Abstract
Carrot (Daucus carota L.) is a highly consumed vegetable rich in carotenoids, known for their potent antioxidant, anti-inflammatory, and immune-protecting properties. While genetic and molecular studies have largely focused on wild and Western carrot cultivars (cvs), little is known about the evolutionary interactions between closely related Eastern and Western cvs. In this study, we conducted comparative transcriptome profiling of root tissues from Eastern (UHSBC-23-1) and Western (UHSBC-100) carrot cv. to better understand differentially expressed genes (DEGs) associated with storage root development and vascular cambium (VC) tissue patterning. Through reference-guided TopHat mapping, we achieved an average mapping rate of 73.87% and identified a total of 3544 DEGs (p < 0.05). Functional annotation and gene ontology classification revealed 97 functional categories, including 33 biological processes, 19 cellular components, 45 metabolic processes, and 26 KEGG pathways. Notably, Eastern cv. exhibited enrichment in cell wall, plant-pathogen interaction, and signal transduction terms, while Western cv. showed dominance in photosynthesis, metabolic process, and carbon metabolism terms. Moreover, constructed gene regulatory network (GRN) for both cvs. obtained orthologs with 1222 VC-responsive genes of Arabidopsis thaliana. In Western cv, GRN revealed VC-responsive gene clusters primarily associated with photosynthetic processes and carbon metabolism. In contrast, Eastern cv. exhibited a higher number of stress-responsive genes, and transcription factors (e.g., MYB15, WRKY46, AP2/ERF TF connected via signaling pathways with NAC036) were identified as master regulators of xylem vessel differentiation and secondary cell wall thickening. By elucidating the comparative transcriptome profiles of Eastern and Western cvs. for the first time, our study provides valuable insights into the differentially expressed genes involved in root development and VC tissue patterning. The identification of key regulatory genes and their roles in these processes represents a significant advancement in our understanding of the evolutionary relations and molecular mechanisms underlying secondary growth of carrot and regulation by vascular cambium.
1. Introduction
Carrot (Daucus carota L.) is widely cultivated spp. from Apiaceae family [1], with a diploid chromosome number of 2n = 2x = 18 and an estimated genome size of 473 Mb [2]. This crop holds a prominent position among primary vegetables, ranking in the top ten alongside tomato, onion, cabbage, cucumbers, and eggplant [3,4,5]. Moreover, carrot is recognized as the second-most important vegetable crop after potatoes, due to its nutritional composition and economic significance [6,7]. Over the past five decades, the global demand for carrots has steadily increased, with substantial growth in cultivated area (1.1 Mha) and production (40.0 Mt) [4]. Carrots are gaining popularity as a source of carbohydrates, minerals and vitamins, notably provitamin A [8]. In addition, carrots are a valuable source of carotenoids, important bioactive compounds that contribute to human health by offering a multitude of benefits, including antioxidant and anti-inflammatory properties, as well as immune and cardiovascular protection [9,10].
Considering morphological traits, cultivated carrots are classified into two types/cultivars: (1) Eastern, which include yellow, purple, or pale orange carrots, and (2) Western, primarily consisting of white, deeply pigmented orange or red carrots [8,11]. This classification is supported by genomics and transcriptomics studies as well [11,12,13,14,15,16,17]. Importantly, it was suggested that both carrot types have ancestral relationships with the domestication history [18]. For instance, Iorizzo et al. [2,17] suggest that Eastern carrot cvs. originated approximately 1100 years ago from wild populations in South Asia. The development of the modern dark orange Western carrot cvs. involved secondary domestication from the natural hybridization between yellow Eastern carrots and white-rooted accessions of wild carrots (Daucus carota L. ssp. Carota) [6,19]. Selective breeding of Western carrots has led to the enhancement of desirable characteristics, including a higher content of provitamin A and carbohydrates in uniform, juicy and smooth storage roots. However, Western carrot varieties exhibit a biennial flowering habit and require vernalization for optimal growth and development [11,18,19]. In contrast, Eastern carrot cvs. exhibit uneven shapes, larger core sizes, lower provitamin A content, coarser texture, and annual flowering, with minimal or no vernalization requirement for flowering [8].
Even though many genetic studies support a Central Asian origin of carrot domestication, specific molecular mechanisms driving domestication and underlying genotypic variability responsible for phenotypic changes, still remain poorly understood. While breeders have capitalized on the extensive phenotypic diversity present in carrots [20], a comprehensive investigation into the genotypic basis of these traits is still warranted [2]. Previous findings by Machaj et al. [21] provided insight into storage root development and genetic complexity between wild accession of D. carota subsp. Commutatus and domesticated orange-rooted Western carrots (Daucus carota ssp. Sativus var. sativus). However, there are still few comparative studies examining both the morphological and molecular aspects of these two carrot types. Namely, in carrots, the highest diversity has been found related to root thickening and vascular tissue differentiation, leading to genetic variation in root shape, color, length, and yield. In rooty crops, the development of storage roots and their yields primarily depends on the vascular cambium (VC) tissue. The VC acts as the key regulator, determining the thickening of xylem (internal) and phloem (external) tissues during the formation of secondary storage roots [22,23]. Continuous cell division (i.e., anticlinal type of cell division in VC) provides precursor cells for the secondary growth of the xylem inside the VC and phloem extension outside the VC by their radial growth [24]. In this process, several stress-responsive transcription factors (TF), VC and carbon partitioning genes play a crucial role. In coordination with environmental changes, a dynamic and complex gene regulatory network (GRN) operates in the VC for xylem and phloem formation [24]. In addition, cultivated plant species are exposed to a variety of abiotic [25] and biotic stresses [26], with negative impacts on their growth performances [27] and evolution [28]. However, it is believed that plant growth has an antagonistic relationship with stress resistance. Moreover, plants can prioritize their defense responses for survival by inducing stress-responsive gene cascades and suppressing growth-promoting regulators at the molecular level [29,30]. The molecular mechanisms underlying stress responses and growth modulation have widely been investigated in Arabidopsis thaliana, a model plant in stress biology [31], shedding light on the intricate balance between growth and stress adaptation [32].
In this study, we focused on analyzing the transcriptome of the storage root system in two important carrot cultivars: Eastern (Daucus carota ssp. sativus var. atrorubens Alef.) and Western (Daucus carota ssp. sativus var. sativus), with distinct domestication histories and valuable genetic resources for further investigation. In addition, GRN constructed with the VC regulatory genes separately for Eastern and Western carrot cvs. was compared with Arabidopsis thaliana from recent studies [24,33]. As a result, identified novel regulators of VC activity in response to growth and stress could (i) accelerate the genetic improvement of the carrot taproot, as well as (ii) elucidate molecular regulatory mechanisms governing modulation of key genes for stress resistance and growth.
2. Results and Discussion
2.1. Carrot Root Yield Depends on the Secondary Growth
A wide range of root plasticity parameters were recorded in the test carrot cultivars and are presented in Table 1. Eastern cv. has contrasting domestication traits with the Western cv. (Table 1) in terms of root shape, xylem-phloem patterning, growth, and flowering habit, which is in line with some previous studies [8,11]. Significant variations between test cvs. were found in morphometric parameters such as root yield, shoulder width, xylem and phloem color and size, carotenoid accumulation, growth habit, and blooming behavior (Table 1). Western cv. obtained the highest phloem width (86 mm) with uniform pigmentation and little xylem patterning (40 mm), whereas Eastern cv. showed the highest xylem width (84 mm), but lower phloem width (50 mm) (Table 1). Uniform exterior and interior colors are important characteristics receiving attention in carrot breeding programs. The average shoot weight (70.91 ± 12 g) of Eastern cv. was higher in comparison to Western cv. (24.44 ± 3.46 g). However, average root weight of Western cv. (40.66 ± 4.11 g) was lower than in Eastern cv. (72.70 ± 5.88 g). Western cv. exhibited significantly higher β-carotene content (8.5 mg/100 mg) than Eastern cv. (6.5 mg/100 mg; Table 1). The total soluble solids (TSS) varied from 6.86° Brix in Western cv. to 7.08° Brix in Eastern cv. These morphometric variations are useful tools in the (pre)selection of desirable cvs. [34].

Table 1.
Morphological traits distinguishing Eastern and Western carrot cultivars.
In addition, the average root width showed a strong positive correlation with average root weight (R2 = 0.675) (Table 2). The vegetative weight has a positive correlation with xylem width (R2 = 0.440); however, it has a negative correlation with phloem width (R2 = −0.166). The root width was influenced positively by xylem (R2 = 0.432) and phloem width (R2 = 0.44). As shown by a strong correlation, the xylem width will increase the root weight (R2 = 0.726) markedly more than the phloem width (R2 = 0.342) since the xylem core is harder and heavier. Both the xylem and phloem obtained a positive correlation with root weight and root width, indicating that radial root growth is an important factor determining root yield in carrots. An increase in shoulder width is correlated with higher xylem width (R2 = 0.869) than phloem width (R2 = 0.450). Hence, Eastern cv. Has a higher root weight than Western cv. (Table 1); however, the energy is mostly diverted to the shoulder portion and to the xylem core formation, making it coarser in texture. The poor negative correlation between shoot length and root width (R2 = −0.091) and phloem width (R2 = −0.115) was recorded. In addition, phloem width had a negative association with plant vegetative weight (R2 = −0.166), number of petioles (R2 = −0.178), shoot length (R2 = −0.115), and root length (R2 = −0.059), whereas xylem width had significant positive correlations with plant height (R2 = 0.692), petiole length (R2 = 0.432), root width (R2 = 0.726), and vegetative weight (R2 = 0.440) (Table 2). This result highlights the importance of achieving a uniform phloem pattern in carrots, which is associated with lower vegetative weight, shorter petiole length, and reduced shoulder width. Western cvs. Evolved with these desirable characteristics through careful selection during domestication [19], whereas Eastern cv. Had limited exposure to the domestication of such traits. Therefore, to gain further insights into the genetic basis of secondary root growth and VC patterning in Eastern and Western carrot cvs., transcriptome profiling was employed to explore genes and regulatory pathways involved.
