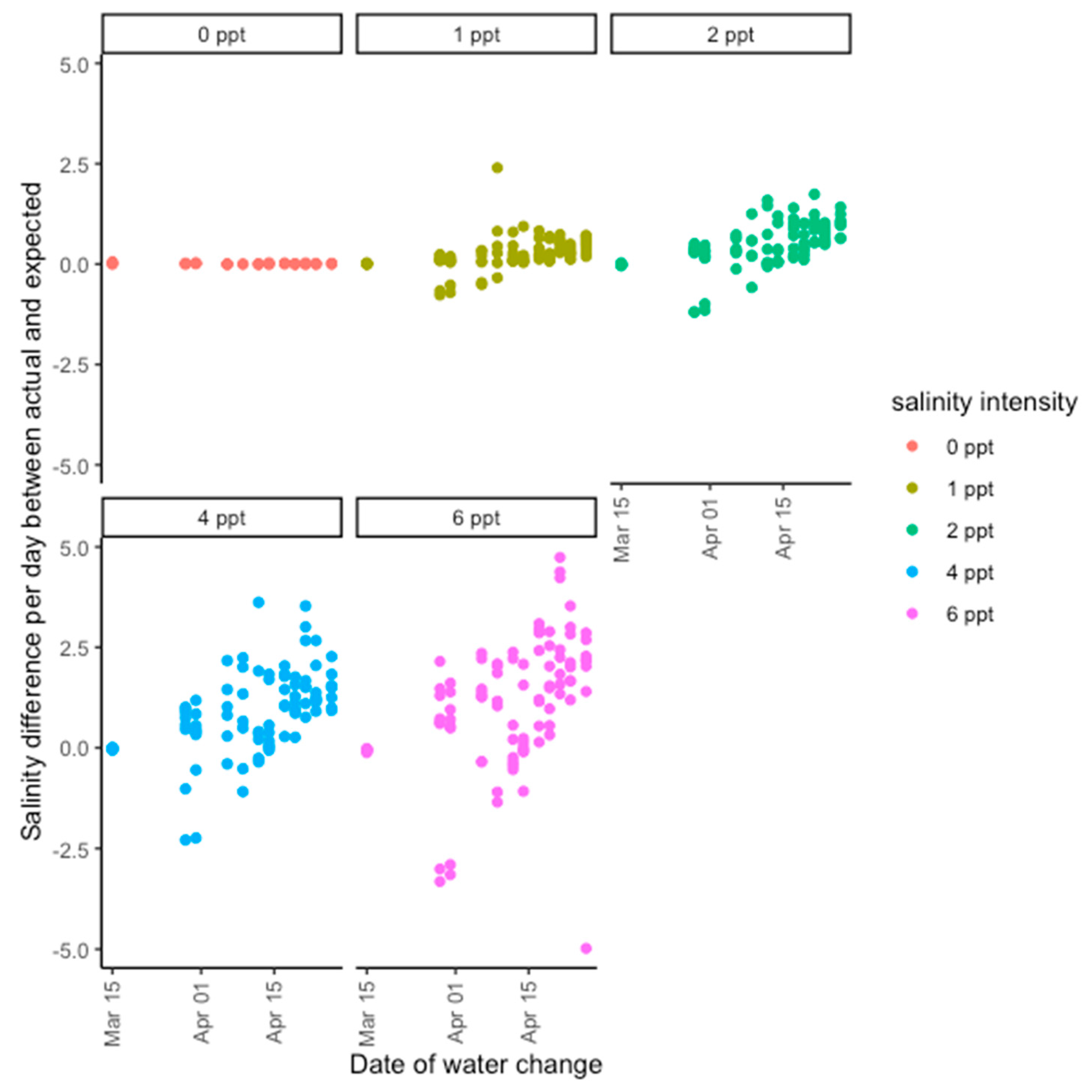
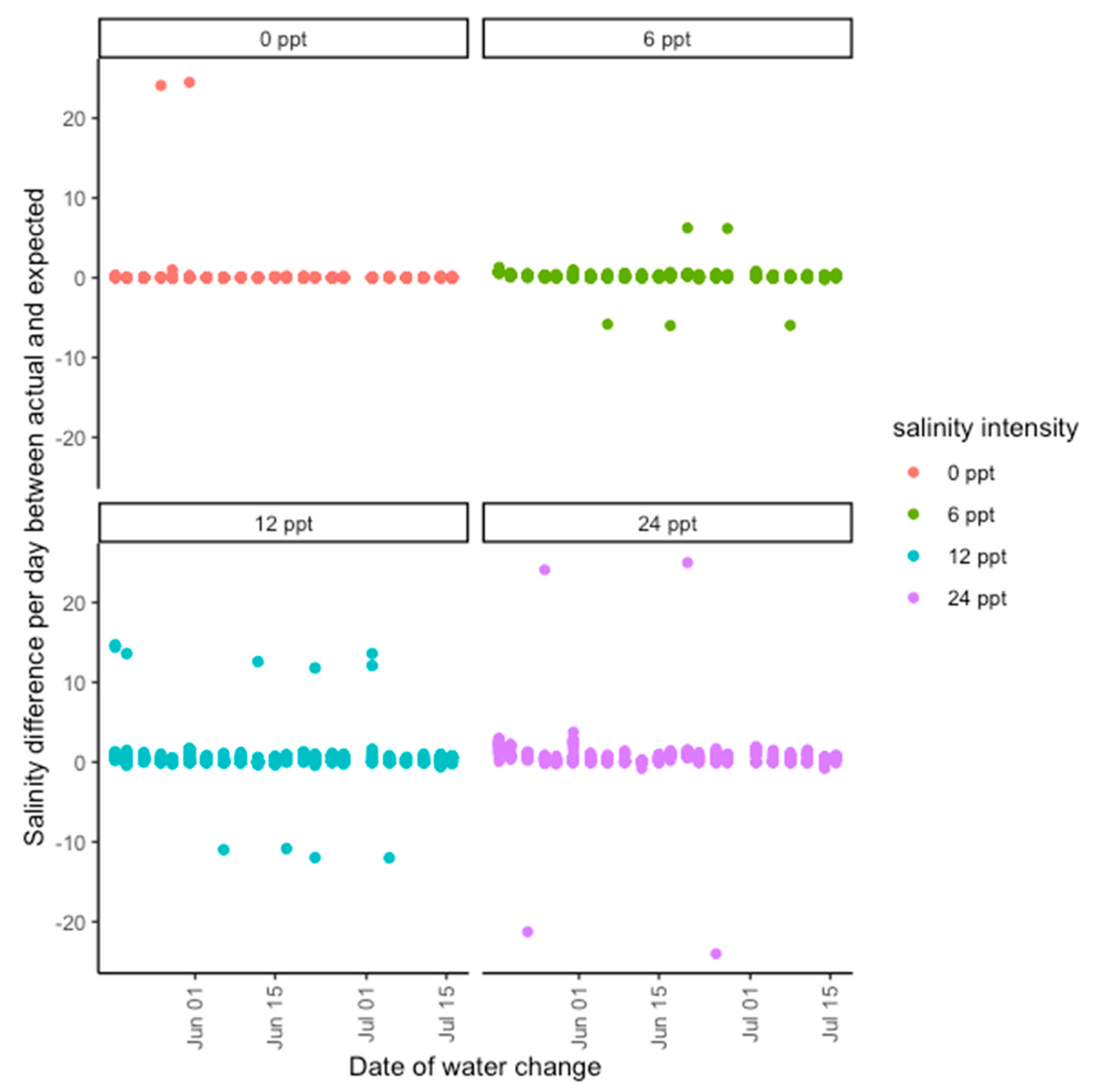
Figure 4). Saltwater was made by mixing Instant Ocean with deionized water in large containers to reach the target concentration. Plants were bottom-watered, and water was changed every 3 days to prevent salinity buildup in soils due to evaporation. During the water change for the halophyte experiment, soil was gently flushed with freshwater for 15 s to prevent the buildup of salinity in the soil, which proved a challenge during the glycophyte experiment (
Figure A1 and
Figure A2). In both experiments, trays were rotated among greenhouse benches at each watering to prevent greenhouse position effects. A handheld salinity meter (Pocket Pro+ Multi 1) was used to check the source water salinity of each treatment. The glycophyte experiment ran for 60 days, or two 30-day cycles, and the halophyte experiment ran for 90 days, or three 30-day cycles.
