
Application of SVR-Mediated GWAS for Identification of Durable Genetic Regions Associated with Soybean Seed Quality Traits

 ,
,  , and
, and 
Abstract
:1. Introduction
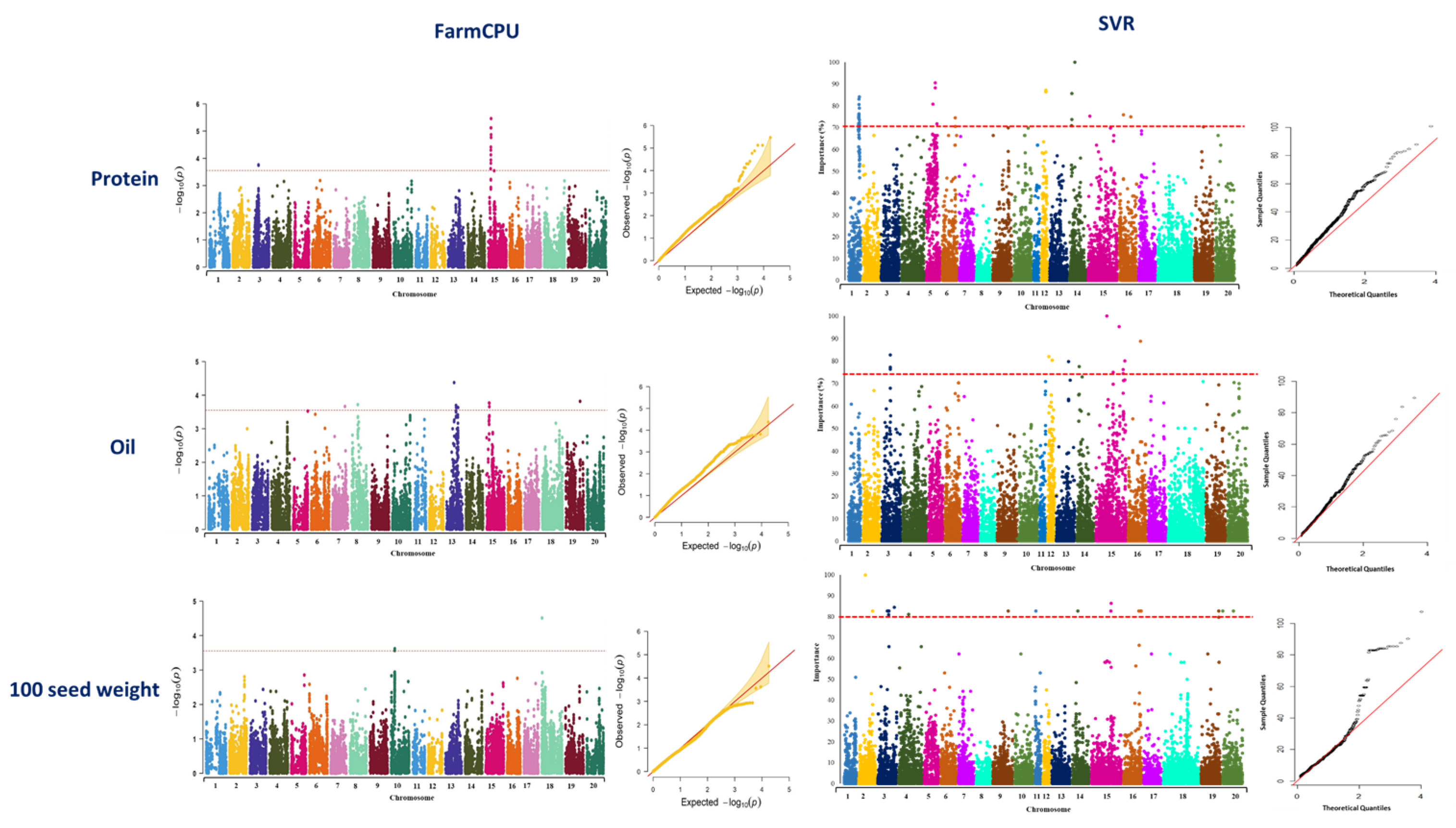
2. Results
2.1. Phenotyping Evaluation
2.1.1. Genotyping
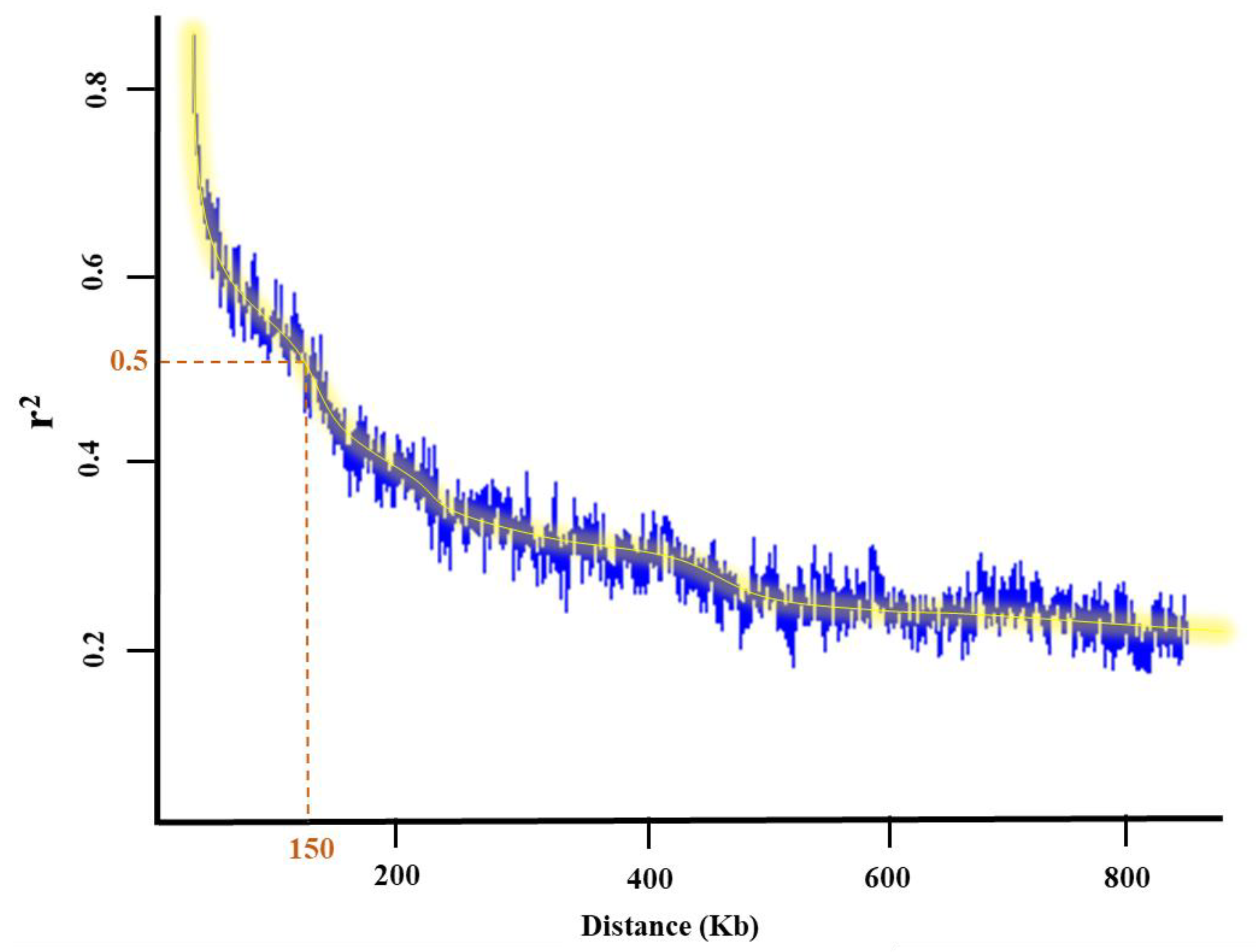
2.1.2. Population Structure and Kinship
2.1.3. GWAS Analysis
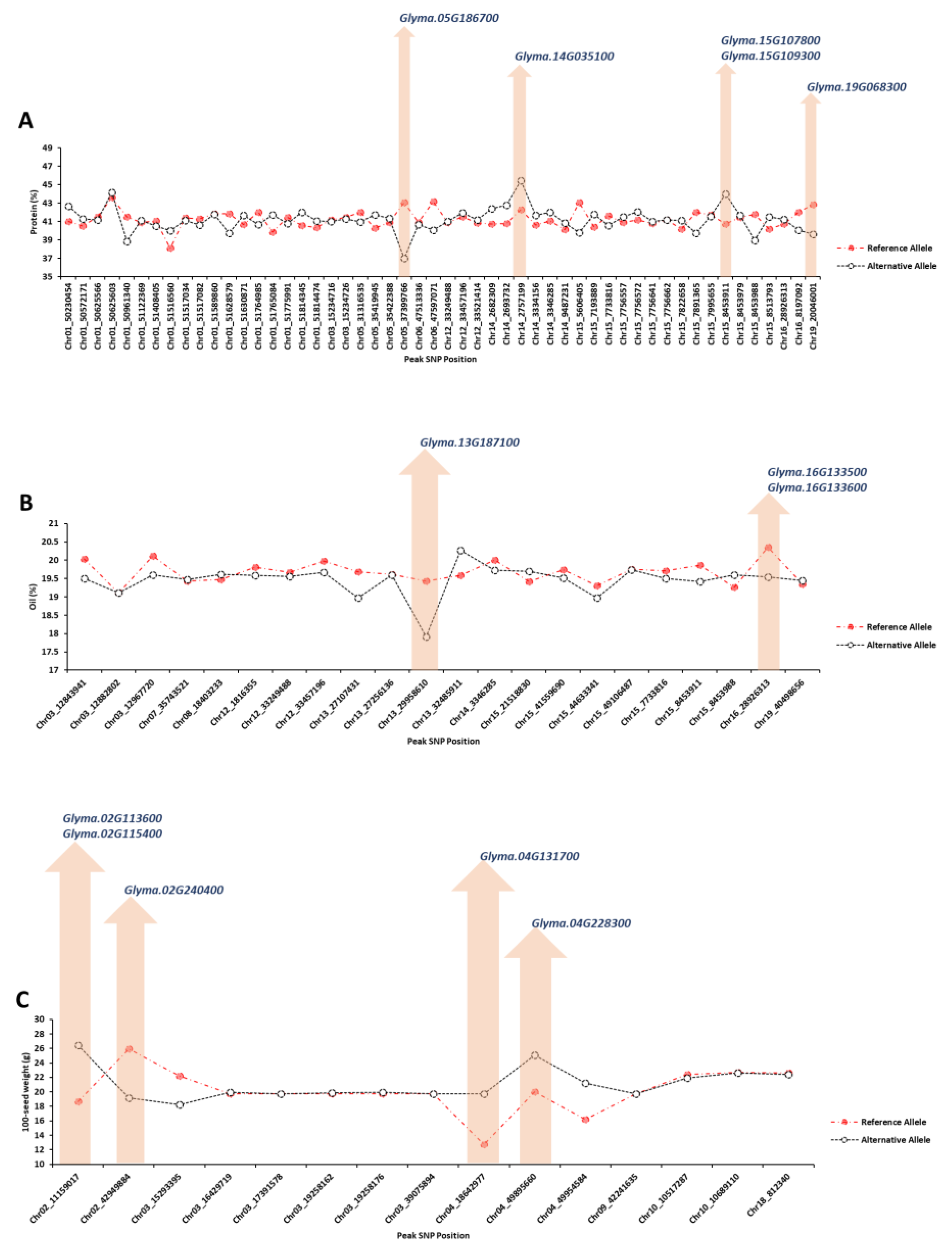
2.1.4. Extracting Candidate Genes Undelaying Detected QTLs
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Field Experiments
4.2. Phenotypic Data and Analysis
4.3. Genotyping
4.4. Analysis of Population Structure
4.5. Association Analysis
4.6. Extracting Candidate Genes Undelaying Detected QTLs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Jin, J.; Wang, G.; Herbert, S.J. Soybean yield physiology and development of high-yielding practices in Northeast China. Field Crops Res. 2008, 105, 157–171. [Google Scholar] [CrossRef]
- Joshi, V.; Kumar, S. Meat Analogues: Plant based alternatives to meat products-A review. Int. J. Food Ferment. Technol. 2015, 5, 107. [Google Scholar] [CrossRef]
- Eskandari, M.; Cober, E.R.; Rajcan, I. Genetic control of soybean seed oil: II. QTL and genes that increase oil concentration without decreasing protein or with increased seed yield. Theor. Appl. Genet. 2013, 126, 1677–1687. [Google Scholar] [CrossRef]
- Hong, H.; Yoosefzadeh Najafabadi, M.; Rajcan, I. Correlations between soybean seed quality traits using a genome-wide association study panel grown in Canadian and Ukrainian mega-environments. Can. J. Plant Sci. 2022, 102, 1040–1052. [Google Scholar] [CrossRef]
- Yoosefzadeh-Najafabadi, M.; Rajcan, I. Six decades of soybean breeding in Ontario, Canada: A tradition of innovation. Can. J. Plant Sci. 2022. [Google Scholar] [CrossRef]
- Zhu, X.; Leiser, W.L.; Hahn, V.; Würschum, T. Identification of seed protein and oil related QTL in 944 RILs from a diallel of early-maturing European soybean. Crop J. 2021, 9, 238–247. [Google Scholar] [CrossRef]
- Hong, H.; Najafabadi, M.Y.; Torkamaneh, D.; Rajcan, I. Identification of quantitative trait loci associated with seed quality traits between Canadian and Ukrainian mega-environments using genome-wide association study. Theor. Appl. Genet. 2022, 135, 2515–2530. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qin, D.; Piersanti, A.; Zhang, Q.; Miceli, C.; Wang, P. Genome-wide association study identifies candidate genes related to oleic acid content in soybean seeds. BMC Plant Biol. 2020, 20, 399. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, T.; Wang, L.; Jiang, B.; Zhen, C.; Yuan, S.; Hou, W.; Wu, C.; Han, T.; Sun, S. A combined linkage and GWAS analysis identifies QTLs linked to soybean seed protein and oil content. Int. J. Mol. Sci. 2019, 20, 5915. [Google Scholar] [CrossRef] [Green Version]
- Bhat, J.A.; Yu, D. High-throughput NGS-based genotyping and phenotyping: Role in genomics-assisted breeding for soybean improvement. Legume Sci. 2021, 3, e81. [Google Scholar] [CrossRef]
- Yoosefzadeh-Najafabadi, M.; Eskandari, M.; Belzile, F.; Torkamaneh, D. Genome-Wide Association Study Statistical Models: A Review. In Genome-Wide Association Studies; Humana: New York, NY, USA, 2022; pp. 43–62. [Google Scholar]
- Alqudah, A.M.; Sallam, A.; Baenziger, P.S.; Börner, A. GWAS: Fast-forwarding gene identification and characterization in temperate Cereals: Lessons from Barley—A review. J. Adv. Res. 2020, 22, 119–135. [Google Scholar] [CrossRef]
- Yoosefzadeh Najafabadi, M.; Hesami, M.; Eskandari, M. Machine Learning-Assisted Approaches in Modernized Plant Breeding Programs. Genes 2023, 14, 777. [Google Scholar] [CrossRef]
- Tibbs Cortes, L.; Zhang, Z.; Yu, J. Status and prospects of genome-wide association studies in plants. Plant Genome 2021, 14, e20077. [Google Scholar] [CrossRef]
- Bush, W.S.; Moore, J.H. Genome-wide association studies. PLoS Comput. Biol. 2012, 8, e1002822. [Google Scholar] [CrossRef] [Green Version]
