

Abiotic Stress in Rice: Visiting the Physiological Response and Its Tolerance Mechanisms

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Morphophysiological and Biochemical Impacts and Tolerance Mechanisms in Response to Different Abiotic Stressors
2.1. Drought Stress
2.1.1. Morphophysiological and Biochemical Responses to Drought Stress
2.1.2. Molecular Response to Drought Stress
2.2. Heat Stress
2.2.1. Morphophysiological and Biochemical Responses to Heat Stress
2.2.2. Molecular Response to Heat Stress
2.3. Cold or Low-Temperature Stress
2.3.1. Morphophysiological and Biochemical Responses to Cold or Low-Temperature Stress
2.3.2. Molecular Response to Cold or Low-Temperature Stress
2.4. Submergence Stress
2.4.1. Morphophysiological and Biochemical Responses to Submergence Stress
2.4.2. Molecular Response to Submergence Stress
2.5. Salinity Stress
2.5.1. Morphophysiological and Biochemical Responses to Salinity Stress
2.5.2. Molecular Response to Salinity Stress
2.6. Heavy Metal Stress
2.6.1. Morphological and Physiological Responses to Heavy Metals
- i.
- Arsenic
- ii.
- Cadmium
- iii.
- Lead
- iv.
- Mercury
2.6.2. Biochemical Responses to Heavy Metals
2.6.3. Molecular Responses to Heavy Metals
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yadav, G.S.; Lal, R.; Meena, R.S.; Babu, S.; Das, A.; Bhowmik, S.N.; Datta, M.; Layak, J.; Saha, P. Conservation tillage and nutrient management effects on productivity and soil carbon sequestration under double cropping of rice in north eastern region of India. Ecol. Indic. 2019, 105, 303–315. [Google Scholar] [CrossRef]
- Mahanta, K.; Bhattacharyya, P.N.; Sharma, A.K.; Rajkhowa, D.; Lesueur, D.; Verma, H.; Parit, R.; Deka, J.; Medhi, B.K.; Kohli, A. Residue and soil dissipation kinetics of chloroacetanilide herbicides on rice (Oryzae sativa L.) and assessing the impact on soil microbial parameters and enzyme activity. Environ. Monit. Assess. 2023, 195, 910. [Google Scholar] [CrossRef] [PubMed]
- Hanafiah, N.M.; Mispan, M.S.; Lim, P.E.; Baisakh, N.; Cheng, A. The 21st century agriculture: When rice research draws attention to climate variability and how weedy rice and underutilized grains come in handy. Plants 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, P.K.; Sahu, B.B. Diversity of Panicle Architecture and Traits Influencing Grain Filling. In Panicle Architecture of Rice and Its Relationship with Grain Filling; Springer International Publishing: Cham, Switzerland, 2022; pp. 107–128. ISBN 978-3-030-67897-5. [Google Scholar]
- Pickson, R.B.; He, G.; Boateng, E. Impacts of climate change on rice production: Evidence from 30 Chinese provinces. Environ. Dev. Sustain. 2022, 24, 3907–3925. [Google Scholar] [CrossRef]
- Dar, M.H.; Bano, D.A.; Waza, S.A.; Zaidi, N.W.; Majid, A.; Shikari, A.B.; Ahangar, M.A.; Hossain, M.; Kumar, A.; Singh, U.S. Abiotic Stress Tolerance-Progress and Pathways of Sustainable Rice Production. Sustainability 2021, 13, 2078. [Google Scholar] [CrossRef]
- Saud, S.; Wang, D.; Fahad, S.; Alharby, H.F.; Bamagoos, A.A.; Mjrashi, A.; Alabdallah, N.M.; AlZahrani, S.S.; AbdElgawad, H.; Adnan, M.; et al. Comprehensive Impacts of Climate Change on Rice Production and Adaptive Strategies in China. Front. Microbiol. 2022, 13, 926059. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Rivera, N.; Michel-Cuello, C.; Cárdenas-González, J.F. Green Revolution and Sustainable Development. In Encyclopedia of Sustainability in Higher Education; Leal Filho, W., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 833–850. ISBN 978-3-030-11352-0. [Google Scholar]
- Iqbal, J.; Zia-ul-Qamar; Yousaf, U.; Asgher, A.; Dilshad, R.; Qamar, F.M.; Bibi, S.; Rehman, S.U.; Haroon, M. Sustainable Rice Production Under Biotic and Abiotic Stress Challenges. In Sustainable Agriculture in the Era of the OMICs Revolution; Prakash, C.S., Fiaz, S., Nadeem, M.A., Baloch, F.S., Qayyum, A., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 241–268. ISBN 978-3-031-15568-0. [Google Scholar]
- Yang, Y.; Yu, J.; Qian, Q.; Shang, L. Enhancement of Heat and Drought Stress Tolerance in Rice by Genetic Manipulation: A Systematic Review. Rice 2022, 15, 67. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Ali, B.; Hussain, H.A. Temperature Extremes: Impact on Rice Growth and Development. In Plant Abiotic Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.; Kolapo, K. Drought Resistance in Rice from Conventional to Molecular Breeding: A Review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef]
- Fahad, S.; Adnan, M.; Hassan, S.; Saud, S.; Hussain, S.; Wu, C.; Wang, D.; Hakeem, K.R.; Alharby, H.F.; Turan, V.; et al. Chapter 10—Rice Responses and Tolerance to High Temperature; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 201–224. ISBN 978-0-12-814332-2. [Google Scholar]
- Bhattacharya, A. Effect of Low Temperature Stress on Photosynthesis and Allied Traits: A Review; Springer: Singapore, 2022; ISBN 978-981-16-9037-2. [Google Scholar]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef]
- Mahmood-ur-Rahman; Ijaz, M.; Qamar, S.; Bukhari, S.A.; Malik, K. Chapter 27—Abiotic Stress Signaling in Rice Crop; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 551–569. ISBN 978-0-12-814332-2. [Google Scholar]
- Suwanmontri, P.; Kamoshita, A.; Fukai, S. Recent changes in rice production in rainfed lowland and irrigated ecosystems in Thailand. Plant Prod. Sci. 2020, 24, 15–28. [Google Scholar] [CrossRef]
- Arif, N.; Sharma, N.C.; Yadav, V.; Ramawat, N.; Dubey, N.K.; Tripathi, D.K.; Chauhan, D.K.; Sahi, S. Understanding Heavy Metal Stress in a Rice Crop: Toxicity, Tolerance Mechanisms, and Amelioration Strategies. J. Plant Biol. 2019, 62, 239–253. [Google Scholar] [CrossRef]
- Melo, F.V.; Oliveira, M.M.; Saibo, N.J.M.; Lourenço, T.F. Modulation of Abiotic Stress Responses in Rice by E3-Ubiquitin Ligases: A Promising Way to Develop Stress-Tolerant Crops. Front. Plant Sci. 2021, 12, 640193. [Google Scholar] [CrossRef]
- Das, G.; Patra, J.K.; Baek, K.H. Insight into MAS: A Molecular Tool for Development of Stress Resistant and Quality of Rice through Gene Stacking. Front. Plant Sci. 2017, 8, 985. [Google Scholar] [CrossRef]
- Pandey, V.; Shukla, A. Acclimation and Tolerance Strategies of Rice under Drought Stress. Rice Sci. 2015, 22, 147–161. [Google Scholar] [CrossRef]
- Miyan, M.A. Droughts in asian least developed countries: Vulnerability and sustainability. Weather Clim. Extrem. 2015, 7, 8–23. [Google Scholar] [CrossRef]
- Yang, X.; Wang, B.; Chen, L.; Li, P.; Cao, C. The Different Influences of Drought Stress at the Flowering Stage on Rice Physiological Traits, Grain Yield, and Quality. Sci. Rep. 2019, 9, 3742. [Google Scholar] [CrossRef]
- Venuprasad, R.; Lafitte, H.R.; Atlin, G.N. Response to direct selection for grain yield under drought stress in rice. Crop Sci. 2007, 47, 285–293. [Google Scholar] [CrossRef]
- Lafitte, H.; Ismail, A.; Bennett, J. Abiotic Stress Tolerance in Rice for Asia: Progress and the Future. In Proceedings of the 4th International Crop Science Congress, Brisbane, Australia, 26 September–1 October 2004. [Google Scholar]
- Upadhyaya, H.; Panda, S.K. Chapter 9—Drought Stress Responses and Its Management in Rice; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J.K., Eds.; Woodhead Publishing: Sawston, UK, 2019; ISBN 978-0-12-814332-2. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Zampieri, E.; Pesenti, M.; Nocito, F.F.; Sacchi, G.A.; Valè, G. Rice Responses to Water Limiting Conditions: Improving Stress Management by Exploiting Genetics and Physiological Processes. Agriculture 2023, 13, 464. [Google Scholar] [CrossRef]
- Fukai, S.; Cooper, M. Development of drought-resistant cultivars using physiomorphological traits in rice. Field Crop. Res. 1995, 40, 67–86. [Google Scholar] [CrossRef]
- Moonmoon, S.; Islam, M. Effect of Drought Stress at Different Growth Stages on Yield and Yield Components of Six Rice (Oryza sativa L.) Genotypes. Fundam. Appl. Agric. 2017, 2, 285–289. [Google Scholar] [CrossRef]
- Ranjan, A.; Sinha, R.; Singla-Pareek, S.L.; Pareek, A.; Singh, A.K. Shaping the root system architecture in plants for adaptation to drought stress. Physiol. Plant. 2022, 174, e13651. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.-H.K.-H. Root Response to Drought Stress in Rice (Oryza Sativa L.). Int. J. Mol. Sci 2020, 21, 1513. [Google Scholar] [CrossRef] [PubMed]
- Champoux, M.C.; Wang, G.; Sarkarung, S.; Mackill, D.J.; O’Toole, J.C.; Huang, N.; McCouch, S.R. Locating genes associated with root morphology and drought avoidance in rice via linkage to molecular markers. Theor. Appl. Genet. 1995, 90, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Bañoc, D.M.; Yamauchi, A.; Kamoshita, A.; Wade, L.J.; Pardales, J.R. Dry Matter Production and Root System Development of Rice Cultivars under Fluctuating Soil Moisture. Plant Prod. Sci. 2000, 3, 197–207. [Google Scholar] [CrossRef]
- Bañoc, D.M.; Yamauchi, A.; Kamoshita, A.; Wade, L.J.; Pardales, J.R. Genotypic Variations in Response of Lateral Root Development to Fluctuating Soil Moisture in Rice. Plant Prod. Sci. 2000, 3, 335–343. [Google Scholar] [CrossRef]
- Henry, A.; Cal, A.J.; Batoto, T.C.; Torres, R.O.; Serraj, R. Root attributes affecting water uptake of rice (Oryza sativa) under drought. J. Exp. Bot. 2012, 63, 4751–4763. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Farooq, M.; Kobayashi, N.; Ito, O.; Wahid, A.; Serraj, R. Broader leaves result in better performance of indica rice under drought stress. J. Plant Physiol. 2010, 167, 1066–1075. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Sangam, S.; Amrutha, R.N.; Sri Laxmi, P.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Zhu, R.; Wu, F.; Zhou, S.; Hu, T.; Huang, J.; Gao, Y. Cumulative effects of drought–flood abrupt alternation on the photosynthetic characteristics of rice. Environ. Exp. Bot. 2020, 169, 103901. [Google Scholar] [CrossRef]
- Mishra, S.S.; Behera, P.K.; Kumar, V.; Lenka, S.K.; Panda, D. Physiological characterization and allelic diversity of selected drought tolerant traditional rice (Oryza sativa L.) landraces of Koraput, India. Physiol. Mol. Biol. Plants 2018, 24, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Kobayashi, N.; Wahid, A.; Ito, O.; Basra, S.M.A. Strategies for Producing More Rice with Less Water. In Advances in Agronomy; Elsevier: San Diego, CA, USA, 2009; Volume 101, p. e1. [Google Scholar] [CrossRef]
- Panda, D.; Mishra, S.S.; Behera, P.K. Drought Tolerance in Rice: Focus on Recent Mechanisms and Approaches. Rice Sci. 2021, 28, 119–132. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Zhang, F.-J.; Zhang, K.-K.; Du, C.-Z.; Li, J.; Xing, Y.-X.; Yang, L.-T.; Li, Y.-R. Effect of Drought Stress on Anatomical Structure and Chloroplast Ultrastructure in Leaves of Sugarcane. Sugar Tech 2015, 17, 41–48. [Google Scholar] [CrossRef]
- Ayyaz, A.; Fang, R.; Ma, J.; Hannan, F.; Huang, Q.; Athar, H.-R.; Sun, Y.; Javed, M.; Ali, S.; Zhou, W.; et al. Calcium nanoparticles (Ca-NPs) improve drought stress tolerance in Brassica napus by modulating the photosystem II, nutrient acquisition and antioxidant performance. NanoImpact 2022, 28, 100423. [Google Scholar] [CrossRef]
- Zahra, N.; Hafeez, M.B.; Kausar, A.; Al Zeidi, M.; Asekova, S.; Siddique, K.H.M.M.; Farooq, M.; Al Zeidi, M.; Asekova, S.; Siddique, K.H.M.M.; et al. Plant photosynthetic responses under drought stress: Effects and management. J. Agron. Crop Sci. 2023, 209, 651–672. [Google Scholar] [CrossRef]
- Lum, M.S.; Hanafi, M.M.; Rafii, Y.M.; Akmar, A.S.N. Effect of drought stress on growth, proline and antioxidant enzyme activities of upland rice. J. Anim. Plant Sci. 2014, 24, 1487–1493. [Google Scholar]
- Schlicke, H.; Hartwig, A.S.; Firtzlaff, V.; Richter, A.S.; Glässer, C.; Maier, K.; Finkemeier, I.; Grimm, B. Induced Deactivation of Genes Encoding Chlorophyll Biosynthesis Enzymes Disentangles Tetrapyrrole-Mediated Retrograde Signaling. Mol. Plant 2014, 7, 1211–1227. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.W.U.; Montgomery, B.L. Interdependence of tetrapyrrole metabolism, the generation of oxidative stress and the mitigative oxidative stress response. Redox Biol. 2015, 4, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Hossain, M.A.; da Silva, J.A.T.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense Is a Key Factor. In Crop Stress and its Management: Perspectives and Strategies; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. [Google Scholar] [CrossRef]
