
Iodine-Biofortified Microgreens as High Nutraceutical Value Component of Space Mission Crew Diets and Candidate for Extraterrestrial Cultivation

 ,
,  ,
,  ,
,  , ,
, ,  ,
, 
 and
and 
Abstract
1. Introduction
2. Results
2.1. Microgreens Yield
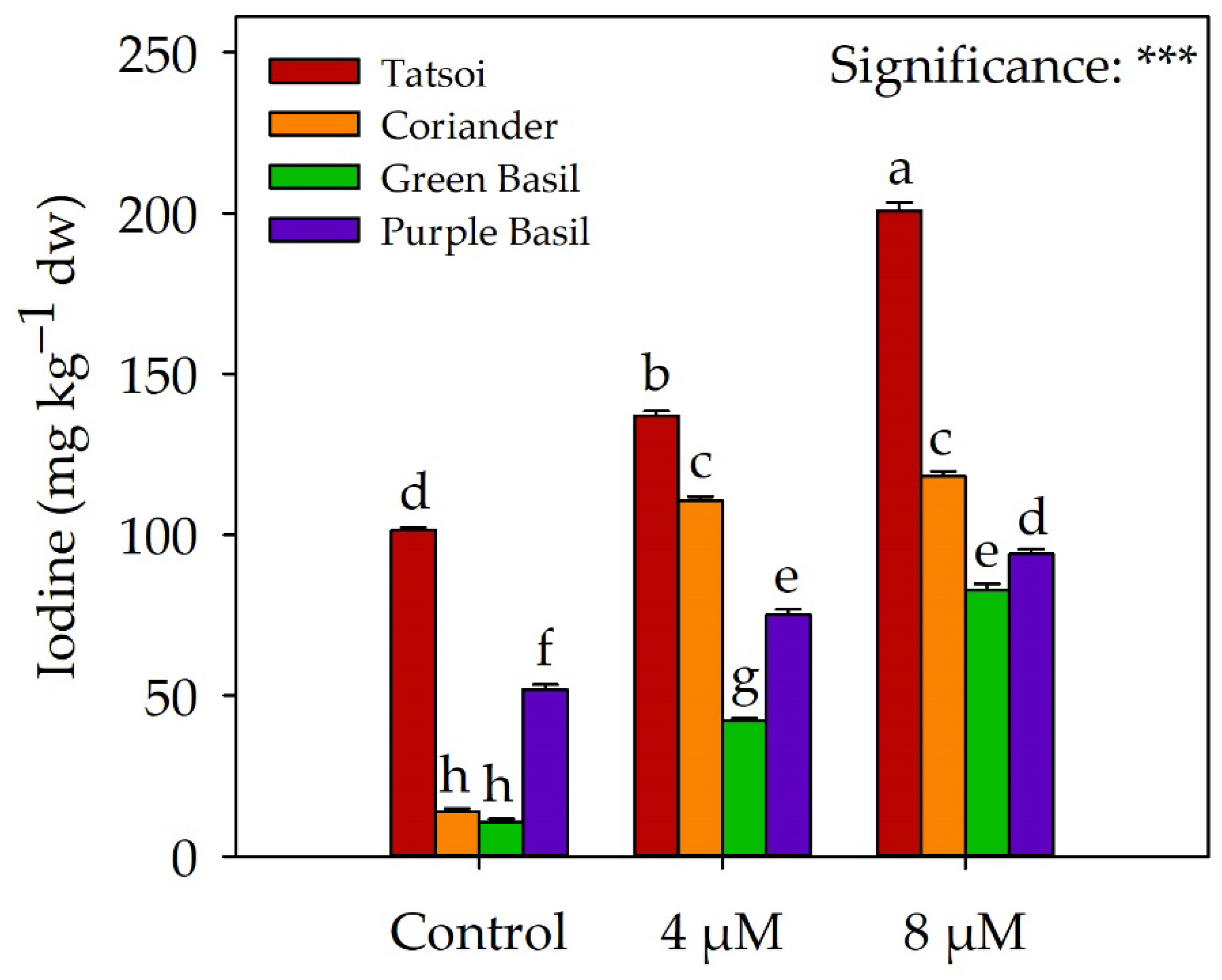
2.2. Iodine Biofortification
2.3. Pigments Concentration and Antioxidant Activity
2.4. Phenolic Acids
2.5. Total Polyphenols and Flavonoids
3. Discussion
3.1. Effect of Iodine Biofortification on Yield and Growth Parameters
3.2. Effect of Iodine Biofortification on Microgreens Iodine Concentration
3.3. Effect of Iodine Biofortification on Pigments and Antioxidants Activities
3.4. Effect of Iodine Biofortification on Phenolics
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Plant Material Collection, Yield, and Hypocotyl Length
4.3. Iodine Concentration Determination
4.4. Chlorophyll and Carotenoids Concentration
4.5. Antioxidant Activity
4.6. Determination of Polyphenols
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Douglas, G.L.; Zwart, S.R.; Smith, S.M. Space food for thought: Challenges and considerations for food and nutrition on exploration missions. J. Nutr. 2020, 150, 2242–2244. [Google Scholar] [CrossRef] [PubMed]
- Enrico, C. Nutraceutical and Nutrients Development for Space Travel. In Handbook of Space Pharmaceuticals; Springer: Berlin/Heidelberg, Germany, 2022; pp. 747–769. [Google Scholar]
- Kulkarni, S.; Gandhi, D.; Mehta, P.J. Nutraceuticals for Reducing Radiation Effects During Space Travel. In Handbook of Space Pharmaceuticals; Springer: Berlin/Heidelberg, Germany, 2022; pp. 915–933. [Google Scholar]
- Smith, S.M.; Zwart, S.R.; Heer, M. Human Adaptation to Space Flight: The Role of Nutrition; National Aeronautics and Space Administration, Lyndon B. Johnson Space Center: Houston, TX, USA, 2014. [Google Scholar]
- Nguyen, M.; Knowling, M.; Tran, N.N.; Burgess, A.; Fisk, I.; Watt, M.; Escribà-Gelonch, M.; This, H.; Culton, J.; Hessel, V. Space farming: Horticulture systems on spacecraft and outlook to planetary space exploration. Plant Physiol. Biochem. 2022, 194, 708–721. [Google Scholar] [CrossRef]
- Monje, O.; Stutte, G.; Goins, G.D.; Porterfield, D.; Bingham, G. Farming in space: Environmental and biophysical concerns. Adv. Space Res. 2003, 31, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Galieni, A.; Falcinelli, B.; Stagnari, F.; Datti, A.; Benincasa, P. Sprouts and microgreens: Trends, opportunities, and horizons for novel research. Agronomy 2020, 10, 1424. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Z.; Ager, E.; Kong, L.; Tan, L. Nutritional quality and health benefits of microgreens, a crop of modern agriculture. J. Future Foods 2021, 1, 58–66. [Google Scholar] [CrossRef]
- Bulgari, R.; Baldi, A.; Ferrante, A.; Lenzi, A. Yield and quality of basil, Swiss chard, and rocket microgreens grown in a hydroponic system. N. Z. J. Crop Hortic. Sci. 2017, 45, 119–129. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Ciriello, M.; Formisano, L.; Pannico, A.; Giordano, M.; Gentile, B.R.; Fusco, G.M.; Kyriacou, M.C.; Carillo, P.; Rouphael, Y. Protein hydrolysate combined with hydroponics divergently modifies growth and shuffles pigments and free amino acids of carrot and dill microgreens. Horticulturae 2021, 7, 279. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; De Pascale, S.; Kyratzis, A.; Rouphael, Y. Microgreens as a component of space life support systems: A cornucopia of functional food. Front. Plant Sci. 2017, 8, 1587. [Google Scholar] [CrossRef]
- Mortazavi, S.; Foadi, M.; Mozdarani, H.; Haghani, M.; Mosleh, S.M.; Abolghasemi, P.; Nematollahi, S.; Sharifzadeh, S. Future role of vitamin C in radiation mitigation and its possible applications in manned deep space missions: Survival study and the measurement of cell viability. Phys. Medica 2015, 13, 55–60. [Google Scholar] [CrossRef]