Table 2.
Correlation pattern among the morphological characters in tested carrot cultivars.
2.2. Reference-Based Assembly and DEGs in Storage Root Transcriptome
RNAseq technology was employed in the current study to retain 130.62 M (92.71%) high-quality (Phred score > 30), cleaned paired-end reads out of a total of 140.88 M reads. The cleaned reads from each library, with the range of 17.63 M to 24.93 M reads per sample, were mapped using reference-based assembly (Table 3), resulting in more reads than previous studies on the transcriptomics of carrots [21,35] or other storage rooty crops such as radishes [36]. The TopHat assembly mapping rate varied from 71.6 to 82.8%, with an average mapping percentage of 73.87%. Mapping rates in Western cv. ranged from 71.6 to 82.8% and in Eastern cv. from 68.5 to 72.7% (Table 3), which is markedly higher than in some similar studies, e.g., from 0.06 to 66.61%, [37].
Table 3.
Summary of the cleaned reads of root transcriptome aligned to the reference genome by TopHat v2.0.6.
A total of 3544 DEGs were identified by comparing expression levels of genes or other genomic features between test carrot cvs. using storage root transcriptome analysis. In Eastern cv., 1817 genes were up-regulated, while 1727 genes were found to be up-regulated in Western cv. (Figure 1a,b). Conversely, genes that are up-regulated in Eastern cv. are down-regulated in Western cv., and vice versa. Gene expression coverage across the chromosomal regions is a useful feature for analyzing DEGs spread across the genome (Figure 1c). Difference in DEGs between tested cultivars reflects the possible molecular mechanisms that control storage root development and VC patterning or their significance in domestication and evolution [6,19].
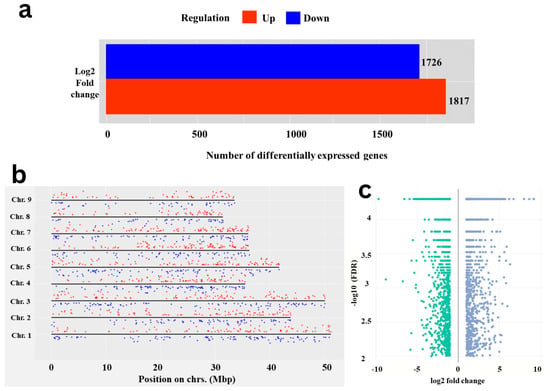
Figure 1.
Differentially expressed genes in Eastern and Western cultivars root transcriptome. (a) Bar plot representing the total number of up- and down-regulated DEGs in the root transcriptome by RNAseq analysis; (b) Chromosome-wise distribution of 3544 DEGs on 9 haploid sets of chromosomes of Carrot reference genome; up- (red dots) and down-regulated (blue dots) DEGs across Eastern and Western cv. are highlighted with different colors. (c) Volcano plot showing the up- (blue dots) and down-regulated (green dots) DEGs in Eastern and Western cv. of carrot, respectively.
2.3. Functional Annotation of DEGs
In the DAVID v6.8 web server, using Daucus carota subsp. sativus L. as a reference, a total of 2776 (78.33%) of the 3544 DEGs in the root transcriptome were functionally annotated. Among these, 70.83% (1287) and 86.21% (1489) of the 1817 up-regulated genes in Eastern cv. (Table S1) and 1727 up-regulated genes in Western cv. (Table S2), respectively, were successfully annotated. The remaining 768 genes were either uncharacterized or had no function assigned. The up-regulated DEGs in Eastern cv. were associated with “innate and adaptive immune response” inclusive of WRKY transcription factors, ABC transporters, and Mitogen-activated protein Kinase (MAPK) signaling. Furthermore, cellular process (cell cycle checkpoint protein, cell wall protein, cellulose synthase-like protein D3, cell division control protein, xyloglucan endotransglucosylase), cell wall synthesis (cell wall protein RBR3, repetitive proline-rich cell wall, pectin esterase), pathogenesis-related genes, phenylpropanoid, Ca signaling, and various chaperon proteins were also up-regulated (Table S2). Likewise, cytochrome P450-like, ubiquitin-protein ligase, gibberellin-regulated protein, kinesin-like protein (KIN-like), leaf rust disease resistance, leucine-rich receptor protein-related DEGs were higher in the Eastern cultivar. Heavy metal-associated isoprenylated plant proteins (HIPPs) that are involved in heavy metal homeostasis and detoxification in cells, serine-threonine protein kinase, NAC, WRKY TF, and U-box domain-containing protein, had higher accumulation in Eastern cv. In addition, various plant hormones responsive transcripts (auxin-responsive protein-SAUR32, IAA26, auxin response factors 3, 4, auxin-induced proteins, cytokinin dehydrogenase 7-like, gibberellin-regulated protein, ethylene-responsive transcription factors-ERF-061, 014, 25, CRF4-like, abscisic acid-insensitive, etc.), NRT1/PTR gene family in response to salt stress, etc., were highest in Eastern cv. that is necessary for wider adaptation to climate change. A total of 58 TF of different classes that play key roles in gene expression were found in Eastern cv. (Table S1). A significantly higher number of protein kinases (104) was detected in Eastern vs. Western cv. (36) (Tables S1 and S2). Throughout the plant cycle, root growth during biotic stress responses and MAPK signals are extremely important [36,38,39,40]. Phenylpropanoid biosynthesis is also important in releasing the phenols against the water stress and wound stress in carrots, which helps in pre-/post-harvest management [41].
Possible selection signatures are identified by the higher accumulation of photosynthesis-related genes (photosystem-I reaction center and photosystem II protein, photosynthetic NDH subunit), carotenoid biosynthetic pathway genes (phytoene synthase, phytoene desaturase), and chlorophyll a-b binding proteins were more highly accumulated in Western cv. than in Eastern cv. (Table S1). A higher accumulation of carotenoids is essential for photosynthesis and has previously shown prominent diversity among the yellow, red, and orange carrot roots [5]. Our results with Western cv. are also supported by a carrot genome study [2] showing the upregulation of photosynthetic pathway genes and Photosystem-I and II in orange carrots. Photosynthates reach roots by sieve components, companion cells, and plasmodesmata to facilitate secondary root development [42,43]. The uniform dark orange storage root in Western cv. is associated with photosynthetic efficiency [2,5] and could be a rate-limiting pathway in breeding for higher carotenoid accumulation in Eastern cvs. In addition, heat shock proteins, auxin-responsive proteins, thylakoid luminal protein, F-box protein, F-box-Kelch repeat protein were found to be higher in Western cv. Similarly, Dof zinc finger protein, MYB TF, probable WRKY TF, oxygen-evolving enhancer protein, Zn finger proteins and pentatricopeptide repeat-containing protein that facilitates processing, splicing, editing, stability and translation of RNAs [44] were also found to be higher in Western cv. (Table S2).
Important transcription factors such as ERF, heat stress TF, NAC-29 TF, WRKY, bHLH, CYCLOIDEA TF, DIVARICATA, MYB108, MYC2, RAX2, TCP4, trihelix TF GT-3b-like, bZIP, AP2/ERF, ethylene-responsive TF-CRF4 were up-regulated in Eastern cv. (Table S2). These TF play an important role in the regulation of storage root development, defense mechanisms, signal transduction, and abiotic stress tolerance [21]. Parallel, in Western cv., TF such as bHLH, DIVARICATA, MYB, E2F, HBP-1b, PCL1, PIF7, SPATULA, TCP, TGA, VIP-1 like, RAV-1 like, B3 domain containing VRN1 like, ERF, RAP2, TINY-like, NF-YA, GATA, and WRKY were up-regulated (Table S2). PIF TF regulates photomorphogenesis in carrot, plays an important role in hypocotyl elongation in the dark, and enhances carotenoid accumulation [45]. VRN1 plays a crucial role during vernalization to repress the FLC gene for a successful vegetative to reproductive transition [46]. The divergence of the type and class of transcriptional factors between test cultivars would be the result of selection signatures of two domestication events in carrots and play a possible role in the differential expression of domesticated traits, especially regarding root quality in terms of carotenoid accumulation and defense response to various stresses.
Protein kinases are a class of proteins important in post-translation modification by phosphorylation, protein conformation, stability, localization, signal transduction, and cell regulation, and responses to various environmental stresses [47]. Interestingly, various classes of protein kinases were found to be higher in Eastern cv. (104; Table S1) than in Western cv. (only 36; Table S2), which can be interrelated to a higher accumulation of signaling pathways in Eastern cv. Furthermore, Machaj et al. [21] observed a significant increase in protein kinase accumulation during the storage root development of the wild carrot relative to Western cv.