- Asada, K. The Water-Water Cycle In Chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential Activity of the Antioxidant Defence System and Alterations in the Accumulation of Osmolyte and Reactive Oxygen Species under Drought Stress and Recovery in Rice (Oryza Sativa L.) Tillering. Sci. Rep. 2019, 9, 8543. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; Dey, N. Redox metabolic and molecular parameters for screening drought tolerant indigenous aromatic rice cultivars. Physiol. Mol. Biol. Plants 2018, 24, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.; Bhattacharjee, S. Accumulation of Polyphenolic Compounds and Osmolytes under Dehydration Stress and Their Implication in Redox Regulation in Four Indigenous Aromatic Rice Cultivars. Rice Sci. 2020, 27, 329–344. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Capell, T.; Bassie, L.; Christou, P. Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress. Proc. Natl. Acad. Sci. USA 2004, 101, 9909–9914. [Google Scholar] [CrossRef]
- Kuru, İ.; Işıkalan, Ç.; Akbaş, F. Physiological and Biochemical Responses of Rice (Oryza Sativa L.) Varieties Against Drought Stress. Bangladesh J. Bot. 2021, 50, 335–342. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Hoang, X.L.T.; Nhi, D.N.H.; Thu, N.B.A.; Thao, N.P.; Tran, L.-S.P. Transcription Factors and Their Roles in Signal Transduction in Plants under Abiotic Stresses. Curr. Genom. 2017, 18, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.-P.; Chao, D.-Y.; Lin, H.-X. Toward understanding molecular mechanisms of abiotic stress responses in rice. Rice 2008, 1, 36–51. [Google Scholar] [CrossRef]
- Maruyama, K.; Todaka, D.; Mizoi, J.; Yoshida, T.; Kidokoro, S.; Matsukura, S.; Takasaki, H.; Sakurai, T.; Yamamoto, Y.Y.; Yoshiwara, K.; et al. Identification of Cis-Acting Promoter Elements in Cold- and Dehydration-Induced Transcriptional Pathways in Arabidopsis, Rice, and Soybean. DNA Res. 2012, 19, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Kashtoh, H.; Wang, Q.; Zhen, G.; Li, Q.; Tang, L.; Gao, H.; Zhang, C.; Qin, L.; Su, M.; et al. Structure and activity of SLAC1 channels for stomatal signaling in leaves. Proc. Natl. Acad. Sci. USA 2021, 118, e2015151118. [Google Scholar] [CrossRef] [PubMed]
- Kashtoh, H.; Baek, K.-H. Structural and Functional Insights into the Role of Guard Cell Ion Channels in Abiotic Stress-Induced Stomatal Closure. Plants 2021, 10, 2774. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C Phosphatase Activity Function as Abscisic Acid Sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef]
- Park, S.-Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.F.; et al. Abscisic Acid Inhibits Type 2C Protein Phosphatases via the PYR/PYL Family of START Proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef]
- Kim, H.; Lee, K.; Hwang, H.; Bhatnagar, N.; Kim, D.-Y.; Yoon, I.S.; Byun, M.-O.; Kim, S.T.; Jung, K.-H.; Kim, B.-G. Overexpression of PYL5 in rice enhances drought tolerance, inhibits growth, and modulates gene expression. J. Exp. Bot. 2014, 65, 453–464. [Google Scholar] [CrossRef]
- Matsukura, S.; Mizoi, J.; Yoshida, T.; Todaka, D.; Ito, Y.; Maruyama, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol. Genet. Genom. 2010, 283, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crop. Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.; Ramanathan, V.; Nallathambi, J.; Duraialagaraja, S.; Muthurajan, R. Over-expression of a NAC 67 transcription factor from finger millet (Eleusine coracana L.) confers tolerance against salinity and drought stress in rice. BMC Biotechnol. 2016, 16, 35. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Li, X.; Hicks, L.M.; Xiong, L. A raf-like MAPKKK gene DSM1 mediates drought resistance through reactive oxygen species scavenging in rice. Plant Physiol. 2010, 152, 876–890. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium Supplementation Improves Na+/K+ Ratio, Antioxidant Defense and Glyoxalase Systems in Salt-Stressed Rice Seedlings. Front. Plant Sci. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, Y.S.; Redillas, M.C.F.R.; Jang, G.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.H.; Reuzeau, C.; Kim, J.K. OsNAC5 overexpression enlarges root diameter in rice plants leading to enhanced drought tolerance and increased grain yield in the field. Plant Biotechnol. J. 2013, 11, 101–114. [Google Scholar] [CrossRef]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef]
- Xiao, B.; Huang, Y.; Tang, N.; Xiong, L. Over-expression of a LEA gene in rice improves drought resistance under the field conditions. Theor. Appl. Genet. 2007, 115, 35–46. [Google Scholar] [CrossRef]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a Key Player of the Basic Leucine Zipper Transcription Factor Family for Conferring Abscisic Acid Sensitivity and Salinity and Drought Tolerance in Rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef]
- Lu, G.; Gao, C.; Zheng, X.; Han, B. Identification of OsbZIP72 as a positive regulator of ABA response and drought tolerance in rice. Planta 2009, 229, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Kim, Y.S.; Kwon, C.W.; Park, H.K.; Jeong, J.S.; Kim, J.K. Overexpression of the Transcription Factor AP37 in Rice Improves Grain Yield under Drought Conditions. Plant Physiol. 2009, 150, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.H.; Choi, Y.D.Y.D.; Kim, M.; Reuzeau, C.; Kim, J.K. Root-Specific Expression of OsNAC10 Improves Drought Tolerance and Grain Yield in Rice under Field Drought Conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, X.; Wang, Z.; Wang, S.; Wang, Y.; Zhu, Q.; Li, S.; Xiang, C. Arabidopsis Enhanced Drought Tolerance1/HOMEODOMAIN GLABROUS11 Confers Drought Tolerance in Transgenic Rice without Yield Penalty. Plant Physiol. 2013, 162, 1378–1391. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, G.; Manimaran, P.; Voleti, S.R.; Subrahmanyam, D.; Sundaram, R.M.; Bansal, K.C.; Viraktamath, B.C.; Balachandran, S.M. Stress-inducible expression of AtDREB1A transcription factor greatly improves drought stress tolerance in transgenic indica rice. Transgenic Res. 2014, 23, 421–439. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Hu, W.; Deng, X.; Zhang, Y.; Liu, X.; Zhao, X.; Luo, Q.; Jin, Z.; Li, Y.; Zhou, S.; et al. A Rice Calcium-Dependent Protein Kinase OsCPK9 Positively Regulates Drought Stress Tolerance and Spikelet Fertility. BMC Plant Biol. 2014, 14, 133. [Google Scholar] [CrossRef]
- You, J.; Hu, H.; Xiong, L. An ornithine δ-aminotransferase gene OsOAT confers drought and oxidative stress tolerance in rice. Plant Sci. 2012, 197, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-W.; Zang, B.-S.; Deng, X.-W.; Wang, X.-P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef]
- Zhu, B.; Su, J.; Chang, M.; Verma, D.P.S.; Fan, Y.-L.; Wu, R. Overexpression of a Δ1-pyrroline-5-carboxylate synthetase gene and analysis of tolerance to water- and salt-stress in transgenic rice. Plant Sci. 1998, 139, 41–48. [Google Scholar] [CrossRef]
- Chandra Babu, R.; Zhang, J.; Blum, A.; David Ho, T.-H.; Wu, R.; Nguyen, H. HVA1, a LEA gene from barley confers dehydration tolerance in transgenic rice (Oryza sativa L.) via cell membrane protection. Plant Sci. 2004, 166, 855–862. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, S.; Li, W.; Feng, W.; Li, J.; Wu, Z.; Gao, X.; Liu, F.; Shao, M. Overexpression of a Harpin-encoding gene hrf1 in rice enhances drought tolerance. J. Exp. Bot. 2011, 62, 4229–4238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, F.; Li, D.; Zhang, H.; Huang, R. Expression of ethylene response factor JERF1 in rice improves tolerance to drought. Planta 2010, 232, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.; Kim, S.K.; Cho, S.K.; Kang, B.G.; Kim, W.T. Overexpression of OsRDCP1, a rice RING domain-containing E3 ubiquitin ligase, increased tolerance to drought stress in rice (Oryza sativa L.). Plant Sci. 2011, 180, 775–782. [Google Scholar] [CrossRef]
- Gao, T.; Wu, Y.; Zhang, Y.; Liu, L.; Ning, Y.; Wang, D.; Tong, H.; Chen, S.; Chu, C.; Xie, Q. OsSDIR1 overexpression greatly improves drought tolerance in transgenic rice. Plant Mol. Biol. 2011, 76, 145–156. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Zong, W.; Li, X.; Ning, J.; Hu, H.; Li, X.; Xiao, J.; Xiong, L. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 2013, 64, 569–583. [Google Scholar] [CrossRef] [PubMed]
- Samota, M.K.; Sasi, M.; Awana, M.; Yadav, O.P.; Amitha Mithra, S.V.; Tyagi, A.; Kumar, S.; Singh, A. Elicitor-Induced Biochemical and Molecular Manifestations to Improve Drought Tolerance in Rice (Oryza Sativa L.) through Seed-Priming. Front. Plant Sci. 2017, 8, 934. [Google Scholar] [CrossRef]
- Pandey, V.; Ansari, M.W.; Tula, S.; Yadav, S.; Sahoo, R.K.; Shukla, N.; Bains, G.; Badal, S.; Chandra, S.; Gaur, A.K.; et al. Dose-dependent response of Trichoderma harzianum in improving drought tolerance in rice genotypes. Planta 2016, 243, 1251–1264. [Google Scholar] [CrossRef]
- Wu, C.; Cui, K.; Wang, W.; Li, Q.; Fahad, S.; Hu, Q.; Huang, J.; Nie, L.; Peng, S. Heat-Induced Phytohormone Changes Are Associated with Disrupted Early Reproductive Development and Reduced Yield in Rice. Sci. Rep. 2016, 6, 34978. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Kilasi, N.L.; Singh, J.; Vallejos, C.E.; Ye, C.; Jagadish, S.V.K.; Kusolwa, P.; Rathinasabapathi, B. Heat Stress Tolerance in Rice (Oryza Sativa L.): Identification of Quantitative Trait Loci and Candidate Genes for Seedling Growth Under Heat Stress. Front. Plant Sci. 2018, 9, 1578. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Zhou, J.; Hu, S.; Chen, H.; Xiang, J.; Zhang, Y.Y.Y.; Zeng, Y.; Shi, Q.; Zhu, D.; et al. Research Progress on Heat Stress of Rice at Flowering Stage. Rice Sci. 2019, 26, 1–10. [Google Scholar] [CrossRef]
- Ishimaru, T.; Hirabayashi, H.; Kuwagata, T.; Ogawa, T.; Kondo, M. The Early-Morning Flowering Trait of Rice Reduces Spikelet Sterility under Windy and Elevated Temperature Conditions at Anthesis. Plant Prod. Sci. 2012, 15, 19–22. [Google Scholar] [CrossRef]
- Fahad, S.; Ihsan, M.Z.; Khaliq, A.; Daur, I.; Saud, S.; Alzamanan, S.; Nasim, W.; Abdullah, M.; Khan, I.A.; Wu, C.; et al. Consequences of high temperature under changing climate optima for rice pollen characteristics-concepts and perspectives. Arch. Agron. Soil Sci. 2018, 64, 1473–1488. [Google Scholar] [CrossRef]
- Ranathunge, K.; Steudle, E.; Lafitte, R. Control of water uptake by rice (Oryza sativa L.): Role of the outer part of the root. Planta 2003, 217, 193–205. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Ashraf, M.Y.; Khaliq, B.; Sun, M.; Hussain, S.; Gao, Z.; Noor, H.; Alam, S. Mechanisms and Adaptation Strategies to Improve Heat Tolerance in Rice. A Review. Plants 2019, 8, 508. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Bao, J.; Gao, Z.; Sun, D.; Zheng, S.; Bai, J. How rice adapts to high temperatures. Front. Plant Sci. 2023, 14, 1137923. [Google Scholar] [CrossRef] [PubMed]
- González-García, M.P.; Conesa, C.M.; Lozano-Enguita, A.; Baca-González, V.; Simancas, B.; Navarro-Neila, S.; Sánchez-Bermúdez, M.; Salas-González, I.; Caro, E.; Castrillo, G.; et al. Temperature changes in the root ecosystem affect plant functionality. Plant Commun. 2023, 4, 100514. [Google Scholar] [CrossRef] [PubMed]
- Arai-Sanoh, Y.; Ishimaru, T.; Ohsumi, A.; Kondo, M. Effects of Soil Temperature on Growth and Root Function in Rice. Plant Prod. Sci. 2010, 13, 235–242. [Google Scholar] [CrossRef]
- Yamakawa, Y.; Kishikawa, H. On the Effect of Temperature upon the Division and Elongation of Cells in the Root of Rice Plant. Jpn. J. Crop Sci. 1957, 26, 94–95. [Google Scholar] [CrossRef]
- Liu, J.; Hasanuzzaman, M.; Wen, H.; Zhang, J.; Peng, T.; Sun, H.; Zhao, Q. High temperature and drought stress cause abscisic acid and reactive oxygen species accumulation and suppress seed germination growth in rice. Protoplasma 2019, 256, 1217–1227. [Google Scholar] [CrossRef]
- Liu, J.; Sun, X.; Xu, F.; Zhang, Y.; Zhang, Q.; Miao, R.; Zhang, J.; Liang, J.; Xu, W. Suppression of OsMDHAR4 Enhances Heat Tolerance by Mediating H2O2-Induced Stomatal Closure in Rice Plants. Rice 2018, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, T.N.; Ghildyal, B.P. Germination Response of Rice Seeds to Constant and Alternating Temperatures. Agron. J. 1969, 61, 328–330. [Google Scholar] [CrossRef]
- Baker, J.T. Yield responses of southern US rice cultivars to CO2 and temperature. Agric. For. Meteorol. 2004, 122, 129–137. [Google Scholar] [CrossRef]
