
Phenotypic Diversity in Leaf Cuticular Waxes in Brassica carinata Accessions


Abstract
:1. Introduction
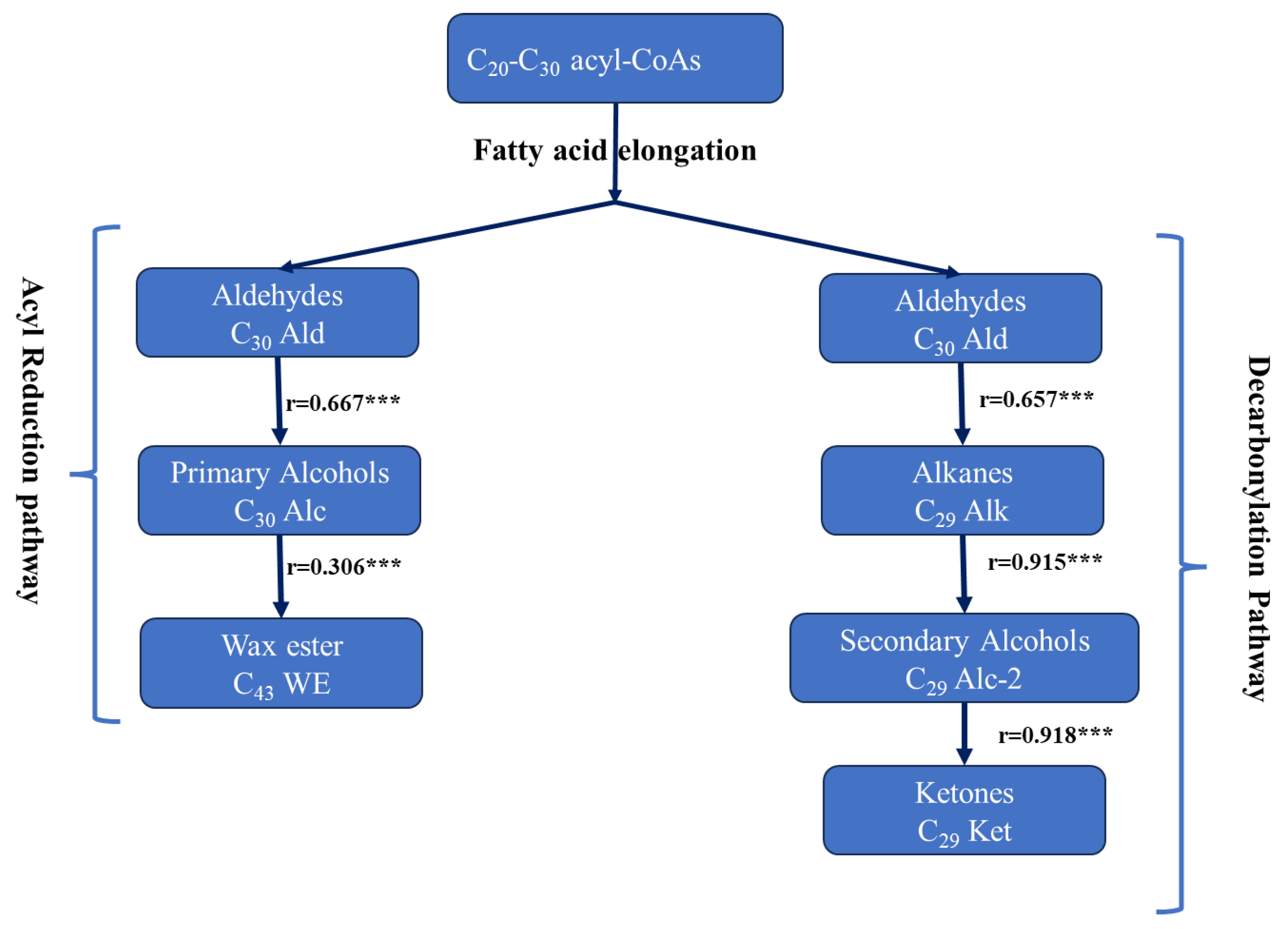
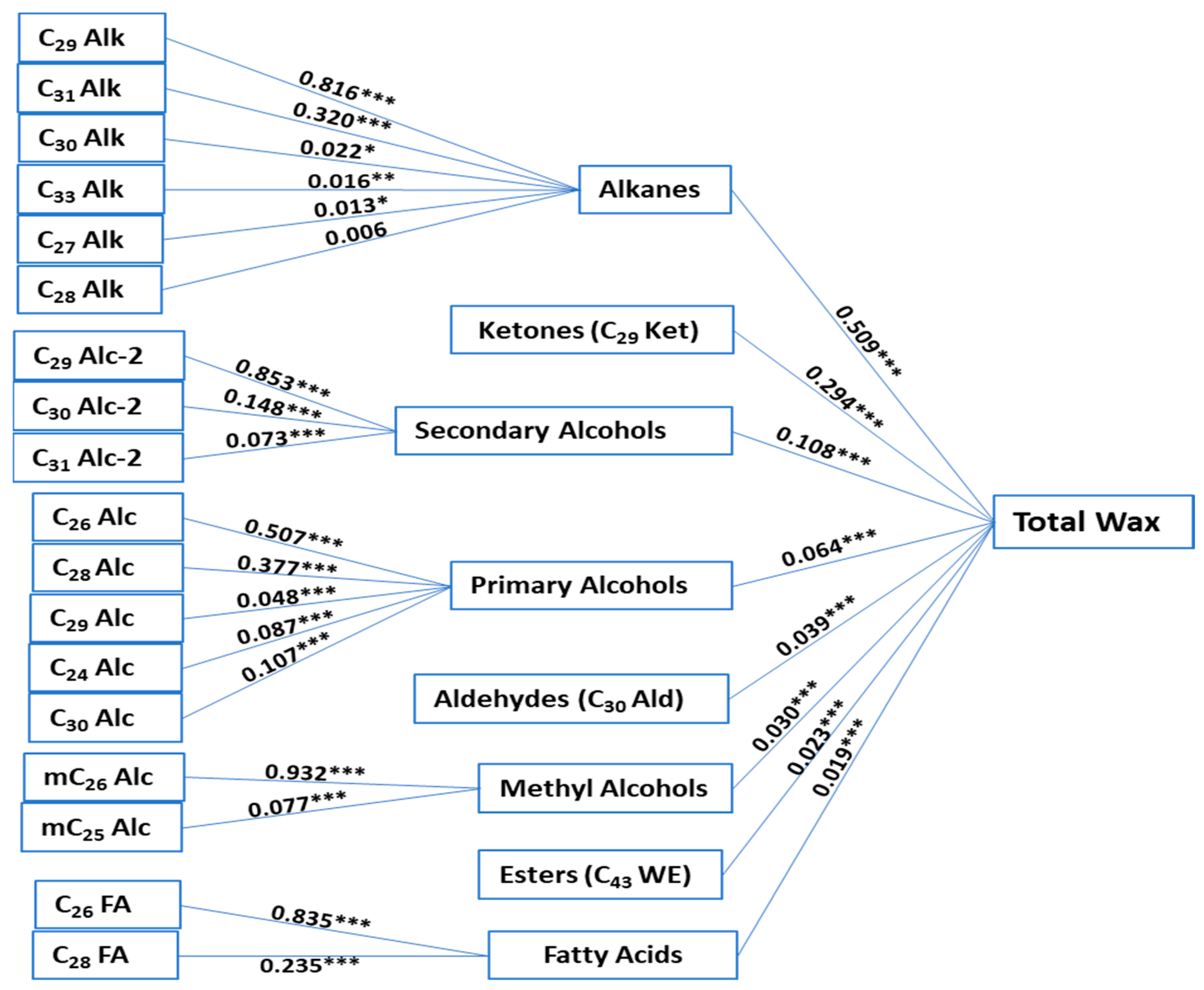
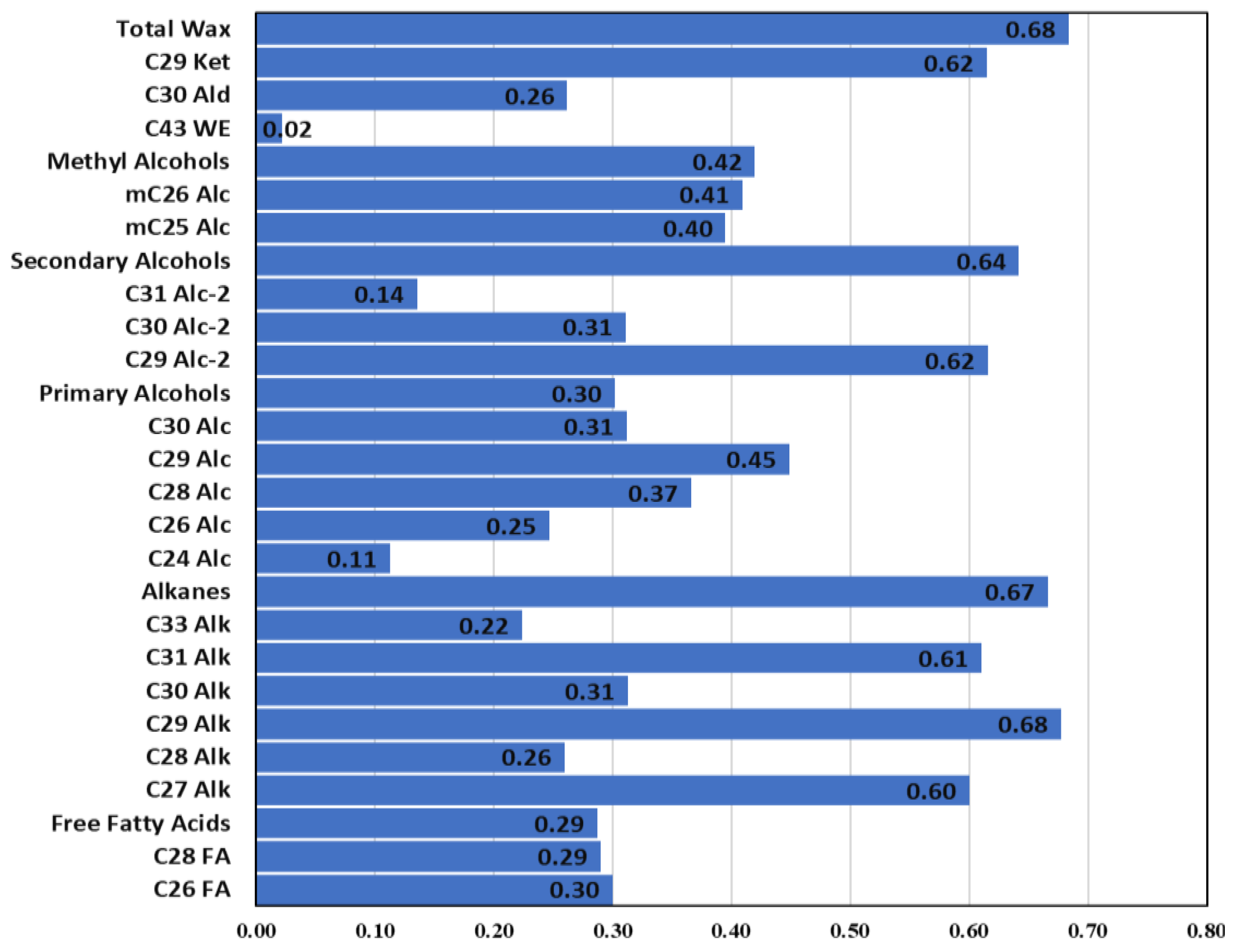
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Wax Extraction and Analyses
4.3. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drenth, A.C.; Olsen, D.B.; Cabot, P.E.; Johnson, J.J. Compression ignition engine performance and emission evaluation of industrial oilseed biofuel feedstocks camelina, carinata, and pennycress across three fuel pathways. Fuel 2014, 136, 143–155. [Google Scholar] [CrossRef]
- Moser, B.R. Biodiesel production, properties, and feedstocks. Vitr. Cell. Dev. Biol. Plant 2009, 45, 229–266. [Google Scholar] [CrossRef]
- Pinzi, S.; Garcia, I.L.; Lopez-Gimenez, F.J.; de Castro, M.D.L.; Dorado, G.; Dorado, M.P. The Ideal Vegetable Oil-based Biodiesel Composition: A Review of Social, Economical and Technical Implications. Energy Fuels 2009, 23, 2325–2341. [Google Scholar] [CrossRef]
- Cardone, M.; Prati, M.V.; Rocco, V.; Seggiani, M.; Senatore, A.; Vitolo, S. Brassica carinata as an Alternative Oil Crop for the Production of Biodiesel in Italy: Engine Performance and Regulated and Unregulated Exhaust Emissions. Environ. Sci. Technol. 2002, 36, 4656–4662. [Google Scholar] [CrossRef] [PubMed]
- Dorado, M.P.; Ballesteros, E.; López, F.J.; Mittelbach, M. Optimization of Alkali-Catalyzed Transesterification of Brassica Carinata Oil for Biodiesel Production. Energy Fuels 2004, 18, 77–83. [Google Scholar] [CrossRef]
- Moeller, D.; Sieverding, H.L.; Stone, J.J. Comparative Farm-Gate Life Cycle Assessment of Oilseed Feedstocks in the Northern Great Plains. Biophys. Econ. Resour. Qual. 2017, 2, 13. [Google Scholar] [CrossRef]
- Gesch, R.W.; Isbell, T.A.; Oblath, E.A.; Allen, B.L.; Archer, D.W.; Brown, J.; Hatfield, J.L.; Jabro, J.D.; Kiniry, J.R.; Long, D.S.; et al. Comparison of several Brassica species in the north central U.S. for potential jet fuel feedstock. Ind. Crops Prod. 2015, 75, 2–7. [Google Scholar] [CrossRef]