2.4. Gene Ontology (GO) Terms and KEGG Pathway Annotation of Differentially Expressed Genes
To further establish the biological function of DEGs responsible for storage root development and VC patterning in carrots, the functional annotation was performed by mapping all DEGs to Gene Ontology (GO). A total of 3544 DEGs in the root transcriptome were classified into 97 functional groups consisting of 33 biological processes (BP), 19 cellular processes (CC), and 45 molecular functions (MF) subcategories (Figure 2a–c & Table S3). The BP terms in Eastern cv includes phosphate ion transport, metabolic processes (sucrose metabolism, fructose metabolism, xyloglucan), cell wall organization, cell wall biogenesis, microtubule-based movement, and other processes. The structure of the cell wall plays critical functions in plant development, cell differentiation, cell expansion, intercellular communication, and defense [48]. CC terms were related to integral components of membrane, cytosol, plasma membrane, plasmodesmata, and cell wall specifying nuclear and inter-cellular transport and localization. MF terms were shared with ATP binding, oxidoreductase activity, protein serine/threonine kinase activity, xyloglucan transferase sequence-specific DNA binding, and others. This GO analysis was implemented with Bonferroni corrected FDR ≤ 0.01 and fold enrichment 1.0-11.51 (Table S3). In the KEGG, important signaling pathways (MAPK signaling, phenyl propanoid pathway), metabolic processes, ABC transporters, and plant-pathogen interaction enrichment in Eastern cv. support the strong root development that has wider buffering capacity and climatic adaptation to various (a)biotic stresses (Figure 2d). Plants have signal transduction pathways, which are complex networks of interactions involving signal elements transmitting through the plant cell and allowing them to respond appropriately to a specific environmental stimulus. Cell signaling influences nearly every aspect of plant cell structure and function [49]. The absence of these GO terms during the subsequent secondary domestication event in Western carrots would be hypothesized as a domestication bottleneck.
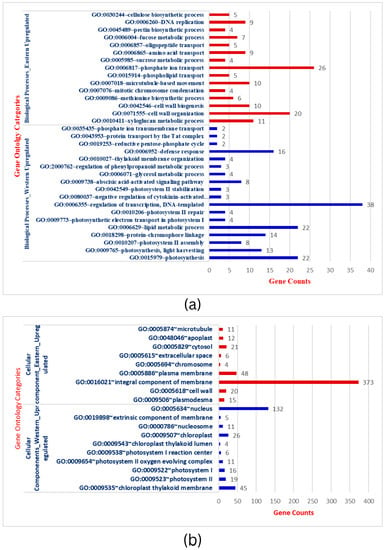
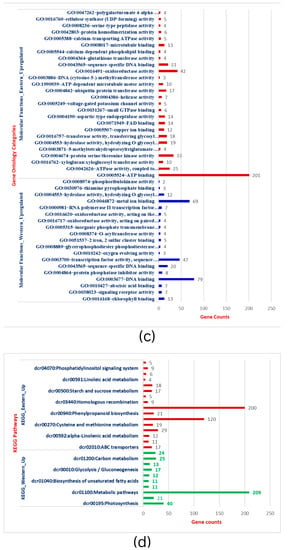
Figure 2.
(a): Gene ontology categorization of Biological processes (BP) across Eastern and Western carrot cultivars and the number of genes showing respective BP. (b): Gene ontology categorization of Cellular Components (CC) across Eastern and Western carrot cultivars and the respective genes involved in CC. (c): Gene ontology categorization of Molecular Functions (MF) across Eastern and Western carrots and the respective number of genes involved in MF. (d): Comparison of DEGs involved in KEGG. Red bars indicate KEGG pathways upregulated in Eastern cv, while blue/green bars indicate KEGG pathways upregulated in Western cv.
Conversely, up-regulated BP in Western cv. was related to the regulation of transcription, photosynthesis (photosystem-I, photosystem-II, oxygen-evolving complex), metabolic process (lipid metabolism, carbon metabolism, biosynthesis of secondary metabolites), and defense response (Figure 2a–c & Table S4). Genes in photosynthesis were variably expressed and substantially enriched during storage root formation in cultivated orange carrots and repressed in wild carrots in previous comparative transcriptomics research [21]. The carotenoid pathway is enriched in early or developing roots, whereas in adult roots, carotenoid biosynthesis genes are less common and co-expressed with chloroplast genes [21]. Increased carotenoid accumulation in mature carrot roots is correlated with greater expression of genes involved in photosynthesis, such as LHC-II [2]. CC terms in Western cv. were confined to the nucleus, chloroplast thylakoid membrane, photosystem II, photosystem I, oxygen-evolving complex, photosystem I reaction center, chloroplast thylakoid lumen, and nucleus. MF expressions in Western cultivars were DNA binding, metal binding, transcription factor activity, and chlorophyll binding (Table S4). In carrots, genes involved in plastid formation, photosystem I and II assembly, and carotenoid biosynthesis are co-expressed with isoprenoid pathways [2,21,50]. In summary, extensive analysis has demonstrated that the gene expression patterns and enriched pathways in Eastern and Western cvs. are unequivocally distinct from each other.
To further identify the differentially expressed transcripts that are involved in the same or different biological pathways and to gain deeper insight into the enriched pathways between test carrot cultivars, KEGG pathway analysis was carried out to emphasize the network of gene products (Figure 3a,b). It was found that metabolic pathways (200 genes) and biosynthesis of secondary metabolites (120), plant-pathogen interaction (29), phenylpropanoid biosynthesis (21), MAPK signaling, and ABC transporters were up-regulated pathways in Eastern cv. (Figure 2d and Table S3). For Western cv., metabolic pathways (209 genes) were followed by photosynthesis (40), carbon metabolism (25), plant hormone signal transduction (24), along with six other KEGG pathways (Figure 2d and Table S4). This shows the systematic evolution of the photosynthetic pathway for higher carotenoid accumulation in Western cv. as supported by previous findings [2,5] and our KEGG pathway enrichment analysis (Figure 3b). Further KEGG pathway enrichment analysis confirmed the enrichment of pathways viz., “alpha-linoleic acid metabolism”, “ABC transporters”, “plant-pathogen interaction”, “phenylpropanoid pathways”, ‘metabolic processes’ in Eastern cv. (Figure 3a), and “metabolic processes” followed by “photosynthetic pathways” and “carbon metabolism” in Western cv. (Figure 3b). These results suggest that differentially expressed transcripts likely have a significant impact on the development of secondary storage roots and the transportation of photosynthates from leaves to roots but in distinct patterns between tested cvs.
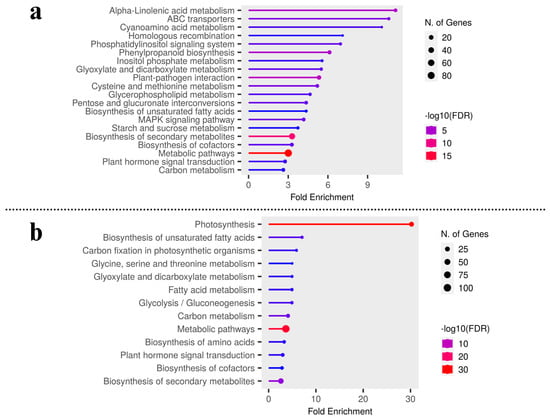
Figure 3.
KEGG pathways enrichment analysis by gene set enrichment analysis in (a) Eastern cv. and (b) Western cv.
2.4.1. DEGs of Cell Cycle and Cell Wall Metabolism Transcript Profiles
Genes associated with the cell cycle were observed as differentially regulated in carrot storage root (Tables S5 and S6). For instance, cell division control protein 2 (CDC2), cyclin-dependent kinase (CDKs), cyclin-D1-1-like, B2-1-like, and cyclin-dependent kinase inhibitor (CDKI) 4-like SMR were up-regulated in Eastern cv. In addition, glycine-rich cell wall structural protein, putative cell division cycle ATPase, repetitive proline-rich cell wall protein, programmed cell death protein, and accelerated cell death related DEGs were up-regulated in Eastern carrot (Table S5). The cell wall not only strengthens the plant body, but also has key roles in plant growth, cell differentiation, cell expansion, intercellular communication, water transport, and defense [51,52].
In the present study, transcripts encoding key enzymes that are involved in the cell wall synthesis and degradation, such as cellulose synthase, galactosyltransferase, UDP-glycosyltransferase, and caffeoyl-CoA O-methyltransferase, endoglucanase, pectin acetyl esterase, pectin esterase, Xyloglucan endotransglucosylase protein-like-related DEGs were up-regulated during Eastern secondary storage root development compared to Western carrot (Tables S5 and S6). The cell cycle and division of cells determine the size of cells and organs [53]. Interestingly, higher numbers of transcripts homologous to cell division and cell wall metabolism were associated with Eastern carrot cv. (Figure 4a).
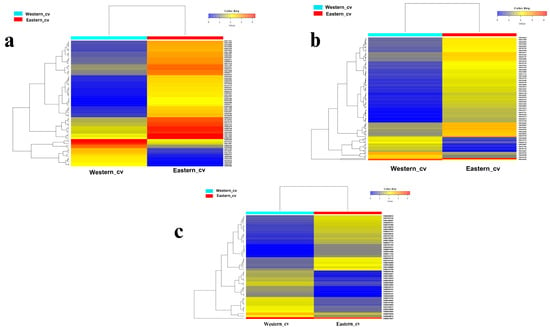
Figure 4.