- de la Fuente, B.; López-García, G.; Máñez, V.; Alegría, A.; Barberá, R.; Cilla, A. Evaluation of the bioaccessibility of antioxidant bioactive compounds and minerals of four genotypes of Brassicaceae microgreens. Foods 2019, 8, 250. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef]
- Xiao, Z.; Codling, E.E.; Luo, Y.; Nou, X.; Lester, G.E.; Wang, Q. Microgreens of Brassicaceae: Mineral composition and content of 30 varieties. J. Food Compos. Anal. 2016, 49, 87–93. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Palladino, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Phenolic constitution, phytochemical and macronutrient content in three species of microgreens as modulated by natural fiber and synthetic substrates. Antioxidants 2020, 9, 252. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Jankauskienė, J.; Viršilė, A.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskaitė, J.; Duchovskis, P. LED irradiance level affects growth and nutritional quality of Brassica microgreens. Cent. Eur. J. Biol. 2013, 8, 1241–1249. [Google Scholar] [CrossRef]
- Teng, Z.; Luo, Y.; Pearlstein, D.J.; Wheeler, R.M.; Johnson, C.M.; Wang, Q.; Fonseca, J.M. Microgreens for Home, Commercial, and Space Farming: A Comprehensive Update of the Most Recent Developments. Annu. Rev. Food Sci. Technol. 2023, 14, 539–562. [Google Scholar] [CrossRef]
- Eftychia, G.K.; Roupas, N.D.; Markou, K.B. Effect of excess iodine intake on thyroid on human health. Minerva Med. 2017, 108, 136–146. [Google Scholar]
- Krzepilko, A.; Prazak, R.; Skwarylo-Bednarz, B.; Molas, J. Agronomic biofortification as a means of enriching plant foodstuffs with iodine. Acta Agrobot. 2019, 72, 1766. [Google Scholar] [CrossRef]
- Hays, S.M.; Poddalgoda, D.; Macey, K.; Aylward, L.; Nong, A. Biomonitoring Equivalents for interpretation of urinary iodine. Regul. Toxicol. Pharmacol. 2018, 94, 40–46. [Google Scholar] [CrossRef]
- Doggui, R.; El Atia, J. Iodine deficiency: Physiological, clinical and epidemiological features, and pre-analytical considerations. Ann. D’endocrinologie 2015, 76, 59–66. [Google Scholar] [CrossRef]
- Ershow, A.G.; Skeaff, S.A.; Merkel, J.M.; Pehrsson, P.R. Development of databases on iodine in foods and dietary supplements. Nutrients 2018, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- van der Reijden, O.L.; Zimmermann, M.B.; Galetti, V. Iodine in dairy milk: Sources, concentrations and importance to human health. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium biofortification impacts the nutritive value, polyphenolic content, and bioactive constitution of variable microgreens genotypes. Antioxidants 2020, 9, 272. [Google Scholar] [CrossRef] [PubMed]
- Ciriello, M.; Formisano, L.; El-Nakhel, C.; Zarrelli, A.; Giordano, M.; De Pascale, S.; Kyriacou, M.; Rouphael, Y. Iodine biofortification of four microgreens species and its implications for mineral composition and potential contribution to the recommended dietary intake of iodine. Sci. Hortic. 2023, 320, 112229. [Google Scholar] [CrossRef]
- Golob, A.; Novak, T.; Maršić, N.K.; Šircelj, H.; Stibilj, V.; Jerše, A.; Kroflič, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes) and increases their contents in tubers. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Smoleń, S.; Sady, W. Influence of iodine form and application method on the effectiveness of iodine biofortification, nitrogen metabolism as well as the content of mineral nutrients and heavy metals in spinach plants (Spinacia oleracea L.). Sci. Hortic. 2012, 143, 176–183. [Google Scholar] [CrossRef]
- Gonnella, M.; Renna, M.; D’Imperio, M.; Santamaria, P.; Serio, F. Iodine biofortification of four brassica genotypes is effective already at low rates of potassium iodate. Nutrients 2019, 11, 451. [Google Scholar] [CrossRef]