All plants used in this study were germinated from seed in George Washington University’s Harlan Greenhouse. To stimulate germination, seeds were spread on top of a wet potting mix (2:1 of potting mix:sand) and placed into a warm and humid germination chamber. Seeds for the halophyte experiment were treated with fungicide prior to germination. Trays of germinated seedlings were placed on heated mats until developing true leaves, at which point the plants were individually repotted and tagged with a unique identifier to facilitate tracking throughout the experiment.
The glycophytes were repotted into 10 × 10 × 10 cm cubic pots and placed into flat trays (
Figure A3). Three individuals of each of the six glycophyte species were placed into each tray. In the halophyte experiment, seedlings were repotted into conical pots, and placed in a plant stand set atop six cylindrical containers (
Figure A4). Each cylindrical container held five cones with one individual of each of the five halophytic species. All plants within a tray or container received the same water, and therefore the same stress treatments. The tray or container served as an experimental replicate and was randomly assigned to a combination of salt intensity and duration. Plants were haphazardly assigned a treatment combination. In each experiment, there were ten replicates of each treatment combination.
4.3. Response Variables—Survival, Biomass, and Chlorophyll
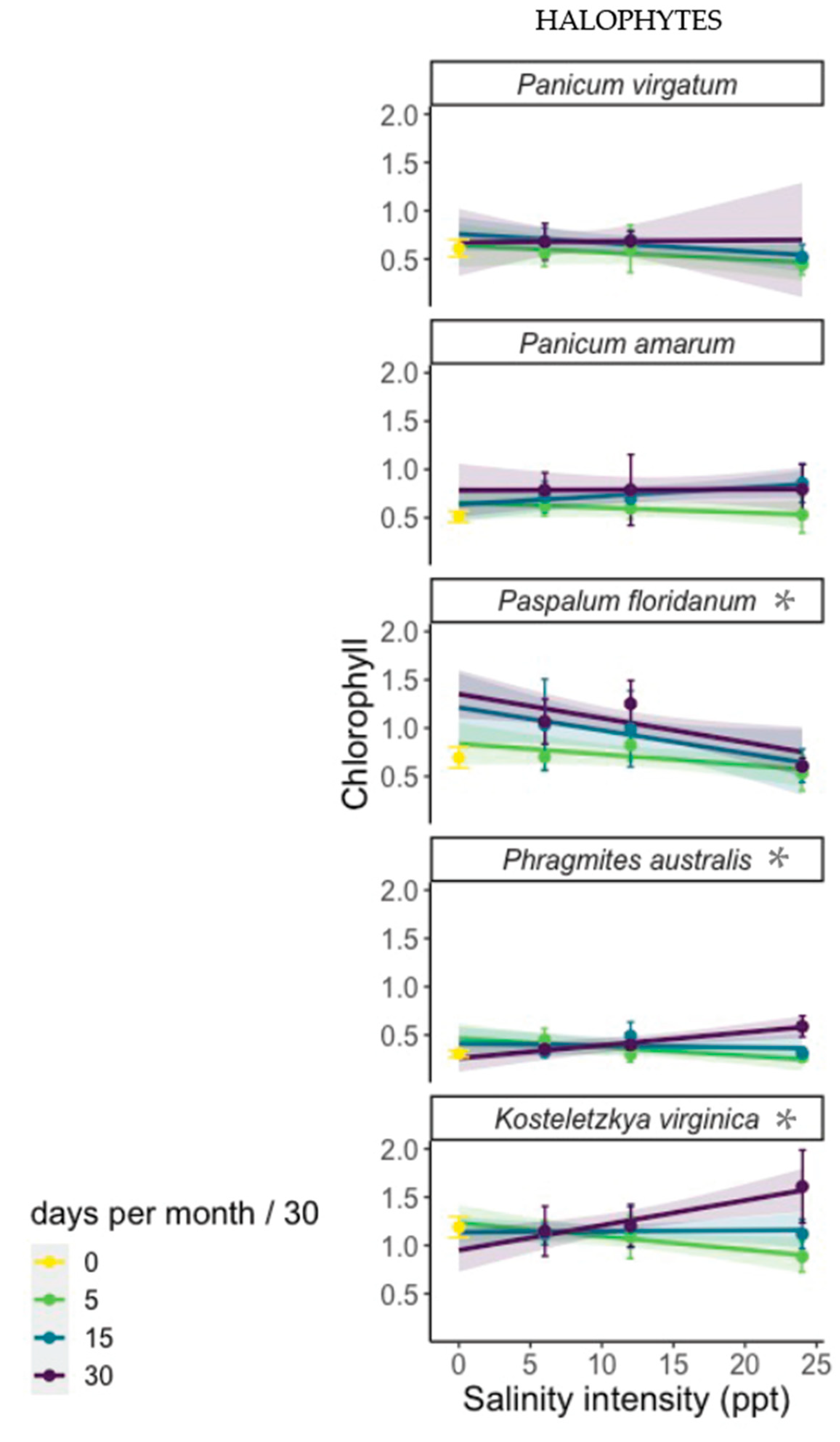
We screened for salt tolerance by measuring survival, biomass, and chlorophyll response to increasing salinity. We monitored survival every three days. Plants with no green aboveground tissue were scored as dead and removed from the experiment. At the end of the experiment, plants were removed from their pots, and gently agitated in water to remove the soil from the roots. Biomass was separated into aboveground live, aboveground dead, and belowground components. All biomass was dried at 60 ° for at least 72 h before being weighed. Chlorophyll measurements were made in the halophyte experiment only, in the week prior to harvest, using a chlorophyll meter (model CCM-200 plus, Opti-Sciences). We made three measurements of chlorophyll concentrations in the third fully expanded leaf, or in the second if the plant had only two leaves.
4.4. Statistical Analysis