Heat map constructed based on Euclidean distance depicting the significant differentially expressed transcripts across Eastern and Western cvs. involved in (a) cell cycle and cell wall metabolism, (b) signal transduction, (c) starch and sucrose metabolism. Heatmaps are drawn based on FPKM values of significant DEGs across cvs.
2.4.2. DEGs with Signal Transduction and Related Transcriptome Profiles
The mitogen-activated protein kinase (MAPK) was reported to play a key role in regulating the cell cycle and developmental processes [35]. Two MAPK (MAPK-9 and NPK-1-like) were found in Western cv. (Table S6), and four (MAPK-3, 9, MMK2 and NTF6) were found in Eastern cv. (Table S7). In addition, KEGG has also been enriched with MAPK signaling pathway in Eastern cv. (Table S3). Ca-regulated transduction pathway is another key signaling transduction category in cell development [35]. In Western cv., only two Ca-binding proteins (CML49 and CML36) were identified (Table S8). However, in Eastern cv, eight Ca-binding proteins and calmodulin (CaM) proteins (CML30, CML18, CML49, CML20, CML25, CML45, CML23, CML24) were found (Table S7). This suggests that pathways involved in Ca transport are more extensive in tested Eastern cv. than in Western cv. (Tables S7 and S8). Furthermore, the up-regulation of various genes in Eastern cv. (Table S7) indicates their potential role in important physiological processes. These include ABC transporters, WRKY transcription factors, plant pathogen interacting respiratory burst oxidase protein (RbohD) responsible for signaling reactive oxygen species (ROS), and caffeoyl-CoA O-methyltransferase-like enzymes involved in the phenylpropanoid pathway (Figure 4b, Tables S7 and S8).
2.4.3. DEGs of Starch and Sucrose Metabolism Transcript Profiles
Carbon-related metabolism genes (e.g., beta-amylase, sucrose synthase, sucrose phosphate synthase (SPS), UDP-glucose transferase, glucose phosphate, glucose dehydrogenase, beta-glucosidase, beta-galactosidase, beta-fructofuranosidase, and beta-xylosidase) as suggested by Treves et al. [54] exhibited down-regulation in Western cv. in contrast to Eastern cv. (Figure 4c, Tables S9 and S10), suggesting distinct differences in carbon metabolism and accumulation between test cultivars. However, in Western cv., KEGG analysis revealed a higher enrichment in metabolic pathways (vs. signal transduction pathways) such as photosynthesis, carbon fixation, glycolysis, gluconeogenesis, and carbon metabolism (Figure 3b). In addition, in Eastern cv., synthesis of higher levels of carbohydrates may also be involved in various signal transduction pathways and other biological processes, which is evident from KEGG analysis (Figure 3a). Higher accumulation of sucrose is necessary during storage root development [55]. These results suggest that many key functional genes are differing in the storage root between test carrot cultivars. Therefore, the GRN was constructed to find the predominant active pathway in the storage root of carrot cultivars and master regulators involved in the swapping of root growth and stress response (Figure 4c).
2.5. GRN Associated with VC Patterning between Test Carrot Cultivars
The GRN analysis revealed a higher number of stress-responsive genes and transcription factors (LOX2, LOX3, LOX4, WRKY46, MYB15, ERF4, PUB24, KIN3, NAC036, NLA etc.) in Eastern (vs Western) cultivars, along with a few genes related to root growth (PERK10, AT2G26380 Leucine-rich repeat (LRR, LSH6, HCA2) and carbohydrate metabolism (NANA)), with no significant impact on photosynthesis network genes in Eastern cv. (Figure 5a, Tables S11 and S12). In addition, GO enrichment analysis highlighted 83 biological process (BP) terms, 12 molecular function (MF) terms, 7 cellular component (CC) terms, and 10 KEGG pathway enrichments (Table S13) associated with VC-responsive genes specific to Eastern cv. Several of the target genes in GRN were implicated in cell cycle events, plant hormone regulation, and various signal transduction and metabolic processes, including cell division, differentiation, expansion, auxin signaling, sucrose metabolism, and energy metabolism (Figure 5a). These findings suggest their crucial role in taproot thickening during carrot development.
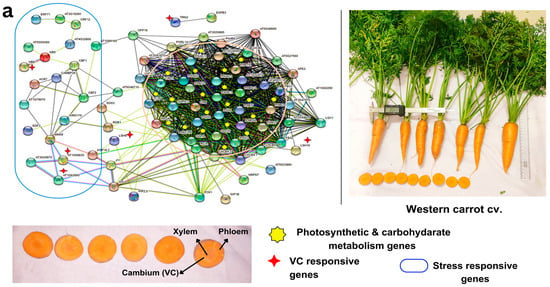
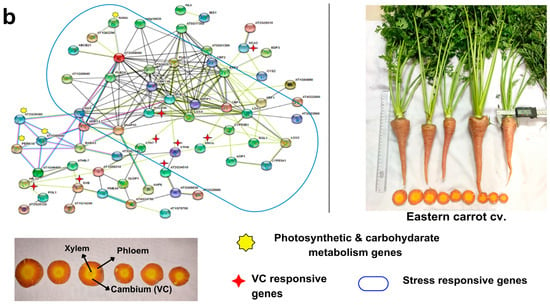
Figure 5.
Gene regulatory network showing the interconnections of vascular cambium (VC) responsive genes and DEGs with stress-responsive genes and photosynthesis and carbon metabolism pathways in (a) Eastern carrot and (b) Western carrot based on an orthology search with A. thaliana and network drawn in STRING v. 11. Western and Eastern cv. root morphology and the vascular tissue (xylem, phloem and cambium and color) that are distinct across the cultivars are highlighted in the carrot vascular tissue.
VC-responsive genes crucially play a role in maintaining the uniform xylem phloem pattering with the help of various regulatory mechanisms. In Eastern cv. XTH 7, XTH9, SVB, PhyA, HCA2, HB-12, and NAC036 are abundantly expressed in cambial regions during the secondary root growth (Figure 5a). These not only regulate xylem cell expansion but also influence several characteristics of secondary growth, including secondary xylem production and secondary wall deposition [56]. HCA2 is a Dof-type zinc finger DNA-binding family protein, which induces the formation of interfascicular cambium and regulates VC tissue development [24]. NAC036 present in Eastern cv. was identified as the master regulator of xylem vessel differentiation and secondary cell wall thickening [57]. WOX4 (AT1G46480) present in Eastern cv. promotes differentiation and/or maintenance of the vascular procambium. Down-regulation of WOX4 in Arabidopsis generated small plants with undifferentiated ground tissue and exhibited severe reductions in differentiated xylem and phloem [58]. The genome editing approach for this gene would facilitate uniform patterning in Eastern carrot cvs. Integration of DEGs and gene interactions in GRN provided insights into the intricate regulatory network associated with carrot taproot development, thickening, and VC patterning.
In Western cv., there was an up-regulation of photosynthetic cascades and photosynthesis-related genes, as observed in GO subcategories of BP and CC (Figure 5b, Tables S14–S16). Similarly, the GRN using up-regulated VC-responsive genes in Western cv. revealed the clustering of genes involved in photosynthetic activities (ex: LSH10, PSAH, PSAN, PSAD, LHCB, PSB, etc.), along with a few stress-responsive genes (WRKY70, ERF-71, MKK9, MYB15, CAMBP25) and root vascular developmental genes (TRN2, LSH10, SP1L2, HB6, AT1G66830) (Figure 5b, Tables S14 and S15). GO enrichment analysis of 109 up-regulated DEGs in Western cv. identified 82 enriched terms in BP, 18 in MF, 33 in CC, and 6 enriched KEGG pathways (Table S9) related to the VC-responsive genes (Table S16). Such results show that VC-responsive genes were co-expressed with photosynthetic processes leading to higher and more uniform carotenoid accumulation. It is well known that Western carrot cvs. have reduced genetic diversity [11,17] with increased desirable traits during domestication [8] and are highly susceptible to various pathogens [8,59]. This is also supported by our results because the root development and vascular tissue development utilizes higher photosynthates instead of diversion to other stress-responsive pathways, as confirmed in Eastern cv. TRN2 regulation is required for initial meristematic divisions in the epidermal layer and to maintain the radial phloem pattern of cell specification in the root [60]. LSH10 is a developmental regulator involved in deeper carbohydrate assimilation in storage roots [61]. AT1G66830 is an LRR-protein kinase found as a master regulator that cooperates with phloem intercalated XYLEM (PXY)/TDIF receptor, promoting cell divisions in the cambium, inhibiting xylem differentiation, and controlling VC patterning [62]. Several edible secondary storage rooty crops, such as potato, radish, yam, cassava, and sweet potato, have comparable results [24]. The inclusion of these two cultivars in a genomic-assisted quality improvement program could help in (i) the development of climate-resilient Western carrot cvs, and (ii) higher carotenoid accumulation with uniform ‘self-core’ and high-quality roots in Eastern carrot types.