- Smoleń, S.; Kowalska, I.; Sady, W. Assessment of biofortification with iodine and selenium of lettuce cultivated in the NFT hydroponic system. Sci. Hortic. 2014, 166, 9–16. [Google Scholar] [CrossRef]
- Blasco, B.; Rios, J.; Cervilla, L.; Sánchez-Rodrigez, E.; Ruiz, J.; Romero, L. Iodine biofortification and antioxidant capacity of lettuce: Potential benefits for cultivation and human health. Ann. Appl. Biol. 2008, 152, 289–299. [Google Scholar] [CrossRef]
- Blasco, B.; Rios, J.; Leyva, R.; Melgarejo, R.; Constán-Aguilar, C.; Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.; Romero, L.; Ruiz, J. Photosynthesis and metabolism of sugars from lettuce plants (Lactuca sativa L. var. longifolia) subjected to biofortification with iodine. Plant Growth Regul. 2011, 65, 137–143. [Google Scholar] [CrossRef]
- Blasco, B.; Rios, J.J.; Cervilla, L.M.; Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.M.; Rosales, M.A.; Romero, L.; Ruiz, J.M. Iodine application affects nitrogen-use efficiency of lettuce plants (Lactuca sativa L.). Acta Agric. Scand. Sect. B-Soil Plant Sci. 2011, 61, 378–383. [Google Scholar]
- Germ, M.; Stibilj, V.; Šircelj, H.; Jerše, A.; Kroflič, A.; Golob, A.; Maršić, N.K. Biofortification of common buckwheat microgreens and seeds with different forms of selenium and iodine. J. Sci. Food Agric. 2019, 99, 4353–4362. [Google Scholar] [CrossRef]
- Puccinelli, M.; Landi, M.; Maggini, R.; Pardossi, A.; Incrocci, L. Iodine biofortification of sweet basil and lettuce grown in two hydroponic systems. Sci. Hortic. 2021, 276, 109783. [Google Scholar] [CrossRef]
- Kiferle, C.; Martinelli, M.; Salzano, A.M.; Gonzali, S.; Beltrami, S.; Salvadori, P.A.; Hora, K.; Holwerda, H.T.; Scaloni, A.; Perata, P. Evidences for a nutritional role of iodine in plants. Front. Plant Sci. 2021, 12, 616868. [Google Scholar] [CrossRef]
- Dai, J.-L.; Zhu, Y.-G.; Zhang, M.; Huang, Y.-Z. Selecting iodine-enriched vegetables and the residual effect of iodate application to soil. Biol. Trace Elem. Res. 2004, 101, 265–276. [Google Scholar] [CrossRef]
- Strzelecki, P.; Smoleń, S.; Rożek, S.; Sady, W.o. Effect of differentiated fertilization and foliar application of iodine on yielding and antioxidant properties in radish (Raphanus sativus L.) plants. Ecol. Chem. Eng. A 2010, 17, 1189–1196. [Google Scholar]
- Smoleń, S.; Ledwożyw-Smoleń, I.; Sady, W. The role of exogenous humic and fulvic acids in iodine biofortification in spinach (Spinacia oleracea L.). Plant Soil 2016, 402, 129–143. [Google Scholar] [CrossRef]
- Weng, H.-X.; Yan, A.-L.; Hong, C.-L.; Xie, L.-L.; Qin, Y.-C.; Cheng, C.Q. Uptake of different species of iodine by water spinach and its effect to growth. Biol. Trace Elem. Res. 2008, 124, 184–194. [Google Scholar] [CrossRef]
- Kiferle, C.; Gonzali, S.; Holwerda, H.T.; Ibaceta, R.R.; Perata, P. Tomato fruits: A good target for iodine biofortification. Front. Plant Sci. 2013, 4, 205. [Google Scholar] [CrossRef]
- Landini, M.; Gonzali, S.; Perata, P. Iodine biofortification in tomato. J. Plant Nutr. Soil Sci. 2011, 174, 480–486. [Google Scholar] [CrossRef]
- Smoleń, S.; Baranski, R.; Ledwożyw-Smoleń, I.; Skoczylas, Ł.; Sady, W. Combined biofortification of carrot with iodine and selenium. Food Chem. 2019, 300, 125202. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, M.; Carroll, Y.; Corridan, B.; Olmedilla, B.; Granado, F.; Blanco, I.; Van den Berg, H.; Hininger, I.; Rousell, A.-M.; Chopra, M. A European carotenoid database to assess carotenoid intakes and its use in a five-country comparative study. Br. J. Nutr. 2001, 85, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Rapp, L.M.; Maple, S.S.; Choi, J.H. Lutein and zeaxanthin concentrations in rod outer segment membranes from perifoveal and peripheral human retina. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1200–1209. [Google Scholar]
- Naves, M.M.V.; Moreno, F.S. β-carotene and cancer chemoprevention: From epidemiological associations to cellular mechanisms of action. Nutr. Res. 1998, 18, 1807–1824. [Google Scholar] [CrossRef]