- Kaler, A.S.; Gillman, J.D.; Beissinger, T.; Purcell, L.C. Comparing different statistical models and multiple testing corrections for association mapping in soybean and maize. Front. Plant Sci. 2020, 10, 1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltaher, S.; Baenziger, P.S.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.; Alqudah, A.M.; Sallam, A. GWAS revealed effect of genotype× environment interactions for grain yield of Nebraska winter wheat. BMC Genom. 2021, 22, 2. [Google Scholar] [CrossRef]
- Li, S.; Zhang, C.; Yang, D.; Lu, M.; Qian, Y.; Jin, F.; Liu, X.; Wang, Y.; Liu, W.; Li, X. Detection of QTNs for kernel moisture concentration and kernel dehydration rate before physiological maturity in maize using multi-locus GWAS. Sci. Rep. 2021, 11, 1764. [Google Scholar] [CrossRef]
- Yoosefzadeh Najafabadi, M. Using Advanced Proximal Sensing and Genotyping Tools Combined with Bigdata Analysis Methods to Improve Soybean Yield. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2021. [Google Scholar]
- Somegowda, V.K.; Rayaprolu, L.; Rathore, A.; Deshpande, S.P.; Gupta, R. Genome-Wide Association Studies (GWAS) for Traits Related to Fodder Quality and Biofuel in Sorghum: Progress and Prospects. Protein Pept. Lett. 2021, 28, 843–854. [Google Scholar] [CrossRef]
- Yoosefzadeh-Najafabadi, M.; Torabi, S.; Tulpan, D.; Rajcan, I.; Eskandari, M. Genome-Wide Association Studies of Soybean Yield-Related Hyperspectral Reflectance Bands Using Machine Learning-Mediated Data Integration Methods. Front. Plant Sci. 2021, 12, 777028. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Gaquerel, E. Next-Generation Mass Spectrometry Metabolomics Revives the Functional Analysis of Plant Metabolic Diversity. Annu. Rev. Plant Biol. 2021, 72, 867–891. [Google Scholar] [CrossRef] [PubMed]
- Yoosefzadeh Najafabadi, M.; Hesami, M.; Rajcan, I. Unveiling the Mysteries of Non-Mendelian Heredity in Plant Breeding. Plants 2023, 12, 1956. [Google Scholar] [CrossRef]
- Leonelli, S. Scientific Research and Big Data; Stanford Encyclopedia of Philosophy: Stanford, CA, USA, 2020. [Google Scholar]
- Yoosefzadeh-Najafabadi, M.; Rajcan, I.; Eskandari, M. Optimizing genomic selection in soybean: An important improvement in agricultural genomics. Heliyon 2022, 8, e11873. [Google Scholar] [CrossRef] [PubMed]
- Nasser, T.; Tariq, R. Big data challenges. J. Comput. Eng. Inf. Technol. 2015, 4, 3. [Google Scholar] [CrossRef]
- Yoosefzadeh-Najafabadi, M.; Singh, K.D.; Pourreza, A.; Sandhu, K.S.; Rajcan, I. Remote and proximal sensing: How far has it come to help plant breeders? In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Lee, S.; Liang, X.; Woods, M.; Reiner, A.S.; Concannon, P.; Bernstein, L.; Lynch, C.F.; Boice, J.D.; Deasy, J.O.; Bernstein, J.L. Machine learning on genome-wide association studies to predict the risk of radiation-associated contralateral breast cancer in the WECARE Study. PLoS ONE 2020, 15, e0226157. [Google Scholar] [CrossRef]
- Yoosefzadeh-Najafabadi, M.; Tulpan, D.; Eskandari, M. Application of machine learning and genetic optimization algorithms for modeling and optimizing soybean yield using its component traits. PLoS ONE 2021, 16, e0250665. [Google Scholar] [CrossRef]
- Pepe, M.; Hesami, M.; Jones, A.M.P. Machine learning-mediated development and optimization of disinfection protocol and scarification method for improved in vitro germination of cannabis seeds. Plants 2021, 10, 2397. [Google Scholar] [CrossRef]
- Vapnik, V.N. The support vector method. In Proceedings of the International Conference on Artificial Neural Networks, Lausanne, Switzerland, 8–10 October 1997; pp. 261–271. [Google Scholar]
- Drucker, H.; Burges, C.J.; Kaufman, L.; Smola, A.; Vapnik, V. Support vector regression machines. Adv. Neural Inf. Process. Syst. 1997, 9, 155–161. [Google Scholar]
- Yoosefzadeh-Najafabadi, M.; Earl, H.J.; Tulpan, D.; Sulik, J.; Eskandari, M. Application of Machine Learning Algorithms in Plant Breeding: Predicting Yield From Hyperspectral Reflectance in Soybean. Front. Plant Sci. 2021, 11, 624273. [Google Scholar] [CrossRef]