- Sandhu, J.; Irvin, L.; Liu, K.; Staswick, P.; Zhang, C.; Walia, H. Endoplasmic Reticulum Stress Pathway Mediates the Early Heat Stress Response of Developing Rice Seeds. Plant. Cell Environ. 2021, 44, 2604–2624. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.V.K.; Murty, M.V.R.; Quick, W.P. Rice Responses to Rising Temperatures--Challenges, Perspectives and Future Directions. Plant. Cell Environ. 2015, 38, 1686–1698. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.S.; Farooq, M.; Asch, F.; Krishna, J.S.V.; Prasad, P.V.V.; Siddique, K.H.M. Thermal stress impacts reproductive development and grain yield in rice. Plant Physiol. Biochem. 2017, 115, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Soda, N.; Gupta, B.K.; Anwar, K.; Sharan, A.; Govindjee, S.-P.L.S.; Pareek, A.; Govindjee; Singla-Pareek, S.L.; Pareek, A. Rice Intermediate Filament, OsIF, Stabilizes Photosynthetic Machinery and Yield under Salinity and Heat Stress. Sci. Rep. 2018, 8, 4072. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Wang, H.H.; Shao, H.; Tang, X. Recent Advances in Utilizing Transcription Factors to Improve Plant Abiotic Stress Tolerance by Transgenic Technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef]
- Miyahara, K.; Wada, T.; Sonoda, J.-Y.; Tsukaguchi, T.; Miyazaki, M.; Tsubone, M.; Yamaguchi, O.; Ishibashi, M.; Iwasawa, N.; Umemoto, T.; et al. Detection and validation of QTLs for milky-white grains caused by high temperature during the ripening period in Japonica rice. Breed. Sci. 2017, 67, 333–339. [Google Scholar] [CrossRef]
- Kaneko, K.; Sasaki, M.; Kuribayashi, N.; Suzuki, H.; Sasuga, Y.; Shiraya, T.; Inomata, T.; Itoh, K.; Baslam, M.; Mitsui, T. Proteomic and Glycomic Characterization of Rice Chalky Grains Produced Under Moderate and High-Temperature Conditions in Field System. Rice 2016, 9, 26. [Google Scholar] [CrossRef]
- Nevame, A.Y.M.; Emon, R.M.; Malek, M.A.; Hasan, M.M.; Alam, M.A.; Muharam, F.M.; Aslani, F.; Rafii, M.Y.; Ismail, M.R. Relationship between High Temperature and Formation of Chalkiness and Their Effects on Quality of Rice. Biomed Res. Int. 2018, 1653721, 1653721. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Miyahara, K.; Sonoda, J.-Y.; Tsukaguchi, T.; Miyazaki, M.; Tsubone, M.; Ando, T.; Ebana, K.; Yamamoto, T.; Iwasawa, N.; et al. Detection of QTLs for white-back and basal-white grains caused by high temperature during ripening period in japonica rice. Breed. Sci. 2015, 65, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.S.; Yang, C.Y. Ethylene-Mediated Signaling Confers Thermotolerance and Regulates Transcript Levels of Heat Shock Factors in Rice Seedlings under Heat Stress. Bot. Stud. 2019, 60, 23. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, H.; Wang, J.; Wang, Y.; Pan, S.; Tian, H.; Duan, M.; Wang, S.; Tang, X.; Mo, Z. Responses of Plant Growth, Physiological, Gas Exchange Parameters of Super and Non-Super Rice to Rhizosphere Temperature at the Tillering Stage. Sci. Rep. 2019, 9, 10618. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-M.; Chao, D.-Y.; Wu, Y.; Huang, X.; Chen, K.; Cui, L.-G.; Su, L.; Ye, W.-W.; Chen, H.; Chen, H.-C.; et al. Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice. Nat. Genet. 2015, 47, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.; Canales, E.; Borras-Hidalgo, O. Molecular aspects of abiotic stress in plants. Biotecnol. Apl. 2005, 22, 1–10. [Google Scholar]
- Djanaguiraman, M.; Annie Sheeba, J.; Durga Devi, D.; Bangarusamy, U. Cotton Leaf Senescence Can Be Delayed by Nitrophenolate Spray through Enhanced Antioxidant Defence System. J. Agron. Crop Sci. 2009, 195, 213–224. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhou, L.; Liu, J.; Du, X.; Asad, M.-A.-U.; Huang, F.; Pan, G.; Cheng, F. Relationship of ROS accumulation and superoxide dismutase isozymes in developing anther with floret fertility of rice under heat stress. Plant Physiol. Biochem. PPB 2018, 122, 90–101. [Google Scholar] [CrossRef]
- Awasthi, R.; Bhandari, K.; Nayyar, H. Temperature stress and redox homeostasis in agricultural crops. Front. Environ. Sci. 2015, 3, 11. [Google Scholar] [CrossRef]
- Soengas, P.; Rodríguez, V.M.; Velasco, P.; Cartea, M.E. Effect of Temperature Stress on Antioxidant Defenses in Brassica Oleracea. ACS Omega 2018, 3, 5237–5243. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Liu, C.; Chen, X. Proteomics of rice in response to heat stress and advances in genetic engineering for heat tolerance in rice. Plant Cell Rep. 2011, 30, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-H.; Won, S.-H.; Lee, H.-S.; Miyao, M.; Chung, W.-I.; Kim, I.-J.; Jo, J. Expression of the chloroplast-localized small heat shock protein by oxidative stress in rice. Gene 2000, 245, 283–290. [Google Scholar] [CrossRef]
- Wu, X.; Shiroto, Y.; Kishitani, S.; Ito, Y.; Toriyama, K. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 2009, 28, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Liao, K.; Du, H.; Xu, Y.; Song, H.; Li, X.; Xiong, L. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2015, 66, 6803–6817. [Google Scholar] [CrossRef] [PubMed]
- El-kereamy, A.; Bi, Y.M.; Ranathunge, K.; Beatty, P.H.; Good, A.G.; Rothstein, S.J. The Rice R2R3-MYB Transcription Factor OsMYB55 Is Involved in the Tolerance to High Temperature and Modulates Amino Acid Metabolism. PLoS ONE 2012, 7, 52030. [Google Scholar] [CrossRef]
- Ambavaram, M.M.R.; Basu, S.; Krishnan, A.; Ramegowda, V.; Batlang, U.; Rahman, L.; Baisakh, N.; Pereira, A. Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress. Nat. Commun. 2014, 5, 5302. [Google Scholar] [CrossRef]
- Lim, S.D.; Cho, H.Y.; Park, Y.C.; Ham, D.J.; Lee, J.K.; Jang, C.S. The rice RING finger E3 ligase, OsHCI1, drives nuclear export of multiple substrate proteins and its heterogeneous overexpression enhances acquired thermotolerance. J. Exp. Bot. 2013, 64, 2899–2914. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, S.; Zhang, Y.; Xu, T.; Guo, F.; Tang, H.; Li, X.; Wang, P.; Qian, W.; Xue, Y. A High Temperature-Dependent Mitochondrial Lipase EXTRA GLUME1 Promotes Floral Phenotypic Robustness against Temperature Fluctuation in Rice (Oryza sativa L.). PLOS Genet. 2016, 12, e1006152. [Google Scholar] [CrossRef]
- Hossain, M.A.; Cho, J.I.; Han, M.; Ahn, C.H.; Jeon, J.S.; An, G.; Park, P.B. The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J. Plant Physiol. 2010, 167, 1512–1520. [Google Scholar] [CrossRef]
- Liu, J.G.; Qin, Q.-L.; Zhang, Z.; Peng, R.H.; Xiong, A.S.; Chen, J.M.; Yao, Q.H. OsHSF7 Gene in Rice, Oryza Sativa L., Encodes a Transcription Factor That Functions as a High Temperature Receptive and Responsive Factor. BMB Rep. 2009, 42, 16–21. [Google Scholar] [CrossRef]
- Lin, M.Y.; Chai, K.H.; Ko, S.S.; Kuang, L.Y.; Lur, H.S.; Charng, Y.Y. A Positive Feedback Loop between Heat Shock Protein101 And Heat Stress-Associated 32-Kd Protein Modulates Long-Term Acquired Thermotolerance Illustrating Diverse Heat Stress Responses in Rice Varieties. Plant Physiol. 2014, 164, 2045–2053. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, C.; Wei, C.; Liu, X.; Wang, M.; Yu, F.; Xie, Q.; Tu, J. The RING Finger Ubiquitin E3 Ligase OsHTAS Enhances Heat Tolerance by Promoting H2O2-Induced Stomatal Closure in Rice. Plant Physiol. 2016, 170, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Zhao, J.; Lin, D.; Chen, J.; Xu, J.; Zhou, H.; Teng, S.; Dong, Y. The Rice TCM5 Gene Encoding a Novel Deg Protease Protein Is Essential for Chloroplast Development under High Temperatures. Rice 2016, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Qin, B.; Li, X.; Tang, D.; Zhang, Y.; Cheng, Z.; Xue, Y. Nucleolar DEAD-Box RNA Helicase TOGR1 Regulates Thermotolerant Growth as a Pre-rRNA Chaperone in Rice. PLoS Genet. 2016, 12, e1005844. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Liu, H.; Cui, Y.; Yu, H.; Rao, Y.; Yan, Y.; Zeng, D.; Hu, J.; Zhang, G.; Gao, Z.; et al. UDP-N-Acetylglucosamine Pyrophosphorylase Enhances Rice Survival at High Temperature. New Phytol. 2022, 233, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Guo, T.; Li, X.M.; Zhang, Y.M.; Yang, Y.B.; Ye, W.W.; Dong, N.Q.; Shi, C.L.; Kan, Y.; Xiang, Y.H.; et al. Translational Regulation of Plant Response to High Temperature by a Dual-Function TRNAHis Guanylyltransferase in Rice. Mol. Plant 2019, 12, 1123–1142. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lyu, Y.; Yang, W.; Yang, Z.; Lu, S.; Liu, J. A membrane-associated NAC transcription factor OsNTL3 is involved in thermotolerance in rice. Plant Biotechnol. J. 2020, 18, 1317–1329. [Google Scholar] [CrossRef]
- Singh, A.; Mittal, D.; Lavania, D.; Agarwal, M.; Mishra, R.C.; Grover, A. OsHsfA2c and OsHsfB4b are involved in the transcriptional regulation of cytoplasmic OsClpB (Hsp100) gene in rice (Oryza sativa L.). Cell Stress Chaperones 2012, 17, 243–254. [Google Scholar] [CrossRef]
- Tang, Y.; Gao, C.-C.; Gao, Y.; Yang, Y.; Shi, B.; Yu, J.-L.; Lyu, C.; Sun, B.-F.; Wang, H.-L.; Xu, Y.; et al. OsNSUN2-Mediated 5-Methylcytosine mRNA Modification Enhances Rice Adaptation to High Temperature. Dev. Cell 2020, 53, 272–286.e7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, J.-F.; Kan, Y.; Shan, J.-X.; Ye, W.-W.; Dong, N.-Q.; Guo, T.; Xiang, Y.-H.; Yang, Y.-B.; Li, Y.-C.; et al. A genetic module at one locus in rice protects chloroplasts to enhance thermotolerance. Science 2022, 376, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Qiao, B.; Zhang, Q.; Liu, D.; Wang, H.; Yin, J.; Wang, R.; He, M.; Cui, M.; Shang, Z.; Wang, D.; et al. A calcium-binding protein, rice annexin OsANN1, enhances heat stress tolerance by modulating the production of H2O2. J. Exp. Bot. 2015, 66, 5853–5866. [Google Scholar] [CrossRef] [PubMed]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, I. The Role of Sugars in Plant Responses to Stress and Their Regulatory Function during Development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Araki, M.; Okamura, K.; Ishibashi, Y.; Yuasa, T.; Iwaya-Inoue, M. Assimilate translocation and expression of sucrose transporter, OsSUT1, contribute to high-performance ripening under heat stress in the heat-tolerant rice cultivar Genkitsukushi. J. Plant Physiol. 2013, 170, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Savada, R.P.; Ozga, J.A.; Jayasinghege, C.P.A.; Waduthanthri, K.D.; Reinecke, D.M. Heat stress differentially modifies ethylene biosynthesis and signaling in pea floral and fruit tissues. Plant Mol. Biol. 2017, 95, 313–331. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Páez, S.E.; Cueto-Niño, Y.A.; Sánchez-Reinoso, A.D.; Garces-Varon, G.; Chávez-Arias, C.C.; Restrepo-Díaz, H. Foliar boron compounds applications mitigate heat stress caused by high daytime temperatures in rice (Oryza sativa L.) Boron mitigates heat stress in rice. J. Plant Nutr. 2021, 44, 2514–2527. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Z.; Chong, K.; Xu, Y. Chilling tolerance in rice: Past and present. J. Plant Physiol. 2022, 268, 153576. [Google Scholar] [CrossRef]
- Bhattacharya, A. Effect of Low-Temperature Stress on Germination, Growth, and Phenology of Plants: A Review. In Physiological Processes in Plants Under Low Temperature Stress; Springer: Singapore, 2022; pp. 1–106. [Google Scholar] [CrossRef]
- Hsu, C.H.; Hsu, Y.T. Biochemical Responses of Rice Roots to Cold Stress. Bot. Stud. 2019, 60, 14. [Google Scholar] [CrossRef]
- Reddy, K.R.; Seghal, A.; Jumaa, S.; Bheemanahalli, R.; Kakar, N.; Redoña, E.D.; Wijewardana, C.; Alsajri, F.A.; Chastain, D.; Gao, W.; et al. Morpho-Physiological Characterization of Diverse Rice Genotypes for Seedling Stage High- and Low-Temperature Tolerance. Agronomy 2021, 11, 112. [Google Scholar] [CrossRef]
- Freitas, G.M.; Thomas, J.; Liyanage, R.; Lay, J.O.; Basu, S.; Ramegowda, V.; Amaral, M.N.; Benitez, L.C.; Bolacel Braga, E.J.; Pereira, A.; et al. Cold tolerance response mechanisms revealed through comparative analysis of gene and protein expression in multiple rice genotypes. PLoS ONE 2019, 14, e0218019. [Google Scholar] [CrossRef]
- Baruah, A.R.; Ishigo-Oka, N.; Adachi, M.; Oguma, Y.; Tokizono, Y.; Onishi, K.; Sano, Y. Cold tolerance at the early growth stage in wild and cultivated rice. Euphytica 2009, 165, 459–470. [Google Scholar] [CrossRef]
- González-Schain, N.; Roig-Villanova, I.; Kater, M.M. Early Cold Stress Responses in Post-Meiotic Anthers from Tolerant and Sensitive Rice Cultivars. Rice 2019, 12, 94. [Google Scholar] [CrossRef] [PubMed]
- Nurhasanah Ritonga, F.; Chen, S. Physiological and Molecular Mechanism Involved in Cold Stress Tolerance in Plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Song, S.; Xiao, Y.; Fan, F.; Zhou, Y.; Jia, G.; Tang, W.; Peng, J. Circadian Clock-Coordinated Response to Chilling Stress in Rice. Environ. Exp. Bot. 2021, 185, 104398. [Google Scholar] [CrossRef]
- Shimono, H.; Fujimura, S.; Nishimura, T.; Hasegawa, T. Nitrogen Uptake by Rice (Oryza sativa L.) Exposed to Low Water Temperatures at Different Growth Stages. J. Agron. Crop Sci. 2012, 198, 145–151. [Google Scholar] [CrossRef]