- Smoleń, S.; Sady, W.; Ledwożyw-Smoleń, I.; Strzetelski, P.; Liszka-Skoczylas, M.; Rożek, S. Quality of fresh and stored carrots depending on iodine and nitrogen fertilization. Food Chem. 2014, 159, 316–322. [Google Scholar] [CrossRef]
- Osuna, H.T.G.; Mendoza, A.B.; Morales, C.R.; Rubio, E.M.; Star, J.V.; Ruvalcaba, R.M. Iodine application increased ascorbic acid content and modified the vascular tissue in Opuntia Ficus-Indica L. Pak. J. Bot. 2014, 46, 127–134. [Google Scholar]
- Incrocci, L.; Carmassi, G.; Maggini, R.; Poli, C.; Saidov, D.; Tamburini, C.; Kiferle, C.; Perata, P.; Pardossi, A. Iodine accumulation and tolerance in sweet basil (Ocimum basilicum L.) with green or purple leaves grown in floating system technique. Front. Plant Sci. 2019, 10, 1494. [Google Scholar] [CrossRef]
- Kiferle, C.; Ascrizzi, R.; Martinelli, M.; Gonzali, S.; Mariotti, L.; Pistelli, L.; Flamini, G.; Perata, P. Effect of Iodine treatments on Ocimum basilicum L.: Biofortification, phenolics production and essential oil composition. PLoS ONE 2019, 14, e0226559. [Google Scholar] [CrossRef]
- Blasco, B.; Ríos, J.J.; Leyva, R.; Cervilla, L.M.; Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.M.; Rosales, M.A.; Ruiz, J.M.; Romero, L. Does iodine biofortification affect oxidative metabolism in lettuce plants? Biol. Trace Elem. Res. 2011, 142, 831–842. [Google Scholar] [CrossRef]
- Crucian, B.E.; Choukèr, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A. Immune system dysregulation during spaceflight: Potential countermeasures for deep space exploration missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef]
- Xiao, Z.; Lester, G.E.; Park, E.; Saftner, R.A.; Luo, Y.; Wang, Q. Evaluation and correlation of sensory attributes and chemical compositions of emerging fresh produce: Microgreens. Postharvest Biol. Technol. 2015, 110, 140–148. [Google Scholar] [CrossRef]
- Smoleń, S.; Ledwożyw-Smoleń, I.; Halka, M.; Sady, W.; Kováčik, P. The absorption of iodine from 5-iodosalicylic acid by hydroponically grown lettuce. Sci. Hortic. 2017, 225, 716–725. [Google Scholar] [CrossRef]
- Smoleń, S.; Wierzbińska, J.; Sady, W.; Kołton, A.; Wiszniewska, A.; Liszka-Skoczylas, M. Iodine biofortification with additional application of salicylic acid affects yield and selected parameters of chemical composition of tomato fruits (Solanum lycopersicum L.). Sci. Hortic. 2015, 188, 89–96. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Formisano, L.; Ciriello, M.; El-Nakhel, C.; Poledica, M.; Starace, G.; Graziani, G.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Pearl Grey Shading Net Boosts the Accumulation of Total Carotenoids and Phenolic Compounds that Accentuate the Antioxidant Activity of Processing Tomato. Antioxidants 2021, 10, 1999. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Fresh Biomass | Dry Biomass | Hypocotyl Length |
|---|---|---|---|
| kg m−2 | cm | ||
| Species (S) | |||
| Tatsoi | 1.52 ± 0.03 b | 0.11 ± 0.00 b | 3.18 ± 0.03 c |
| Coriander | 1.78 ± 0.03 a | 0.20 ± 0.00 a | 4.30 ± 0.06 a |
| Green basil | 1.10 ± 0.03 c | 0.10 ± 0.00 c | 3.79 ± 0.03 b |
| Purple basil | 1.04 ± 0.02 d | 0.09 ± 0.00 d | 4.24 ± 0.08 a |
| *** | *** | *** | |
| Biofortification (B) | |||
| Control | 1.44 ± 0.09 a | 0.13 ± 0.01 a | 3.91 ± 0.16 |
| 4 µM | 1.31 ± 0.10 b | 0.12 ± 0.01 b | 3.81 ± 0.13 |
| 8 µM | 1.34 ± 0.09 b | 0.12 ± 0.01 b | 3.90 ± 0.14 |
| *** | *** | n.s. | |
| S × B | |||
| Tatsoi × control | 1.60 ± 0.01 c | 0.12 ± 0.00 | 3.17 ± 0.03 d |
| Tatsoi × 4 µM | 1.39 ± 0.04 d | 0.11 ± 0.00 | 3.13 ± 0.10 d |
| Tatsoi × 8 µM | 1.58 ± 0.01 c | 0.12 ± 0.00 | 3.23 ± 0.03 d |
| Coriander × control | 1.85 ± 0.02 a | 0.20 ± 0.00 | 4.20 ± 0.12 ab |
| Coriander × 4 µM | 1.81 ± 0.06 ab | 0.19 ± 0.01 | 4.21 ± 0.03 ab |
| Coriander × 8 µM | 1.69 ± 0.04 bc | 0.19 ± 0.00 | 4.49 ± 0.09 a |