- Pirooznia, M.; Han, S.; Lee, R.S. Machine Learning and Network-Driven Integrative Genomics. Front. Genet. 2021, 12, 327. [Google Scholar] [CrossRef]
- Hesami, M.; Yoosefzadeh Najafabadi, M.; Adamek, K.; Torkamaneh, D.; Jones, A.M.P. Synergizing off-target predictions for in silico insights of CENH3 knockout in cannabis through CRISPR/CAS. Molecules 2021, 26, 2053. [Google Scholar] [CrossRef]
- Jafari, M.; Daneshvar, M.H.; Jafari, S.; Hesami, M. Machine learning-assisted in vitro rooting optimization in passiflora caerulea. Forests 2022, 13, 2020. [Google Scholar] [CrossRef]
- Shen, X.; Gong, X.; Cai, Y.; Guo, Y.; Tu, J.; Li, H.; Zhang, T.; Wang, J.; Xue, F.; Zhu, Z.-J. Normalization and integration of large-scale metabolomics data using support vector regression. Metabolomics 2016, 12, 89. [Google Scholar] [CrossRef]
- de Oliveira, F.C.; Borges, C.C.H.; Almeida, F.N.; e Silva, F.F.; da Silva Verneque, R.; da Silva, M.V.G.; Arbex, W. SNPs selection using support vector regression and genetic algorithms in GWAS. BMC Genom. 2014, 15, S4. [Google Scholar] [CrossRef] [Green Version]
- Dhanapal, A.P.; Ray, J.D.; Smith, J.R.; Purcell, L.C.; Fritschi, F.B. Identification of Novel Genomic Loci Associated with Soybean Shoot Tissue Macro and Micronutrient Concentrations. Plant Genome 2018, 11, 170066. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Shi, X.H.; Li, H.H.; Reif, J.C.; Wang, J.J.; Liu, Z.X.; He, S.; Yu, B.S.; Qiu, L.J. Dissecting the genetic basis of resistance to soybean cyst nematode combining linkage and association mapping. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Lü, H.; Chu, S.; Zhang, H.; Zhang, H.; Yang, Y.; Li, H.; Yu, D. The genetic architecture of water-soluble protein content and its genetic relationship to total protein content in soybean. Sci. Rep. 2017, 7, 5053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanapal, A.P.; Ray, J.D.; Singh, S.K.; Hoyos-Villegas, V.; Smith, J.R.; Purcell, L.C.; King, C.A.; Fritschi, F.B. Association mapping of total carotenoids in diverse soybean genotypes based on leaf extracts and high-throughput canopy spectral reflectance measurements. PLoS ONE 2015, 10, e0137213. [Google Scholar] [CrossRef] [Green Version]
- Kaler, A.S.; Dhanapal, A.P.; Ray, J.D.; King, C.A.; Fritschi, F.B.; Purcell, L.C. Genome-wide association mapping of carbon isotope and oxygen isotope ratios in diverse soybean genotypes. Crop Sci. 2017, 57, 3085–3100. [Google Scholar] [CrossRef]
- Zhang, J.; Song, Q.; Cregan, P.B.; Nelson, R.L.; Wang, X.; Wu, J.; Jiang, G.-L. Genome-wide association study for flowering time, maturity dates and plant height in early maturing soybean (Glycine max) germplasm. BMC Genom. 2015, 16, 217. [Google Scholar] [CrossRef] [Green Version]
- Xavier, A.; Hall, B.; Hearst, A.A.; Cherkauer, K.A.; Rainey, K.M. Genetic architecture of phenomic-enabled canopy coverage in Glycine max. Genetics 2017, 206, 1081–1089. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, X.; Lu, Y.; Bhusal, S.J.; Song, Q.; Cregan, P.B.; Yen, Y.; Brown, M.; Jiang, G.-L. Genome-wide Scan for Seed Composition Provides Insights into Soybean Quality Improvement and the Impacts of Domestication and Breeding. Mol. Plant 2018, 11, 460–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Zhang, D.; Zhang, G.; Kan, G.; Hong, D.; Yu, D. Association mapping of yield-related traits and SSR markers in wild soybean (Glycine soja Sieb. and Zucc.). Breed. Sci. 2014, 63, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Sonah, H.; O’Donoughue, L.; Cober, E.; Rajcan, I.; Belzile, F. Identification of loci governing eight agronomic traits using a GBS-GWAS approach and validation by QTL mapping in soya bean. Plant Biotechnol. J. 2015, 13, 211–221. [Google Scholar] [CrossRef]