Root development and response to the environment are controlled by the GRN [63]. To further understand the enhanced regulatory network pathways between test cvs, GRN was built utilizing DEGs of our root transcriptome orthologous to VC of Arabidopsis. The availability of a higher number of homologous genes with the major evolutionary closeness between carrot and Arabidopsis unfolds the major regulatory genes in this comparative investigation [64,65,66,67]. Signal transduction pathways, such as those involved in hormone, calcium, and MAPK signaling, as well as metabolic processes related to cell wall, carbohydrates, storage, and energy metabolism, play crucial roles in energy and nutrient absorption by cells. Regulatory elements influence the differentiation, division, and expansion of secondary xylem and phloem in root development [24]. To devise strategies for sustainable yield in rooty crops, it is essential to understand the cambium regulatory programs that drive storage root development.
2.6. Expression Validation of DEGs by qPCR
A total of 9 DEGs were randomly chosen and validated by RT-qPCR analysis to assess the validity and reproducibility of our DEG findings (Table S17). For each sample, the Log2 ratio of the FPKM value was utilized to reflect the fold changes in comparison to the Log2FC estimated based on the CT value of qPCR (Figure 6). For eight of the nine genes (G1 to G9), the fold change values from RNAseq and qPCR showed a concordant tendency of gene expression. Eight genes matched the direction of the change fold. The expression of G1 was discordantly expressed in opposing directions by RNAseq and qPCR. Current annotation indicates that the root growth is regulated by the genes: G1 (Mechano-sensitive ion channel protein 6-like), G2 (ubiquitin-conjugating enzyme E2 20-like; LOC108208449), G3 (polyubiquitin; LOC108227972), and G4 (probable WRKY transcription factor53; LOC108209169). Then, G6 (cysteine proteinase inhibitor B; LOC108211283) is engaged in seed germination, while G5 (probable glycosyltransferase At5g03795; LOC108193533) is likely involved in carbon metabolism. The innate immune response involves G7 (ATPase WRNIP1; LOC108207021), G8 (DNA repair protein XRCC2 homolog; LOC108216678), and G9 (histone deacetylase 6; LOC108200546) (Plant pathogen interaction). G1 (mechanosensitive ion channel protein 6-like; LOC108226635) participates in signaling pathways and homeostasis, but their interactions with other genes are involved in growth and development and are sensitive to hormones, light, and stress. With a few exceptions, it was discovered that relative expression changes based on qRT-PCR results were consistent with the RNAseq data (Figure 6). Results obtained through RNA sequencing (RNA-seq) offer a higher degree of precision because this method provides comprehensive gene expression data at the genome-wide level, allowing for the quantification of gene expression levels across the entire genome. RNA-seq is considered reliable due to its reliance on a substantial number of biological replicates, usually three or more, which enhances its credibility. In contrast, qRT-PCR techniques are typically employed to measure the expression levels of a limited number of genes. While there are situations where this approach may offer advantages, it is worth noting that RNA-seq methodologies and data analysis procedures have reached a level of reliability that often renders the validation of results by qPCR or other methods unnecessary [68,69,70].
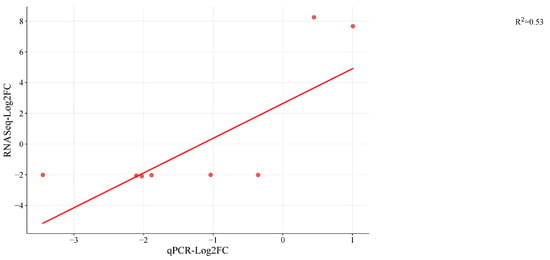
Figure 6.
Validation of eastern vs. western DEGs identified in RNA seq by quantitative real-time PCR (qRT-PCR) in storage root transcriptome of cultivars of carrot by Log2FC. Red dots indicate the candidate genes used for RNAseq (Table S1) the linear line is the regression line explaining qPCR and RNAseq results and R2 is the regression coefficient.
Our findings contribute to the improvement of Eastern cv. by identifying genes responsible for key domesticated properties such as enhanced carotenoids with consistent xylem-phloem color [11] and Western cv. improvement against different diseases and pests [8,55]. To our knowledge, this is the first report to identify likely candidate genes responsible for VC patterning in Eastern and Western carrot cvs. in secondary storage roots. The markers developed for the candidate genes of the present study are valuable for genetic diversity, genomic-assisted breeding, or genomic selection in carrot and related crop species [71,72].
3. Materials and Methods
3.1. Plant Material Preparation for Transcriptome Profiling
The vegetative phase of the study was conducted in Bagalkot, Karnataka, India (16°12′ N, 75°45′ E), during 2020 rabi season in a greenhouse. The average elevation in this area reaches approximately 610 m, with an average annual rainfall of 318 mm and a semi-arid tropical climate. For the storage root phenotypic evaluation and RNA sequencing, two carrot cultivars were selected: Eastern (Daucus carota ssp. Sativus var. atrorubens Alef) and Western (Daucus carota ssp. Sativus var. sativus) subgroup of the association panel [11]. Eastern cultivar UHSBC-23-1 is characterized by light orange pigmentation, clear vascular tissue differentiation, and non-uniformity in the xylem and phloem patterning. Western cultivar UHSBC-100 was a commercial dark orange ‘Kuruda’ type with a completely uniform xylem and phloem with a thin cambium line. Test cultivars were sown in sandy loam soil, with a pH between 6.0 to 7.0. At 90 days after sowing (DAS), fully matured and well-developed roots from both cultivars were harvested. For measuring various quantitative and qualitative traits related to root plasticity and other root morphometric characters, 100 plants were randomly selected from each cv. from the bulk. Quantitative traits such as plant height (cm), root length (cm), shoot length (cm), xylem width (mm), and phloem width (mm) were recorded with a measuring scale. Root width (cm) and shoulder width (cm) were measured using a digital vernier caliper. Root weight (g), shoot weight (g), and plot yield were measured using a digital weighing balance. Root-to-shoot ratios were estimated from the root length (cm) or weight (g) and shoot length (cm) or weight (g). Total soluble solids (TSS in °brix) were estimated using a refractometer, and β-carotene was estimated using the acetone method [73]. Qualitative parameters were recorded with the help of the minimum characterization descriptor of carrot [74]. The tendency of flowering was observed by growing the root shoulders in a polyhouse in Bagalkot, Karnataka, India. Eight weeks of vernalization was performed with a temperature of 0 to 4 °C. A Student t-test assuming equal variance was performed to find the significant differences between test cultivars for individual traits. Furthermore, Pearson’s correlation coefficient analysis was performed to understand the correlation between the quantitative traits using the software package PAST v4.03 [75]. For transcriptome profiling, to ensure accurate analysis, three biological replicates were collected, each replicate consisting of two roots from each cultivar. The harvested roots were promptly frozen in liquid nitrogen to preserve their molecular integrity. The cryopreserved samples were then transported in dry ice on the same day of collection to proceed with the sequencing process.
3.2. RNA Isolation, Library Preparation, and Sequencing
Samples with RNA integrity number (RIN) > 8.0 were subjected to cDNA synthesis and library construction for RNAseq. cDNA libraries were prepared for transcriptome profiling using Hiseq Illumina 4000. Approximately 5–10 ug of total RNA was used to prepare the RNAseq library using the TrueSeq RNA sample preparation Kit (Illumina). In short, poly A-containing mRNA molecules were purified using poly-T oligo-attached magnetic beads. Following purification, the mRNA was fragmented using divalent cations under elevated temperatures. The cleaved RNA fragments were used to synthesize first-strand cDNA using reverse transcriptase and random primers followed by second-strand cDNA synthesis using DNA polymerase I and RNase H. These cDNA fragments underwent an end repair process, the addition of a single ‘A’ base, and then ligation of the adapters. The products were purified and enriched with PCR to create the final cDNA library. Bioanalyzer plots were used at every step to assess mRNA quality, enrichment success, fragmentation, and final library sizes. The size distribution of the sequencing library was determined by gel electrophoresis. Qubit (Q) was used for measuring the quantity of the library before sequencing. For sequencing, constructed libraries were processed on HiSeq 4000, resulting in paired-end (PE) reads of 2 × 100 base pairs. Each library generated a substantial number of reads, ranging from 25 to 30 M. Over 90% of the reads exhibited a Q value of 30 or higher, indicating high-quality sequencing data.
3.3. Reference Based Assembly & Functional Annotation of Differential Expressed Genes (DEG)
Raw reads of six paired-end libraries were passed through quality check using the FASTQC (Version 0.11.9) (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ accessed on 20 May 2021) and analyzed for parameters such as basic statistics, adapter content, sequence length distribution, sequence duplication level, per base sequence quality, and Guanine: Cytosine (GC) content. The raw data of six RNAseq libraries have been submitted to Sequence Read Archives (SRA) in the NCBI database under the project ID PRJNA913450. Depending on the errors reported by FASTQC, additional pre-processing steps were carried out. The adapter was trimmed for each sample individually by applying Trimmomatic v0.36 [76]. The parameters with min quality 30, min length 50, LEADING:28, TRAILING:28, SLIDINGWINDOW:10:28, MINLEN:50 was assigned. After filtering duplicate and low-quality reads, 130,618,433 (92.71%) high-quality cleaned reads remained out of 140,882,952. Bowtie 2 (v.2.4.4) [77] was used to get the index files of the carrot reference genome assembly (ASM162521v1-Refseq GCF_001625215.1) downloaded from ncbi.nlm.nih.gov accessed on 20 May 2021. TopHat [78] was used for mapping the individual libraries with the help of generated index files and reference assembly. A total of 2 mismatches and up to 3 bp indels were allowed in alignment. Mapped RNAseq was assembled with Cufflink’s suite v. 0.17.3 [79] for normalization and transcript quantification by Fragments Per Kilobase of exon per Million (FPKM) fragments mapped values. Differentially expressed gene (DEG) analysis was performed with the cuff-diff tool of the Cufflinks suite v. 0.17.3 [79].