- Del Gatto, A.; Melilli, M.G.; Raccuia, S.A.; Pieri, S.; Mangoni, L.; Pacifico, D.; Signor, M.; Duca, D.; Foppa Pedretti, E.; Mengarelli, C. A comparative study of oilseed crops (Brassica napus L. subsp. oleifera and Brassica carinata A. Braun) in the biodiesel production chain and their adaptability to different Italian areas. Ind. Crops Prod. 2015, 75, 98–107. [Google Scholar] [CrossRef]
- Marillia, E.-F.; Francis, T.; Falk, K.C.; Smith, M.; Taylor, D.C. Palliser’s promise: Brassica carinata, An emerging western Canadian crop for delivery of new bio-industrial oil feedstocks. Biocatal. Agric. Biotechnol. 2014, 3, 65–74. [Google Scholar] [CrossRef]
- Hossain, Z.; Johnson, E.N.; Wang, L.; Blackshaw, R.E.; Cutforth, H.; Gan, Y. Plant establishment, yield and yield components of Brassicaceae oilseeds as potential biofuel feedstock. Ind. Crops Prod. 2019, 141, 111800. [Google Scholar] [CrossRef]
- Seepaul, R. Carinata, the Jet Fuel Cover Crop: 2016 Production Recommendations for the Southeastern United States; SS-AGR-384; Agronomy Department, UF/IFAS Extension; University of Florida: Gainesville, FL, USA, 2015; pp. 1–8. [Google Scholar]
- Gasol, C.M.; Gabarrell, X.; Anton, A.; Rigola, M.; Carrasco, J.; Ciria, P.; Solano, M.L.; Rieradevall, J. Life cycle assessment of a Brassica carinata bioenergy cropping system in southern Europe. Biomass Bioenergy 2007, 31, 543–555. [Google Scholar] [CrossRef]
- Gugel, R.K.; Falk, K.C. Agronomic and seed quality evaluation of Camelina sativa in western Canada. Can. J. Plant Sci. 2006, 86, 1047–1058. [Google Scholar] [CrossRef]