Outliers were removed from the dataset via visual analysis prior to data transformations and model analysis. Plant mortality was coded as a binary variable (1 indicating dead and 0 indicating alive). To facilitate comparisons across species that varied in size and growth rate, total biomass and aboveground biomass were transformed to a standard normal distribution using the following equation:
where x is biomass of species y.
For each experiment, we ran a series of models that included intensity (PPT), duration (DPM), and their interaction, or only the salt stress exposure index (SSE), to identify the metric that best explained patterns of plant fitness and survival in response to experimental salinity stress in greenhouse manipulations. SSE was calculated by multiplying the PPT treatment by the DPM treatment to generate a single linear scale of salinity stress. Generalized linear mixed-models were built using the glm() function in R. In addition to these predictor variables, all models included species as a fixed factor to allow patterns to vary between species, and the tray as a random factor with a zero intercept.
In order to determine the metric(s) of salinity that best explained variation in survival and ecophysiology, we performed model selection based on the model with the lowest AIC value as the model of best fit for each experiment. We selected models for each experiment independently, as we expected glycophyte and halophyte groups to exhibit different responses to salinity stress. After selecting the model of best fit for the tolerance class, we ran the model for each of the species to identify the species-specific significance of predictor coefficients.
Standardized total biomass and standardized aboveground biomass met assumptions of normality and homogeneity of variance by definition; however, chlorophyll data were log-transformed prior to analysis to meet these assumptions. Total biomass was tested for assumptions of normality and homogeneity of variance prior to linear regression or ANOVA. Where ANOVA indicated a significant effect, pairwise comparisons were made using Tukey’s Highly Significant Difference tests.
Lastly, simple linear regression models were used to predict total biomass as a function of PPT within the highest duration treatment, 30 DPM, as a measure of each species’ sensitivity to salinity. A more negative regression coefficient indicated greater salinity sensitivity. Results were compared to species’ salt tolerances in the literature.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