Statistical significance (p-values) was estimated by computing the False Discovery Rate (FDR) using the Benjamini–Hochberg correction [80]. Differences in gene expression were statistically significant when the q-value (FDR-adjusted p-value) was <less than 0.01. The criteria considered for filtering significant DEG included Log2Fold Change (Log2FC) ≥ +1 as up-regulated in Eastern cv. and ≤−1 as up-regulated in Western cv. The list of DEGs with Entrez gene IDs was imported into DAVID 2021 bioinformatics resource [81]. The annotated genes and functionally classified gene clusters (based on enrichment score) were downloaded to understand the biological meaning of the DEGs. The up- (Log2FC ≥ 1) and down-regulated (Log2FC ≤ −1) DEGs were separately imported to DAVID and ShinyGo [82] for functional annotation and Gene Ontology (GO) analysis. GO categories such as Biological Processes (BP), cellular components (CC), MF (Molecular Functions), and KEGG (Kyoto Encyclopaedia of Genes and Genomes) pathways for up- and down-regulated genes were downloaded and compared with our results.
3.4. Gene Regulatory Network (GRN) Analysis with Arabidopsis thaliana Homologs
GRN analysis was carried out with the list of genes identified as VC-responsive genes in Arabidopsis thaliana (AT) [24,33]. The upregulated genes of test carrot cvs. were extracted from the DEGs and subjected to Arabidospis thaliana (AT) orthologs search in “g:orth” in g:Profiler web browser. In brief, a total of 1794 out of 3544 DEGs were successfully obtained with AT gene ID. Then, obtained AT gene IDs were searched for common genes in 1222 AT genes that are considered key VC regulatory genes [33]. The common genes from this study and by Zhang et al. [33] were further sorted for up-and down-regulated genes for corresponding test carrot cvs. DEGs. A total of 198 DEGs out of 1794 DEGs perfectly matched the list of VC regulatory genes of Zhang et al. [33]. A total of 89 DEGs up-regulated in Eastern cv. (Tables S11 and S12) and 109 DEGs up-regulated in Western cv. (Tables S14 and S15) were identified as VC regulatory genes/transcriptional factors for GRN construction.
3.5. Expression Validation of DEGs by qPCR
A total of nine genes identified as DEGs were selected for the validation of gene expression using quantitative real-time PCR. The most stably expressed reference gene “Actin” was used as an endogenous control in the root qPCR experiment to validate RNAseq target genes. The details of genes used for qPCR validation are presented in Table S1. The relative expression levels of target genes were calculated using the 2−ΔΔCt method [83] with Actin as an internal control. The RNA pools used in the qRT-PCR analyses were extracted from three independent samples from Eastern (UHSBC-23-1) and Western (UHSBC-100) carrot cvs. storage root tissue.
4. Conclusions
The present study provides a comprehensive understanding of the development of storage roots in Western carrot cv. by elucidating how carbon flow influences phenylpropanoid biosynthesis and stress response pathways, while also contributing to carbohydrate metabolism, starch biosynthesis, and secondary metabolite formation. These processes play crucial roles in the initiation and development of carrot storage roots during domestication and adaptation in ecologically distinct regions. The transcript data generated in the present study is valuable for global carrot breeders to understand the domestication genes between evolutionarily related test cvs.
The enrichment of DEGs associated with photosynthesis, carbon metabolism, and carotenoid-related genes in Western cv. underscores its potential for nutritious and uniformly deep-colored carrots. Conversely, Eastern cv. exhibits a significant enrichment in genes responding to various (a)biotic stresses, along with VC genes. For effective plant breeding programs, a comprehensive understanding of VC-responsive genes controlling xylem and phloem patterning pathways is essential. Consequently, genetic engineering of master regulators of VC patterning genes would bring a uniform xylem-phloem patterning in Eastern carrot cvs.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12193449/s1, Table S1: List of Eastern cultivar Upregulated DEGs Annotated in David annotation web browser using Daucus carota subsp. sativus as a reference; Table S2: List of Western Upregulated DEGs Annotated in David Annotation web browser using Daucus carota subsp. sativus as a reference; Table S3: Gene ontology categorization into Biological Processes (BP), Cellular Component (CC) and Molecular functions (MF) and KEGG in Eastern cultivar; Table S4: Gene ontology categorization into Biological Processes (BP), Cellular Component (CC) and Molecular functions (MF) and KEGG in Western cultivar; Table S5: DEGs involved in cell cycle and Cell wall biosynthesis in Eastern carrots; Table S6: DEGs involved in cell cycle and Cell wall biosynthesis in Western carrots; Table S7: DEGs involved in signal transduction pathways in Eastern carrots; Table S8: DEGs involved in signal transduction pathways in Western carrots; Table S9: DEGs involved in starch and sucrose metabolism in Eastern carrots; Table S10: DEGs involved in starch and sucrose metabolism in Western carrots; Table S11: AT orthology search for Upregulated DEGS in Eastern cultivar and identification of Common genes from Zhang et al., 2019; Table S12: GRN analysis for 89 Vascular cambium regulatory genes upregulated in Eastern cultivar in comparison to AT orthology (Zhang et al., 2019); Table S13; Gene Ontology terms enriched in Eastern cultivar with Vascular Cambium Regulatory genes for BP, CC, MF and KEGG pathways; Table S14: AT orthology search for Uregulated DEGS in Western cultivar and Identification of Common genes from Zhang et al., 2019; Table S15: RN Analysis of 109 Vascular cambium regulatory genes upregulated in Western cultivar in comparison to AT orthology (Zhang et al., 2019); Table S16: Gene Ontology terms enriched in Western carrots with Vascular Cambium Regulatory genes for BP, CC, MF and KEGG pathways; Table S17: List of genes and primer-pairs used for RNAseq validation of root tissue transcriptome by quantitative real-time PCR.
Author Contributions
C.C.K. conducted the experiment and prepared the manuscript; S.S.C. conceived the idea, received funding, analyzed the data, and was involved in manuscript preparation with C.C.K. and G.O.; A.K.B. assisted S.S.C. computational analysis up to DEG identification; G.O. edited, proofread, and improved the quality of the manuscript; S.R. read the manuscript and was involved in the improvement of the manuscript; R.K.M. and H.B.P. facilitated qPCR experimentation and read the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Biotechnology (DBT), Govt of India (DBT-BIOCARe-102/IFD/SAN/3308/2014-15) and Vision Group of Science and Technology, Govt of Karnataka (KSTePS/VGST-RGS-F/2018-19/GRD No. 840/315).
Data Availability Statement
The raw data is available at the NCBI database in the project ID PRJNA913450.
Acknowledgments
Department of Biotechnology (DBT), Govt of India (DBT-BIOCARe-102/IFD/SAN/3308/2014-15) and Vision Group of Science and Technology, Govt of Karnataka (KSTePS/VGST-RGS-F/2018-19/GRD No. 840/315) for financial support; C.C.K. thankful to Department of Science and Technology, Govt of Karnataka (AGR08:2019-20) for doctoral fellowship. C.C.K. and S.S.C. are thankful to M. S. Kulkarni for academic support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; Donoghue, M.J. Plant systematics: A phylogenetic approach. Ecol. Med. 1999, 25, 215. [Google Scholar] [CrossRef]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Bowman, M.; Iovene, M.; Sanseverino, W.; Cavagnaro, P.; et al. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef]
- FAO. The State of Food Security and Nutrition in Africa: Building Resilience for Peace and Food Security; FAO: Rome, Italy, 2017. [Google Scholar]
- FAO. 2020. Available online: https://www.fao.org/faostat/#data/QCL/visualize (accessed on 10 September 2023).