- Padbury, G.; Waltman, S.; Caprio, J.; Coen, G.; McGinn, S.; Mortensen, D.; Nielsen, G.; Sinclair, R. Agroecosystems and Land Resources of the Northern Great Plains. Agron. J. 2002, 94, 251–261. [Google Scholar] [CrossRef]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.M.; Turner, N.C.; Osmond, C.B. Mechanisms of drought resistance. In The Physiology and Biochemistry of Drought Resistance in Plants; Paleg, L.G., Aspinall, D., Eds.; Academic Press: New York, NY, USA, 1981; pp. 15–37. [Google Scholar]
- Jenks, M.A.; Joly, R.J.; Peters, P.J.; Rich, P.J.; Axtell, J.D.; Ashworth, E.N. Chemically Induced Cuticle Mutation Affecting Epidermal Conductance to Water Vapor and Disease Susceptibility in Sorghum bicolor (L.) Moench. Plant Physiol. 1994, 105, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Yeats, T.H.; Rose, J.K.C. The Formation and Function of Plant Cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, T.; Wynne Griffiths, D. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef] [PubMed]
- Kosma, D.K.; Bourdenx, B.; Bernard, A.; Parsons, E.P.; Lü, S.; Joubès, J.; Jenks, M.A. The Impact of Water Deficiency on Leaf Cuticle Lipids of Arabidopsis. Plant Physiol. 2009, 151, 1918–1929. [Google Scholar] [CrossRef]
- Shaheenuzzamn, M.; Shi, S.D.; Sohail, K.; Wu, H.Q.; Liu, T.X.; An, P.P.; Wang, Z.H.; Hasanuzzaman, M. Regulation of cuticular wax biosynthesis in plants under abiotic stress. Plant Biotechnol. Rep. 2021, 15, 1–12. [Google Scholar] [CrossRef]
- Shaheenuzzamn, M.; Liu, T.X.; Shi, S.D.; Wu, H.Q.; Wang, Z.H. Research Advances on Cuticular Waxes Biosynthesis in Crops: A Review. Int. J. Agric. Biol. 2019, 21, 911–921. [Google Scholar] [CrossRef]
- Ormeno, E.; Ruffault, J.; Gutigny, C.; Madrigal, J.; Guijarro, M.; Hernando, C.; Ballini, C. Increasing cuticular wax concentrations in a drier climate promote litter flammability. For. Ecol. Manag. 2020, 473, 118242. [Google Scholar] [CrossRef]
- Buschhaus, C.; Jetter, R. Composition and Physiological Function of the Wax Layers Coating Arabidopsis Leaves: β-Amyrin Negatively Affects the Intracuticular Water Barrier. Plant Physiol. 2012, 160, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Tomasi, P.; Dyer, J.M.; Jenks, M.A.; Abdel-Haleem, H. Characterization of leaf cuticular wax classes and constituents in a spring Camelina sativa diversity panel. Ind. Crops Prod. 2018, 112, 247–251. [Google Scholar] [CrossRef]