- Moellers, T.C.; Singh, A.; Zhang, J.; Brungardt, J.; Kabbage, M.; Mueller, D.S.; Grau, C.R.; Ranjan, A.; Smith, D.L.; Chowda-Reddy, R. Main and epistatic loci studies in soybean for Sclerotinia sclerotiorum resistance reveal multiple modes of resistance in multi-environments. Sci. Rep. 2017, 7, 3554. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Tan, R.; Yuan, J.; Bales, C.; Du, W.; Zhang, S.; Chilvers, M.I.; Schmidt, C.; Song, Q.; Cregan, P.B. Genome-wide association mapping of quantitative resistance to sudden death syndrome in soybean. BMC Genom. 2014, 15, 809. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-X.; Lipka, A.E.; Domier, L.L.; Hartman, G.L. Characterization of disease resistance loci in the USDA soybean germplasm collection using genome-wide association studies. Phytopathology 2016, 106, 1139–1151. [Google Scholar] [CrossRef] [Green Version]
- Vuong, T.; Sonah, H.; Meinhardt, C.; Deshmukh, R.; Kadam, S.; Nelson, R.; Shannon, J.; Nguyen, H. Genetic architecture of cyst nematode resistance revealed by genome-wide association study in soybean. BMC Genom. 2015, 16, 593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamidi, S.; Lee, R.K.; Goos, J.R.; McClean, P.E. Genome-wide association studies identifies seven major regions responsible for iron deficiency chlorosis in soybean (Glycine max). PLoS ONE 2014, 9, e107469. [Google Scholar] [CrossRef]
- Mamidi, S.; Chikara, S.; Goos, R.J.; Hyten, D.L.; Annam, D.; Moghaddam, S.M.; Lee, R.K.; Cregan, P.B.; McClean, P.E. Genome-wide association analysis identifies candidate genes associated with iron deficiency chlorosis in soybean. Plant Genome 2011, 4. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Ma, Y.; Wu, S.; Liu, Z.; Wang, Z.; Yang, R.; Hu, G.; Zhou, Z.; Yu, H.; Zhang, M. Genome-wide association studies dissect the genetic networks underlying agronomical traits in soybean. Genome Biol. 2017, 18, 161. [Google Scholar] [CrossRef] [Green Version]
- Kan, G.; Zhang, W.; Yang, W.; Ma, D.; Zhang, D.; Hao, D.; Hu, Z.; Yu, D. Association mapping of soybean seed germination under salt stress. Mol. Genet. Genom. 2015, 290, 2147–2162. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Lal, S. Molecular analysis of soybean varying in water use efficiency using SSRs markers. J. Environ. Biol. 2015, 36, 1011–1016. [Google Scholar]
- Yoosefzadeh-Najafabadi, M.; Eskandari, M.; Torabi, S.; Torkamaneh, D.; Tulpan, D.; Rajcan, I. Machine-Learning-Based Genome-Wide Association Studies for Uncovering QTL Underlying Soybean Yield and Its Components. Int. J. Mol. Sci. 2022, 23, 5538. [Google Scholar] [CrossRef]
- Copley, T.R.; Duceppe, M.-O.; O’Donoughue, L.S. Identification of novel loci associated with maturity and yield traits in early maturity soybean plant introduction lines. BMC Genom. 2018, 19, 167. [Google Scholar] [CrossRef] [Green Version]
- Korir, P.C.; Zhang, J.; Wu, K.; Zhao, T.; Gai, J. Association mapping combined with linkage analysis for aluminum tolerance among soybean cultivars released in Yellow and Changjiang River Valleys in China. Theor. Appl. Genet. 2013, 126, 1659–1675. [Google Scholar] [CrossRef] [PubMed]
- Priolli, R.H.G.; Campos, J.; Stabellini, N.; Pinheiro, J.; Vello, N. Association mapping of oil content and fatty acid components in soybean. Euphytica 2015, 203, 83–96. [Google Scholar] [CrossRef]
- Zhang, S.; Hao, D.; Zhang, S.; Zhang, D.; Wang, H.; Du, H.; Kan, G.; Yu, D. Genome-wide association mapping for protein, oil and water-soluble protein contents in soybean. Mol. Genet. Genom. 2021, 296, 91–102. [Google Scholar] [CrossRef]