| Green basil × control | 1.20 ± 0.03 e | 0.11 ± 0.00 | 3.76 ± 0.05 c |
| Green basil × 4 µM | 1.03 ± 0.03 f | 0.10 ± 0.00 | 3.79 ± 0.01 c |
| Green basil × 8 µM | 1.09 ± 0.00 ef | 0.10 ± 0.00 | 3.81 ± 0.10 c |
| Purple basil × control | 1.10 ± 0.01 ef | 0.09 ± 0.00 | 4.52 ± 0.06 a |
| Purple basil × 4 µM | 0.99 ± 0.01 f | 0.09 ± 0.00 | 4.11 ± 0.09 bc |
| Purple basil × 8 µM | 1.02 ± 0.01 f | 0.09 ± 0.00 | 4.09 ± 0.04 bc |
| *** | n.s. | ** | |
| Treatment | Caffeic Acid Hexoside | Coumaroyl Quinic Acid | Sinapinic Acid Hexose | Feruloyl Quinic Acid | Caffeoyl Quinic Acid | Caffeic Acid |
|---|---|---|---|---|---|---|
| µg 100 g−1 fw | ||||||
| Species (S) | ||||||
| Tatsoi | 85.26 ± 5.43 a | 21.84 ± 1.58 a | 168.24 ± 4.76 a | 5.38 ± 0.34 a | 10.12 ± 0.60 b | 4.02 ± 0.22 c |
| Coriander | 9.72 ± 0.45 b | 0.17 ± 0.04 b | 17.45 ± 0.67 b | 662.51 ± 17.73 c | 3600.43 ± 182.27 a | 6.15 ± 0.27 c |
| Green basil | 9.03 ± 0.61 b | 0.22 ± 0.02 b | 16.4 ± 0.54 b | 6.33 ± 0.53 c | 62.38 ± 10.07 b | 36.20 ± 2.81 a |
| Purple basil | 13.54 ± 0.34 b | 0.23 ± 0.03 b | 9.55 ± 0.23 c | 48.95 ± 1.91 b | 18.30 ± 0.95 b | 25.11 ± 0.97 b |
| *** | *** | *** | *** | *** | *** | |
| Biofortification (B) | ||||||
| Control | 25.71 ± 7.66 b | 4.42 ± 2.21 b | 51.18 ± 19.28 b | 167.73 ± 76.75 b | 926.29 ± 464.57 | 16.13 ± 3.36 |
| 4 µM | 32.40 ± 11.57 a | 6.09 ± 3.12 b | 57.50 ± 22.43 a | 187.52 ± 88.09 a | 868.86 ± 449.57 | 18.31 ± 4.53 |
| 8 µM | 30.05 ± 10.31 ab | 8.39 ± 4.14 a | 50.05 ± 18.62 b | 187.13 ± 87.64 a | 973.27 ± 501.64 | 19.17 ± 4.65 |
| * | * | *** | * | n.s. | n.s. | |
| S × B | ||||||
| Tatsoi × control | 69.25 ± 4.40 b | 16.92 ± 1.70 b | 161.79 ± 1.79 b | 4.45 ± 0.32 c | 11.65 ± 0.70 | 4.24 ± 0.53 c |
| Tatsoi × 4 µM | 97.85 ± 9.23 a | 23.82 ± 2.06 a | 186.18 ± 2.30 a | 6.25 ± 0.42 c | 10.29 ± 0.63 | 4.28 ± 0.33 c |
| Tatsoi × 8 µM | 88.66 ± 5.99 a | 24.78 ± 2.12 a | 156.74 ± 3.93 b | 5.44 ± 0.56 c | 8.43 ± 0.88 | 3.52 ± 0.14 c |
| Coriander × control | 9.18 ± 0.81 c | 0.25 ± 0.01 c | 16.14 ± 0.61 cd | 607.27 ± 8.16 b | 3574.32 ± 266.25 | 6.03 ± 0.26 c |
| Coriander × 4 µM | 9.47 ± 1.14 c | 0.09 ± 0.00 c | 18.14 ± 1.74 c | 691.84 ± 25.46 a | 3401.9 ± 410.60 | 5.77 ± 0.77 c |
| Coriander × 8 µM | 10.51 ± 0.17 c | nd | 18.08 ± 0.86 c | 688.41 ± 27.70 a | 3825.07 ± 337.51 | 6.63 ± 0.20 c |
| Green basil × control | 10.95 ± 0.25 c | 0.29 ± 0.02 c | 17.21 ± 0.99 cd | 7.17 ± 1.09 c | 98.91 ± 13.69 | 27.60 ± 2.10 b |
| Green basil × 4 µM | 9.11 ± 0.13 c | 0.13 ± 0.00 c | 16.52 ± 0.34 cd | 6.99 ± 0.68 c | 46.91 ± 2.68 | 39.28 ± 5.64 a |
| Green basil × 8 µM | 7.03 ± 0.70 c | 0.25 ± 0.00 c | 15.47 ± 1.30 cd | 4.81 ± 0.05 c | 41.33 ± 3.60 | 41.73 ± 1.33 a |
| Purple basil × control | 13.45 ± 0.70 c | 0.23 ± 0.00 c | 9.58 ± 0.33 d | 52.01 ± 4.13 c | 20.28 ± 0.35 | 26.65 ± 0.55 b |
| Purple basil × 4 µM | 13.18 ± 0.29 c | 0.33 ± 0.01 c | 9.17 ± 0.35 d | 45.00 ± 2.88 c | 16.34 ± 1.15 | 23.89 ± 2.58 b |
| Purple basil × 8 µM | 14.00 ± 0.79 c | 0.14 ± 0.00 c | 9.91 ± 0.49 cd | 49.85 ± 2.30 c | 18.26 ± 2.37 | 24.79 ± 1.52 b |
| ** | ** | *** | ** | n.s. | ** | |
| Treatment | Ferulic Acid | Rosmarinic Acid | Caffeoyl–Feruloyl–Tartaric Acid | Cichoric Acid | Total Phenolic Acids | |
| µg 100 g−1 fw | ||||||
| Species (S) | ||||||
| Tatsoi | 0.97 ± 0.03 d | nd | nd | nd | 295.82 ± 9.80 c | |
| Coriander | 65.74 ± 2.02 a | nd | nd | nd | 4362.10 ± 180.80 b | |
| Green basil | 14.37 ± 1.33 b | 14,696.01 ± 606.65 | 325.85 ± 15.49 a | 1249.37 ± 105.63 | 16,416.16 ± 609.75 a | |
| Purple basil | 6.95 ± 0.38 c | 14,274.07 ± 776.5 | 199.22 ± 9.25 b | 1135.39 ± 99.57 | 15,731.32 ± 798.21 a | |
| *** | n.s. | *** | n.s. | *** | ||
| Biofortification (B) | ||||||
| Control | 22.12 ± 7.80 | 12,613.59 ± 179.26 b | 281.66 ± 35.25 | 1335.59 ± 127.16 | 8328.99 ± 1880.87 | |
| 4 µM | 22.19 ± 7.42 | 14,945.96 ± 1028.18 a | 240.86 ± 27.82 | 1097.97 ± 119.92 | 9335.28 ± 2240.83 | |