- Simon, P.W.; Geoffriau, E.; Ellison, S.; Iorizzo, M. Carrot carotenoid genetics and genomics. In The Carrot Genome; Springer: Cham, Switzerland, 2019; pp. 247–260. [Google Scholar] [CrossRef]
- Heywood, V.H. Relationships and evolution in the Daucus carota complex. Israel J. Plant Sci. 1983, 32, 51–65. [Google Scholar] [CrossRef]
- Spooner, D.M.; Simon, W.; Senalik, D.; Iorizzo, M. Carrot organelle genomes: Organization, diversity, and inheritance. In The Carrot Genome; Springer: Cham, Switzerland, 2019; pp. 205–223. [Google Scholar] [CrossRef]
- Que, F.; Hou, X.L.; Wang, G.L.; Xu, Z.S.; Tan, G.F.; Li, T.; Wang, Y.H.; Khadr, A.; Xiong, A.S. Advances in research on the carrot, an important root vegetable in the Apiaceae family. Hortic. Res. 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Pérez, M.B.; Carvajal, S.; Beretta, V.; Bannoud, F.; Fangio, M.F.; Berli, F.; Fontana, A.; Salomón, M.V.; Gonzalez, R.; Valerga, L.; et al. Characterization of Purple Carrot Germplasm for Antioxidant Capacity and Root Concentration of Anthocyanins, Phenolics, and Carotenoids. Plants 2023, 12, 1796. [Google Scholar] [CrossRef]
- Purkiewicz, A.; Ciborska, J.; Tańska, M.; Narwojsz, A.; Starowicz, M.; Przybyłowicz, K.E.; Sawicki, T. The Impact of the Method Extraction and Different Carrot Variety on the Carotenoid Profile, Total Phenolic Content and Antioxidant Properties of Juices. Plants 2020, 9, 1759. [Google Scholar] [CrossRef]
- Chaitra, K.C.; Sarvamangala, C.; Manikanta, D.S.; Chaitra, P.A.; Fakrudin, B. Insights into genetic diversity and population structure of Indian carrot (Daucus carota L.) accessions. J. Appl. Genet. 2020, 61, 303–312. [Google Scholar] [CrossRef]
- Clotault, J.; Geoffriau, E.; Lionneton, E.; Briard, M.; Peltier, D. Carotenoid biosynthesis genes provide evidence of geographical subdivision and extensive linkage disequilibrium in the carrot. Theor. Appl. Genet. 2010, 121, 659–672. [Google Scholar] [CrossRef]
- Baranski, R.; Maksylewicz-Kaul, A.; Nothnagel, T.; Cavagnaro, F.; Simon, W.; Grzebelus, D. Genetic diversity of carrot (Daucus carota L.) cultivars revealed by analysis of SSR loci. Genet. Resour. Crop Evol. 2012, 59, 163–170. [Google Scholar] [CrossRef]
- Soufflet-Freslon, V.; Jourdan, M.; Clotault, J.; Huet, S.; Briard, M.; Peltier, D.; Geoffriau, E. Functional gene polymorphism to reveal species history: The case of the CRTISO gene in cultivated carrots. PLoS ONE 2013, 8, e70801. [Google Scholar] [CrossRef]
- Maksylewicz, A.; Baranski, R. Intra-population genetic diversity of cultivated carrot (Daucus carota L.) assessed by analysis of microsatellite markers. Acta Biochim. Pol. 2013, 60, 58–63. [Google Scholar] [CrossRef]
- Grzebelus, D.; Iorizzo, M.; Senalik, D.; Ellison, S.; Cavagnaro, P.; Macko-Podgorni, A.; Heller-Uszynska, K.; Kilian, A.; Nothnagel, T.; Allender, C.; et al. Diversity, genetic mapping, and signatures of domestication in the carrot (Daucus carota L.) genome, as revealed by Diversity Arrays Technology (DArT) markers. Mol. Breed. 2014, 33, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Senalik, D.A.; Ellison, S.L.; Grzebelus, D.; Cavagnaro, F.; Allender, C.; Brunet, J.; Spooner, D.M.; Van Deynze, A.; Simon, W. Genetic structure and domestication of carrot (Daucus carota subsp. sativus) (Apiaceae). Am. J. Bot. 2013, 100, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Ellison, S.L.; Luby, C.H.; Corak, K.E.; Coe, K.M.; Senalik, D.; Iorizzo, M.; Goldman, I.L.; Simon, W.; Dawson, J.C. Carotenoid presence is associated with the or gene in domesticated carrots. Genetics 2018, 210, 1497–1508. [Google Scholar] [CrossRef]
- Ellison, S. Carrot domestication. In The Carrot Genome; Springer: Cham, Switzerland, 2019; pp. 77–91. [Google Scholar] [CrossRef]
- Simon, W.; Freeman, R.E.; Vieira, J.V.; Boiteux, L.S.; Briard, M.; Nothnagel, T.; Michalik, B.; Kwon, Y.S. Carrot. In Handbook of Plant Breeding: Vegetables II: Fabaceae, Liliaceae, Solanaceae, and Umbelliferae; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; pp. 327–357. [Google Scholar] [CrossRef]
- Machaj, G.; Bostan, H.; Macko-Podgórni, A.; Iorizzo, M.; Grzebelus, D. Comparative transcriptomics of root development in wild and cultivated carrots. Genes 2018, 9, 431. [Google Scholar] [CrossRef]
- Ondrasek, G.; Clode, P.L.; Kilburn, M.R.; Guagliardo, P.; Romić, D.; Rengel, Z. Zinc and Cadmium Mapping in the Apical Shoot and Hypocotyl Tissues of Radish by High-Resolution Secondary Ion Mass Spectrometry (NanoSIMS) after Short-Term Exposure to Metal Contamination. Int. J. Environ. Res. Public Health 2019, 16, 373. [Google Scholar] [CrossRef]
- Ondrasek, G.; Rengel, Z.; Clode, P.L.; Kilburn, M.R.; Guagliardo, P.; Romic, D. Zinc and cadmium mapping by NanoSIMS within the root apex after short-term exposure to metal contamination. Ecotoxicol. Environ. Saf. 2019, 30, 571–578. [Google Scholar] [CrossRef]
- Hoang, N.V.; Choe, G.; Zheng, Y.; Fandiño, A.C.A.; Sung, I.; Hur, J.; Kamran, M.; Park, C.; Kim, H.; Ahn, H.; et al. Identification of conserved gene-regulatory networks that integrate environmental sensing and growth in the root cambium. Curr. Biol. 2020, 30, 2887–2900. [Google Scholar] [CrossRef]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic stresses: General defenses of land plants and chances for engineering multi-stress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef]
- Ondrasek, G.; Rengel, Z.; Maurović, N.; Kondres, N.; Filipović, V.; Savić, R.; Blagojević, B.; Tanaskovik, V.; Gergichevich, C.M.; Romić, D. Growth and Element Uptake by Salt-Sensitive Crops under Combined NaCl and Cd Stresses. Plants 2021, 10, 1202. [Google Scholar] [CrossRef]
- Karlova, R.; Boer, D.; Hayes, S.; Testerink, C. Root plasticity under abiotic stress. Plant Physiol. 2021, 187, 1057–1070. [Google Scholar] [CrossRef]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth–defense tradeoffs in plants: A balancing act to optimize fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef] [PubMed]
- Kudo, M.; Kidokoro, S.; Yoshida, T.; Mizoi, J.; Kojima, M.; Takebayashi, Y.; Sakakibara, H.; Fernie, A.R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. A gene-stacking approach to overcome the trade-off between drought stress tolerance and growth in Arabidopsis. Plant J. 2019, 97, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Duruflé, H.; Déjean, S. Multi-omics data integration in the context of plant abiotic stress signaling. In Plant Abiotic Stress Signaling; Humana: New York, NY, USA, 2023; pp. 295–318. [Google Scholar] [CrossRef]
- Zachgo, S. Nuclear redox processes in land plant development and stress adaptation. Biol. Chem. 2023, 404, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Eswaran, G.; Alonso-Serra, J.; Kucukoglu, M.; Xiang, J.; Yang, W.; Helariutta, Y. Transcriptional regulatory framework for vascular cambium development in Arabidopsis roots. Nat. Plants 2019, 5, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.R.; Rhee, J.; Choi, C.S.; Jo, J.S.; Shin, Y.K.; Song, J.W.; Kim, S.-H.; Lee, J.G. Morphological and Biochemical Variation in Carrot Genetic Resources Grown under Open Field Conditions: The Selection of Functional Genotypes for a Breeding Program. Agronomy 2022, 12, 553. [Google Scholar] [CrossRef]
- Ma, J.; Li, J.; Xu, Z.; Wang, F.; Xiong, A. Transcriptome profiling of genes involving in carotenoid biosynthesis and accumulation between leaf and root of carrot (Daucus carota L.). Acta Biochim. Biophys. Sin. 2018, 50, 481–490. [Google Scholar] [CrossRef]
- Yu, R.; Wang, J.; Xu, L.; Wang, Y.; Wang, R.; Zhu, X.; Sun, X.; Luo, X.; Xie, Y.; Everlyne, M.; et al. Transcriptome Profiling of Taproot Reveals Complex Regulatory Networks during Taproot Thickening in Radish (Raphanus sativus L.). Front. Plant Sci. 2016, 7, 1210. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Clausen, S.K.; Rasmussen, S.K. Transcriptome analysis reveals candidate genes related to anthocyanin biosynthesis in different carrot genotypes and tissues. Plants 2020, 9, 344. [Google Scholar] [CrossRef]
- Gohar, T.; Payal, A.; Grant, M.; Anil, K. MAPK machinery in plants: Recognition and response to different stresses through multiple signal transduction pathways. Plant Signal. Behav. 2010, 5, 1372–1380. [Google Scholar] [CrossRef]
- Yao, X.; Feng, H.; Yu, Y.; Dong, A.; Shen, W.H. SDG2-mediated H3K4 methylation is required for proper Arabidopsis root growth and development. PLoS ONE 2013, 8, e56537. [Google Scholar] [CrossRef] [PubMed]