- Alaswad, A.A.; Song, B.; Oehrle, N.W.; Wiebold, W.J.; Mawhinney, T.P.; Krishnan, H.B. Development of soybean experimental lines with enhanced protein and sulfur amino acid content. Plant Sci. 2021, 308, 110912. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, P.; Shi, X.; Yang, N.; Yan, L.; Zhao, Q.; Yang, C.; Guan, Y. Primary metabolite contents are correlated with seed protein and oil traits in near-isogenic lines of soybean. Crop J. 2019, 7, 651–659. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, G.; Li, P.; Yang, J.; Guo, L.; Benning, C.; Wang, X.; Zhao, J. Multiple GmWRI1s are redundantly involved in seed filling and nodulation by regulating plastidic glycolysis, lipid biosynthesis and hormone signalling in soybean (Glycine max). Plant Biotechnol. J. 2020, 18, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.; Komatsu, S. Characterization of proteins in soybean roots under flooding and drought stresses. J. Proteom. 2015, 114, 161–181. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, S.; Dubreucq, B.; Miquel, M.; Rochat, C.; Lepiniec, L. Storage reserve accumulation in Arabidopsis: Metabolic and developmental control of seed filling. Arab. Book/Am. Soc. Plant Biol. 2008, 6, e0113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi, M.; Xavier, A.; Beckett, T.; Beyer, S.; Chen, L.; Chikssa, H.; Cross, V.; Moreira, F.F.; French, E.; Gaire, R. Identification, Deployment, and Transferability of Quantitative Trait Loci from Genome-Wide Association Studies in Plants. Curr. Plant Biol. 2020, 24, 100145. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.; Yang, J.; Zhao, T. Dissecting the genetic architecture of seed protein and oil content in soybean from the Yangtze and Huaihe River valleys using multi-locus genome-wide association studies. Int. J. Mol. Sci. 2019, 20, 3041. [Google Scholar] [CrossRef] [Green Version]
- Szymczak, S.; Biernacka, J.M.; Cordell, H.J.; González-Recio, O.; König, I.R.; Zhang, H.; Sun, Y.V. Machine learning in genome-wide association studies. Genet. Epidemiol. 2009, 33, S51–S57. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Bellis, E.S.; Stubblefield, J.; Causey, J.; Qualls, J.; Walker, K.; Huang, X. Minor QTLs mining through the combination of GWAS and machine learning feature selection. BioRxiv 2019, 2019, 712190. [Google Scholar]
- Yoosefzadeh-Najafabadi, M.; Tulpan, D.; Eskandari, M. Using Hybrid Artificial Intelligence and Evolutionary Optimization Algorithms for Estimating Soybean Yield and Fresh Biomass Using Hyperspectral Vegetation Indices. Remote Sens. 2021, 13, 2555. [Google Scholar] [CrossRef]
- González-Camacho, J.M.; Ornella, L.; Pérez-Rodríguez, P.; Gianola, D.; Dreisigacker, S.; Crossa, J. Applications of machine learning methods to genomic selection in breeding wheat for rust resistance. Plant Genome 2018, 11, 170104. [Google Scholar] [CrossRef] [Green Version]
- Hesami, M.; Jones, A.M.P. Modeling and optimizing callus growth and development in Cannabis sativa using random forest and support vector machine in combination with a genetic algorithm. Appl. Microbiol. Biotechnol. 2021, 105, 5201–5212. [Google Scholar] [CrossRef]
- Ziliak, S. P values and the search for significance. Nat. Methods 2017, 14, 3–4. [Google Scholar]
- Di Leo, G.; Sardanelli, F. Statistical significance: P value, 0.05 threshold, and applications to radiomics—Reasons for a conservative approach. Eur. Radiol. Exp. 2020, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, G.; Mian, R.; Vuong, T.; Pantalone, V.; Song, Q.; Chen, P.; Shannon, G.J.; Carter, T.C.; Nguyen, H.T. Molecular mapping and genomics of soybean seed protein: A review and perspective for the future. Theor. Appl. Genet. 2017, 130, 1975–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoosefzadeh-Najafabadi, M.; Rajcan, I.; Vazin, M. High-throughput plant breeding approaches: Moving along with plant-based food demands for pet food industries. Front. Vet. Sci. 2022, 9, 991844. [Google Scholar] [CrossRef]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef]
- Torabi, S.; Sukumaran, A.; Dhaubhadel, S.; Johnson, S.E.; LaFayette, P.; Parrott, W.A.; Rajcan, I.; Eskandari, M. Effects of type I Diacylglycerol O-acyltransferase (DGAT1) genes on soybean (Glycine max L.) seed composition. Sci. Rep. 2021, 11, 2556. [Google Scholar] [CrossRef]
- Napier, J.A.; Haslam, R.P.; Beaudoin, F.; Cahoon, E.B. Understanding and manipulating plant lipid composition: Metabolic engineering leads the way. Curr. Opin. Plant Biol. 2014, 19, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Kanai, M.; Yamada, T.; Hayashi, M.; Mano, S.; Nishimura, M. Soybean (Glycine max L.) triacylglycerol lipase GmSDP1 regulates the quality and quantity of seed oil. Sci. Rep. 2019, 9, 8924. [Google Scholar] [CrossRef] [Green Version]