| 8 µM | 21.70 ± 8.20 | 15,895.57 ± 356.20 a | 265.10 ± 31.65 | 1143.58 ± 124.94 | 9939.79 ± 2312.86 | |
| n.s. | ** | n.s. | n.s. | * | ||
| S × B | ||||||
| Tatsoi × control | 1.03 ± 0.07 | nd | nd | nd | 269.33 ± 6.56 d | |
| Tatsoi × 4 µM | 0.90 ± 0.05 | nd | nd | nd | 329.57 ± 9.61 d | |
| Tatsoi × 8 µM | 0.98 ± 0.03 | nd | nd | nd | 288.56 ± 8.34 d | |
| Coriander × control | 65.76 ± 4.34 | nd | nd | nd | 4278.96 ± 261.22 c | |
| Coriander × 4 µM | 63.18 ± 3.29 | nd | nd | nd | 4190.39 ± 385.82 c | |
| Coriander × 8 µM | 68.27 ± 3.58 | nd | nd | nd | 4616.96 ± 352.85 c | |
| Green basil × control | 15.04 ± 0.49 | 12,780.35 ± 293.32 | 355.25 ± 26.44 | 1431.68 ± 200.64 | 14,744.46 ± 491.6 ab | |
| Green basil × 4 µM | 18.47 ± 0.57 | 16,063.09 ± 1111.59 | 294.38 ± 30.83 | 1171.19 ± 235.18 | 17,666.06 ± 1313.64 a | |
| Green basil × 8 µM | 9.59 ± 0.72 | 15,244.58 ± 418.17 | 327.93 ± 17.38 | 1145.25 ± 115.00 | 16,837.97 ± 476.27 ab | |
| Purple basil × control | 6.65 ± 0.80 | 12,446.82 ± 216.39 | 208.06 ± 9.89 | 1239.51 ± 177.09 | 14,023.23 ± 217.80 b | |
| Purple basil × 4 µM | 6.22 ± 0.47 | 13,828.84 ± 1673.96 | 187.34 ± 7.42 | 1024.76 ± 106.00 | 15,155.07 ± 1781.08 ab | |
| Purple basil × 8 µM | 7.97 ± 0.23 | 16,546.56 ± 189.04 | 202.26 ± 27.56 | 1141.90 ± 254.60 | 18,015.65 ± 438.35 a | |
| n.s. | n.s. | n.s. | n.s. | * | ||
| Treatment | Luteolin-7-O-glucoside | Qn 3-sophoroside-7-glucoside | Km 3-sophoroside-7-glucoside | Qn 3-caffeoyl-sophoroside-7-glucoside | Km 3-sinapoyl-sophoroside-7-glucoside | Km 3-sinapoyl-sophorotrioside-7-glucoside | Km 3-O-caffeoyl-spophoroside-7-glucoside | Quercetin-3-sophoroside | Isorhamnetin-3-gentiobioside |
|---|---|---|---|---|---|---|---|---|---|
| µg 100 g−1 fw | |||||||||
| Species (S) | |||||||||
| Tatsoi | nd | 0.40 ± 0.03 | 1.6 ± 0.15 a | 6.14 ± 0.49 b | 88.29 ± 2.35 a | 0.47 ± 0.06 | 36.7 ± 2.06 a | 1.34 ± 0.18 b | 4.94 ± 0.18 |
| Coriander | nd | nd | 1.00 ± 0.18 b | 29.76 ± 1.74 a | 0.71 ± 0.11 b | nd | nd | 2.05 ± 0.18 a | nd |
| Green basil | 1.37 ± 0.15 a | nd | nd | nd | nd | nd | nd | nd | nd |
| Purple basil | 0.86 ± 0.11 b | nd | nd | nd | 1.97 ± 0.12 b | nd | 0.57 ± 0.05 b | 1.22 ± 0.13 b | nd |
| *** | - | ** | *** | *** | - | *** | ** | - | |
| Biofortification (B) | |||||||||
| Control | 1.58 ± 0.17 a | 0.32 ± 0.03 b | 1.23 ± 0.33 | 16.63 ± 5.73 | 32.47 ± 15.65 | 0.33 ± 0.01 b | 15.59 ± 6.70 b | 1.61 ± 0.23 | 5.26 ± 0.23 |
| 4 µM | 0.95 ± 0.09 b | 0.41 ± 0.02 ab | 1.17 ± 0.22 | 18.60 ± 5.70 | 29.20 ± 13.98 | 0.41 ± 0.01 b | 19.45 ± 8.60 ab | 1.43 ± 0.20 | 4.49 ± 0.28 |
| 8 µM | 0.81 ± 0.11 b | 0.48 ± 0.03 a | 1.49 ± 0.14 | 18.62 ± 5.06 | 29.30 ± 13.94 | 0.67 ± 0.07 a | 20.85 ± 9.17 a | 1.58 ± 0.19 | 5.07 ± 0.33 |
| *** | * | n.s. | n.s. | n.s. | ** | * | n.s. | n.s. | |
| S×B | |||||||||
| Tatsoi × control | nd | nd | 1.92 ± 0.17 a | 4.75 ± 0.33 b | 95.04 ± 1.27 a | nd | 30.57 ± 0.54 b | 0.97 ± 0.11 | nd |
| Tatsoi × 4 µM | nd | nd | 1.50 ± 0.35 abc | 6.00 ± 0.44 b | 85.01 ± 3.18 b | nd | 38.37 ± 3.49 ab | 1.38 ± 0.42 | nd |
| Tatsoi × 8 µM | nd | nd | 1.37 ± 0.23 abc | 7.67 ± 0.63 b | 84.81 ± 4.49 b | nd | 41.15 ± 2.88 a | 1.68 ± 0.25 | nd |
| Coriander × control | nd | nd | 0.55 ± 0.23 c | 28.51 ± 4.81 a | 0.44 ± 0.20 c | nd | nd | 2.38 ± 0.17 | nd |
| Coriander × 4 µM | nd | nd | 0.85 ± 0.08 bc | 31.21 ± 1.89 a | 0.72 ± 0.13 c | nd | nd | 1.90 ± 0.25 | nd |
| Coriander × 8 µM | nd | nd | 1.61 ± 0.17 ab | 29.57 ± 2.79 a | 0.97 ± 0.13 c | nd | nd | 1.87 ± 0.48 | nd |
| Green basil × control | 1.94 ± 0.02 | nd | nd | nd | nd | nd | nd | nd | nd |
| Green basil × 4 µM | 1.11 ± 0.09 | nd | nd | nd | nd | nd | nd | nd | nd |
| Green basil × 8 µM | 1.04 ± 0.01 | nd | nd | nd | nd | nd | nd | nd | nd |
| Purple basil × control | 1.22 ± 0.14 | nd | nd | nd | 1.92 ± 0.12 c | nd | 0.62 ± 0.07 c | 1.48 ± 0.32 | nd |