- Banakar, P.; Hada, A.; Papolu, K.; Rao, U. Simultaneous RNAi knockdown of three FMRFamide-like peptide genes, Mi-flp1, Mi-flp12, and Mi-flp18 provides resistance to root-knot nematode, Meloidogyne incognita. Front. Microbiol. 2020, 11, 2690. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Moreno, A.; Redondo-Gil, M.; Benavides, J.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Combined effect of water loss and wounding stress on gene activation of metabolic pathways associated with phenolic biosynthesis in carrot. Front. Plant Sci. 2015, 6, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Usuda, H.; Rouhier, H.; Demura, T.; Fukuda, H. Development of sink capacity of the “storage root” in a radish variety with a low ratio of “storage root” to shoot. Plant Cell Physiol. 1999, 40, 1210–1218. [Google Scholar] [CrossRef][Green Version]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of waterlogging tolerance in plants: Research progress and prospects. Front. Plant Sci. 2021, 11, 627331. [Google Scholar] [CrossRef]
- Wang, X.; An, Y.; Xu, P.; Xiao, J. Functioning of PPR proteins in organelle RNA metabolism and chloroplast biogenesis. Front. Plant Sci. 2021, 12, 627501. [Google Scholar] [CrossRef]
- Arias, D.; Maldonado, J.; Silva, H.; Stange, C. A de novo transcriptome analysis revealed that photomorphogenic genes are required for carotenoid synthesis in the dark-grown carrot taproot. Mol. Genet. Genom. 2020, 295, 1379–1392. [Google Scholar] [CrossRef]
- Sung, S.; Amasino, R.M. Vernalization and epigenetics: How plants remember winter. Curr. Opin. Plant Biol. 2004, 7, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Hsu, C.C.; Du, Y.; Zhu, P.; Zhao, C.; Fu, X.; Zhu, J.K. Mapping proteome-wide targets of protein kinases in plant stress responses. Proc. Natl. Acad. Sci. USA 2020, 117, 3270–3280. [Google Scholar] [CrossRef]
- Wolf, S. Cell wall signaling in plant development and defense. Annu. Rev. Plant Biol. 2022, 73, 323–353. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.T.; Heidari, P. Cell Signaling in Model Plants. Int. J. Mol. Sci. 2020, 21, 6062. [Google Scholar] [CrossRef] [PubMed]
- Rong, J.; Lammers, Y.; Strasburg, J.L.; Schidlo, N.S.; Ariyurek, Y.; De Jong, T.J.; Klinkhamer, G.; Smulders, M.J.; Vrieling, K. New insights into domestication of carrot from root transcriptome analyses. BMC Genom. 2014, 15, 895. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Paque, S.; Mouille, G.; Grandont, L.; Alabadí, D.; Gaertner, C.; Goyallon, A.; Muller, P.; Primard-Brisset, C.; Sormani, R.; Blázquez, M.A.; et al. AUXIN BINDING PROTEIN1 links cell wall remodeling, auxin signaling, and cell expansion in Arabidopsis. Plant Cell 2014, 26, 280–295. [Google Scholar] [CrossRef]
- Guo, M.; Simmons, C.R. Cell number counts--the fw2.2 and CNR genes and implications for controlling plant fruit and organ size. Plant Sci. Int. J. Exp. Plant Biol. 2011, 181, 1–7. [Google Scholar] [CrossRef]
- Treves, H.; Küken, A.; Arrivault, S.; Ishihara, H.; Hoppe, I.; Erban, A.; Höhne, M.; Moraes, T.A.; Kopka, J.; Szymanski, J.; et al. Carbon flux through photosynthesis and central carbon metabolism show distinct patterns between algae, C3 and C4 plants. Nat. Plants 2022, 8, 78–91. [Google Scholar] [CrossRef]
- Yu, R.; Xu, L.; Zhang, W.; Wang, Y.; Luo, X.; Wang, R.; Zhu, X.; Xie, Y.; Karanja, B.; Liu, L. De novo taproot transcriptome sequencing and analysis of major genes involved in sucrose metabolism in radish (Raphanus sativus L.). Front. Plant Sci. 2016, 7, 585–589. [Google Scholar] [CrossRef]
- Xiao, X.; Hu, Y.; Zhang, M.; Si, S.; Zhou, H.; Zhu, W.; Ge, F.; Wu, C.; Fan, S. Transcriptome profiling reveals the genes involved in tuberous root expansion in Pueraria (Pueraria montana var. thomsonii). BMC Plant Biol. 2023, 23, 338. [Google Scholar] [CrossRef]
- Nakano, Y.; Yamaguchi, M.; Endo, H.; Rejab, N.A.; Ohtani, M. NAC-MYB-based transcriptional regulation of secondary cell wall biosynthesis in land plants. Front. plant sci. 2015, 6, 288. [Google Scholar] [CrossRef]
- Wang, X.; Mäkilä, R.; Mähönen, A.P. From procambium patterning to cambium activation and maintenance in the Arabidopsis root. Curr. Opin. Plant Biol. 2023, 75, 102404. [Google Scholar] [CrossRef] [PubMed]
- Chrapačienė, S.; Rasiukevičiūtė, N.; Valiuškaitė, A. Biocontrol of Carrot Disease-Causing Pathogens Using Essential Oils. Plants 2021, 10, 2231. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Niu, X.; Li, S.; Li, Q.; Fu, S.; Wang, C.; Wu, S. Intercellular signaling across plasmodesmata in vegetable species. Veg. Res. 2023, 3, 22. [Google Scholar] [CrossRef]
- Zhao, L.; Nakazawa, M.; Takase, T.; Manabe, K.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Matsui, M. Overexpression of LSH1, a member of an uncharacterised gene family, causes enhanced light regulation of seedling development. Plant J. 2004, 37, 694–706. [Google Scholar] [CrossRef]
- Xu, K.; Jourquin, J.; Njo, M.F.; Nguyen, L.; Beeckman, T.; Fernandez, A.I. The Phloem Intercalated with Xylem-Correlated 3 Receptor-Like Kinase Constitutively Interacts with Brassinosteroid Insensitive 1-Associated Receptor Kinase 1 and Is Involved in Vascular Development in Arabidopsis. Front. Plant Sci. 2022, 12, 706633. [Google Scholar] [CrossRef]
- Patrick, J.W.; Offler, C.E. Post-sieve element transport of photoassimilates in sink regions. J. Exp. Bot. 1996, 47, 1165–1177. [Google Scholar] [CrossRef]
- Li, S. The Arabidopsis thaliana TCP transcription factors: A broadening horizon beyond development. Plant Signal. Behav. 2015, 10, 1044192. [Google Scholar] [CrossRef]
- Huang, X.; Chen, X.G.; Armbruster, A. Comparative performance of transcriptome assembly methods for non-model organisms. BMC Genom. 2016, 17, 523. [Google Scholar] [CrossRef]
- Hoang, N.V.; Park, C.; Kamran, M.; Lee, J.Y. Gene regulatory network guided investigations and engineering of storage root development in root crops. Front. Plant Sci. 2020, 11, 762. [Google Scholar] [CrossRef]
- Macko-Podgórni, A.; Stelmach, K.; Kwolek, K.; Machaj, G.; Ellison, S.; Senalik, D.A.; Simon, P.W.; Grzebelus, D. Mining for candidate genes controlling secondary growth of the carrot storage root. Int. J. Mol. Sci. 2020, 21, 4263. [Google Scholar] [CrossRef]
- Coenye, T. Do results obtained with RNA-sequencing require independent verification? Biofilm 2021, 3, 100043. [Google Scholar] [CrossRef] [PubMed]
- Stupnikov, A.; McInerney, C.E.; Savage, K.I.; McIntosh, S.A.; Emmert-Streib, F.; Kennedy, R.; Salto-Tellez, M.; Prise, K.M.; McArt, D.G. Robustness of differential gene expression analysis of RNA-seq. Comput. Struct. Biotechnol. J. 2021, 19, 3470–3481. [Google Scholar] [CrossRef] [PubMed]
- Sampathkumar, N.K.; Sundaram, V.K.; Danthi, P.S.; Barakat, R.; Solomon, S.; Mondal, M.; Carre, I.; El Jalkh, T.; Padilla-Ferrer, A.; Grenier, J.; et al. RNA-Seq is not required to determine stable reference genes for qPCR normalization. PLoS Comput. Biol. 2022, 18, 1009868. [Google Scholar] [CrossRef] [PubMed]
- Cholin, S.S.; Poleshi, C.A.; Manikanta, D.S.; Christopher, C. Exploring the genomic resources of carrot for cross-genera transferability and phylogenetic assessment among orphan spices and vegetables of Apiaceae family. Hortic. Environ. Biotechnol. 2019, 60, 81–93. [Google Scholar] [CrossRef]
- Perveen, N.; Cholin, S.S.; Hipparagi, K.; Prabhuling, G.; Murthy, B.N.S.; Peerjade, D. Molecular diversity assessment among the pomegranate genotypes belonging to diverse genetic background using microsatellite markers. Acta Physiol. Plant. 2023, 45, 92. [Google Scholar] [CrossRef]
- Harborne, J.B.; Harborne, J.B. Phenolic compounds. In Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis; Springer: Dordrecht, The Netherlands, 1984; pp. 37–99. [Google Scholar] [CrossRef]
- IPGRI. Descriptors for Wild and Cultivated Carrots (Daucus carota L.); International Plant Genetic Resources Institute: Rome, Italy, 1998. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Trapnell, C.; Salzberg, S.L. How to map billions of short reads onto genomes. Nat. Biotechnol. 2009, 27, 455–457. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J. Tran-script assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple hypothesis testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Quail, M.A.; Kozarewa, I.; Smith, F.; Scally, A.; Stephens, J.; Durbin, R. A large genome center’s improvement to the Illumina sequencing system. Nat. Methods 2008, 5, 1005–1010. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).