- Tomasi, P.; Wang, H.; Lohrey, G.; Park, S.; Dyer, J.; Jenks, M.; Abdel-Haleem, H.J.I.C. Characterization of leaf cuticular waxes and cutin monomers of Camelina sativa and closely-related Camelina species. Ind. Crops Prod. 2017, 98, 130–138. [Google Scholar] [CrossRef]
- Cameron, K.D.; Teece, M.A.; Smart, L.B. Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco. Plant Physiol. 2006, 140, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Samdur, M.Y.; Manivel, P.; Jain, V.K.; Chikani, B.M.; Gor, H.K.; Desai, S.; Misra, J.B. Genotypic differences and water-deficit induced enhancement in epicuticular wax load in peanut. Crop Sci. 2003, 43, 1294–1299. [Google Scholar] [CrossRef]
- Kim, K.S.; Park, S.H.; Kim, D.K.; Jenks, M.A. Influence of water deficit on leaf cuticular waxes of soybean (Glycine max [L.] Merr.). Int. J. Plant Sci. 2007, 168, 307–316. [Google Scholar] [CrossRef]
- Mohammed, S.; Huggins, T.; Mason, E.; Beecher, F.; Chick, C.; Sengodon, P.; Paudel, A.; Ibrahim, A.; Tilley, M.; Hays, D. Mapping the genetic loci regulating leaf epicuticular wax, canopy temperature, and drought susceptibility index in Triticum aestivum. Crop Sci. 2021, 61, 2294–2305. [Google Scholar] [CrossRef]
- Mohammed, S.; Huggins, T.D.; Beecher, F.; Chick, C.; Sengodon, P.; Mondal, S.; Paudel, A.; Ibrahim, A.M.H.; Tilley, M.; Hays, D.B. The Role of Leaf Epicuticular Wax in the Adaptation of Wheat (Triticum aestivum L.) to High Temperatures and Moisture Deficit Conditions. Crop Sci. 2018, 58, 679–689. [Google Scholar] [CrossRef]
- Bondada, B.R.; Oosterhuis, D.M.; Murphy, J.B.; Kim, K.S. Effect of water stress on the epicuticular wax composition and ultrastructure of cotton (Gossypium hirsutum L.) leaf, bract, and boll. Environ. Exp. Bot. 1996, 36, 61–69. [Google Scholar] [CrossRef]