- Feng, H.; Yan, M.; Fan, X.; Li, B.; Shen, Q.; Miller, A.J.; Xu, G. Spatial expression and regulation of rice high-affinity nitrate transporters by nitrogen and carbon status. J. Exp. Bot. 2011, 62, 2319–2332. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Yuan, Y.; Zhang, C.; Ou, J.; Zhou, W.; Lin, Q. Modulation of key enzymes involved in ammonium assimilation and carbon metabolism by low temperature in rice (Oryza sativa L.) roots. Plant Sci. 2005, 169, 295–302. [Google Scholar] [CrossRef]
- Guy, C.L. Cold Acclimation and Freezing Stress Tolerance: Role of Protein Metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 187–223. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant Cold Acclimation: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef]
- Janmohammadi, M.; Zolla, L.; Rinalducci, S. Low temperature tolerance in plants: Changes at the protein level. Phytochemistry 2015, 117, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Dikilitas, M.; Karakas, S.; Simsek, E.; Yadav, A.N. Microbes from cold deserts and their applications in mitigation of cold stress in plants. In Microbiomes of Extreme Environments; CRC Press: Boca Raton, FL, USA, 2021; pp. 126–152. ISBN 9780429328633. [Google Scholar]
- Steponkus, P.L.; Uemura, M.; Joseph, R.A.; Gilmour, S.J.; Thomashow, M.F. Mode of action of the COR15a gene on the freezing tolerance of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1998, 95, 14570–14575. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Kopka, J.; Sung, D.Y.; Zhao, W.; Popp, M.; Porat, R.; Guy, C.L. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 2007, 50, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Ruelland, E.; Vaultier, M.-N.N.; Zachowski, A.; Hurry, V. Chapter 2 Cold Signalling and Cold Acclimation in Plants. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2009; Volume 49, pp. 35–150. [Google Scholar]
- Yu, X.; Peng, Y.H.; Zhang, M.H.; Shao, Y.J.; Su, W.A.; Tang, Z.C. Water relations and an expression analysis of plasma membrane intrinsic proteins in sensitive and tolerant rice during chilling and recovery. Cell Res. 2006, 16, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, A.; Murai-Hatano, M.; Ishikawa-Sakurai, J.; Hayashi, H.; Kawamura, Y.; Uemura, M. Cold Stress-Induced Acclimation in Rice is Mediated by Root-Specific Aquaporins. Plant Cell Physiol. 2012, 53, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Boursiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Javot, H.; Lauvergeat, V.; Gerbeau, P.; Tournaire, C.; Santoni, V.; Heyes, J. Molecular physiology of aquaporins in plants. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 2002; Volume 215, pp. 105–148. [Google Scholar] [CrossRef]
- Javot, H.; Lauvergeat, V.; Santoni, V.; Martin-Laurent, F.; Güçlü, J.; Vinh, J.; Heyes, J.; Franck, K.I.; Schäffner, A.R.; Bouchez, D.; et al. Role of a Single Aquaporin Isoform in Root Water Uptake. Plant Cell 2003, 15, 509–522. [Google Scholar] [CrossRef]
- Postaire, O.; Tournaire-Roux, C.; Grondin, A.; Boursiac, Y.; Morillon, R.; Schäffner, A.R.; Maurel, C. A PIP1 Aquaporin Contributes to Hydrostatic Pressure-Induced Water Transport in Both the Root and Rosette of Arabidopsis. Plant Physiol. 2010, 152, 1418–1430. [Google Scholar] [CrossRef]
- Murai-Hatano, M.; Kuwagata, T.; Sakurai, J.; Nonami, H.; Ahamed, A.; Nagasuga, K.; Matsunami, T.; Fukushi, K.; Maeshima, M.; Okada, M. Effect of Low Root Temperature on Hydraulic Conductivity of Rice Plants and the Possible Role of Aquaporins. Plant Cell Physiol. 2008, 49, 1294–1305. [Google Scholar] [CrossRef]
- Aroca, R.; Amodeo, G.; Fernández-Illescas, S.; Herman, E.M.; Chaumont, F.; Chrispeels, M.J. The Role of Aquaporins and Membrane Damage in Chilling and Hydrogen Peroxide Induced Changes in the Hydraulic Conductance of Maize Roots. Plant Physiol. 2005, 137, 341–353. [Google Scholar] [CrossRef]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 Rice Aquaporin Genes and Analysis of Their Expression and Function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S. Heat and chilling induced disruption of redox homeostasis and its regulation by hydrogen peroxide in germinating rice seeds (Oryza sativa L., Cultivar Ratna). Physiol. Mol. Biol. Plants 2013, 19, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.T.; Cheng, D.G.; Hsu, Y.T.; Kao, C.H. Abscisic acid-induced hydrogen peroxide is required for anthocyanin accumulation in leaves of rice seedlings. J. Plant Physiol. 2008, 165, 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- Seifi, H.S.; Van Bockhaven, J.; Angenon, G.; Höfte, M. Glutamate Metabolism in Plant Disease and Defense: Friend or Foe? Mol. Plant-Microbe Interact. 2013, 26, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant. Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Charagh, S.; Abbas, S.; Hassan, M.U.; Saeed, F.; Haider, S.; Sharif, R.; Anand, A.; Corpas, F.J.; Jin, W.; et al. Assessment of proline function in higher plants under extreme temperatures. Plant Biol. 2023, 25, 379–395. [Google Scholar] [CrossRef] [PubMed]
- da Cruz, R.P.; Sperotto, R.A.; Cargnelutti, D.; Adamski, J.M.; FreitasTerra, T.; Fett, J.P. Avoiding damage and achieving cold tolerance in rice plants. Food Energy Secur. 2013, 2, 96–119. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, Q.; Wang, S.; Hong, Y.; Wang, Z. Rice and cold stress: Methods for its evaluation and summary of cold tolerance-related quantitative trait loci. Rice 2014, 7, 24. [Google Scholar] [CrossRef]
- Ma, Y.; Dai, X.; Xu, Y.; Luo, W.; Zheng, X.; Zeng, D.; Pan, Y.; Lin, X.; Liu, H.; Zhang, D.; et al. COLD1 Confers Chilling Tolerance in Rice. Cell 2015, 160, 1209–1221. [Google Scholar] [CrossRef]
- Deng, X.; Hu, W.; Wei, S.; Zhou, S.; Zhang, F.; Han, J.; Chen, L.; Li, Y.; Feng, J.; Fang, B.; et al. TaCIPK29, a CBL-Interacting Protein Kinase Gene from Wheat, Confers Salt Stress Tolerance in Transgenic Tobacco. PLoS ONE 2013, 8, e69881. [Google Scholar] [CrossRef]
- Tian, X.; Wang, Z.; Li, X.; Lv, T.; Liu, H.; Wang, L.; Niu, H.; Bu, Q. Characterization and Functional Analysis of Pyrabactin Resistance-Like Abscisic Acid Receptor Family in Rice. Rice 2015, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, J.; Li, F.; Liu, H.; Yang, W.; Chong, K.; Xu, Y. OsMAPK3 Phosphorylates OsbHLH002/OsICE1 and Inhibits Its Ubiquitination to Activate OsTPP1 and Enhances Rice Chilling Tolerance. Dev. Cell 2017, 43, 731–743.e5. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, S.; Qin, J.; Sun, C.; Liu, F. The lipid transfer protein OsLTPL159 is involved in cold tolerance at the early seedling stage in rice. Plant Biotechnol. J. 2020, 18, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yang, L.; Wang, J.; Liu, H.; Zheng, H.; Xie, D.; Zhang, M.; Feng, M.; Jia, Y.; Zhao, H.; et al. Identification of a Cold-Tolerant Locus in Rice (Oryza Sativa L.) Using Bulked Segregant Analysis with a next-Generation Sequencing Strategy. Rice 2018, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wang, L.; Xu, Y.; Chen, N.; Ma, Q.; Li, F.; Chong, K. Overexpression of OsCOIN, a Putative Cold Inducible Zinc Finger Protein, Increased Tolerance to Chilling, Salt and Drought, and Enhanced Proline Level in Rice. Planta 2007, 226, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, D.; Mao, D.; Liu, X.; Ji, C.; Li, X.; Zhao, X.; Cheng, Z.; Chen, C.; Zhu, L. Overexpression of MicroRNA319 Impacts Leaf Morphogenesis and Leads to Enhanced Cold Tolerance in Rice (Oryza Sativa L.). Plant Cell Environ. 2013, 36, 2207–2218. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Chiappetta, A.; Bruno, L.; Bitonti, M.B. In Posidonia oceanica cadmium induces changes in DNA methylation and chromatin patterning. J. Exp. Bot. 2012, 63, 695–709. [Google Scholar] [CrossRef]
- Ma, Q.; Dai, X.; Xu, Y.; Guo, J.; Liu, Y.; Chen, N.; Xiao, J.; Zhang, D.; Xu, Z.; Zhang, X.; et al. Enhanced Tolerance to Chilling Stress in OsMYB3R-2 Transgenic Rice Is Mediated by Alteration in Cell Cycle and Ectopic Expression of Stress Genes. Plant Physiol. 2009, 150, 244–256. [Google Scholar] [CrossRef]
- Hu, H.; You, J.; Fang, Y.; Zhu, X.; Qi, Z.; Xiong, L. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2008, 67, 169–181. [Google Scholar] [CrossRef]
- Wang, Q.; Guan, Y.; Wu, Y.; Chen, H.; Chen, F.; Chu, C. Overexpression of a rice OsDREB1F gene increases salt, drought, and low temperature tolerance in both Arabidopsis and rice. Plant Mol. Biol. 2008, 67, 589–602. [Google Scholar] [CrossRef]
- Joo, J.; Lee, Y.H.; Kim, Y.K.; Nahm, B.H.; Song, S.I. Abiotic stress responsive rice ASR1 and ASR3 exhibit different tissue-dependent sugar and hormone-sensitivities. Mol. Cells 2013, 35, 421–435. [Google Scholar] [CrossRef]
- Shi, J.; Cao, Y.; Fan, X.; Li, M.; Wang, Y.; Ming, F. A rice microsomal delta-12 fatty acid desaturase can enhance resistance to cold stress in yeast and Oryza sativa. Mol. Breed. 2012, 29, 743–757. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, J.Y.; Kim, S.J.; An, K.S.; An, G.; Kim, S.R. Isolation of Cold Stress-Responsive Genes in the Reproductive Organs, and Characterization of the OsLti6b Gene from Rice (Oryza Sativa L.). Plant Cell Rep. 2007, 26, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Kou, Y.; Liu, H.; Li, X.; Xiao, J.; Wang, S. OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice. J. Exp. Bot. 2011, 62, 4863–4874. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Xu, Y.; Wang, X.; Du, C.; Du, J.; Yuan, M.; Xu, Z.; Chong, K. OsRAN2, essential for mitosis, enhances cold tolerance in rice by promoting export of intranuclear tubulin and maintaining cell division under cold stress. Plant Cell Environ. 2011, 34, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wei, Q.; Zhang, K.; Wang, L.; Liu, F.; Zhao, L.; Tan, Y.; Di, C.; Yan, H.; Yu, J.; et al. Down-Regulation of OsSPX1 Causes High Sensitivity to Cold and Oxidative Stresses in Rice Seedlings. PLoS ONE 2013, 8, 81849. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Kang, I.S.; Han, J.S.; Lee, C.H.; An, G.; Moon, Y.H. OsDEG10 encoding a small RNA-binding protein is involved in abiotic stress signaling. Biochem. Biophys. Res. Commun. 2009, 380, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Sakata, N.; Suzuki, Y.; Shikanai, T.; Makino, A. Cyclic Electron Flow around Photosystem i via Chloroplast NAD(P)H Dehydrogenase (NDH) Complex Performs a Significant Physiological Role during Photosynthesis and Plant Growth at Low Temperature in Rice. Plant J. 2011, 68, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-W.G.-W.; Zhang, M.-H.M.-H.; Cai, W.-M.W.-M.; Sun, W.-N.W.-N.; Su, W.-A.W.-A. Characterization of OsPIP2: 7, a Water Channel Protein in Rice. Plant Cell Physiol. 2008, 49, 1851–1858. [Google Scholar] [CrossRef]
- Gothandam, K.M.; Nalini, E.; Karthikeyan, S.; Shin, J.S. OsPRP3, a flower specific Proline-rich protein of rice, determines extracellular matrix structure of floral organs and its overexpression confers cold-tolerance. Plant Mol. Biol. 2010, 72, 125–135. [Google Scholar] [CrossRef]
- Kim, S.-J.; Lee, S.-C.; Hong, S.K.; An, K.; An, G.; Kim, S.-R. Ectopic expression of a cold-responsive OsAsr1 cDNA gives enhanced cold tolerance in transgenic rice plants. Mol. Cells 2009, 27, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-F.; Wang, Y.-C.; Hsieh, T.-H.; Lu, C.-A.; Tseng, T.-H.; Yu, S.-M. A Novel MYBS3-Dependent Pathway Confers Cold Tolerance in Rice. Plant Physiol. 2010, 153, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, J.; Wang, X.; Chen, J. OVP1, a Vacuolar H+-translocating inorganic pyrophosphatase (V-PPase), overexpression improved rice cold tolerance. Plant Physiol. Biochem. 2011, 49, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Jiang, M.; Zhang, H.; Li, R. Zinc Oxide Nanoparticles Alleviate Chilling Stress in Rice (Oryza Sativa L.) by Regulating Antioxidative System and Chilling Response Transcription Factors. Molecules 2021, 26, 2196. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.H.; Huang, B.; Ding, C.B.; Zhang, Z.W.; Chen, Y.E.; Hu, C.; Zhou, L.J.; Huang, Y.; Liao, J.Q.; Yuan, S.; et al. Effects of Melatonin on Anti-Oxidative Systems and Photosystem II in Cold-Stressed Rice Seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, S.B.; Pires, S.N.; Ávila, G.E.; Silva, B.E.P.; Schmitz, V.N.; Deuner, C.; Silva Armesto, R.; Silva Moura, D.; Deuner, S.; da Silva Armesto, R.; et al. Application of Vigor Indexes to Evaluate the Cold Tolerance in Rice Seeds Germination Conditioned in Plant Extract. Sci. Rep. 2021, 11, 11038. [Google Scholar] [CrossRef]
- Bui, L.T.; Ella, E.S.; Dionisio-Sese, M.L.; Ismail, A.M. Morpho-Physiological Changes in Roots of Rice Seedling upon Submergence. Rice Sci. 2019, 26, 167–177. [Google Scholar] [CrossRef]