- Pádua, G.P.d.; Carvalho, M.L.M.D.; França-Neto, J.D.B.; Guerreiro, M.C.; Guimarães, R.M. Response of soybean genotypes to the expression of green seed under temperature and water stresses. Rev. Bras. Sementes 2009, 31, 140–149. [Google Scholar] [CrossRef]
- Veas, R.E.; Ergo, V.V.; Vega, C.R.; Lascano, R.H.; Rondanini, D.P.; Carrera, C.S. Soybean seed growth dynamics exposed to heat and water stress during the filling period under field conditions. J. Agron. Crop Sci. 2021, 208, 472–485. [Google Scholar] [CrossRef]
- Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino acid transporters in plants: Identification and function. Plants 2020, 9, 972. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, G.; Xu, Z.; Li, J.; Sun, M.; Guo, J.; Ji, W. Organization and regulation of soybean SUMOylation system under abiotic stress conditions. Front. Plant Sci. 2017, 8, 1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandasamy, P.; Gyimesi, G.; Kanai, Y.; Hediger, M.A. Amino acid transporters revisited: New views in health and disease. Trends Biochem. Sci. 2018, 43, 752–789. [Google Scholar] [CrossRef]
- Clemente, T.E.; Cahoon, E.B. Soybean oil: Genetic approaches for modification of functionality and total content. Plant Physiol. 2009, 151, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- Ghassemi-Golezani, K.; Farhangi-Abriz, S. Changes in Oil Accumulation and Fatty Acid Composition of Soybean Seeds under Salt Stress in Response to Salicylic Acid and Jasmonic Acid. Russ. J. Plant Physiol. 2018, 65, 229–236. [Google Scholar] [CrossRef]
- Singh, B.; Usha, K. Salicylic acid induced physiological and biochemical changes in wheat seedlings under water stress. Plant Growth Regul. 2003, 39, 137–141. [Google Scholar] [CrossRef]
- Stevenson, D.G.; Doorenbos, R.K.; Jane, J.L.; Inglett, G.E. Structures and functional properties of starch from seeds of three soybean (Glycine max (L.) Merr.) varieties. Starch-Stärke 2006, 58, 509–519. [Google Scholar] [CrossRef]
- Potts, R.O.; Tamada, J.A.; Tierney, M.J. Glucose monitoring by reverse iontophoresis. Diabetes/Metab. Res. Rev. 2002, 18, S49–S53. [Google Scholar] [CrossRef]
- Geigenberger, P.; Stitt, M.; Fernie, A. Metabolic control analysis and regulation of the conversion of sucrose to starch in growing potato tubers. Plant Cell Environ. 2004, 27, 655–673. [Google Scholar] [CrossRef]
- Lee, S.-K.; Jeon, J.-S. Crucial role of inorganic pyrophosphate in integrating carbon metabolism from sucrose breakdown to starch synthesis in rice endosperm. Plant Sci. 2020, 298, 110572. [Google Scholar] [CrossRef] [PubMed]
- Stroup, W.; Mulitze, D. Nearest neighbor adjusted best linear unbiased prediction. Am. Stat. 1991, 45, 194–200. [Google Scholar]
- Katsileros, A.; Drosou, K.; Koukouvinos, C. Evaluation of nearest neighbor methods in wheat genotype experiments. Commun. Biometry Crop Sci. 2015, 10, 115–123. [Google Scholar]
- Bowley, S. A Hitchhiker’s Guide to Statistics in Plant Biology; Any Old Subject Books: Guelph, ON, Canada, 1999. [Google Scholar]
- Hurburgh, C.R. Measurement of fatty acids in whole soybeans with near infrared spectroscopy. Lipid Technol. 2007, 19, 88–90. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Walker, E.R.; Young, L.D. Soybean seed composition as affected by seeding rates and row spacing. Crop Sci. 2014, 54, 1782–1795. [Google Scholar] [CrossRef]
- Goldberger, A.S. Best linear unbiased prediction in the generalized linear regression model. J. Am. Stat. Assoc. 1962, 57, 369–375. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Najafabadi, M.Y.; Heidari, A.; Rajcan, I. AllInOne Pre-processing: A comprehensive preprocessing framework in plant field phenotyping. SoftwareX 2023, 23, 101464. [Google Scholar] [CrossRef]