- Lee, S.B.; Suh, M.C. Advances in the understanding of cuticular waxes in Arabidopsis thaliana and crop species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Joubès, J. Arabidopsis cuticular waxes: Advances in synthesis, export and regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef] [PubMed]
- Baales, J.; Zeisler-Diehl, V.V.; Schreiber, L. Analysis of Extracellular Cell Wall Lipids: Wax, Cutin, and Suberin in Leaves, Roots, Fruits, and Seeds. In Plant Lipids: Methods and Protocols; Bartels, D., Dörmann, P., Eds.; Springer US: New York, NY, USA, 2021; pp. 275–293. [Google Scholar]
- Samuels, L.; Kunst, L.; Jetter, R. Sealing Plant Surfaces: Cuticular Wax Formation by Epidermal Cells. Annu. Rev. Plant Biol. 2008, 59, 683–707. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.A. The influence of environment on leaf wax development in Brassica oleracea var. gemmifera. New Phytol. 1974, 73, 955–966. [Google Scholar] [CrossRef]
- Lee, S.B.; Kim, H.; Kim, R.J.; Suh, M.C. Overexpression of Arabidopsis MYB96 confers drought resistance in Camelina sativa via cuticular wax accumulation. Plant Cell Rep. 2014, 33, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.A.; Clemens, S.; Zachgo, S.; Giblin, E.M.; Taylor, D.C.; Kunst, L. CUT1, an Arabidopsis Gene Required for Cuticular Wax Biosynthesis and Pollen Fertility, Encodes a Very-Long-Chain Fatty Acid Condensing Enzyme. Plant Cell 1999, 11, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Zhang, X.; Lu, X.; Chen, G.; Chen, Z.-H. Molecular and Evolutionary Mechanisms of Cuticular Wax for Plant Drought Tolerance. Front. Plant Sci. 2017, 8, 621. [Google Scholar] [CrossRef] [PubMed]
- Kolattukudy, P.E. Enzymatic synthesis of fatty alcohols in Brassica oleracea. Arch. Biochem. Biophys. 1971, 142, 701–709. [Google Scholar] [CrossRef]