- Kumar, A.; Nayak, A.K.; Hanjagi, P.S.; Kumari, K.S.V.; Mohanty, S.; Tripathi, R.; Panneerselvam, P. Submergence stress in rice: Adaptive mechanisms, coping strategies and future research needs. Environ. Exp. Bot. 2021, 186, 104448. [Google Scholar] [CrossRef]
- Nishiuchi, S.; Yamauchi, T.; Takahashi, H.; Kotula, L.; Nakazono, M. Mechanisms for coping with submergence and waterlogging in rice. Rice 2012, 5, 2. [Google Scholar] [CrossRef]
- Kato, Y.; Collard, B.C.Y.; Septiningsih, E.M.; Ismail, A.M. Increasing flooding tolerance in rice: Combining tolerance of submergence and of stagnant flooding. Ann. Bot. 2019, 124, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms associated with tolerance to flooding during germination and early seedling growth in rice (Oryza sativa). Ann. Bot. 2009, 103, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.B. Physiological and Molecular Basis of Susceptibility and Tolerance of Rice Plants to Complete Submergence. Ann. Bot. 2003, 91, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, R.K.; Bhattacharjee, B. Rice Genotypes with SUB1 QTL Differ in Submergence Tolerance, Elongation Ability during Submergence and Re-generation Growth at Re-emergence. Rice 2011, 5, 7. [Google Scholar] [CrossRef]
- Colmer, T.D.; Armstrong, W.; Greenway, H.; Ismail, A.M.; Kirk, G.J.D.; Atwell, B.J. Physiological Mechanisms of Flooding Tolerance in Rice: Transient Complete Submergence and Prolonged Standing Water. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2014; Volume 75, pp. 255–307. [Google Scholar] [CrossRef]
- Singh, H.P.; Singh, B.B.; Ram, P.C. Submergence tolerance of rainfed lowland rice: Search for physiological marker traits. J. Plant Physiol. 2001, 158, 883–889. [Google Scholar] [CrossRef]
- Setter, T.L.; Ingram, K.T.; Tuong, T.P. Environmental characterization requirements for strategic research in rice grown under adverse conditions of drought, flooding and salinity. Int. Rice Res. Inst. 1995, 3–18. [Google Scholar]
- Bailey-Serres, J.; Fukao, T.; Ronald, P.; Ismail, A.; Heuer, S.; Mackill, D. Submergence Tolerant Rice: SUB1’s Journey from Landrace to Modern Cultivar. Rice 2010, 3, 138–147. [Google Scholar] [CrossRef]
- Sarkar, R.K.; Panda, D. Distinction and characterisation of submergence tolerant and sensitive rice cultivars, probed by the fluorescence OJIP rise kinetics. Funct. Plant Biol. 2009, 36, 222. [Google Scholar] [CrossRef]
- Emes, M.J.; Wilkins, C.P.; Smith, P.A.; Kupkanchanakul, K.; Hawker, K.; Charlton, W.A.; Cutter, E.G. Starch utilization by deepwater rices during submergence. Int. Deep. Rice Work. 1988, 26–30. [Google Scholar]
- Fukao, T.; Yeung, E.; Bailey-Serres, J. The Submergence Tolerance Regulator SUB1A Mediates Crosstalk between Submergence and Drought Tolerance in Rice. Plant Cell 2011, 23, 412–427. [Google Scholar] [CrossRef]
- Setter, T.L.; Bhekasut, P.; Greenway, H. Desiccation of leaves after de-submergence is one cause for intolerance to complete submergence of the rice cultivar IR 42. Funct. Plant Biol. 2010, 37, 1096. [Google Scholar] [CrossRef]
- Upadhyay, R.; Panda, S.; Dutta, B. Growth, Chlorophyll and Electric Conductivity responses of rice cultivars to different levels of Submergence and Post-submergence Stress. J. Phytol. 2009, 1, 425–432. [Google Scholar]
- Upadhyay, R.K. Oxidative Injury and Its Detoxification in Rice Plants after Submergence Stress. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 15–21. [Google Scholar] [CrossRef]
- Ribeiro, C.W.; Korbes, A.P.; Garighan, J.A.; Jardim-Messeder, D.; Carvalho, F.E.L.L.; Sousa, R.H.V.V.; Caverzan, A.; Teixeira, F.K.; Silveira, J.A.G.G.; Margis-Pinheiro, M. Rice Peroxisomal Ascorbate Peroxidase Knockdown Affects ROS Signaling and Triggers Early Leaf Senescence. Plant Sci. 2017, 263, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Voesenek, L. Flooding Stress: Acclimations and Genetic Diversity. Annu. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D.; Voesenek, L.A.C.J. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665. [Google Scholar] [CrossRef] [PubMed]
- Drew, M.C.; He, C.-J.; Morgan, P.W. Programmed cell death and aerenchyma formation in roots. Trends Plant Sci. 2000, 5, 123–127. [Google Scholar] [CrossRef]
- Shiono, K.; Takahashi, H.; Colmer, T.D.; Nakazono, M. Role of ethylene in acclimations to promote oxygen transport in roots of plants in waterlogged soils. Plant Sci. 2008, 175, 52–58. [Google Scholar] [CrossRef]
- Steffens, B.; Sauter, M. Epidermal Cell Death in Rice Is Confined to Cells with a Distinct Molecular Identity and Is Mediated by Ethylene and H2O2 through an Autoamplified Signal Pathway. Plant Cell 2009, 21, 184–196. [Google Scholar] [CrossRef]
- Hattori, Y.; Hattori, Y.; Nagai, K.; Nagai, K.; Furukawa, S.; Furukawa, S.; Song, X.-J.X.-J.; Song, X.-J.X.-J.; Kawano, R.; Kawano, R.; et al. The Ethylene Response Factors SNORKEL1 and SNORKEL2 Allow Rice to Adapt to Deep Water. Nature 2009, 460, 1026. [Google Scholar] [CrossRef]
- Colmer, T.D.; Pedersen, O. Oxygen dynamics in submerged rice (Oryza sativa). New Phytol. 2008, 178, 326–334. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A Is an Ethylene-Response-Factor-like Gene That Confers Submergence Tolerance to Rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Xu, K.; Ronald, P.; Bailey-Serres, J. A Variable Cluster of Ethylene Response Factor-like Genes Regulates Metabolic and Developmental Acclimation Responses to Submergence in Rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Bailey-Serres, J. Ethylene—A key regulator of submergence responses in rice. Plant Sci. 2008, 175, 43–51. [Google Scholar] [CrossRef]
- Jackson, M.; Armstrong, W. Formation of Aerenchyma and the Processes of Plant Ventilation in Relation to Soil Flooding and Submergence. Plant Biol. 1999, 1, 274–287. [Google Scholar] [CrossRef]
- COLMER, T.D. Aerenchyma and an Inducible Barrier to Radial Oxygen Loss Facilitate Root Aeration in Upland, Paddy and Deep-water Rice (Oryza sativa L.). Ann. Bot. 2003, 91, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Kuroha, T.; Nagai, K.; Gamuyao, R.; Wang, D.R.; Furuta, T.; Nakamori, M.; Kitaoka, T.; Adachi, K.; Minami, A.; Mori, Y.; et al. Ethylene-gibberellin signaling underlies adaptation of rice to periodic flooding. Science 2018, 361, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Mori, Y.; Ishikawa, S.; Furuta, T.; Gamuyao, R.; Niimi, Y.; Hobo, T.; Fukuda, M.; Kojima, M.; Takebayashi, Y.; et al. Antagonistic regulation of the gibberellic acid response during stem growth in rice. Nature 2020, 584, 109–114. [Google Scholar] [CrossRef]
- Reynoso, M.A.; Kajala, K.; Bajic, M.; West, D.A.; Pauluzzi, G.; Yao, A.I.; Hatch, K.; Zumstein, K.; Woodhouse, M.; Rodriguez-Medina, J.; et al. Evolutionary flexibility in flooding response circuitry in angiosperms. Science 2019, 365, 1291–1295. [Google Scholar] [CrossRef]
- Straeten, D.; Zhou, Z.; Prinsen, E.; Onckelen, H.A.; Montagu, M.C.; Van Der Straeten, D.; Zhou, Z.; Prinsen, E.; Van Onckelen, H.A.; Van Montagu, M.C. A Comparative Molecular-Physiological Study of Submergence Response in Lowland and Deepwater Rice. Plant Physiol. 2001, 125, 955–968. [Google Scholar] [CrossRef]
- Zarembinski, T.I.; Theologis, A. Expression characteristics of OS-ACS1 and OSACS2, two members of the 1-aminocyclopropane-1-carboxylate synthase gene family in rice (Oryza sativa L. Cv. Habiganj Aman II) during partial submergence. Plant Mol. Biol 1997, 33, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, Y.; Nagai, K.; Huan, P.D.; Shimazaki, K.; Qu, H.; Mori, Y.; Toda, Y.; Kuroha, T.; Hayashi, N.; Aiga, S.; et al. Rice leaf hydrophobicity and gas films are conferred by a wax synthesis gene (LGF 1) and contribute to flood tolerance. New Phytol. 2018, 218, 1558–1569. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.R.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef] [PubMed]
- Panda, D.; Sarkar, R.K. Mechanism associated with nonstructural carbohydrate accumulation in submergence tolerant rice (Oryza sativa L.) cultivars. J. Plant Interact. 2014, 9, 62–68. [Google Scholar] [CrossRef]
- Jisha, V.; Dampanaboina, L.; Vadassery, J.; Mithöfer, A.; Kappara, S.; Ramanan, R. Overexpression of an AP2/ERF Type Transcription Factor OsEREBP1 Confers Biotic and Abiotic Stress Tolerance in Rice. PLoS ONE 2015, 10, e0127831. [Google Scholar] [CrossRef]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated Responses to Oxygen and Sugar Deficiency Allow Rice Seedlings to Tolerate Flooding. Sci. Signal. 2009, 2, 61. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Kurokawa, Y.; Mori, Y.; Minami, A.; Reuscher, S.; Wu, J.; Matsumoto, T.; Ashikari, M. SNORKEL Genes Relating to Flood Tolerance Were Pseudogenized in Normal Cultivated Rice. Plants 2022, 11, 376. [Google Scholar] [CrossRef]
- Nemoto, K.; Ukai, Y.; Tang, D.-Q.; Kasai, Y.; Morita, M. Inheritance of early elongation ability in floating rice revealed by diallel and QTL analyses. Theor. Appl. Genet. 2004, 109, 42–47. [Google Scholar] [CrossRef]
- Tang, D.-Q.; Kasai, Y.; Miyamoto, N.; Ukai, Y.; Nemoto, K. Comparison of QTLs for Early Elongation Ability between Two Floating Rice Cultivars with a Different Phylogenetic Origin. Breed. Sci. 2005, 55, 1–5. [Google Scholar] [CrossRef]
- Hattori, Y.; Miura, K.; Asano, K.; Yamamoto, E.; Mori, H.; Kitano, H.; Matsuoka, M.; Ashikari, M. A Major QTL Confers Rapid Internode Elongation in Response to Water Rise in Deepwater Rice. Breed. Sci. 2007, 57, 305–314. [Google Scholar] [CrossRef]
- Kawano, R.; Doi, K.; Yasui, H.; Mochizuki, T.; Yoshimura, A. Mapping of QTLs for floating ability in rice. Breed. Sci. 2008, 58, 47–53. [Google Scholar] [CrossRef]
- Septiningsih, E.M.; Pamplona, A.M.; Sanchez, D.L.; Neeraja, C.N.; Vergara, G.V.; Heuer, S.; Ismail, A.M.; Mackill, D.J. Development of submergence-tolerant rice cultivars: The Sub1 locus and beyond. Ann. Bot. 2009, 103, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Yeung, E.; Bailey-Serres, J. The Submergence Tolerance Gene SUB1A Delays Leaf Senescence under Prolonged Darkness through Hormonal Regulation in Rice. Plant Physiol. 2012, 160, 1795–1807. [Google Scholar] [CrossRef] [PubMed]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s Role in Abiotic and Biotic Plant Stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Zhang, J.; He, L.; Hafeez, A.; Ning, C.; Cai, K. Silicon Enhances Plant Resistance of Rice against Submergence Stress. Plants 2021, 10, 767. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Du, H.; Wang, D.; Ma, M. Effects of Mercury Stress on Methylmercury Production in Rice Rhizosphere, Methylmercury Uptake in Rice and Physiological Changes of Leaves. Sci. Total Environ. 2021, 765, 142682. [Google Scholar] [CrossRef] [PubMed]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot. 1995, 46, 1843–1852. [Google Scholar] [CrossRef]
- Bundó, M.; Martín-Cardoso, H.; Pesenti, M.; Gómez-Ariza, J.; Castillo, L.; Frouin, J.; Serrat, X.; Nogués, S.; Courtois, B.; Grenier, C.; et al. Integrative Approach for Precise Genotyping and Transcriptomics of Salt Tolerant Introgression Rice Lines. Front. Plant Sci. 2022, 12, 797141. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A.; Rubio, F. High-affinity potassium and sodium transport systems in plants. J. Exp. Bot. 2006, 57, 1149–1160. [Google Scholar] [CrossRef]
- Ali, A.; Raddatz, N.; Pardo, J.M.; Yun, D. HKT sodium and potassium transporters in Arabidopsis thaliana and related halophyte species. Physiol. Plant. 2021, 171, 546–558. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na(+) and K(+) Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Khatun, S.; Rizzo, C.A.; Flowers, T.J. Genotypic variation in the effect of salinity on fertility in rice. Plant Soil 1995, 173, 239–250. [Google Scholar] [CrossRef]
- Rao, P.S.; Mishra, B.; Gupta, S.R. Effects of Soil Salinity and Alkalinity on Grain Quality of Tolerant, Semi-Tolerant and Sensitive Rice Genotypes. Rice Sci. 2013, 20, 284–291. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhang, H.; Li, Z.; Shen, S.; Sun, J.; Wang, M.; Liao, D.; Liu, X.; Wang, X.; Xiao, F.; et al. Evaluation of Genetic Diversity of Rice Landraces (Oryza sativa L.) in Yunnan, China. Breed. Sci. 2007, 57, 91–99. [Google Scholar] [CrossRef]