- Kaur, P.; Bayer, P.E.; Milec, Z.; Vrána, J.; Yuan, Y.; Appels, R.; Edwards, D.; Batley, J.; Nichols, P.; Erskine, W. An advanced reference genome of Trifolium subterraneum L. reveals genes related to agronomic performance. Plant Biotechnol. J. 2017, 15, 1034–1046. [Google Scholar] [CrossRef] [Green Version]
- Torkamaneh, D.; Laroche, J.; Bastien, M.; Abed, A.; Belzile, F. Fast-GBS: A new pipeline for the efficient and highly accurate calling of SNPs from genotyping-by-sequencing data. BMC Bioinform. 2017, 18, 5. [Google Scholar] [CrossRef] [Green Version]
- Raj, A.; Stephens, M.; Pritchard, J.K. fastSTRUCTURE: Variational inference of population structure in large SNP data sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yeh, C.-T.E.; Ramamurthy, R.K.; Qi, X.; Fernando, R.L.; Dekkers, J.C.; Garrick, D.J.; Nettleton, D.; Schnable, P.S. Empirical comparisons of different statistical models to identify and validate kernel row number-associated variants from structured multi-parent mapping populations of maize. G3 Genes Genomes Genet. 2018, 8, 3567–3575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X. rmvp: A memory-efficient, visualization-enhanced, and parallel-accelerated tool for genome-wide association study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef]
- Weston, J.; Mukherjee, S.; Chapelle, O.; Pontil, M.; Poggio, T.; Vapnik, V. Feature selection for SVMs. In Advances in Neural Information Processing Systems; MIT Press: Cambridge, MA, USA, 2000; pp. 668–674. [Google Scholar]
- Siegmann, B.; Jarmer, T. Comparison of different regression models and validation techniques for the assessment of wheat leaf area index from hyperspectral data. Int. J. Remote Sens. 2015, 36, 4519–4534. [Google Scholar] [CrossRef]
- Doerge, R.W.; Churchill, G.A. Permutation tests for multiple loci affecting a quantitative character. Genetics 1996, 142, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Churchill, G.A.; Doerge, R.W. Empirical threshold values for quantitative trait mapping. Genetics 1994, 138, 963–971. [Google Scholar] [CrossRef]
- Kuhn, M.; Wing, J.; Weston, S.; Williams, A.; Keefer, C.; Engelhardt, A.; Cooper, T.; Mayer, Z.; Kenkel, B.; Team, R.C. Package ‘caret’. R J. 2020. Available online: https://github.com/topepo/caret/ (accessed on 10 July 2023).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GWAS Method | Chromosome | MTA (Peak SNP Position) | Co-Located QTL | Environments a | Reference |
|---|---|---|---|---|---|
| FarmCPU | S15 | 7068549 | Shoot Fe 1-g43 | NA | [39] |
| 7288161 | SCN 5-g32 | NA | [40] | ||
| 7705443 | Seed protein 7-g13 | NA | [41] | ||
| Leaf carotenoid content 1-g11 | NA | [42] | |||
| WUE 2-g34 | NA | [43] | |||
| 8304621 | Shoot Zn 1-g24 | NA | [39] | ||
| 8554284 | Shoot Zn 1-g25 | NA | [39] | ||
| 8620771 | Shoot Zn 1-g26 | NA | [39] | ||
| SVR | S01 | 50879523 | Ureide content 1-g1.1 | NA | [42] |
| 50933494 | Ureide content 1-g1.2 | NA | [42] | ||
| 50945345 | Ureide content 1-g1.3 | NA | [42] | ||
| 50947984 | Ureide content 1-g1.4 | NA | [42] | ||
| 51104169 | First flower 2-g1 | NA | [44] | ||
| 51797141 | Canopy cover 1-g1 | NA | [45] | ||
| 51104169 | First flower 7-g1 | NA | [44] | ||
| 51679239 | Seed Trp 1-g1 | NA | [46] | ||
| S05 | 37483313 | Shoot Mg 1-g4 | 2&4 | [39] | |
| 37414768 | Shoot Cu 1-g6 | 2&4 | [39] | ||
| 31380926 | Seed oil 5-g1 | 2&4 | [42] | ||
| 35536817 | Pod number 3-g4 | 2&4 | [47] | ||
| 31380926 | Seed protein 4-g1 | 2&4 | [48] | ||
| 37297357 | Shoot Zn 1-g10.1 | 2&4 | [39] | ||
| 37347763 | Shoot Zn 1-g11 | 2&4 | [39] | ||
| 37289637 | Shoot P 1-g7 | 2&4 | [39] | ||
| Shoot Zn 1-g9 | 2&4 | [39] | |||
| 37297357 | Shoot P 1-g8.1 | 2&4 | [39] | ||
| 37317508 | Shoot P 1-g8.2 | 2&4 | [39] | ||
| Shoot Zn 1-g10.2 | 2&4 | [39] | |||
| 37347763 | Shoot P 1-g9 | 2&4 | [39] | ||
| S14 | 2919862 | First flower 2-g20 | NA | [44] | |
| 3198128 | Sclero 3-g56 | NA | [49] | ||
| 3419976 | Sclero 3-g57 | NA | [49] | ||
| S16 | 28851611 | Seed protein 7-g25 | 1,2&4 | [41] |