- Tassone, E.E.; Lipka, A.E.; Tomasi, P.; Lohrey, G.T.; Qian, W.; Dyer, J.M.; Gore, M.A.; Jenks, M.A. Chemical variation for leaf cuticular waxes and their levels revealed in a diverse panel of Brassica napus L. Ind. Crops Prod. 2016, 79, 77–83. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Campo, C.; Tortosa, M.E.; Tewari, I.; Tewari, J.P. Epicuticular Wax Columns in Cultivated Brassica Species and in their Close Wild Relatives. Ann. Bot. 1999, 83, 515–519. [Google Scholar] [CrossRef]
- Gunasinghe, N.; You, M.P.; Clode, P.L.; Barbetti, M.J. Mechanisms of resistance in Brassica carinata, B. napus and B. juncea to Pseudocercosporella capsellae. Plant Pathol. 2016, 65, 888–900. [Google Scholar] [CrossRef]
- Bodnaryk, R.P. Leaf epicuticular wax, an antixenotic factor in Brassicaceae that affects the rate and pattern of feeding of flea beetles, Phyllotreta cruciferae (Goeze). Can. J. Plant Sci. 1992, 72, 1295–1303. [Google Scholar] [CrossRef]
- Ni, Y.; Guo, Y.J.; Han, L.; Tang, H.; Conyers, M. Leaf cuticular waxes and physiological parameters in alfalfa leaves as influenced by drought. Photosynthetica 2012, 50, 458–466. [Google Scholar] [CrossRef]
- Bourdenx, B.; Bernard, A.; Domergue, F.; Pascal, S.; Léger, A.; Roby, D.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; et al. Overexpression of Arabidopsis ECERIFERUM1 Promotes Wax Very-Long-Chain Alkane Biosynthesis and Influences Plant Response to Biotic and Abiotic Stresses. Plant Physiol. 2011, 156, 29–45. [Google Scholar] [CrossRef]
- Kim, K.S.; Park, S.H.; Jenks, M.A. Changes in leaf cuticular waxes of sesame (Sesamum indicum L.) plants exposed to water deficit. J. Plant Physiol. 2007, 164, 1134–1143. [Google Scholar] [CrossRef]
- Xu, X.; Xiao, L.; Feng, J.; Chen, N.; Chen, Y.; Song, B.; Xue, K.; Shi, S.; Zhou, Y.; Jenks, M.A. Cuticle lipids on heteromorphic leaves of Populus euphratica Oliv. growing in riparian habitats differing in available soil moisture. Physiol. Plant. 2016, 158, 318–330. [Google Scholar] [CrossRef]