- Yeo, A.R.; Yeo, M.E.; Flowers, S.A.; Flowers, T.J. Screening of rice (Oryza sativa L.) genotypes for physiological characters contributing to salinity resistance, and their relationship to overall performance. Theor. Appl. Genet. 1990, 79, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Ali, A.; Safdar, L.B.; Zafar, M.M.; Rui, Y.; Shakeel, A.; Shaukat, A.; Ashraf, M.; Gong, W.; Yuan, Y. Salt stress induces physiochemical alterations in rice grain composition and quality. J. Food Sci. 2020, 85, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Zhou, H. Plant salt response: Perception, signaling, and tolerance. Front. Plant Sci. 2023, 13, 1053699. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Zhang, J.; Hua, J.; Zhong, C.; Zhu, L.; Feng, L.F.; Cao, X.; Chuan, X.; Chuang, Y.S.; Miao, S.; et al. Effects of Salt Stress on Rice Growth, Development Characteristics, and the Regulating Ways: A Review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Cha-Um, S.; Supaibulwattana, K.; Kirdmanee, C. Comparative Effects of Salt Stress and Extreme pH Stress Combined on Glycinebetaine Accumulation, Photosynthetic Abilities and Growth Characters of Two Rice Genotypes. Rice Sci. 2009, 16, 274–282. [Google Scholar] [CrossRef]
- Ran, X.; Wang, X.; Huang, X.; Ma, C.; Liang, H.; Liu, B. Study on the Relationship of Ions (Na, K, Ca) Absorption and Distribution to Photosynthetic Response of Salix matsudana Koidz Under Salt Stress. Front. Plant Sci. 2022, 13, 860111. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.M.L.; Tran, T.N.; Nguyen, T.K.T.; Williams, B.; Wurm, P.; Bellairs, S.; Mundree, S. Improvement of Salinity Stress Tolerance in Rice: Challenges and Opportunities. Agronomy 2016, 6, 54. [Google Scholar] [CrossRef]
- Vaidyanathan, H.; Sivakumar, P.; Chakrabarty, R.; Thomas, G. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)—Differential response in salt-tolerant and sensitive varieties. Plant Sci. 2003, 165, 1411–1418. [Google Scholar] [CrossRef]
- Yamane, K.; Mitsuya, S.; Kawasaki, M.; Taniguchi, M.; Miyake, H. Antioxidant Capacity and Damages Caused by Salinity Stress in Apical and Basal Regions of Rice Leaf. Plant Prod. Sci. 2009, 12, 319–326. [Google Scholar] [CrossRef]
- Schmidt, R.; Schippers, J.H.M.; Mieulet, D.; Watanabe, M.; Hoefgen, R.; Guiderdoni, E.; Mueller-Roeber, B. SALT-RESPONSIVE ERF1 Is a Negative Regulator of Grain Filling and Gibberellin-Mediated Seedling Establishment in Rice. Mol. Plant 2014, 7, 404–421. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, K.; Tester, M.; Roy, S.J. Quantifying the three main components of salinity tolerance in cereals. Plant. Cell Environ. 2009, 32, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Moradi, F.; Ismail, A.M. Responses of Photosynthesis, Chlorophyll Fluorescence and ROS-Scavenging Systems to Salt Stress During Seedling and Reproductive Stages in Rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef]
- Maathuis, F. K + Nutrition and Na + Toxicity: The Basis of Cellular K+/Na+ Ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Rodrigues, C.R.F.; Silva, E.N.; Ferreira-Silva, S.L.; Voigt, E.L.; Viégas, R.A.; Silveira, J.A.G. High K + supply avoids Na + toxicity and improves photosynthesis by allowing favorable K + : Na + ratios through the inhibition of Na + uptake and transport to the shoots of Jatropha curcas plants. J. Plant Nutr. Soil Sci. 2013, 176, 157–164. [Google Scholar] [CrossRef]
- Lee, K.-S.; Choi, W.-Y.; Ko, J.-C.; Kim, T.-S.; Gregorio, G.B. Salinity tolerance of japonica and indica rice (Oryza sativa L.) at the seedling stage. Planta 2003, 216, 1043–1046. [Google Scholar] [CrossRef]
- Ghosh, N.; Adak, M.K.; Ghosh, P.D.; Gupta, S.; Sen Gupta, D.N.; Mandal, C. Differential responses of two rice varieties to salt stress. Plant Biotechnol. Rep. 2011, 5, 89–103. [Google Scholar] [CrossRef]
- Kader, M.A.; Lindberg, S. Uptake of sodium in protoplasts of salt-sensitive and salt-tolerant cultivars of rice, Oryza sativa L. determined by the fluorescent dye SBFI. J. Exp. Bot. 2005, 56, 3149–3158. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.K.; Johnson, E.J.; MacElroy, R.D.; Speer, H.L.; Bruff, B.S. Effects of Salts on the Halophilic Alga Dunaliella viridis. J. Bacteriol. 1968, 95, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with Water Stress: Evolution of Osmolyte Systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.K.; Kim, J.; Owens, T.G.; Ranwala, A.P.; Choi, Y.D.; Kochian, L.V.; Wu, R.J. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA 2002, 99, 15898–15903. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Choudhary, K.K.; Chaudhary, N.; Gupta, S.; Sahu, M.; Tejaswini, B.; Sarkar, S. Salt Stress Resilience in Plants Mediated through Osmolyte Accumulation and Its Crosstalk Mechanism with Phytohormones. Front. Plant Sci. 2022, 13, 1006617. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.T. Programmed cell death: A way of life for plants. Proc. Natl. Acad. Sci. USA 1996, 93, 12094–12097. [Google Scholar] [CrossRef]
- Liu, S.-H.; Fu, B.-Y.; Xu, H.-X.; Zhu, L.-H.; Zhai, H.-Q.; Li, Z.-K. Cell death in response to osmotic and salt stresses in two rice (Oryza sativa L.) ecotypes. Plant Sci. 2007, 172, 897–902. [Google Scholar] [CrossRef]
- Li, J.; Jiang, A.; Zhang, W. Salt Stress-induced Programmed Cell Death in Rice Root Tip Cells. J. Integr. Plant Biol. 2007, 49, 481–486. [Google Scholar] [CrossRef]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef]
- Gumi, A.M.; Guha, P.K.; Mazumder, A.; Jayaswal, P.; Mondal, T.K. Characterization of OglDREB2A gene from African rice (Oryza glaberrima), comparative analysis and its transcriptional regulation under salinity stress. Biotech 2018, 8, 91. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2014, 84, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Jan, A.; Maruyama, K.; Todaka, D.; Kidokoro, S.; Abo, M.; Yoshimura, E.; Shinozaki, K.; Nakashima, K.; Yamaguchi-Shinozaki, K. OsTZF1, a CCCH-Tandem Zinc Finger Protein, Confers Delayed Senescence and Stress Tolerance in Rice by Regulating Stress-Related Genes. Plant Physiol. 2013, 161, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Xiao, J.; Xie, W.; Liu, H.; Li, X.; Xiong, L.; Wang, S. Rice Gene Network Inferred from Expression Profiling of Plants Overexpressing OsWRKY13, a Positive Regulator of Disease Resistance. Mol. Plant 2008, 1, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Xiao, J.; Ding, X.; Xiong, M.; Cai, M.; Cao, Y.; Li, X.; Xu, C.; Wang, S. OsWRKY13 Mediates Rice Disease Resistance by Regulating Defense-Related Genes in Salicylate- and Jasmonate-Dependent Signaling. Mol. Plant-Microbe Interact. 2007, 20, 492–499. [Google Scholar] [CrossRef]
- Liu, C.; Chen, K.; Zhao, X.; Wang, X.; Shen, C.; Zhu, Y.; Dai, M.; Qiu, X.; Yang, R.; Xing, D.; et al. Identification of Genes for Salt Tolerance and Yield-Related Traits in Rice Plants Grown Hydroponically and under Saline Field Conditions by Genome-Wide Association Study. Rice 2019, 12, 88. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Guo, R.; Shi, D.; Liu, B.; Lin, X.; Yang, C. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012, 12, 194. [Google Scholar] [CrossRef]
- Chen, G.; Hu, Q.; Luo, L.; Yang, T.; Zhang, S.; Hu, Y.; Yu, L.; Xu, G. Rice Potassium Transporter OsHAK1 Is Essential for Maintaining Potassium-Mediated Growth and Functions in Salt Tolerance over Low and High Potassium Concentration Ranges. Plant Cell Environ. 2015, 38, 2747–2765. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef]
- Kurotani, K.I.; Hayashi, K.; Hatanaka, S.; Toda, Y.; Ogawa, D.; Ichikawa, H.; Ishimaru, Y.; Tashita, R.; Suzuki, T.; Ueda, M.; et al. Elevated Levels of CYP94 Family Gene Expression Alleviate the Jasmonate Response and Enhance Salt Tolerance in Rice. Plant Cell Physiol. 2015, 56, 779–789. [Google Scholar] [CrossRef]
- Asano, T.; Hayashi, N.; Kobayashi, M.; Aoki, N.; Miyao, A.; Mitsuhara, I.; Ichikawa, H.; Komatsu, S.; Hirochika, H.; Kikuchi, S.; et al. A Rice Calcium-Dependent Protein Kinase OsCPK12 Oppositely Modulates Salt-Stress Tolerance and Blast Disease Resistance. Plant J. 2012, 69, 26–36. [Google Scholar] [CrossRef]
- Guan, Q.; Ma, H.; Wang, Z.Z.; Wang, Z.Z.; Bu, Q.; Liu, S. A rice LSD1-like-type ZFP gene OsLOL5 enhances saline-alkaline tolerance in transgenic Arabidopsis thaliana, yeast and rice. BMC Genom. 2016, 17, 142. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.-J.; Lee, S.-K.; Kim, B.-G.; Kwon, T.-R.; Cho, W.-S.; Park, Y.-T.; Lee, J.-O.; Kwon, H.-B.; Byun, M.-O.; Park, S.-C. A rice (Oryza sativa L.) MAP kinase gene, OsMAPK44, is involved in response to abiotic stresses. Plant Cell. Tissue Organ Cult. 2006, 85, 151–160. [Google Scholar] [CrossRef]
- Gao, Q.; Yin, X.; Wang, F.; Hu, S.; Liu, W.; Chen, L.; Dai, X.; Liang, M. OsJRL40, a Jacalin-Related Lectin Gene, Promotes Salt Stress Tolerance in Rice. Int. J. Mol. Sci. 2023, 24, 7441. [Google Scholar] [CrossRef] [PubMed]
- Diédhiou, C.J.; Popova, O.V.; Dietz, K.-J.; Golldack, D. The SNF1-type serine-threonine protein kinase SAPK4regulates stress-responsive gene expression in rice. BMC Plant Biol. 2008, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Kitamoto, H.K.; Nakamura, A.; Fukuda, A.; Tanaka, Y. Rice Shaker Potassium Channel OsKAT1 Confers Tolerance to Salinity Stress on Yeast and Rice Cells. Plant Physiol. 2007, 144, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Vishal, B.; Krishnamurthy, P.; Ramamoorthy, R.; Kumar, P.P. OsTPS8 controls yield-related traits and confers salt stress tolerance in rice by enhancing suberin deposition. New Phytol. 2019, 221, 1369–1386. [Google Scholar] [CrossRef] [PubMed]
- Hasthanasombut, S.; Ntui, V.; Supaibulwatana, K.; Mii, M.; Nakamura, I. Expression of Indica rice OsBADH1 gene under salinity stress in transgenic tobacco. Plant Biotechnol. Rep. 2010, 4, 75–83. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.; Liu, X.; Du, H.; Dai, M.; Zhou, D.X.; Yang, W.; Zhao, Y. The R2R3-Type MYB Gene OsMYB91 Has a Function in Coordinating Plant Growth and Salt Stress Tolerance in Rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef]
- Liu, S.; Zheng, L.; Xue, Y.; Zhang, Q.; Wang, L.; Shou, H. Overexpression of OsVP1 and OsNHX1 Increases Tolerance to Drought and Salinity in Rice. J. Plant Biol. 2010, 53, 444–452. [Google Scholar] [CrossRef]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Yamaji, N.; Costa, A.; Okuma, E.; Kobayashi, N.I.; Kashiwagi, T.; Katsuhara, M.; Wang, C.; Tanoi, K.; Murata, Y.; et al. OsHKT1: 4-Mediated Na+ Transport in Stems Contributes to Na+ Exclusion from Leaf Blades of Rice at the Reproductive Growth Stage upon Salt Stress. BMC Plant Biol. 2016, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Jing, W.; Xiao, L.; Jin, Y.; Shen, L.; Zhang, W. The Rice High-Affinity Potassium Transporterl: L Is Involved in Salt Tolerance and Regulated by an MYB-Type Transcription Factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Sugawara, M.; Okada, T.; Taira, K.; Kaothien-Nakayama, P.; Katsuhara, M.; Shinmyo, A.; Nakayama, H. Rice sodium-insensitive potassium transporter, OsHAK5, confers increased salt tolerance in tobacco BY2 cells. J. Biosci. Bioeng. 2011, 111, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Polispitak, K.; Thongpoem, P.; Singh, H.P.; Cha-um, S. Alleviation of Salt Stress in Upland Rice (Oryza sativa L. Ssp. Indica Cv. Leum Pua) Using Arbuscular Mycorrhizal Fungi Inoculation. Front. Plant Sci. 2020, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Jallad, K.N. Heavy metal exposure from ingesting rice and its related potential hazardous health risks to humans. Environ. Sci. Pollut. Res. 2015, 22, 15449–15458. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, R.; Tamás, M.J. How Saccharomyces cerevisiae copes with toxic metals and metalloids. FEMS Microbiol. Rev. 2010, 34, 925–951. [Google Scholar] [CrossRef]
- Sharma, S.K.; Goloubinoff, P.; Christen, P. Non-native Proteins as Newly-Identified Targets of Heavy Metals and Metalloids. In Cellular Effects of Heavy Metals; Springer: Dordrecht, The Netherlands, 2011; pp. 263–274. [Google Scholar] [CrossRef]
- Hu, T.; Zhu, S.; Tan, L.; Qi, W.; He, S.; Wang, G. Overexpression of OsLEA4 enhances drought, high salt and heavy metal stress tolerance in transgenic rice (Oryza sativa L.). Environ. Exp. Bot. 2016, 123, 68–77. [Google Scholar] [CrossRef]
- Liang, J.; Zhou, M.; Zhou, X.; Jin, Y.; Xu, M.; Lin, J. JcLEA, a Novel LEA-Like Protein from Jatropha curcas, Confers a High Level of Tolerance to Dehydration and Salinity in Arabidopsis thaliana. PLoS ONE 2013, 8, e83056. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Guillod-Magnin, R.; Brüschweiler, B.J.; Aubert, R.; Haldimann, M. Arsenic species in rice and rice-based products consumed by toddlers in Switzerland. Food Addit. Contam. Part A 2018, 35, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, V.; Zeibig, F.; Anumalla, M.; Siddiq, S.A.; Frei, M.; Murugaiyan, J.; Ali, J. Arsenic Stress Responses and Accumulation in Rice. In Rice Improvement; Springer: Berlin/Heidelberg, Germany, 2021; pp. 281–313. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hasegawa, H.; Rahman, M.M.; Miah, M.A.M.; Tasmin, A. Straighthead disease of rice (Oryza sativa L.) induced by arsenic toxicity. Environ. Exp. Bot. 2008, 62, 54–59. [Google Scholar] [CrossRef]