| Purple basil × 4 µM | 0.79 ± 0.05 | nd | nd | nd | 1.88 ± 0.32 c | nd | 0.53 ± 0.14 c | 1.00 ± 0.13 | nd |
| Purple basil × 8 µM | 0.58 ± 0.07 | nd | nd | nd | 2.11 ± 0.24 c | nd | 0.55 ± 0.07 c | 1.19 ± 0.16 | nd |
| n.s. | - | * | n.s. | * | - | * | n.s. | - | |
| Treatment | Km 3-feruloylsophoroside-7-glucoside | Hyperoside | Rutin | Quercetin-3-glucoside | Km 3-p-coumaroylsophoroside-7-glucoside | Luteolin-malonil-hexose | Quercetin Rhamnoside | ||
| µg 100 g−1 fw | |||||||||
| Species (S) | |||||||||
| Tatsoi | 13.4 ± 0.55 | nd | 12.86 ± 0.56 b | nd | 12.95 ± 1.10 b | 0.13 ± 0.01 a | 0.71 ± 0.06 c | ||
| Coriander | nd | 26.92 ± 1.86 a | 6430.29 ± 292.91 a | 20.58 ± 1.46 a | nd | 0.08 ± 0.01 bc | 0.46 ± 0.13 c | ||
| Green basil | nd | 1.48 ± 0.13 c | 1.77 ± 0.63 b | 1.37 ± 0.12 c | nd | nd | 12.37 ± 0.65 a | ||
| Purple basil | nd | 18.07 ± 0.59 b | 2.66 ± 0.13 b | 16.91 ± 0.56 b | 31.87 ± 1.30 a | nd | 8.89 ± 0.30 b | ||
| - | *** | *** | *** | *** | *** | *** | |||
| Biofortification (B) | |||||||||
| Control | 12.19 ± 0.96 | 14.89 ± 3.53 | 1347.60 ± 701.30 c | 12.79 ± 2.94 ab | 21.89 ± 5.86 | 0.13 ± 0.02 a | 5.57 ± 1.47 | ||
| 4 µM | 14.55 ± 0.78 | 14.13 ± 3.53 | 2224.83 ± 1110.47 a | 11.45 ± 2.71 b | 22.83 ± 3.78 | 0.10 ± 0.02 b | 5.38 ± 1.59 | ||
| 8 µM | 13.45 ± 0.88 | 17.46 ± 4.45 | 1819.02 ± 947.73 b | 14.62 ± 3.44 a | 22.51 ± 3.46 | 0.09 ± 0.01 b | 5.87 ± 1.70 | ||
| n.s. | n.s. | *** | * | n.s. | ** | n.s. | |||
| S × B | |||||||||
| Tatsoi × control | nd | nd | 11.29 ± 0.78 c | nd | 9.10 ± 1.19 b | 0.17 ± 0.02 a | 0.53 ± 0.02 | ||
| Tatsoi × 4 µM | nd | nd | 14.52 ± 0.55 c | nd | 14.61 ± 0.43 b | 0.14 ± 0.00 a | 0.66 ± 0.01 | ||
| Tatsoi × 8 µM | nd | nd | 12.78 ± 0.49 c | nd | 15.15 ± 1.35 b | 0.08 ± 0.01 bc | 0.95 ± 0.03 | ||
| Coriander × control | nd | 25.74 ± 1.77 | 5373.4 ± 123.58 c | 21.2 ± 1.91 ab | nd | 0.08 ± 0.00 bc | 0.97 ± 0.02 | ||
| Coriander × 4 µM | nd | 23.18 ± 4.03 | 6656.93 ± 255.3 b | 16.50 ± 2.49 b | nd | 0.06 ± 0.01 c | 0.21 ± 0.01 | ||
| Coriander × 8 µM | nd | 31.83 ± 1.48 | 7260.54 ± 142.22 a | 24.03 ± 1.08 a | nd | 0.10 ± 0.01 b | 0.20 ± 0.02 | ||
| Green basil × control | nd | 1.87 ± 0.13 | 3.17 ± 0.20 c | 1.72 ± 0.09 c | nd | nd | 11.13 ± 0.81 | ||
| Green basil × 4 µM | nd | 1.13 ± 0.08 | nd | 1.03 ± 0.04 c | nd | nd | 12.24 ± 1.55 | ||
| Green basil × 8 µM | nd | 1.44 ± 0.21 | 0.37 ± 0.02 c | 1.36 ± 0.19 c | nd | nd | 13.74 ± 0.56 | ||
| Purple basil × control | nd | 17.05 ± 0.43 | 2.55 ± 0.17 c | 15.46 ± 0.38 b | 34.67 ± 2.58 a | nd | 9.66 ± 0.32 | ||
| Purple basil × 4 µM | nd | 18.08 ± 0.53 | 3.05 ± 0.22 c | 16.81 ± 0.60 b | 31.05 ± 1.91 a | nd | 8.41 ± 0.64 | ||
| Purple basil × 8 µM | nd | 19.09 ± 1.62 | 2.39 ± 0.03 c | 18.45 ± 0.99 ab | 29.87 ± 1.94 a | nd | 8.60 ± 0.38 | ||
| - | n.s. | *** | * | * | *** | n.s. | |||
| Treatment | Apigenin-malonil-glucoside | Apigenin-7-rutinoside | Kaempferol-rutinoside | Kaempferol-glucoside | Apigenin-7-rhamnoside-4-rutinoside | Apigenin-7-glucoside | Luteolin-7-rutinoside | Total Flavonoids | Total Polyphenols |
| µg 100 g−1 fw | |||||||||
| Species (S) | |||||||||
| Tatsoi | 0.38 ± 0.03 c | 0.87 ± 0.12 c | 0.49 ± 0.08 b | 0.35 ± 0.04 c | 2.59 ± 0.24 | nd | nd | 184.62 ± 4.66 b | 480.44 ± 12.73 c |
| Coriander | nd | 1.35 ± 0.43 c | 43.55 ± 1.55 a | 0.19 ± 0.02 c | nd | nd | 51.00 ± 2.18 a | 6607.93 ± 296.30 a | 10,970.03 ± 375.70 b |
| Green basil | 6.25 ± 0.39 a | 18.57 ± 0.83 b | 2.33 ± 0.17 b | 8.60 ± 0.29 a | nd | 3.63 ± 0.18 a | 0.23 ± 0.03 b | 57.38 ± 0.88 b | 16,473.54 ± 610.04 a |
| Purple basil | 1.94 ± 0.18 b | 21.44 ± 0.84 a | 2.95 ± 0.18 b | 5.81 ± 0.24 b | nd | 1.29 ± 0.09 b | 1.84 ± 0.23 b | 118.30 ± 2.20 b | 15,849.61 ± 799.34 a |
| *** | *** | *** | *** | - | *** | *** | *** | *** | |
| Biofortification (B) | |||||||||
| Control | 3.51 ± 1.10 a | 9.56 ± 2.41 b | 11.68 ± 5.14 | 3.70 ± 1.06 | 2.14 ± 0.19 b | 2.34 ± 0.46 | 15.37 ± 7.40 b | 1473.35 ± 708.96 b | 9802.34 ± 1733.69 b |