- Li, H.; Guo, Y.; Cui, Q.; Zhang, Z.; Yan, X.; Ahammed, G.J.; Yang, X.; Yang, J.; Wei, C.; Zhang, X. Alkanes (C29 and C31)-Mediated Intracuticular Wax Accumulation Contributes to Melatonin- and ABA-Induced Drought Tolerance in Watermelon. J. Plant Growth Regul. 2020, 39, 1441–1450. [Google Scholar] [CrossRef]
- Post-Beittenmiller, D. Biochemistry and Molecular Biology of Wax Production in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 405–430. [Google Scholar] [CrossRef]
- Le Provost, G.; Domergue, F.; Lalanne, C.; Ramos Campos, P.; Grosbois, A.; Bert, D.; Meredieu, C.; Danjon, F.; Plomion, C.; Gion, J.-M. Soil water stress affects both cuticular wax content and cuticle-related gene expression in young saplings of maritime pine (Pinus pinaster Ait). BMC Plant Biol. 2013, 13, 95. [Google Scholar] [CrossRef]
- Nyquist, W.E.; Baker, R. Estimation of heritability and prediction of selection response in plant populations. Crit. Rev. Plant Sci. 1991, 10, 235–322. [Google Scholar] [CrossRef]
- Holland, J.B.; Nyquist, W.E.; Cervantes-Martínez, C.T. Estimating and interpreting heritability for plant breeding: An update. Plant Breed. Rev. 2003, 22, 9–112. [Google Scholar]
- Piepho, H.P.; Mohring, J. Computing heritability and selection response from unbalanced plant breeding trials. Genetics 2007, 177, 1881–1888. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wax Components | Abr. | Min. | Max. | Mean | SD |
|---|---|---|---|---|---|
| Hexacosanoic acid (C26) | C26 FA | 2.35 | 4.49 | 3 | 0.37 |
| Octacosanoic acid (C28) | C28 FA | 5.97 | 12.49 | 7.92 | 1.19 |
| Free Fatty Acids | 8.44 | 16.7 | 10.81 | 1.45 | |
| Heptacosane (C27) | C27 Alk | 2.62 | 13.5 | 5.61 | 1.45 |
| Octacosane (C28) | C28 Alk | 2.88 | 8 | 3.7 | 0.45 |
| Nonacosane (C29) | C29 Alk | 90.48 | 518.82 | 283.21 | 95.55 |
| Triacontane (C30) | C30 Alk | 6.42 | 11.7 | 8.87 | 1.01 |
| Hentriacotane (C31) | C31 Alk | 37.85 | 226.34 | 96.17 | 35.4 |