- Choudhury, B.; Chowdhury, S.; Biswas, A.K. Regulation of growth and metabolism in rice (Oryza sativa L.) by arsenic and its possible reversal by phosphate. J. Plant Interact. 2011, 6, 15–24. [Google Scholar] [CrossRef]
- Dubey, S.; Shri, M.; Misra, P.; Lakhwani, D.; Bag, S.K.; Asif, M.H.; Trivedi, P.K.; Tripathi, R.D.; Chakrabarty, D. Heavy metals induce oxidative stress and genome-wide modulation in transcriptome of rice root. Funct. Integr. Genom. 2014, 14, 401–417. [Google Scholar] [CrossRef]
- Mishra, S.; Jha, A.B.; Dubey, R.S. Arsenite treatment induces oxidative stress, upregulates antioxidant system, and causes phytochelatin synthesis in rice seedlings. Protoplasma 2011, 248, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium Toxicity in Plants: Impacts and Remediation Strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, J.; Song, K.; Sun, Y.; Qin, Q.; Xue, Y. Transcriptome Analysis of Rice (Oryza Sativa L.) Shoots Responsive to Cadmium Stress. Sci. Rep. 2019, 9, 10177. [Google Scholar] [CrossRef]
- Fu, H.; Yu, H.; Li, T.; Wu, Y. Effect of Cadmium Stress on Inorganic and Organic Components in Xylem Sap of High Cadmium Accumulating Rice Line (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2019, 168, 330–337. [Google Scholar] [CrossRef]
- Yang, H.; Yu, H.; Tang, H.; Huang, H.; Zhang, X.; Zheng, Z.; Wang, Y.; Li, T. Physiological responses involved in cadmium tolerance in a high-cadmium-accumulating rice (Oryza sativa L.) line. Environ. Sci. Pollut. Res. 2021, 28, 41736–41745. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M.; Tran, L.S.P. Physiological and Biochemical Mechanisms Associated with Trehalose-Induced Copper-Stress Tolerance in Rice. Sci. Rep. 2015, 5, 11433. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Rahman, M.M.; Ansary, M.M.U.; Fujita, M.; Tran, L.S.P. Interactive Effects of Salicylic Acid and Nitric Oxide in Enhancing Rice Tolerance to Cadmium Stress. Int. J. Mol. Sci. 2019, 20, 5798. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, X.; Li, K.; Wu, M.; Zhang, R.; Zhang, L.; Chen, G. Photosynthetic responses of Oryza sativa L. seedlings to cadmium stress: Physiological, biochemical and ultrastructural analyses. BioMetals 2014, 27, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Adrees, M.; Rizvi, H.; Zia-ur-Rehman, M.; Hannan, F.; Qayyum, M.F.; Hafeez, F.; Ok, Y.S. Cadmium stress in rice: Toxic effects, tolerance mechanisms, and management: A critical review. Environ. Sci. Pollut. Res. 2016, 23, 17859–17879. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Aslam, A.; Sheraz, M.; Ali, B.; Ulhassan, Z.; Najeeb, U.; Zhou, W.; Gill, R.A. Lead Toxicity in Cereals: Mechanistic Insight Into Toxicity, Mode of Action, and Management. Front. Plant Sci. 2021, 11, 2248. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Rolly, N.K.; Al Azzawi, T.N.I.; Imran, M.; Mun, B.G.; Lee, I.J.; Yun, B.W. Lead (Pb)-Induced Oxidative Stress Alters the Morphological and Physio-Biochemical Properties of Rice (Oryza Sativa L.). Agronomy 2021, 11, 409. [Google Scholar] [CrossRef]
- Khan, F.; Hussain, S.; Tanveer, M.; Khan, S.; Hussain, H.A.; Iqbal, B.; Geng, M. Coordinated effects of lead toxicity and nutrient deprivation on growth, oxidative status, and elemental composition of primed and non-primed rice seedlings. Environ. Sci. Pollut. Res. 2018, 25, 21185–21194. [Google Scholar] [CrossRef]
- Li, Y.F.; Zhao, J.; Li, Y.F.; Xu, X.; Zhang, B.; Liu, Y.; Cui, L.; Li, B.; Gao, Y.; Chai, Z. Comparative Metalloproteomic Approaches for the Investigation Proteins Involved in the Toxicity of Inorganic and Organic Forms of Mercury in Rice (Oryza sativa L.) Roots. Metallomics 2016, 8, 663–671. [Google Scholar] [CrossRef]
- Mao, Q.; Tang, L.; Ji, W.; Rennenberg, H.; Hu, B.; Ma, M. Elevated CO2 and Soil Mercury Stress Affect Photosynthetic Characteristics and Mercury Accumulation of Rice. Ecotoxicol. Environ. Saf. 2021, 208, 111605. [Google Scholar] [CrossRef]
- Palmieri, J.; Rzigalinski, B.; Benjamin, B.; Collins, E.; Kaur, G.; Brunette, J.; Council-Troche, M.; Wilson, M.; Meacham, S.; Guthrie, T. Implications and Significance of Mercury in Rice. J. Food Nutr. Metab. 2020, 3, 1–5. [Google Scholar] [CrossRef]
- Meng, B.; Feng, X.; Qiu, G.; Anderson, C.W.N.; Wang, J.; Zhao, L. Localization and Speciation of Mercury in Brown Rice with Implications for Pan-Asian Public Health. Environ. Sci. Technol. 2014, 48, 7974–7981. [Google Scholar] [CrossRef] [PubMed]
- Rao, G.; Ashraf, U.; Huang, S.; Cheng, S.; Abrar, M.; Mo, Z.; Pan, S.; Tang, X. Ultrasonic seed treatment improved physiological and yield traits of rice under lead toxicity. Environ. Sci. Pollut. Res. 2018, 25, 33637–33644. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, U.; Tang, X. Yield and Quality Responses, Plant Metabolism and Metal Distribution Pattern in Aromatic Rice under Lead (Pb) Toxicity. Chemosphere 2017, 176, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Hartley-Whitaker, J.; Ainsworth, G.; Meharg, A.A. Copper- and Arsenate-Induced Oxidative Stress in Holcus Lanatus L. Clones with Differential Sensitivity. Plant Cell Environ. 2001, 24, 713–722. [Google Scholar] [CrossRef]
- Duan, G.-L.; Hu, Y.; Liu, W.-J.; Kneer, R.; Zhao, F.-J.; Zhu, Y.-G. Evidence for a role of phytochelatins in regulating arsenic accumulation in rice grain. Environ. Exp. Bot. 2011, 71, 416–421. [Google Scholar] [CrossRef]
- Kong, D.; Ju, C.; Parihar, A.; Kim, S.; Cho, D.; Kwak, J.M. Arabidopsis Glutamate Receptor Homolog3.5 Modulates Cytosolic Ca2+ Level to Counteract Effect of Abscisic Acid in Seed Germination. Plant Physiol. 2015, 167, 1630–1642. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Jiang, J.; Li, S.; Li, M.; Tan, Y.; Song, S.; Shu, Q.; Huang, J. Glutamate Alleviates Cadmium Toxicity in Rice via Suppressing Cadmium Uptake and Translocation. J. Hazard. Mater. 2020, 384, 121319. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, S.H.; Lee, D.G.; Lee, H.; Lee, S.W.; Bahk, J.D.; Lee, B.H. Physiological and protein profiles alternation of germinating rice seedlings exposed to acute cadmium toxicity. Comptes Rendus Biol. 2007, 330, 735–746. [Google Scholar] [CrossRef]
- Chen, Y.-A.; Chi, W.-C.; Trinh, N.N.; Huang, L.-Y.; Chen, Y.-C.; Cheng, K.-T.; Huang, T.-L.; Lin, C.-Y.; Huang, H.-J. Transcriptome Profiling and Physiological Studies Reveal a Major Role for Aromatic Amino Acids in Mercury Stress Tolerance in Rice Seedlings. PLoS ONE 2014, 9, e95163. [Google Scholar] [CrossRef]
- Kalita, J.; Pradhan, A.K.; Shandilya, Z.M.; Tanti, B. Arsenic Stress Responses and Tolerance in Rice: Physiological, Cellular and Molecular Approaches. Rice Sci. 2018, 25, 235–249. [Google Scholar] [CrossRef]
- Liu, J.; Cao, C.; Wong, M.; Zhang, Z.; Chai, Y. Variations between rice cultivars in iron and manganese plaque on roots and the relation with plant cadmium uptake. J. Environ. Sci. 2010, 22, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Islam, E.; Khan, M.T.; Irem, S. Biochemical mechanisms of signalling: Perspectives in plant under arsenic stress. Ecotoxicol. Environ. Saf. 2015, 114, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Trivedi, P.K. Heavy Metal Stress Signaling in Plants. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier: Amsterdan, The Netherlands, 2016; pp. 585–603. [Google Scholar] [CrossRef]
- Rao, K.P.; Vani, G.; Kumar, K.; Wankhede, D.P.; Misra, M.; Gupta, M. Arsenic stress activates MAP kinase in rice roots and leaves. Arch. Biochem. Biophys. 2011, 506, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Conde, A.; Chaves, M.M.; Geros, H. Membrane transport, sensing and signaling in plant adaptation to environmental stress. Plant Cell Physiol. 2011, 52, 1583–1602. [Google Scholar] [CrossRef] [PubMed]
- Steinhorst, L.; Kudla, J. Calcium and reactive oxygen species rule the waves of signaling. Plant Physiol. 2013, 163, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.F.; Aarts, M.G. The molecular mechanism of zinc and cadmium stress response in plants. Cell. Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Lata, C.; Singh Chauhan, P.; Prasad, V.; Prasad, M. A functional genomic perspective on drought signalling and its crosstalk with phytohormone-mediated signalling pathways in plants. Curr. Genom. 2017, 18, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.M.; Hsiao, L.J.; Huang, H.J. Cadmium activates a mitogen-activated protein kinase gene and MBP kinases in rice. Plant Cell Physiol. 2004, 45, 1306–1312. [Google Scholar] [CrossRef]
- Liu, X.M.; Kim, K.E.C.; Kim, K.E.C.; Nguyen, X.C.; Han, H.J.; Jung, M.S. Cadmium activates Arabidopsis MPK3 and MPK6 via accumulation of reactive oxygen species. Phytochemistry 2010, 71, 614–618. [Google Scholar] [CrossRef]
- Singh, I.; Shah, K. Exogenous application of methyl jasmonate lowers the effect of cadmium-induced oxidative injury in rice seedlings. Phytochemistry 2014, 108, 57–66. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Trivedi, P.K.; Misra, P.; Tiwari, M.; Shri, M.; Shukla, D. Comparative transcriptome analysis of arsenate and arsenite stresses in rice seedlings. Chemosphere 2009, 74, 688–702. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Wang, T.; Chen, Z.; Tang, Z.; Wu, Z.; Salt, D.E.; Chao, D.Y.; Zhao, F.J. OsHAC1;1 and OsHAC1;2 function as arsenate reductases and regulate arsenic accumulation. Plant Physiol. 2016, 172, 1708–1719. [Google Scholar] [CrossRef]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 Using the CRISPR/Cas9 System Produces Low Cd-Accumulating Indica Rice without Compromising Yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, L.; Tang, Z.; Huang, X.Y.; Ma, J.F.; Zhao, F.J. Producing cadmium-free Indica rice by overexpressing OsHMA3. Environ. Int. 2019, 126, 619–626. [Google Scholar] [CrossRef]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The Plant Cuticle: An Ancient Guardian Barrier Set Against Long-Standing Rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Dong, J.; Tan, L.; Ji, Z.; Li, Y.; Sun, Y.; Chen, C.; Lv, Q.; Mao, B.; Hu, Y.; et al. Overexpression of OsLCT2, a Low-Affinity Cation Transporter Gene, Reduces Cadmium Accumulation in Shoots and Grains of Rice. Rice 2021, 14, 89. [Google Scholar] [CrossRef]
- Mishra, N.; Srivastava, A.P.; Esmaeili, N.; Hu, W.; Shen, G. Overexpression of the Rice Gene OsSIZ1 in Arabidopsis Improves Drought-, Heat-, and Salt-Tolerance Simultaneously. PLoS ONE 2018, 13, 201716. [Google Scholar] [CrossRef]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 Functions as a Metal Efflux Transporter Limiting Excess Zinc, Copper and Cadmium Accumulation in Rice. BMC Plant Biol. 2019, 19, 283. [Google Scholar] [CrossRef]
- Rono, J.K.; Le Wang, L.; Wu, X.C.; Cao, H.W.; Zhao, Y.N.; Khan, I.U.; Yang, Z.M. Identification of a New Function of Metallothionein-like Gene OsMT1e for Cadmium Detoxification and Potential Phytoremediation. Chemosphere 2021, 265, 129136. [Google Scholar] [CrossRef]
- Ogo, Y.; Itai, R.N.; Kobayashi, T.; Aung, M.S.; Nakanishi, H.; Nishizawa, N.K. OsIRO2 is responsible for iron utilization in rice and improves growth and yield in calcareous soil. Plant Mol. Biol. 2011, 75, 593–605. [Google Scholar] [CrossRef]
- Tao, J.; Lu, L. Advances in Genes-Encoding Transporters for Cadmium Uptake, Translocation, and Accumulation in Plants. Toxics 2022, 10, 411. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Ueda, Y.; Mukai, A.; Ochiai, K.; Matoh, T. Rice Phytochelatin Synthases OsPCS1 and OsPCS2 Make Different Contributions to Cadmium and Arsenic Tolerance. Plant Direct 2018, 2, e00034. [Google Scholar] [CrossRef] [PubMed]
- Shimo, H.; Ishimaru, Y.; An, G.; Yamakawa, T.; Nakanishi, H.; Nishizawa, N.K. Low cadmium (LCD), a novel gene related to cadmium tolerance and accumulation in rice. J. Exp. Bot. 2011, 62, 5727–5734. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, R.K.; Tuteja, N. OsSUV3 functions in cadmium and zinc stress tolerance in rice (Oryza sativa L. cv IR64). Plant Signal. Behav. 2014, 9, e27389. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Ye, Y.; Jiang, Z.; Wang, Y.; Zhu, C. MicroRNA390 Is Involved in Cadmium Tolerance and Accumulation in Rice. Front. Plant Sci. 2016, 7, 235. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yu, Y.; Chen, Q.; Mu, G.; Shen, Z.; Zheng, L. OsMYB45 plays an important role in rice resistance to cadmium stress. Plant Sci. 2017, 264, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Gong, S.; Wang, Y.; Wang, F.; Bao, H.; Sun, J.; Cai, C.; Yi, K.; Chen, Z.; Zhu, C. MicroRNA166 Modulates Cadmium Tolerance and Accumulation in Rice. Plant Physiol. 2018, 177, 1691–1703. [Google Scholar] [CrossRef]