| 4 µM | 2.32 ± 0.75 c | 10.59 ± 3.03 ab | 13.03 ± 5.84 | 3.71 ± 1.11 | 2.13 ± 0.15 b | 2.37 ± 0.54 | 17.72 ± 8.35 ab | 1796.75 ± 877.88 a | 11,132.02 ± 2040.28 a |
| 8 µM | 2.74 ± 0.80 b | 11.52 ± 3.22 a | 12.28 ± 5.44 | 3.80 ± 1.12 | 3.51 ± 0.15 a | 2.67 ± 0.64 | 19.98 ± 9.34 a | 1956.06 ± 957.39 a | 11,895.85 ± 2108.70 a |
| *** | *** | n.s. | n.s. | *** | n.s. | ** | *** | *** | |
| S × B | |||||||||
| Tatsoi × control | 0.29 ± 0.04 e | 0.46 ± 0.03 d | 0.25 ± 0.02 | 0.23 ± 0.00 d | 2.14 ± 0.19 | nd | nd | 175.81 ± 3.30 d | 445.13 ± 5.21 |
| Tatsoi × 4 µM | 0.37 ± 0.02 e | 0.85 ± 0.02 d | 0.45 ± 0.07 | 0.34 ± 0.05 d | 2.13 ± 0.15 | nd | nd | 186.19 ± 8.44 d | 515.77 ± 17.74 |
| Tatsoi × 8 µM | 0.47 ± 0.01 e | 1.30 ± 0.09 d | 0.77 ± 0.07 | 0.49 ± 0.01 d | 3.51 ± 0.15 | nd | nd | 191.85 ± 10.57 d | 480.42 ± 18.87 |
| Coriander × control | nd | 3.05 ± 0.12 d | 40.97 ± 2.60 | 0.24 ± 0.01 d | nd | nd | 44.79 ± 2.94 b | 5542.3 ± 128.63 c | 9821.26 ± 382.44 |
| Coriander × 4 µM | nd | 0.36 ± 0.06 d | 46.27 ± 3.23 | 0.15 ± 0.01 d | nd | nd | 50.93 ± 2.78 ab | 6829.27 ± 257.49 b | 11,019.66 ± 180.17 |
| Coriander × 8 µM | nd | 0.64 ± 0.01 d | 43.41 ± 2.12 | 0.16 ± 0.02 d | nd | nd | 57.27 ± 1.25 a | 7452.21 ± 148.27 a | 12,069.17 ± 499.91 |
| Green basil × control | 7.71 ± 0.21 a | 15.55 ± 0.38 c | 2.77 ± 0.16 | 7.88 ± 0.41 ab | nd | 3.36 ± 0.08 | 0.33 ± 0.02 c | 57.44 ± 1.07 d | 14,801.90 ± 492.56 |
| Green basil × 4 µM | 5.23 ± 0.34 b | 19.77 ± 1.06 b | 2.12 ± 0.26 | 8.84 ± 0.52 a | nd | 3.50 ± 0.37 | 0.23 ± 0.01 c | 55.21 ± 1.74 d | 17,721.28 ± 1315.35 |
| Green basil × 8 µM | 5.81 ± 0.10 b | 20.38 ± 0.33 b | 2.08 ± 0.35 | 9.08 ± 0.39 a | nd | 4.03 ± 0.38 | 0.13 ± 0.01 c | 59.48 ± 0.76 d | 16,897.45 ± 477.02 |
| Purple basil × control | 2.53 ± 0.15 c | 19.18 ± 0.74 b | 2.73 ± 0.39 | 6.46 ± 0.33 bc | nd | 1.31 ± 0.10 | 1.01 ± 0.11 c | 117.85 ± 2.85 d | 14,141.08 ± 216.17 |
| Purple basil × 4 µM | 1.35 ± 0.13 d | 21.37 ± 1.05 ab | 3.26 ± 0.27 | 5.50 ± 0.40 c | nd | 1.24 ± 0.22 | 1.99 ± 0.08 c | 116.32 ± 2.89 d | 15,271.39 ± 1783.96 |
| Purple basil × 8 µM | 1.95 ± 0.13 cd | 23.77 ± 1.26 a | 2.85 ± 0.28 | 5.47 ± 0.28 c | nd | 1.31 ± 0.19 | 2.53 ± 0.05 c | 120.72 ± 6.07 d | 18,136.36 ± 443.46 |
| *** | *** | n.s. | * | - | n.s. | ** | *** | n.s. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, M.; Ciriello, M.; Formisano, L.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Ritieni, A.; Kyriacou, M.C.; Rouphael, Y.; De Pascale, S. Iodine-Biofortified Microgreens as High Nutraceutical Value Component of Space Mission Crew Diets and Candidate for Extraterrestrial Cultivation. Plants 2023, 12, 2628. https://doi.org/10.3390/plants12142628
Giordano M, Ciriello M, Formisano L, El-Nakhel C, Pannico A, Graziani G, Ritieni A, Kyriacou MC, Rouphael Y, De Pascale S. Iodine-Biofortified Microgreens as High Nutraceutical Value Component of Space Mission Crew Diets and Candidate for Extraterrestrial Cultivation. Plants. 2023; 12(14):2628. https://doi.org/10.3390/plants12142628
Chicago/Turabian StyleGiordano, Maria, Michele Ciriello, Luigi Formisano, Christophe El-Nakhel, Antonio Pannico, Giulia Graziani, Alberto Ritieni, Marios C. Kyriacou, Youssef Rouphael, and Stefania De Pascale. 2023. "Iodine-Biofortified Microgreens as High Nutraceutical Value Component of Space Mission Crew Diets and Candidate for Extraterrestrial Cultivation" Plants 12, no. 14: 2628. https://doi.org/10.3390/plants12142628
APA StyleGiordano, M., Ciriello, M., Formisano, L., El-Nakhel, C., Pannico, A., Graziani, G., Ritieni, A., Kyriacou, M. C., Rouphael, Y., & De Pascale, S. (2023). Iodine-Biofortified Microgreens as High Nutraceutical Value Component of Space Mission Crew Diets and Candidate for Extraterrestrial Cultivation. Plants, 12(14), 2628. https://doi.org/10.3390/plants12142628