| GWAS Method | Chromosome | Peak SNP Position | Co-Located QTL | Environments a | Reference |
|---|---|---|---|---|---|
| FarmCPU | S08 | 18259484 | SDS 1-g54 | NA | [50] |
| 18404800 | SDS 1-g40 | NA | [50] | ||
| SDS 1-g55 | NA | [50] | |||
| S13 | 27301888 | Shoot Fe 1-g33 | NA | [39] | |
| SCN 1-g11 | NA | [51] | |||
| 27325073 | Shoot Fe 1-g34 | NA | [39] | ||
| 33018554 | SCN 4-g11 | NA | [52] | ||
| S15 | 7705443 | Seed protein 7-g13 | NA | [41] | |
| Leaf carotenoid content 1-g11 | NA | [42] | |||
| WUE 2-g34 | NA | [43] | |||
| 8304621 | Shoot Zn 1-g24 | NA | [39] | ||
| 8554284 | Shoot Zn 1-g25 | NA | [39] | ||
| S19 | 40386502 | Iron deficiency chlorosis 4-g27 | NA | [53] | |
| 40550665 | Iron deficiency chlorosis 2-g9 | NA | [54] | ||
| Iron deficiency chlorosis 3-g14 | NA | [54] | |||
| SVR | S03 | 12702388 | Seed long-chain fatty acid 1-g7.2 | 2 | [55] |
| 12704607 | Seed stearic 1-g2.2 | 2 | [55] | ||
| 12917268 | Seed long-chain fatty acid 1-g13.2 | 2 | [55] | ||
| 12954110 | Seed stearic 1-g2.3 | 2 | [55] | ||
| 12958942 | Seed long-chain fatty acid 1-g13.3 | 2 | [55] | ||
| 12989558 | Seed long-chain fatty acid 1-g7.3 | 2 | [55] | ||
| S13 | 30062400 | Hilum color 2-g5.2 | NA | [55] | |
| Hilum color 2-g5.3 | NA | [55] | |||
| 30080662 | Phytoph 3-g21 | NA | [51] | ||
| 29941996 | Soybean mosaic virus 1-g1 | NA | [51] | ||
| 30037573 | Salt tolerance 1-g9 | NA | [56] | ||
| 30062400 | Hilum color 2-g5.1 | NA | [55] | ||
| S14 | 3198128 | Sclero 3-g56 | 3 | [49] | |
| 3419976 | Sclero 3-g57 | 3 | [49] | ||
| S15 | 21479453 | Iron deficiency chlorosis 4-g20 | 3 | [53] | |
| 49067066 | WUE 1-g5 | 3 | [57] | ||
| S16 | 28851611 | Seed protein 7-g25 | 3 | [41] |
| GWAS Method | Chromosome | Peak SNP Position | Co-Located QTL | Environments a | Reference |
|---|---|---|---|---|---|
| FarmCPU | S18 | 703188 | WUE 2-g47 | NA | [43] |
| 713403 | SCN 1-g16 | NA | [51] | ||
| 822049 | SCN 1-g17 | NA | [59] | ||
| SVR | S02 | 11045403 | Seed Trp 1-g5 | 2&4 | [46] |
| 43004026 | WUE 2-g7 | 2&4 | [48] | ||
| S03 | 38932768 | Canopy width 1-g1.1 | NA | [55] | |
| 38936586 | Canopy width 1-g1.2 | NA | [55] | ||
| 39088673 | R8 full maturity 3-g4 | NA | [47] | ||
| S09 | 42132672 | Al tolerance 1-g9 | NA | [60] | |
| 42351295 | Shoot K 1-g19 | NA | [39] | ||
| S11 | 4572326 | SCN 5-g22 | 4 | [40] | |
| S15 | 36329398 | Ureide content 1-g42 | NA | [42] | |
| S16 | 37330986 | Seed linolenic 1-g10 | 2 | [61] | |
| 37153578 | Shoot Cu 1-g15 | 2 | [39] | ||
| 37330986 | Seed palmitic 1-g14 | 2 | [61] | ||
| Seed oleic 1-g23 | 2 | [61] | |||
| Seed linoleic 1-g19 | 2 | [61] | |||
| 37046875 | WUE 2-g38 | 2 | [40] | ||
| Iron deficiency chlorosis 3-g10 | 2 | [54] | |||
| 37079553 | Node number 1-g5.1 | 2 | [55] | ||
| 37079569 | Node number 1-g5.2 | 2 | [55] | ||
| 33018083 | BSR 1-g2 | 2 | [51] | ||
| S19 | 47335622 | Node number 1-g2.3 | NA | [55] | |
| S20 | 276646 | First flower 2-g25 | 1&2 | [44] | |
| First flower 7-g25 | 1&2 | [44] | |||
| 343016 | Iron deficiency chlorosis 3-g15 | 1&2 | [54] | ||
| 376574 | Plant height 1-g26 | 1&2 | [44] | ||
| Plant height 6-g26 | 1&2 | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoosefzadeh-Najafabadi, M.; Torabi, S.; Tulpan, D.; Rajcan, I.; Eskandari, M. Application of SVR-Mediated GWAS for Identification of Durable Genetic Regions Associated with Soybean Seed Quality Traits. Plants 2023, 12, 2659. https://doi.org/10.3390/plants12142659
Yoosefzadeh-Najafabadi M, Torabi S, Tulpan D, Rajcan I, Eskandari M. Application of SVR-Mediated GWAS for Identification of Durable Genetic Regions Associated with Soybean Seed Quality Traits. Plants. 2023; 12(14):2659. https://doi.org/10.3390/plants12142659
Chicago/Turabian StyleYoosefzadeh-Najafabadi, Mohsen, Sepideh Torabi, Dan Tulpan, Istvan Rajcan, and Milad Eskandari. 2023. "Application of SVR-Mediated GWAS for Identification of Durable Genetic Regions Associated with Soybean Seed Quality Traits" Plants 12, no. 14: 2659. https://doi.org/10.3390/plants12142659