{kind=link}
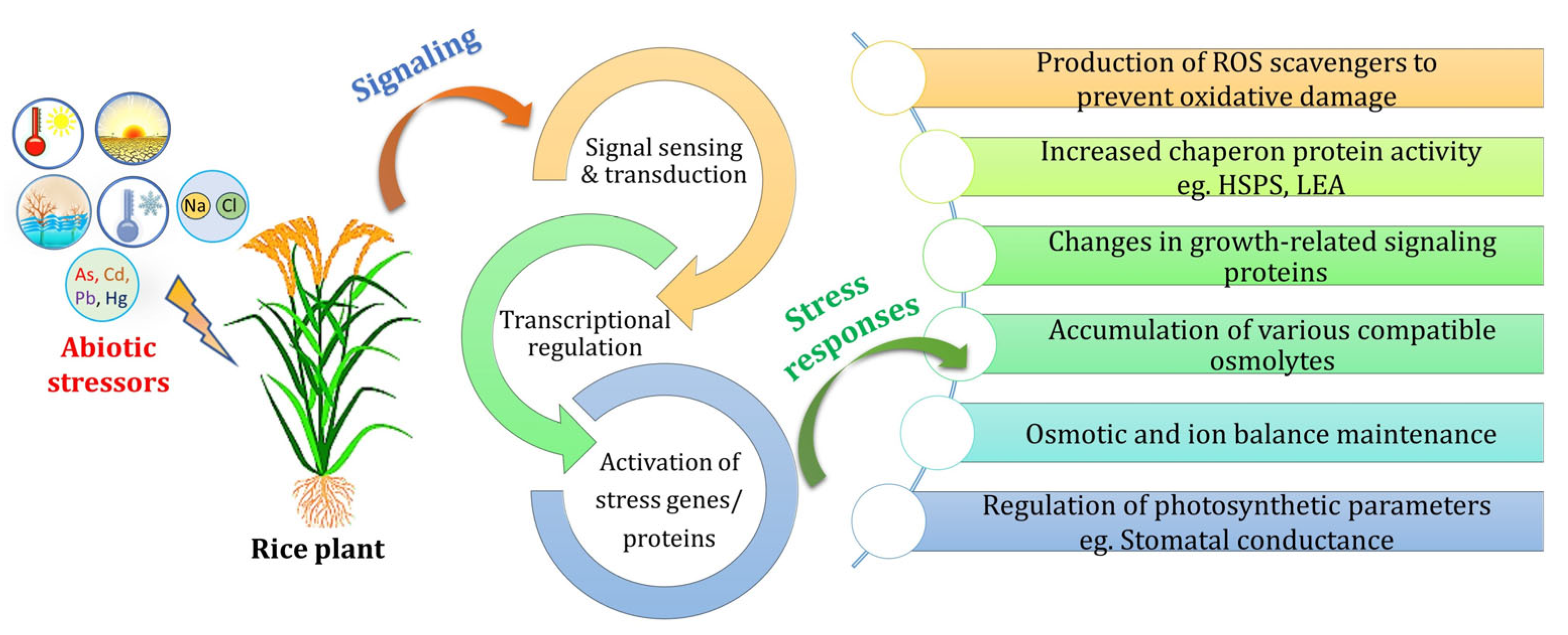{kind=link}
{kind=link}
| Name of Genes | Function | Reference |
|---|---|---|
| DRO1 | Stimulates the growth of roots, resulting in increased length and deeper penetration into the soil | [75] |
| EcNAC67 | Enhances water content, postpones leaf curling, and increases the mass of roots and shoots | [78] |
| DsM1 | Assists in removing reactive oxygen species and enhances drought resistance during the early growth (seedling) phase | [77] |
| OsPYL/RCAR5 | Causes the closure of stomata and controls the weight of leaves | [33] |
| OsDREB2B | Length of roots and the amount of root growth | [73] |
| OsNAC5 | Increases the size of the roots and improves the amount of grain produced | [79] |
| SNAC1 | Enhances spikelet fertility | [80] |
| OsLEA3-1 | Enhances grain yield | [81] |
| OsbZIP23 | Increase grain yield | [82] |
| OsbZIP72 | Enhancing tolerance to drought and increasing sensitivity to ABA (upregulating ABA) | [83] |
| AP37 | Improves the process of seed filling and increases the weight of the grain | [84] |
| OsNAC10 | Enhances resistance to drought during the vegetative phase, enhances root size, and enhances crop productivity | [79,85] |
| EDT1/HDG11 | Increases water use efficiency, the buildup of compatible osmolytes, heightened antioxidant enzymatic activity, and improves photosynthesis | [86] |
| AtDREB1A | Osmolytes accumulation, maintenance of chlorophyll, increment in relative water content, and reduction in ion leakage | [87] |
| OsCPK9 | Enhances drought tolerance in transgenics through improved stomatal closure and osmoregulation | [88] |
| ADC | Enhances resistance to drought by synthesis of polyamines such as putrescine and spermine | [61] |
| OsOAT | Enhances resistance to drought and promotes higher seed production | [89] |
| OsTPS1 | Enhances rice seedling’s tolerance to drought, cold, and salinity stress | [90] |
| P5CS | Enhances biomass production under salinity and drought stresses | [91] |
| HVA1 | Plasma membrane stability, increases leaf relative water content (RWC) and growth under drought stress | [92] |
| Hrf1 | Drought resistance via antioxidants generation, ABA signaling, and regulating stomata closure | [93] |
| JERF1 | Enhances drought resistance | [94] |
| OsRDCP1 | Improves drought stress tolerance | [95] |
| OsSDIR1 | Regulates stomata under drought stress | [96] |
| OsSRO1c | Regulates stomatal closure and enhances oxidative stress tolerance | [97] |
| Name of Genes | Function | Reference |
|---|---|---|
| OsMYB55 | Enhances amino acids’ metabolic process, enhancing the ability to withstand high temperatures | [139] |
| OsAREB1 | Controls abiotic stress-responsive gene expression utilizing an ABA-dependent mechanism | [143] |
| OsHSF7 | Increases the expression of HSPs and other genes that protect against exposure to high temperatures, resulting in enhanced resistance to heat | [144] |
| HSP101 | The effects of heat training in rice seedlings are prolonged by post-transcriptional interactions of HSA32/HSP101 after heat treatment | [145] |
| GAD3 | Participate in the ability to withstand high temperatures | [139] |
| OsHTAS | Improves rice’s ability to withstand heat by mediating stomata closure caused by H2O2 | [146] |
| TCM5 | Plays a vital role in the development of chloroplasts and the maintenance of PSII function in high temperatures | [147] |
| EG1 | Enhances homeostasis in floral organs and the ability to withstand temperature changes by activating a pathway involving mitochondrial lipase in response to high temperatures | [147] |
| OsTT1 | Breaks down poisonous denatured proteins while preserving the high-temperature response process | [127] |
| TOGR1 | Plays a role in the normal processing of rRNA precursors at high temperatures and acts as a chaperone for the nucleolar SSU complex, crucial for cell growth in high-temperature environments | [148] |
| OsHES1 | Plays a crucial part in adjusting to heat stress and ensuring the proper functioning of chloroplasts. | [149] |
| OsAET1 | Plays a dual function in regulating the response to high temperatures through tRNA modification and control of translation | [150] |
| OsNTL3 | Plays a crucial role in thermotolerance by interacting with OsbZIP74 | [151] |
| OsHsfA2c | Involved in regulating the transcription of the HSP100 gene in the cytoplasm of rice | [152] |
| OsHCI1 | Facilitates the nuclear export of target proteins, and its heterologous expression enhanced thermotolerance | [141] |
| OsNSUN2 | Controls the mRNA modification of 5-methylcytosine (m5C), which improves mRNA translation efficiency and sustains normal development at higher temperatures | [153] |
| OsTT3.1 | TT3.2 is ubiquitinated by TT3.1 for vacuolar degradation, and TT3.1 may function as a thermosensor | [154] |
| OsTT3.2 | Chloroplasts rely on mature TT3.2 proteins to protect thylakoids against the detrimental effects of heat stress | [154] |
| OsANN1 | Enhances SOD and CAT activity, controls H2O2 content and redox homeostasis, to provide cell protection against abiotic stress | [155] |
| Name of Genes | Function | Reference |
|---|---|---|
| OsLTPL159 | Reduces the toxic effects of ROS, increases cell wall’s cellulose deposition, and increases osmolyte accumulation in rice, which increases the plant’s ability to withstand cold temperatures in its early seedling stages | [200] |
| qPSST6 | Long-chain fatty acid production, involved in rice’s cold-tolerance during the booting stage | [201] |
| OsCOIN | Protein induced by cold enhances cold, drought, and salt tolerance | [202] |
| Osa-MIR319a | Increased leaf blade width | [203] |
| OsGH3-2 | Regulates ABA and auxin levels during cold and drought stress | [204] |
| OsMYB3R-2 | Regulates cell cycle (especially G2/M phase) to mediate cold tolerance in rice | [205] |
| SNAC2 | Enhances cold and salt tolerance in rice | [206] |
| OsDREB1F | Enhances cold tolerance in rice | [207] |
| ASR3 | Enhances cold/draught tolerance mediated by hormonal/sugar signaling | [208] |
| OsFAD2 | An essential enzyme that raises grain yield and germination rate under LTS (low-temperature stress conditions) | [209] |
| OsLti6b | Produces hydrophobic protein in the ovaries and stamens of flowers undergoing cold treatment | [210] |
| OsWRKY45 | Has a significant role in the signaling of ABA and serves as a means of communication between abiotic and biotic stresses | [211] |
| OsRAN2 | GTPase that enhances cold tolerance through cell cycle regulation | [212] |
| OsSPX1 | Participates in phosphate signaling as well as the interplay between the oxidative and cold stress tolerance mechanisms. | [213] |
| OsDEG10 | Produces RNA-binding protein and has a key role in cold tolerance as well as response to other stresses (anoxia, photooxidative, and salinity) | [214] |
| Oscrr6 | It has a key role in rice growth/photosynthesis at colder temperatures | [215] |
| OsPIP2 | Participates in water homeostasis during cold stress tolerance | [216] |
| OsPRP3 | Involved in the enhancement of cold tolerance in rice | [217] |
| OsAsr1 | Involved in both vegetative and reproductive stages of cold tolerance | [218] |
| MYBS3 | Modulates cold tolerance signaling pathways | [219] |
| OVP1 | Involved in lowering malondialdehyde levels and increasing proline accumulation to increase tolerance to cold | [220] |
| Name of Genes | Function | Reference |
|---|---|---|
| OsACS1 | Involved in ethylene production and the rapid elongation of the stem in submerged rice | [257,258] |
| OsACS5 | Involved in ethylene production and the rapid elongation of the stem in submerged rice | [257,258] |
| SNORKEL1 (SK1) | ERFs that modulate the internode elongation of deepwater rice during submergence | [247] |
| SNORKEL2 (SK2) | ERFs that regulate the internode elongation of deepwater rice during submergence | [247] |
| Submergence 1A (SUB1A) | Plant quiescence and plant survival under complete submergence | [249] |
| SDI | Involved in internode elongation | [254] |
| OsHSD1 | Involved in underwater photosynthesis in submerged rice | [259] |
| OsTPP7 | Involved in anaerobic germination | [260] |
| AGPPase | Promotes increased non-structural carbohydrate (NSC) buildup, which is accessible for a quick recovery after submersion | [261] |
| EREBP1 | enhances resistance to submersion and facilitates better recovery from extended submersion | [262] |
| CIPK15 | Involved in the regulation of sugar and energy production enabling growth of rice under floodwater | [263] |
| Name of Genes | Function | Reference |
|---|---|---|
| OsCPK12 | Increases resistance to high salt levels by decreasing ROS buildup | [319] |
| OsLOL5 | Enhance ROS scavenging and rice tolerance under salinity stress | [320] |
| OsMAPK44 | Participates in ion homeostasis under salinity stress | [321] |
| OsJRL40 | Increases antioxidant enzymatic activities and maintains the balance of Na+/K+ during salinity stress. Manages rice’s salt stress by regulating the expression of genes responsible for transporting Na+/K+, as well as genes involved in salt-responsive transcription factors and proteins | [322] |
| OsSAPK4 | Modulates ion homeostasis as well as the growth and development of rice in a salinized environment | [323] |
| OsKAT1 | Enhances rice’s salinity tolerance by enhancing K+ uptake and thus decreasing Na+ accumulation | [324] |
| OsTPS8 | Controls the ability of rice to tolerate salinity stress by managing the levels of soluble sugars and regulating the activity of genes related to ABA signaling through the regulation of SAPK9 | [325] |
| OsBADH1 | Enhances salinity stress tolerance by positively regulating osmoprotectant biosynthesis | [326] |
| OsMYB91 | Manages the growth of rice and its ability to tolerate salt stress. | [327] |
| OsVP1 and OsNHX1 | Enhances the tolerance of salt by decreasing the accumulation of Na+ in leaves, photosynthesis activity, and increase root biomass | [328] |
| OsHKT1;1, OsHKT1;4 and OsHKT1;5 | Enhance the tolerance of salt by decreasing the accumulation of Na+ in shoots when exposed to salt stress | [329,330,331] |
| OsHAK5 | Enhance rice’s salinity tolerance by contributing to cation homeostasis | [332] |
| Name of Genes | Function | Reference |
|---|---|---|
| OsHAC1;1 and OsHAC1;2 | Drastically influence limiting the accumulation of As in both the shoots and grains of rice | [382] |
| OsNRAMP5 | Enhances resistance to the toxicity of Cd | [383] |
| OsHMA3 | Enhances resistance to the toxicity of Cd | [384] |
| OsABCG31 | Enhances resistance to the toxicity of Cd and Pb | [385] |
| OsLCT1 | Enhances resistance to the toxicity of Cd Al | [386] |
| OsSIZ | Enhances resistance to the toxicity of Cd | [387] |
| OsZIP1 | Enhances resistance to the toxicity of Cd, Zn, | [388] |
| OsNAC5 | Enhances resistance to the toxicity of Cd and Pb | [79] |
| OsMT1e | Encodes a metal-detoxifying protein | [389] |
| OsIRO2 | TF that modulates the activity of genes related to Fe balance in rice | [390] |
| OsIRT1 | Participates in Cd absorption in rice. It is involved in Cd stress tolerance | [391] |
| OsPCS1 | It is involved in detoxifying heavy metals and involved in Cd stress tolerance | [392] |
| OsLCD | Involved in Cd compartmentation | [393] |
| OsSUV3 | Improved Cd and Zn stress tolerance | [394] |
| OsSRK | Increases the uptake and transfer of Cd | [395] |
| OsHMA2 | Improves transfer of Cd from roots to shoots | [395] |
| OsMYB45 | Improves Cd stress tolerance | [396] |
| OsHB4 | Improves Cd accumulation and tolerance | [397] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarma, B.; Kashtoh, H.; Lama Tamang, T.; Bhattacharyya, P.N.; Mohanta, Y.K.; Baek, K.-H. Abiotic Stress in Rice: Visiting the Physiological Response and Its Tolerance Mechanisms. Plants 2023, 12, 3948. https://doi.org/10.3390/plants12233948
Sarma B, Kashtoh H, Lama Tamang T, Bhattacharyya PN, Mohanta YK, Baek K-H. Abiotic Stress in Rice: Visiting the Physiological Response and Its Tolerance Mechanisms. Plants. 2023; 12(23):3948. https://doi.org/10.3390/plants12233948
Chicago/Turabian StyleSarma, Bhaskar, Hamdy Kashtoh, Tensangmu Lama Tamang, Pranaba Nanda Bhattacharyya, Yugal Kishore Mohanta, and Kwang-Hyun Baek. 2023. "Abiotic Stress in Rice: Visiting the Physiological Response and Its Tolerance Mechanisms" Plants 12, no. 23: 3948. https://doi.org/10.3390/plants12233948
APA StyleSarma, B., Kashtoh, H., Lama Tamang, T., Bhattacharyya, P. N., Mohanta, Y. K., & Baek, K. -H. (2023). Abiotic Stress in Rice: Visiting the Physiological Response and Its Tolerance Mechanisms. Plants, 12(23), 3948. https://doi.org/10.3390/plants12233948