| Tritriacontane (C33) | C33 Alk | 2.57 | 5.29 | 3.28 | 0.41 |
| Alkanes | 144.88 | 636.5 | 403.8 | 114.84 | |
| Tetracosanol (C24) | C24 Alc | 4.55 | 5.9 | 5.06 | 0.25 |
| Hexacosanol (C26) | C26 Alc | 27.59 | 46.67 | 34.38 | 2.75 |
| Octacosanol (C28) | C28 Alc | 11.85 | 25.45 | 17.7 | 2.66 |
| Nonacosanol (C29) | C29 Alc | 1.33 | 3.95 | 2.26 | 0.51 |
| Triacontanol (C30) | C30 Alc | 3.81 | 10.83 | 5.35 | 0.93 |
| Primary Alcohols | 47.97 | 85.69 | 63.74 | 6.63 | |
| Nonacosan-(14)15-ol | C29 Alc-2 | 15.13 | 77.41 | 42.13 | 12.73 |
| Triacontan-14(15)-ol | C30 Alc-2 | 3.92 | 9.28 | 6 | 1.11 |
| Hentriacontan-14(15,16)-ol | C31 Alc-2 | 1.71 | 3.55 | 2.07 | 0.27 |
| Secondary Alcohols | 16.47 | 86.74 | 50.1 | 15.55 | |
| 24-methyl-hexacosanol | mC25 Alc | 0.97 | 3.18 | 1.65 | 0.42 |
| 24-methyl-hptacosanol | mC26 Alc | 11.82 | 37.32 | 21.22 | 5.16 |
| Methyl Alcohols | 12.5 | 40.41 | 22.85 | 5.69 | |
| (C43) Wax Ester | C43 WE | 7.28 | 7.94 | 7.44 | 0.09 |
| Triacontanal (C30) Aldehydes | C30 Ald | 19.91 | 51.67 | 27.42 | 4.92 |
| Nonacosane-(14),15-one Ketones | C29 Ket | 78.99 | 453.15 | 220.84 | 70.46 |
| Total Wax | 289.31 | 1356 | 808.48 | 243.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomasi, P.; Abdel-Haleem, H. Phenotypic Diversity in Leaf Cuticular Waxes in Brassica carinata Accessions. Plants 2023, 12, 3716. https://doi.org/10.3390/plants12213716
Tomasi P, Abdel-Haleem H. Phenotypic Diversity in Leaf Cuticular Waxes in Brassica carinata Accessions. Plants. 2023; 12(21):3716. https://doi.org/10.3390/plants12213716
Chicago/Turabian StyleTomasi, Pernell, and Hussein Abdel-Haleem. 2023. "Phenotypic Diversity in Leaf Cuticular Waxes in Brassica carinata Accessions" Plants 12, no. 21: 3716. https://doi.org/10.3390/plants12213716
APA StyleTomasi, P., & Abdel-Haleem, H. (2023). Phenotypic Diversity in Leaf Cuticular Waxes in Brassica carinata Accessions. Plants, 12(21), 3716. https://doi.org/10.3390/plants12213716